0
Performance of common reed constructed
wetland on removal of tetracycline
antibiotic and resistance genes
Thesis for PhD degree in Environmental Engineering, Integrated Graduate
School of Medicine and Engineering, University of Yamanashi
Yamanshi
March, 2018
1
TABLE OF CONTENTS
ACKNOWLEDGEMENT 3
SUMMARY 4
CHAPTER 1. General introduction 7
Antibiotic and antibiotic resistance genes as emergence pollutants 7
Tetracycline antibiotic and tetracycline resistance genes 10
Wastewater treatment plant – a hotspot for spreading antibiotic and
antibiotic resistance genes 13
Constructed wetlands 14
Reference 16
Overall aims of the thesis 21
CHAPTER 2. Removal of tetracycline and tetracycline resistance genes from municipal wastewater in microcosm constructed wetland 22
Abstract 22
Introduction 22
Materials and Methods 24
Results and discussion 31
Conclusion 38
Reference 38
CHAPTER 3. Changes in tetracycline resistance genes populations and microbial community in tetracycline-contaminated and uncontaminated wastewater via constructed wetland treatment
43
Abstract 43
Introduction 43
Materials and Methods 45
Results and discussion 49
Conclusion 56
Reference 56
CHAPTER 4. Microbial acquisition and tetracycline resistance genes enrichment in common reed root planted in different wastewater 60
Abstract 60
2
Materials and Methods 61
Results and discussion 65
Conclusions 84
Reference 85
3
ACKNOWLEDGEMENT
My deep appreciation to Assoc. Prof. Mori Kazuhiro, my supervisor, for his advice and assistance in keeping my progress on schedule.
My sincere gratitude to Assoc. Prof. Toyama Tadashi for his patient guidance and useful critiques of this research work
My grateful thanks are also extended to Prof. Kaneko Hidehiro and Prof. Kazama
Futaba.
My thankfulness to Prof. Taku Misonou and Assoc. Prof. Keiko Hirayama for their work in my thesis adjustment committee.
My acknowledgment to all professors, staffs of ICRE for their support over the years; to my friends for being around.
Many thanks to the MEXT scholarship for giving me a chance to pursue my PhD degree in University of Yamanashi, Japan.
Thanks to all my friends for the support, incentive and patient.
Last but not least, to my parents and my brother for all their support and love. Thanks for standing by me, always.
4
SUMMARY
Tetracycline antibiotics are frequently detected in aquatic environments as a consequence of their extensive use in clinic for human therapy and agriculture for veterinary disease control and growth promoting. Moreover, elevated concentration of TC in the environments are found to favor the selection, acquisition and spread of tetracycline resistance genes (tet genes) in the environments. Because wastewater treatment plant usually are not able to remove properly antibiotics and/or antibiotic resistance genes (ARGs), treated effluent from WWTPs have been recognized as the main sources of antibiotics and ARGs through the environment. Because of many advantages of constructed wetland (CW), like the high efficiency of constructed wetland (CW) in treatment of various pollutant like nutrients, persistence organic compounds, and heavy metals, low maintenance, cost effectiveness, and environmental friendly, CW has been considered as a promising alternative technology for WWTP in removal antibiotic and antibiotic resistance genes.
The aims of this study were: 1) to investigate the ability of constructed wetland planted with common reed (Phragmites australis) in removal of tetracycline and tetracycline resistance genes, and the contribution of different compartments (soil, microorganisms, and plant) into the removal process; 2) The influence of TC in the microbial community and the abundance of tet genes in the wastewater treated through CW were examined; 3) The interaction between common reed and root-associated microorganisms, and their relationship to the microbial sources in the wastewater were dissected. In extension, the influence of common reed roots on the development and distribution of tet genes were investigated.
A high and durable effectiveness of common reed CW in removal of both TC and tet genes (tetC, tetM, tetQ, tetW) have been found when microcosm CWs were used, in greenhouse, to treat municipal wastewater supplied with 230 µg/L of TC. Laboratory experiment using pairwise combination of common reed, soil and wastewater revealed that the soil sorption might be the major removal mechanism over the short-term, but biodegradation by native microorganisms in wastewater also contributed in TC removal in CWs.
5 Antibiotic, even at very low concentration, can disturb microorganism community. Consequently, it not only affects to the removal ability of CW, but further may pose a risk to microbial ecology in CW effluent-receiving environment. On the other hand, sub-inhibitory concentration of antibiotic can favor the selection for resistance genes and induce the resistance genes transference between bacteria. By these reason, the microbial community and tetracycline resistance genes (tetA, tetC, tetQ) populations in effluent from CW were analyzed and compared to examine the influence of tetracycline in wastewater. The results showed that TC in wastewater not only had favorably selective effect on tet genes abundance, but also changed the composition of microbial communities, particularly the propagation of members belong to Chlamydiae and Gammaproteobacteria which include many human and animal pathogens.
CW performance was majorly depend on the activity of inhabited microbial community and the interaction between plant and microorganisms in the rhizosphere including root surface. In this regard, a better understanding on the relationship between wetland vegetation and their associated microbial community, as well as the interaction with the meta-microbiom in the environment can be benefit the removal efficiency and sustainable performance of CW. A laboratory experiment was conducted using the sterile seedling of common reed as model to explore the assemblage and composition of microbial community from domestic and swine wastewater into the root surface. The Illuminate sequencing results showed that the wastewater microbial community, as the source for microorganisms had important effects on the component of microorganisms can colony in root. However, common reed plant was the determinant for the composition of microbial community associated with its root. Common reed also could have an alteration effect on the microbial community in planting water. Because the favorable condition (available nutrients, addition dissolve oxygen, extensive attachment surface) not only increase the metabolism activities but also the horizontal gene transfer among rhizosphere bacteria which may increase antibiotic resistance genes acquisition and dissemination through environment. The abundance of 7 tet genes (tetA, tetC, tetM, tetO, tetQ, tetW, tetX) in the wastewater (no-CR water), wastewater planted with common reed (CR water), and common reed root (CR root) was evaluated using RealTime PCR. The different compositions and abundance of tet genes were found in domestic and swine wastewater in which tetM was specific to swine wastewater while other 6 tet genes were detected in
6 swine wastewater with concentration orders of magnitude higher than in domestic wastewater. The difference influence of CR on the abundance of tet genes in water phase as well as in root surface was found. However, the effects either, enhancement of inhibition, were varied among different tet genes and changed between swine and domestic wastewater.
7
CHAPTER 1
Literature review
1. Antibiotic and antibiotic resistance genes – emerging pollutants 1.1. Antibiotic: source, fate and effects on environment
Antibiotics are natural or semi-synthetic compounds that are used in the treatment and prevention of bacterial disease. Antibiotics are widely used for therapeutic and non-therapy purposes in humans, animals, aquaculture, and agriculture – including use for growth promotion. After administration into the body, a fraction of the drugs is completely metabolized to inactive compounds, but a significant amount (30 to 90%) is excreted as active metabolites via urine and feces being the major source of antibiotic input into the environment (Kummerer, 2009; Chee-Sanford et al., 2009).
Antibiotic have been quantified frequently in aquatic and terrestrial environments, all over the world, demonstrating a strong correlation between human activities and the amount of antibiotic released in the environment. Waste from animal farms, use in aquaculture and wastewater from antibiotic manufacturing, hospitals and municipalities are major sources of antibiotic pollution in the environment (Kummerer, 2009). Antibiotics from the sources can enter aquatic environment via different routes: direct application in aquaculture and/or direct discharge from farming site, indirectly through wastewater treatment plants (urban, hospital, industry); surface and subsurface runoff from agricultural fields that have been amended with manure containing antibiotic residues, and leakage from waste storage containers and landfills (Kummerer, 2009; Luo et al., 2014). On the other hand, water current is the main route for transportation and spread-out of antibiotic through environmental compartments like soil and sediment (Taylor, 2011).
When antibiotic residues enter the environment, the main processes determining their movement and fate are sorption to organic particles and degradation/transformation (Kummerer, 2009). While sorption control the mobility and transport of antibiotic in the environment, results of degradation processes (i.e. recalcitrance, complete mineralization or metabolites formation) can represent the persistence of antibiotic. Both process are driven by the antibiotic‘s physico-chemical properties like size, shape, solubility, speciation, molecular structure and hydrophobicity as well as the characteristics of the
8 natural environment including the pH, organic carbon content of particulate matter, and the charge on the surface of the particulate matter (Thiele-Bruhn, 2003). Antibiotic compounds are complex molecules that exhibit particular properties, such as the dependence of the octanol-water partition coefficient (Kow) on pH. In the range of
conditions encountered in the environment, antibiotics can be neutral, cationic, anionic, or zwitterionic (Kummerer, 2009) making it difficult to predicted the behavior. Sorption is an essential process because it controls the amount of chemical that can mobilized to surface water and groundwater, and the amount that can be degraded by a variety of chemical and biological processes. This value varies over several orders of magnitude results to the large difference in the environmental behavior of different antibiotics (Sarmah et al., 2006).
Residues of antibiotic in the environments can have deleterious effects on non-target organisms and environmental quality. Laboratory experiments show that antibiotics may bio-accumulate and cause adverse effects on different species of flora and fauna like duckweed, algae, daphnia, rotifers, and fish (Carlsson et al., 2009; Ji et al., 2012; Gonzalez-Pleiter et al., 2013). But, the main impact and the most critical concern is the development of antibiotic resistant strains of bacteria that can disturb natural bacterial ecosystems and lead to a serious threat to human health. There is clear evidence that long-term exposure to low-level antibiotics, similar to levels in the environment, can increase the amount of resistance (Knapp et al., 2008) and/or enhance the dissemination of antibiotic resistance in the environment (Heuer and Smalla, 2007). Other impacts that have not yet been fully evaluated include potential effects of antibiotic residues on the abundance and composition of microbial populations (Ding and He, 2010) and their role in ecological functions such as methanogenesis, nitrogen transformation, and sulfate reduction (Keen and Patrick, 2013). Costanzo et al., (2005) reported that antibiotic compounds (erythromycin, clarithromycin and amoxicillin) at 1 mg/L could inhibit denitrification processes in aquatic environments and the resistant genes, especially in eutrophic environments. Yergeau et al (2012) observed that erythromycin and sulfamethoxazole at concentration of 1 and 0.5 µg/L caused the unique expression of several functional genes and variable, antibiotic and taxa-specific changes in the abundance of bacteria. By affecting microbial population, diversity, and functionality,
9 antibiotics in the environment can influence the sustainability and proper performance of an ecosystem.
1.2. Antibiotic resistance genes (ARGs): source, characteristic and effect in environment
The emergence and spread of antibiotic resistant bacteria (ARB) and antibiotic resistance genes (ARGs) represent one of the most threating health care problems with worldwide consequence (Chee-Sanford et al., 2001; Levy, 2002).
As several antibiotics are produced by environmental microorganisms, antibiotic resistance is a natural property of environmental bacteria against to a range of selective pressure in nature (Baquero et al., 2008; Martinez et al., 2008). Nevertheless, the extensive use of antibiotic, for decades, in human, veterinary’s medicine and agriculture have accelerated greatly the development and dissemination of ARGs in the environment (Levy, 2002; Aminov, 2009). In a study by Knapp et al. (2010) in the Netherlands, abundance of resistance genes for different antibiotic class in rural soils was found increase significantly in range of 2 to 15 times between 1970s and 2008, showing an obvious anthropogenic contribution to the current situation.
Recent research has shown that selection of ARGs can occur at very low concentrations of antibiotics (Gullberg et al., 2011) and that pollution by antibiotics may increase the prevalence of ARGs which are in clinical relevant bacteria, in the environment (Tello et al., 2012). Moreover, many of known antibiotic resistance genes associate with plasmid and other mobile genetic elements, therefore they can efficiently spread among environmental bacteria populations and enhance their probability for maintenance in natural ecosystems.
Similar to antibiotic residues, ARGs can enter into aquatic environments through different routes; direct entry through discharge of untreated wastewater, indirect entry through wastewater treatment effluents and sludge discharge (Auerbach et al., 2007). Once spread into the environment, these human- and animal-original bacteria stand a chance to exchange resistance genes with environmental bacteria. The probability that resistance genes in environmental bacteria are transferred to and maintained by pathogenic bacteria may be low. However, as the types and abundance of antibiotic resistance gens in the environment increases, so does the risk that it will happen. There is evidence of the
10 transfer and acquisition of antibiotic resistance genes from environment to human commensal bacteria and pathogenes (Allen et al., 2010). As stated by Baquero et al. (2008), the contact of human microbiota with other types of microbiota in different ecosystems will increase the possibility of genetic variation and the possible emergence of novel mechanisms of resistance that are re-introduced in the human environment.
Because water provides bacteria with protection against dehydration and against UV light (i.e. many bacteria are sensitive for photo degradation), easily accessed nutrients for bacterial activity and gene transfer (Taylor, 2011), aquatic environments such as river, streams, and wastewater effluent have been considered as important in facilitating the transfer of ARGs to native bacteria in these environment and the wide spread in microbial populations through the biosphere (Aminov, 2009).
2. Tetracycline and tetracycline resistance genes 2.1. Tetracycline antibiotic
Tetracyclines are the second most common antibiotic group used for human health and veterinary purpose. Moreover, they are also used extensively as growth promoters in animal farms. They are broad-spectrum antibiotics that have been used since the 1940s against a wide range both Gram-negative and Gram-positive bacteria, atypical organisms and protozoan parasites (Chopra and Roberts, 2001). The chemical structure common to all tetracycline is the tetracycline nucleus consisting of four hydrocarbon rings with varying functional groups attached to them (Fig.1 ) (Chopra et al., 2001). The water solubility of tetracycline ranges between 230 and 5200 (mg L-1) with three pKa values of 3.3, 7.7, and 9.0 (Thiele-Bruhn, 2003). Because of these amphiphilic and amphoteric characteristics, adsorption of tetracycline is strongly influenced by soil properties such as pH, ionic environment, constituents, texture, and organic matter (Thiele-Bruhn, 2003). Distribution coefficients (Kd) for the adsorption of tetracyclines to soil material and aquatic
sediments are from 290 to 1620 L/kg (Thiele-Bruhn, 2003). They are susceptible to photo-degradation and form complexes with chelate divalent and trivalent metal ions such as Mg+2, Ca+2, Fe+2, Zn+2, and Al +2, Fe3+, Al3+, and β-diketones (Oka, 2000; Thiele-Bruhn, 2003).
The antibacterial functionality of tetracyclines is mediated by their ability to bind to the bacterial ribosome, thus preventing protein synthesis. More specifically, tetracycline bind
11 to the 30S ribosomal subunit in the tRNA acceptor site region. This binding sterically hinders the aminoacyl-tRNA from coming into contact with the incoming RNA, preventing translation (Chopra et al., 2001).
Figure 1. The chemical structure of tetracycline antibiotic. Imagine was from
www.ChemSpider.com
Tetracyclines are used frequently in human and veterinary medicine to combat bacterial infections and at sub-therapeutic levels are used to prevent epidemics and to promote growth and weight gain in livestock and aquaculture animals. Tetracycline (TC) is the second most used class of antibiotics after penicillin (Sarmah et al., 2006). Due to the widespread use of TC, they have been introduced in significant amounts to the environment through various routes such as hospital and municipal wastewater, manure from the agriculture industry and agricultural runoff. Recently, TCs have been frequently detected in different environments. Tetracycline were detected in surface water at concentrations between 3 and 10 µg/L (Valverde, 2006). Kulkarni et al. (2017) reported that tetracycline in wastewater can be reduced during wastewater treatment (from 188 µg/L to 23.6 µg/L) but persisted at low level in reclaimed water used for irrigation.
Once present in the environment, TC can modify the local environmental microbiota producing changes in their composition or activity. This impact has different levels and consequences that are not fully understood. Verma et al. (2007) found that tetracycline inhibited bacterial production in river water at a concentration of 5 µg/L. moreover, Quinlan et al., (2011) reported that TC had significant effects on the prevalence of
12 antibiotic resistance bacteria, bacterial abundance and productivity and algal biomass in river biofilms at a concentration of 0.5 µg/L.
2.2. Tetracycline resistance mechanisms and determinants
Bacterial resistance against tetracycline was discovered in less than a year after the discovery of the first tetracycline, although the mechanisms responsible were not elucidated until the 1970s (Nelson et al., 2011). Today, three mechanisms of gene-conferred resistance against tetracyclines are known; efflux, ribosomal protection and enzymatic inactivation (Nelson et al., 2011; Roberts, 2005)
The majority of tetracycline resistance genes code for energy-dependent efflux pumps (n = 30 different genes) which reduces the intracellular concentration of the antibiotic (Roberts and Schawrs., 2016). It is also the most common type of resistance gene among Gram-negative bacteria. Included among the efflux genes are tetA, tetC, and tetH. While the tetA gene is being present in 23 aerobic and facultative Gram-negative genera, the tetH gene has been found in 12 genera from the environment, food-producing animals, humans, and uncultured sludge. The tetC gene is the first acquired antibiotic resistance gene has been found in obligate intracellular bacteria (Chlamydia suits) (Roberts et al., 2016).
Ribosomal protection proteins are proteins which protect the bacterial ribosome from its interaction with tetracycline. By binding to the ribosome, the ribosomal protection protein confers a conformational change to the ribosome which disrupts the tetracycline binding site of the ribosome. Any bound tetracycline molecules are ejected from the ribosome, and protein translation many continue. Ribosomal protection protein genes (n = 12 different genes) are found among both Gram-negative and Gram-positive genera. Among the more common genes coding for this type of proteins are tetM, tetO, tetQ and tetW. The tetM gene is the most common tetracycline resistance which found naturally in 75 different general (including 38 Gram-positive and 37 Gram-negative). The tetO gene has been found in 35 genera (18 Gram-positive and 17 Gram-negative genera), while the tetW gene has been identified in 32 different genera (10 positve and 22 negative), and the tetQ gene has been found in 19 genera (8 positive and 11 Gram-negative) (Robert et al., 2016).
Inactivation of tetracycline by enzymatic activity is more rare type of tetracycline resistance mechanism. Only three genes coding for proteins capable of enzymatic
13 inactivation of tetracyclines are known (Roberts et al., 2016). The tetX gene is found in 11 different Gram negative genera, most of which are associated with the various environments (Roberts et al., 2016).
A total of 46 tetracycline resistance genes, for one gene, the mechanism of action is still unknown, have been encountered among 126 different genera including 76 Gram-negative and 50 Gram-positive from humans, animals, and multiple different environments (Roberts et al., 2016).
While some tetracycline resistance determinants are chromosomally encoded, the majority of tet genes are found on conjugative mobile gene elements (plasmid, transposons, conjugative transposon) (Roberts, 2005). It is likely that great diversity of tet genes and the diversity and mobilisability of the genetic elements in which they reside have contributed significantly to their dissemination among many different bacterial genera (Roberts et al., 2016). It is then not surprising that tetracycline resistance genes have often been found in various environmental compartments and environment can be potential reservoirs for animal and human pathogens (Roberts et al., 2016; Perry et al., 2014).
3. Wastewater treatment plant – a hotspot for spreading antibiotic and antibiotic resistance genes
WWTPs have been specifically individuated not only as the main sources of antibiotics, but also as platforms for the spread of both antibiotic resistant bacteria and ARGs through the environment (Rizzo et al., 2013).
Treated effluents from urban WWTPs have been characterized as hot spots of ARGs ending up in soil and aquatic environmental compartments, spreading and potentially accumulating AR among environmentally relevant bacteria (Rizzo et al., 2013). Consequently, the release of ARGs in WWTP effluents to environment is suspected to be among the main anthropogenic source of antibiotics, antibiotic resistant bacteria (ARB) and ARGs.
WWTP continuously receives wastewater containing antibiotics from different sources like household, hospital and industry. However, WWTPs are usually not equipped or not able to completely remove these special bioactive compounds, thus leaving behind a considerable level of antibiotic residues in WWTP effluent. Previous research also
14 identified that antibiotic removal may occurs through coagulation, activated carbon filtration, ionic treatment or micelle-clay system. However, in all cases a variable percentage of the antibiotics remain in wastewater treatment effluent and can challenge bacterial populations downstream the WWTPs (Li et al., 2009).
Furthermore, the conventional biological treatment like activate sludge creates a potentially idea environment for the development of ARGs, as ARB and antibiotic susceptible bacteria in the WWTP environment are continuously mixed with subinhibitory concentration of antibiotics and other ARB in close proximity, increasing the spread and prevalence of ARG (Michael et al., 2012, Rizzo et al., 2013). Various studies have been conducted in the context of the presence of AR in treated wastewater effluents by conventional activated sludge treatment, showing that ARB and ARGs are not effectively removed by these processes already (Novo et al., 2013; Czekalski et al., 2014). Eventually, the residual concentrations of ARB/ARGs end up as a result in environmental compartments such as soil and surface water environments through the reuse of the treat effluents (Gatica and Cytryn, 2013).
Currently, treated wastewater is widely used to compensate for diminishing water supplies, as it has been recognized to be a reliable alternative source of irrigation water, making wastewater reuse a practice of utmost importance (Kalavrouziotis et al., 2013; Fatta-Kassinos et al., 2015). As a result, the majority of pathogenic microorganisms that may be present in treated wastewater due to the inability to remove them by CAS treatment are introduced into the environments.
The residues of ARB/ARG in treated wastewater reuse worldwide poses a potential risk of microbiological contamination of ground and surface water, wildlife, and food chains, as shown in studies on their inadequate removal during CAS treatment (Beccerra-Castro et al., 2015). In order to prevent the selection pressure, flow, and spread of AR determinants such as antibiotic residues, ARG, and ARB to and from environmental reservoirs, the optimization and incorporation in the conventional urban WWT of new, alternative, and advanced wastewater processes is one important step for the mitigation of the spread and accumulation of AR determinants.
15 Constructed wetlands are engineered systems that use natural processes involving wetland vegetation, substrates (soils, gravel, sand), and their associated microbial assemblages to improve water quality (Vymaza et al., 2005).
The mechanisms that are available to remove pollutants from contaminated water are numerous and often interrelated include sorption, sedimentation, photolysis, hydrolysis, volatilization, plant uptake and accumulation, plant exudation, microbial degradation, filtration, precipitation and adsorption (Table 1) (Cooper et al., 1996). Removal efficiencies of CWs have been reported that highly depend on plant/ microorganisms interactions. These interactions allow the removal of contaminants from wastewater based on the increase of microbial activities and population numbers in the rhizosphere (Salvato et al., 2012). Plant has a strong influence in soil microbial communities. Microbial communities are stimulated by plant root exudates rich in carbon, nutrients and enzymes (Bais et al., 2006; Salvato et al., 2012) allowing the contaminant degradation. However, plant exudates and excreted exogenous enzymes can affect microbial community composition and diversity and, consequently, affect enzyme activity (Salvato et al., 2012). Fester et al. (2012) described that plant associated microorganisms often seem to be the real player mediating the plant impact on contaminant transformation. Plant also give a strong mechanical stability to wetlands in the presence of contaminants (Fester et al., 2014).
Since the 1950s, CWs have been used effectively to treat various types of wastewaters throughout the world, such as domestic and municipal wastewater, agricultural effluents, animal and industrial wastewaters, mine drainage, landfill leachates, urban and agricultural storm-waters, urban and highway runoff, and groundwater remediation (Vymaza and Brezinova, 2015; Vymazal, 2011; Kadlec and Wallence, 2009). CWs have been recognized as potential alternative for WWTPs, since they have associated low cost and low energy requirements, easy operation and maintenance, high removal efficiencies of several contaminants, high rates of water recycling and potential for providing significant wildlife habit. Nowadays, CWs are common alternative treatment systems in Europe and North America in rural areas and over 95% of these wetlands are subsurface flow wetlands (Vymazal, 2011). In addition, thousands of CWs have been applied as a promising alternative for wastewater treatment facilities in developing countries, e.g. in China (Chen et al., 2011).
16
References
Allen H.K., Donato J., Wang H.H., Cloud-Hansen K.A., Davies J., Handelsman J. (2010). Call of the wild: antibiotic resistance genes in natural environments. Natural Reviews Microbiology,8, 251-259
Aminov R.I. (2009). The role of antibiotics and antibiotic resistance in nature. Environmental microbiology, 11, 2970-2988
Baquero F., Martinez J.L., Canton R. (2008). Antibiotics and antibiotic resistance in water environments. Current Opinion in Biotechnology, 19, 260-265
Carlsson G., Orn S., Larsson D.G. (2009). Effluent from bulk drug production is toxic to aquatic vertebrates. Environmental Toxicology and Chemistry, 28, 2656-2662
Chee-Sanford J.C., Mackie R.I., Koike S., Krapac I.G., Lin Y.F., Yannarell A.C., Maxwell S., Aminov R.I. (2009). Fate and transport of antibiotic residues and antibiotic resistance genes following land application of manure waste. Journal of Environmental Quality, 38, 1086-1108
Chopra I., Roberts M. (2001). Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiology and Molecular Biology Reviews, 65, 232-260
Costanzo, S. D., Murby, J., & Bates, J. (2005). Ecosystem response to antibiotics entering the aquatic environment. Marine pollution bulletin, 51(1-4), 218–23
Costanzo S.D., Murby J., Bates J. (2005). Ecosystem response to antibiotics entering the aquatic environment, 51, 218-223
Czekalski N., Berthold T., Caucci S., Egli A., Burgmann H. (2012). Increased levels of multiresistant bacteria and resistance genes after wastewater treatment and their dissemination into Lake Geneva, Switzerland. Frontiers in Microbiology, doi: 10.3389/fmicb.2012.00106
Ding C., He J. (2010). Effect of antibotics in the environment on microbial populations. Application of Microbial Biotechnology, 87,925-941
17 Fester T., Giebler J., Wick L.Y., Schlosser D., Kastner M. (2014). Plant-microbe interactions as drivers of ecosystem functions relevant for the biodegradation of organic contaminants. Current Opinion in Biotechnology, 27, 168-175
Gatica J., Cytryn E. (2013). Impact of treated wastewater irrigation on antibiotic reistance in the soil microbiome. Environmental Science and Pollution Research, 20, 3529-3538
González-Pleiter M., Gonzalo S., Rodea-Palomares I., Leganés F., Rosal R., Boltes K., Marco E., Fernández-Piñas F. (2013). Toxicity of five anitbiotics and their mixtures towards photosynthetic aquatic organisms: implications for environmental risk assessment. Water Research, 47, 2050-2064
Gullberg E., Cao S., Berg O.G., Carolina I., Sandegren L., Hughes D., Andersson D.I (2011). Selection of resistant bacteria at very low antibiotic concentrations. PLoS Pathog 7(7): e1002158. doi:10.1371/journal.ppat.1002158
He L.Y, Ying G.G., Liu Y.S., Su H.C., Chen J., Liu S.S., Zhao J.L. (2016). Discharge of swine wastes risks water quality and foot safety: Antibiotics and antibiotic resistance genes from swine sources to receiving environments. Environment International, 92-93, 210-219
Heuer H., Smalla K. (2007). Horizontal gene transfer between bacteria. Environmental Biosafety Research, 6, 3-13
Ji X., Shen Q., Liu F., Ma J., Xu G., Wang Y., Wu M. (2012). Antibiotic resistance gene abundances associated with antibiotics and heavy metals in animal manures and agricultural soils adjacent to feedlots in Shanghai, China. Journal of Hazardous Material. 235-236, 178-185
Keen P.L., Patrick D.M. (2013). Tracking change: A Look at the ecological footprint of antibiotics and antimicrobial resistance, 2, 191-205
Knapp C.W., Chritiana A.E., Mark L.H., Patricia L.K., Kenneth J.H., David W.H. (2008). Indirect evidence of transposon-mediated selection of antibiotic resistance genes in aquatic systems at low level oxytetracycline exposures. Environmental Science and Technology, 42, 5348-5353.
18 Knapp C.W., Dolfing J., Ehlert P.A., Graham D.W. (2010). Evidence of increasing antibiotic resistance gene abundances in archived Soils since 1940. Environmental Science and Technology, 44, 580-587
Kulkarni P., Olson N.D., Raspanti G.A., Rosenberg Goldstein R.E., Gibbs S.G., Sapkota A., Sapkota A.R. (2017). Antibiotic concentration decrease during wastewater treatment but persist at low levels in reclaimed water. International Journal of Environmental Research and Public Health, 14, 668-671
Kummerer K. (2009). Antibiotics in the aquatic environment – A review – Part I. Chemosphere, 75, 435-441
Kummerer K. (2009). The presence of pharmaceuticals in the environment due to human use – present knowledge and future challenges. Journal of Environmental Management, 90, 2354-2366
Levy S.B. (2002). Factors impacting on the problem of antibiotic resistance. Journal of Antimicrobial Chemotherapy, 49, 25-30
Li D., Yang M., Hu J., Zhang J., Liu R., Gu X., Zhang Y., Wang Z. (2009). Antibiotic resistance profile in environmental bacteria isolated from penicillin production wastewater treatment plant and the receiving river. Environmental Microbiology, 11, 1506-1517
Luo Y., Guo W., Ngo H.H., Nghiem L.D., Hai F.I., Zhang J., Liang S., Wang X.C. (2014). A review on the occurrence of micropollutants in the aquatic environment and their fate and removal during wastewater treatment. Science of the Total Enivironment, 473-474, 619-641
Martinez J.L., Fajardo A., Garmendia L., Hernandez A., Linares J.H., Martinez-Solano L., Sanchez M.B. (2008). A global view of antibiotic resistance. Microbial Review, 33, 44-65
Michael I., Rizzo L., McArdell C.S., Manaia C.M., Merlin C., Schwartz T., Dagot C., Fatta-Kassinos D. (2012). Water Research, 47, 957-995
Nelson M.L., Levy S.B. (2011). The history of the tetracyclines. Antimicrobial Therapeutics Reviews: Antibiotics that target the ribosome, 1241, 17-32
19 Novo A., Andre A., Viana P., Nunes O.C., Manaia C.M. (2013). Antibiotic resistance, antimicrobial residues and bacterial community composisiton in urban wastewater. Water Research, 47, 1875-1887
Perry J.A., Westman E.L., Wright G.D. (2014). The antibiotic resistome: what’s new? Current Opinion in Microbiology, 21, 45-50
Quinlan E.L., Nietch C.T., Blocksom K.A. (2011). Temporal dynamics of periphyton exposed to tetracycline in stream mesocosms. Environmental Science and Technology, 45, 10684-10690
Rizzo L., Manaia C., Merlin C., Schwartz T., Dagot C., Ploy M.C., Michael I., Fatta-Kassinos D. (2013). Urban wastewater treatment plants as hotspots for antibiotic resistant bacteria and genes spread into the environment: A review. Science of Total Enivironment, 447, 345-360
Roberts M.C., Schwarz S. (2016). Tetracycline and Phenicol resistance genes and mechanisms: Importance for agriculture, the environment, and humans. Journal of Environmental Quality, 45, 576-592
Roberts M.C. (2005). Update on acquired tetracycline resistance genes. FEMS Microbiology Letters, 245, 195-203
Salvato M., Borin M., Doni S., Macci C., Ceccanti B., Marinari S., Masciandaro G. (2012). Wetland plants, microorganisms and enzymatic activities interrelations in treating N polluted water. Ecological Engineering, 47, 36-43
Sarmah, A. K., Meyer, M. T., & Boxall, A. B. a. (2006). A global perspective on the use, sales, exposure pathways, occurrence, fate and effects of veterinary antibiotics (VAs) in the environment. Chemosphere, 65, 725–59.
Seveno, N. A., Kalli, D., Smalla, Ã. K., Dirk, J., Elsas, V., Se, N. A., Collard, J., et al. (2002). Occurrence and reservoirs of antibiotic resistance genes in the environment. Reviews in Medical Microbiology, 13, 15–27.
Taylor, N. G. H., Verner-Jeffreys, D. W., & Baker-Austin, C. (2011). Aquatic systems: maintaining, mixing and mobilising antimicrobial resistance? Trends in ecology & evolution, 26(6), 278–84
20 Taylor N.G.H, Verner-Jeffeys D.W., Baker-Austin C. (2011). Aquatic systems: maintaining, mixing and mobilizing antimincrobial resistance? Trends in ecology and Evolution, 26, 278-284
Tello A., Austin B., Telfer T.C. (2012). Selective pressure of antibiotic pollution on bacteria of importance to public health. Environmental Health Perspective, 8, 1100-1106
Thiele-Bruhn S. (2003). Pharmaceutical antibiotic compounds in soils – a review. Journal of Plant Nutrient and Soil Science, 166, 145-167.
Valverde R.S., Garcia M.D.G., Galera M.M., Goicoechea H.C. (2006). Determination of tetracyclines in surface water by partial least squares using multivariate calibration transfer to correct the effect of solid phase preconcentration in photochemically induced fluorescence signals. Analytica Chimica Acta, 562, 85-93
Verma B., Robarts R.D., Headley J.V. (2007). Impacts of tetracycline on planktonic bacterial production in prairie aquatic systems. Microbial Ecology, 54, 52-55
Vymazal J, Brezinova T. (2015). The use of constructed wetlands for removal of pesticides from agriculture runoff and drainage: A review. Environmental International, 75, 11-20
Vymazal J. (2005). Horizontal sub-surface flow and hybrid constructed wetlands systems for wastewater treatment. Ecological Engineering, 25, 478-490
Vymazal J. (2011). Constructed wetlands for wastewater treatment: Five decades of Experience. Environmental Science and Technology, 45, 61-69
Yergeau E., Sanschagrin S., Waiser M.J., Lawrence J.R., Greeer C.W. (2012) Sub-inhibitory concentrations of different pharmaceutical products affect the meta-transcriptome of river biofilm communities cultivated in rotating annular reactors. Environmental Microbial Reports, 4, 350-359
21
OVERALL AIMS OF THE THESIS
The main goals of this research have been:
1. To investigate the ability to remove tetracycline and tetracycline resistance genes from wastewater by CWs, moreover, the contribution of macrophytes on the performance of CWs in treatment process. In order to reach this aim, microcosm CWs either planted with common reed or not were used to treat wastewater spiked tetracycline. The effluent from CWs were used to evaluate and compare removal efficiency between two sets.
2. There are many factors and removal mechanisms contribute to the treatment process in CWs. To exploit the mechanisms of tetracycline removal in constructed wetlands, a laboratory experiment was conducted. Under the control environment, the contribution of 3 major components, plant, microorganisms and soil was examined.
3. Several studies already showed how antibiotic in environment may impact on the bacterial community structure and function. However, it is a little information on the risk of antibiotic on microorganism in CWs treatment process. Therefore, the third objective of this research is to examine if tetracycline at sub-inhibited concentration could alter the microorganism compositions and tetracycline resistance genes population in wastewater treatment through CWs.
4. As microorganisms and plant/microorganisms interaction are key factor and major role in contaminant removal efficiency of CWs, an understanding of the bacterial community composition in rhizosphere/root surface could benefit the treatment effectiveness and sustainable performance of CWs. The relation between common reed and root-associated microbial community, as well as their interaction with microbiome in two wastewater types (domestic and swine wastewater) were investigated.
5. Considering rhizosphere as a hot spot for horizontal gene transfer, a RealTime PCR quantification of seven tet genes in the common reed root planted in domestic wastewater and swine wastewater was conducted to examine the effects of common reed on the abundance and distribution of tet genes between root surface and water phase.
22
CHAPTER 2.
Removal of tetracycline and tetracycline resistance genes from municipal
wastewater in microcosm constructed wetland
ABSTRACT
This study investigated the capacity of fill-and-drain constructed wetlands (CWs) for removal of a common antibiotic, tetracycline (TC), and tetracycline resistance genes (tet genes) from municipal wastewater. TC (230 µg/L) containing wastewater was treated in the CWs planted or non-planted with common reed (Phragmites australis). TC was removed significantly in the planted (95.4 % removal) and non-planted (87.2% removal) CWs with a treatment time of 1 day. Both CWs, with longer treatment times, completely removed TC from the wastewater. Adsorption of TC to soil materials might be the major mechanism of removal by the CWs over the short-term. Biodegradation of TC by native microorganisms present in wastewater also contributed in TC removal in CWs. In addition, the planted CWs showed higher TC removal efficiency than did the non-planted ones. The presence of five tet genes (tetC, tetM, tetO, tetQ, and tetW) was monitored in the planted CWs. The influent wastewater had 1.7 × 102‒2 × 104 copies/mL of these genes. All the tet genes were completely removed from wastewater by the planted CWs with 1 day treatment. In 28 days sequencing batch experiments using planted and non-planted CWs treating TC containing wastewater (250 µg/L) with treatment time of 2 days, the planted CWs completely and repeatedly removed TC from the wastewater. TC removal by non-planted CWs was 98.9‒99.8%, and low concentration of TC persisted in the effluents. The presence of plants provided the effective TC removal in the CWs for a long-term. The planted CWs also maintained about 3 log reduction of tet genes from wastewater during the sequencing batch experiments. These results suggest the potential of planted CWs for use in the removal TC and tet genes from municipal wastewater.
INTRODUCTION
Tetracycline (TC) is a common antibiotic, widely used in the therapy of human and veterinary infection. Besides, it is usually added at sub-therapeutic levels to animal feeds for growth promoting purpose(Daghrir et al., 2013). The intensive and extensive uses of TC result in its release to aquatic environmentswhere this antibiotic can cause acute and
23 chronic toxicities to aquatic organisms(Daghrir et al., 2013; Ji et al., 2012). Moreover, the presence of TC, even at very low concentrations, can facilitate the emergence and spread of tetracycline resistance genes (tet genes) (Gullber et al., 2011). Tet genes reduce the therapeutic potential of TC against human and animal pathogens and, thereby, increase the risk to public health. TC and tet genes are frequently detected in various types of aquatic environments, including surface water, ground water, sediment, and drinking water (Daghrir et al., 2013; Kummerer, 2004; Thomas et al., 2003Chee-Stanford et al., 2001). For these reasons, TC and tet genes are considered as emerging pollutants, responsible for public health problems.
Conventional wastewater treatment plants (WWTPs) continuously receive wastewater from various sources, such as homes, hospitals, and livestock industries, which contains antibiotics and antibiotic resistance genes (ARGs), particularly TC and tet genes. However, these treatment plants are usually not able to adequately remove the antibiotics and ARGs before discharging the water into the environment7). Moreover, some studies have
suggested that biological wastewater treatment processes, such as activated sludge, can provide suitable conditions for the development of ARGs and their spread because bacteria are continuously mixed with antibiotics or ARGs (Rizzo et al., 2013; Gao et al., 2013). WWTPs are considered a “hotspot” for the release of antibiotics and ARGs into the environment (Michael et al., 2013; Rizzo et al., 2013). Therefore, additional advanced treatment processes to remove both antibiotics and ARGs from wastewater effluent are highly desired.
Constructed wetlands (CWs) are engineered wetland systems designed to stimulate the natural processes based on interactions among soil, microorganisms, and plants for the treatment of wastewaters. CWs are low-cost, low-energy consuming, easy-operable, and environment-friendly option and have high potential for application in developing countries and small rural communities. CWs can efficiently remove organic compounds (biochemical oxygen demand (BOD) and chemical oxygen demand (COD)), total suspended solids, and nutrients from wastewaters (Vymazal, 2007; Karathanasis et al., 2003). Some researchers have also demonstrated the removal of xenobiotic chemicals, such as phenols (Kurzbaum et al., 2010) and pesticides (Vymazal and Brezinova, 2015). The mechanisms for removal of these pollutants are microbial degradation, microbial
24 transformation, soil-adsorption, plant uptake, and degradation by plants (phytodegradation)(Imfeld et al., 2009).
Some recent studies have assessed the removal of TC and/or tet genes from wastewater using different types of CWs including subsurface horizontal flow CW (Chen et al., 2016; Liu et al., 2014), vertical flow CW (Liu et al., 2014; Liu et al., 2013), vertical up-flow CW (Huang et al., 2015), integrated CW (Chen et al., 2015). Vertical subsurface flow CWs showed the highly-efficient removal of TC and tet genes fror wastewaters. For example, Huang et al. (2015)reported a removal efficiency of 69.0‒99.7% of TC and about 0.26 to 3.3 log unit of tet genes (tetA, tetO, tetM, tetW, and tetX) while Liu et al. (2014) reported that 97‒99% of TC and about 1 log unit of tet genes (tetM, tetO, and tetW) were removed from wastewater by vertical flow CWs. These studies suggested that adsorption to soil materials might be the primary mechanism for TC removal in CWs (Liu et al., 2014; Liu et al., 2013). Nevertheless, the contributions of each of the CW components – soil, microorganisms, and plants –, especially microorganisms and plants, in removal of TC by CWs are still not clear. Identification of the mechanisms underlying the TC and tet gene removal in CWs is essential for designing a highly-effective CWs for removal of them from wastewaters.
In this study, we set up microcosm-scale CWs and operated them in a fill-and-drain batch mode. The present study was taken up with the following objectives: (i) to assess the removal efficiency of TC and tet genes by CWs in microcosm-scale CW experiments for short and long terms and (ii) to understand the mechanisms and the contributions of microorganisms, soil, and plants in TC removal by CWs in the small-scale CW experiments.
MATERIALS AND METHODS Chemical
TC was purchased from Kanto-Chemical Co., Japan and its stock solution (250 mg/L in pure sterilized water) was made and used in the experiments.
Plant samples
Young potted seedlings of common reed (P. australis) (50 to 70 cm tall; 12 to 16 seedlings per pot; pot diameter, 6 cm) were purchased from the ESPEC MIC Corporation
25 (Aichi, Japan) and used for microcosm-scale CWs. To obtain sterile (bacteria-free) P.
australis plants for the experiment in TC removal mechanisms, we sterilized the seeds by
washing for 5 min in a solution of 0.5% sodium hypochlorite and 0.02% Tween 80; this was followed by a 1-min wash in 70% ethanol and rinses with sterilized water thrice, for 1 min each. The sterilized seeds were then grown aseptically in a flask containing sterile Murashige-Skoog’s (MS) medium (Duchefa, The Netherlands). The seedlings were grown in a growth chamber at 28 ± 1°C under fluorescent lamps (at a photosynthetic photon flux density of 80 μmol/m2/s and a photoperiod of 16L:8D).
Wastewater sample
Secondary effluent sample was collected from a municipal wastewater treatment plant in Kofu, Yamanashi, Japan, and was used as the wastewater sample in this study. The average values for the chemical properties of the effluent samples (n=10) were as follows: pH 7.4 ± 0.5; 4.2 ± 1.4 mg/L ammonium-N; 0.41 ± 0.09 mg/L nitrite-N; 5.7 ± 2.6 mg/L nitrate-N; 2.2 ± 0.7 mg/L phosphate-P; 16.2 ± 4.1 mg/L total dissolved organic carbon (TOC); 8.6 ± 3.7 mg/L suspended solid (SS); 2.6×106−5.7×107 colony-forming units/mL
of total heterotrophic bacteria. The bacterial counts were performed using 0.1×LB agar plates (1.0 g/L Bacto Peptone, 0.5 g/L yeast extract, 1.0 g/L NaCl, pH 7.0, and 2.0% agar (w/v). TC in the wastewater sample was under detection limit (0.0025 µg/L). The wastewater samples were collected weekly and stored at 4°C until used.
Setup of microcosm-scale constructed wetlands
Microcosm-scale CWs (Fig. 1) were set up in 1-L plastic pots (100 mm diameter and 150 mm height), of which half were planted with young seedlings of native P. australis and the remains were not planted. The CWs were filled with 100 g pumice rock (grain size, 10–20 mm) from bottom to 20 mm height and with 200 g red soil (5–10 mm) from 20 to 130 mm height. The subsurface parts of CWs were wrapped in black plastic sheets to prevent the effect of sunlight. All the CWs were set up on March 15th 2016. CWs were operated in a fill-and-drain batch mode. The preparation period was set for 6 weeks before starting experiment on May 10th 2016 in order to construct a microbial community in CW. In this preparation period, each CW was filled with 400 mL wastewater sample to the soil level. The wastewater sample was maintained for 2 days, then, all the wastewater was drained from the bottom drainage port. In the preparation and experimental periods, the
26 CWs were kept in a greenhouse without any air heating and artificial lighting. The daily average temperature during the experimental period was 21–29 °C.
Figure 1. Schematic diagram of microcosm-scale CW
Experiment for the assessment of the removal of TC and tet genes by the microcosm-scale CWs over a short-term
Experiment for assessing the effect of treatment time on TC removal were conducted in the planted and non-planted CWs in parallel. Four different treatment times of 1, 2, 3, and 4 days were set for each planted and non-planted CWs. Just before the experiment beginning, the TC-polluted wastewater sample was prepared by dissolving 230 μg/L TC into the previously described wastewater sample. Each CW was filled with 400 mL wastewater containing 230 μg/L TC and was maintained for the each treatment time. After each treatment period, all the wastewater was collected from the CW pots for TC analysis. The collected effluent water volume was measured. And then, the TC concentration in the effluent sample was normalized to the initial water volume (400 mL). Also, the influent wastewater and collected effluent samples were subjected to DNA extraction and
27 quantification of tet genes. The tet gene quantification was conducted only for the planted CWs.
TC removal experiments using wastewater, soil, or plant materials in flasks
Figure 2. TC removal experiment using water, soil, and common reed in flasks
To examine the contribution of microorganisms present in wastewater, soil materials, and plant materials in TC removal in CWs, three types of TC removal experiments were conducted (Fig. 2).
To examine the biodegradation of TC by microorganisms present in wastewater, 100 mL wastewater or autoclaved (121°C, 30 min) wastewater was added into a flask, and then TC was dissolved at a final concentration of 250 µg/L. The flasks were statically incubated at 28°C under dark conditions.
TC adsorption by soil material and biodegradation by microorganisms attached to the soil material were examined as follows: the soil materials were collected from the planted CWs, and 100 g (wet weight) of the soil or the autoclaved (121°C, 60 min) soil were added in a flask, and 300 mL of sterile deionized water was supplemented with 250 µg/L TC. The flasks were statically incubated at 28°C under dark conditions.
The uptake, degradation, and adsorption of TC by P. australis were examined as follows: the entire 2-month-old sterile P. australis plant (leaves and shoots having a wet weight of about 1.6 g and the roots having a wet weight of about 0.9 g) was rinsed thrice with sterile water to remove any root exudates available in the flask. In addition, the roots
28 (about 0.9 g-wet weight) of 2-month-old sterile plant were cut off from the upper part, autoclaved (121°C, 30min), and the autoclaved root was placed in the flask. Thereafter, 100 mL of MS medium containing 250 µg/L TC was added into the flask. These flasks were statically incubated in a growth chamber at 28 ± 1°C, under fluorescent lamps at a photosynthetic photon flux density of 80 μmol/m2/s and a photoperiod of 16L:8D.
In these experiments, 1-mL water samples were collected at different times for TC analysis. All the experiments were conducted in triplicates.
TC and tet gene removal experiment in microcosm-scale CWs in the long-term
To examine the sustainable removal of TC and tet genes by CWs, sequencing batch experiments were conducted. The CW pot was filled with 400 mL wastewater containing 250 μg/L TC and was incubated for 2 days. After 2 days, all the wastewater was discharged from the CW pot. This sequencing batch experiment at 2-day treatment time was repeated 14 times for 28 days. The effluent samples from both the planted and non-planted CWs collected in each batch were subjected to TC analysis, but only planted CW effluents were used for tet gene quantification. The sequencing batch experiments were conducted in a greenhouse in duplicate.
TC analysis
TC in the collected water samples (300 mL for microcosm-scale CW experiments; 1 mL for flask experiments) were extracted following the USA EPA Method 1694 (2004), with some modifications. In brief, the collected water sample was filtered through 0.22-µm pore size membrane filters (Millipore, Ireland) immediately after collection. The filters withheld the microorganisms in the effluent sample and were used in DNA extraction and for the quantification of tet genes. The filtrates were acidified to a pH of 2.5 with HCl and were spiked with 200 mg/L EDTA·Na2. Thereafter, they were passed through Oasis HLB
extraction cartridges (6 cc, 200 mg) at a flow rate of 5 mL/min. TC was eluted from the cartridge with 1 mL mixture of acetonitrile/75 mM oxalic acid solution (45:55, v/v) and was quantified by high performance liquid chromatography using a UV detector (HPLC/UV). The HPLC/UV analyses were performed using a mixture of acetonitrile/0.01M phosphate buffer (20:80, v/v) as the mobile phase. The wavelength for UV detection was 365 nm. The recovery in the method employed was in the range of 89– 95%, and the limit of detection was 0.0025 µg/L.
29
DNA extraction and quantification of tet and 16S rRNA genes
The membrane filters (0.22 µm pore size) from the above TC analysis method were used for DNA extraction using the NucleoSpin Soil Kit (TakaraBio, Shiga, Japan), according to the manufacturer’s instructions. In this study, we assessed the expression of five tet genes, belonging to two groups: the efflux pump gene tetC and the ribosome protected genes tetM, tetO, tetQ, and tetW. The five tet genes were chosen for monitoring because they are commonly found in wastewater and aquatic environments (Daghrir et al., 2013; Kummerer, 2004; Thomas et al., 2003Chee-Stanford et al., 2001). The five tet genes and 16S rRNA gene were quantified by quantitative PCR (qPCR) using Thermal Cycler Dice RealTime System II, model TP900/960 (TakaraBio). The qPCRs were conducted in 25-μL reaction mixture, which consisted of 12.5 μL SYBR Green Master Mix (TakaraBio), 0.5 μL of each primer (Table 1), 1 μL template DNA, and 10.5 μL ddH2O. The qPCR
temperature program included an initial denaturation at 95°C for 1 min, followed by 40 cycles of 95°C for 10s, annealing at different temperatures (Table 1) for 30s, and an extension at 72°C for 20s. After each qPCR, a melt curve analysis was conducted by increasing the temperature from 65 to 95°C to verify the specificity of amplification. A standard curve for each gene was created by using custom synthesized plasmid carrying the target sequence. All the qPCRs were performed in triplicate.
Statistical analysis
The averages and standard deviations (SD) of all data were calculated, and all the results are expressed as mean ± SD. Significance (P < 0.05) was assessed using the t-test.
30
Table 1. Target genes used for real time PCR (qPCR) analysis, their primer sequences, and annealing temperatures
Target genes Primer Sequence (3ʹ–5ʹ) Annealing
Temperature (ºC) Reference
16S rRNA 341F
534R
5ʹ-CCTACGGGAGGCAGCAG-3ʹ
5ʹ-TACCGCGGCTGCTGGCAC-3ʹ 60 Bru et al., 2008
tetC TetC-FW
TetC-RV
5ʹ-GCGGGATATCGTCCATTCCG-3ʹ
5ʹ-GCGTAGAGGATCCACAGGACG-3ʹ 68 Aminov et al., 2004
tetM TetM-FW
TetM-RV
5ʹ-ACAGAAAGCTTATTATATAAC-3ʹ
5ʹ-TGGCGTGTCTATGATGTTCAC-3ʹ 57 Aminov et al., 2004
tetO TetO-FW
TetO-RV
5ʹ-ACGGARAGTTTATTGTATACC-3ʹ
5ʹ-TGGCGTATCTATAATGTTGAC-3ʹ 60 Aminov et al., 2004
tetQ TetQ-FW
TetQ-RV
5ʹ-AGAATCTGCTGTTTGCCAGTG-3ʹ
5ʹ-CGGAGTGTCAATGATATTGCA-3ʹ 63 Aminov et al., 2004
tetW TetW-FW
TetW-RV
5ʹ-GAGAGCCTGCTATATGCCAGC-3ʹ
31
RESULTS AND DISCUSSION Potential for TC removal by CWs
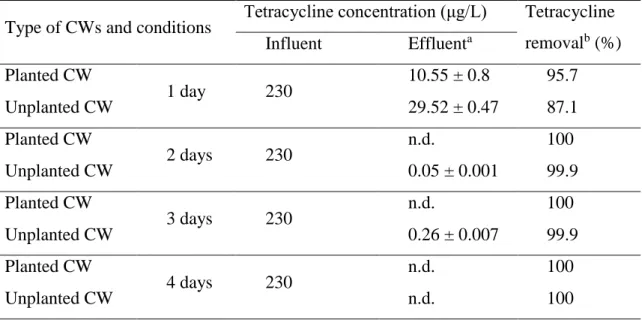
To examine the effect of wastewater retention time on TC removal abilities of CWs, TC removal experiments with planted and non-planted CWs were conducted for different treatment times of 1, 2, 3, and 4 days. The concentrations of TC in the influent and effluent waters from each CW are summarized in Table 2. In CWs with 1 day treatment, 95.4 (remaining TC in the effluent, 10.55 μg/L) and 87.2% (remaining TC in the effluent, 29.52 μg/L) of TC were removed from the contaminated wastewater in the planted and non-planted CWs, respectively. In the other words, there was a significant difference (P<0.05) in TC removal rate between planted CW (219.45 μg/L/day) and non-planted CW (200.48 μg/L/day).
Table 2. Concentrations of TC in planted and unplanted CWs with different
TREATMENT TIMEs
Type of CWs and conditions Tetracycline concentration (μg/L) Tetracycline removalb (%) Influent Effluenta Planted CW 1 day 230 10.55 ± 0.8 95.7 Unplanted CW 29.52 ± 0.47 87.1 Planted CW 2 days 230 n.d. 100 Unplanted CW 0.05 ± 0.001 99.9 Planted CW 3 days 230 n.d. 100 Unplanted CW 0.26 ± 0.007 99.9 Planted CW 4 days 230 n.d. 100 Unplanted CW n.d. 100 n.d., not detected
a Data are the mean of values obtained in duplicate experiments (n=2)
b Tetracycline removal was calculated using the mean values of tetracycline concentration in influent and effluent.
Because of this high removal rate, when TC-containing WW was retained longer in the CWs, from 2 to 4 days, almost 100% of the TC was removed in both the planted and non-planted CWs. The results indicate that TC was rapidly and significantly removed by
32 planted and non-planted CWs. The vegetation of P. australis plants enhanced the TC removal from wastewater in CWs. These results also agree with previous studies. The presence of aquatic plants, such as P. australis (Liu et al., 2014; Carvalho et al, 2013) and
Hybrid Pennisetum(Liu et al., 2013), can be beneficial the effective CWs for TC removal from wastewaters.
Mechanisms of TC removal by CWs
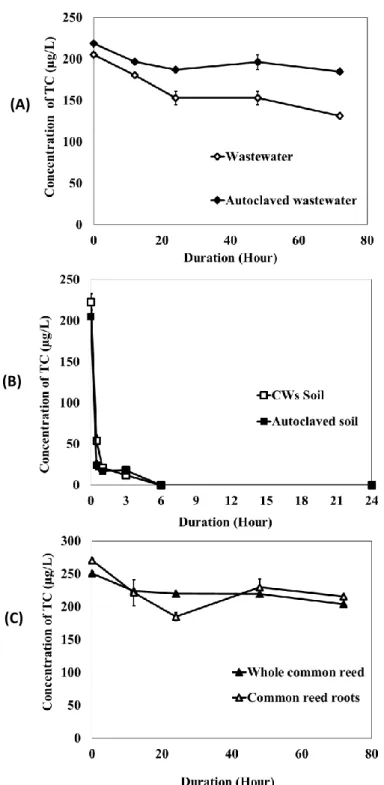
Organic chemicals flowing into CWs can be removed by several factors, such as through biodegradation by microorganisms, by adsorption to solid matrix, through phytodegradation and/or through uptake by plants(Imfeld et al., 2009). However, little is known about the mechanism for removal of antibiotics, including that for TC, by CWs. There is no report describing the comparative evaluation of each factor for TC removal. To completely examine the contribution of each factor, i.e. microorganisms, soil, and plants in the TC removal in CWs, a series of flask experiments was performed using sterile/unsterile wastewater samples, sterile/unsterile CW soils, and sterile plant materials. The results are shown in Fig. 3a, b, c.
In the tests with unsterile/sterile wastewater sample, removal of TC from unsterile wastewater was gradual but significantly more than that from sterile wastewater (P < 0.05), corresponding to 36 and 16% of the TC removed in unsterile or sterile wastewater, respectively, within 3 days (Fig. 3a). TC removal in sterile wastewater might result from TC adsorption to SS in wastewater sample. Although TC persisted in wastewater after 3 days, the biodegradation of TC by native microorganisms in wastewater was significant.
In the tests using soil materials, TC was rapidly and completely removed within 6 h in both the sterile and unsterile soil material tests (Fig. 3b). The results clearly indicate the strong adsorption affinity of red soil materials for TC. Bao et al. (2008) and Guler and Sarioglu (2014) have also revealed the high adsorption ability of red soil and pumice stone for TC. In this flask experiments, TC removal by soil material was more rapid compared to microcosm-scale CW experiments (Table 2). This might result from the differences in experimental/operation conditions between flask and CW experiments.
33 In the TC removal tests using sterile plant materials, the TC concentration was reduced slowly and slightly by both the whole plant and the roots alone; 19 and 20% of TC was removed by autoclave root materials and whole plant, respectively, within 3 days. The
(C) (A)
(B)
Figure 3. Changes in TC concentrations in original wastewater/autoclaved wastewater
experiments (A), in original soil/autoclaved soil material experiments (B), and in sterile whole plant/root material experiments (C). Data are the means of values obtained in triplicate experiments, and error bars indicate the standard deviations.
34 results indicate the adsorption of TC on the root material as the main cause for the TC elimination rather than phyto-degradation and plant uptake.
The TC removal process in a CW system is regulated by various components, such as by microorganisms, soils, and plants. In our experiments, we isolated and evaluated each of these components. The adsorption of TC to soil materials was the main mechanism for TC removal which is attributed to high distribution coefficients (Kd) of TC compounds,
from 290 to 1620 L/kg (Thiele-Bruhn et al., 2013). The biodegradation of TC by native microorganisms present in wastewater was slower than soil sorption but clearly evident. On the other hand, uptake/phytodegradation/adsorption by P. australis plant had a limit effect on TC removal. The results suggest that TC was initially adsorbed to soil material, and was subsequently removed by microorganisms and plant effects. Liu et al. (2013) have also reported that soil materials play a main role in the removal of antibiotics from wastewater in CWs.
Despite the out-performance of soil sorption, in the greenhouse experiment, a better removal of TC by planted CW (95.7%) compared with that by non-planted CW (87.1%) (Table 2) was observed. This result might be a result of the enhanced biodegradation supported with plant. It is a consensus that interface between plant roots and soils is the important area for microbial activity in the wetland system (You et al., 2015). Plants are believed to sustain large rhizosphere associated-microbial populations, and facilitate their growth and metabolism by providing oxygen and organic compounds released from plant roots (Anderson et al., 1993; Shaw et al., 2003). Previous researches reported that planted CWs perform better removal of phenol (Tee et al., 2009), bisphenol A and 4-tert-butylphenol (Dan et al., 2017) than the non-planted CWs because of enhanced biodegradation by plants. Further studies are desired to clarify the effects of enhanced biodegradation supported by plants on TC removal in CWs.
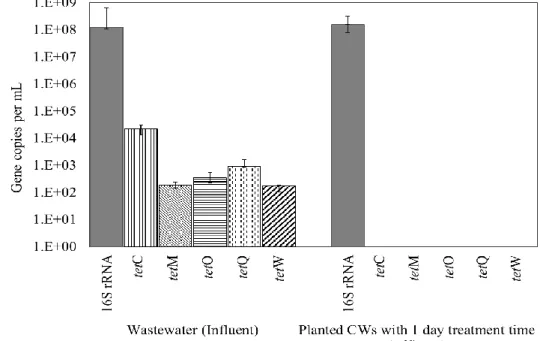
Removal potential of tet genes by CWs
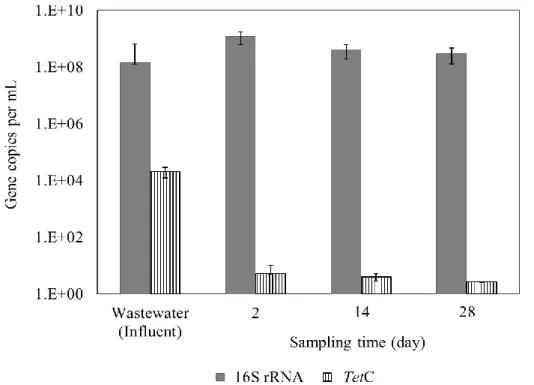
The absolute abundances of 16S rRNA and tet genes (tetC, tetM, tetO, tetQ, and tetW genes) in the influent wastewater and effluents of planted CWs with 1 day treatment are shown in Fig. 4. All the genes were detected in the influent wastewater; the copy numbers of 16S rRNA, tetC, tetM, tetO, tetQ, and tetW genes were 1.2 × 108, 2 × 104, 1.8 × 102, 3.4 × 102, 9.2 × 103, and 1.7 × 102 copies/mL, respectively. The results confirmed that
35 secondary treated wastewater of municipal wastewater treatment plant was a source of the
tet genes (Rizzo et al., 2013; Gao et al., 2012; Michael et al., 2013). After the planted CW
treatment for 1 days, none of the tet genes were detected in the effluent samples. The removal efficiencies of tetC, tetM, tetO, tetQ, and tetW genes were about >4.3, >2.2, >2.5, >3.9, or >2.2 log units. In contrast, the 16S rRNA gene (1.3 × 108) was detected in the effluent samples at the same level found in the influent wastewater. In planted CWs with longer treatment times from 2 to 4 days, none of tet genes were detected in the effluent samples. The results confirmed that the planted CW significantly and selectively removed
these tet genes from the wastewater. A few researchers have reported significant removal of tet genes and other ARGs by CWs. Liu et al. (2013) have reported about one log units removal of tet genes in vertical flow CWs. Chen et al. (2015) have reported about 2‒3 log units removal of tet genes in integrated CWs. Huang et al. (2015) have reported 0.26‒3.3 log units removal of tet genes in vertical up-flow CWs. Removal efficiency of tet genes in our planted CWs was comparable to those reported in the previous researches.
The removal efficiencies of tet genes by our CWs and those of the previous CWs are comparable to the removal efficiencies of ARGs, such as tetG, by chlorination (3.24 log reductions), ultraviolet (2.48 log reductions), and ozonation (2.55 log reductions)
Figure 4. Absolute abundances of 16S rRNA and five tet genes (tetC, tetM, tetO, tetQ,
and tetW) in influent (wastewater sample) and effluent in planted CWs with treatment time of 1 days. Data are the means of values obtained in duplicate experiments, and error bars indicate the standard deviations.
36 processes (Zhuang et al., 2015). The results clearly indicated that CWs appear to be an advanced treatment process to prevent the discharge of large amount of tet genes from the wastewater treatment plants. Chen et al. (2016) reported that the removal of tet genes in CWs was likely due to several factors including microbial activities, soil adsorption, and plant uptake like that of antibiotics. However, the results of the present as well as previous studies do not clearly reveal the removal mechanisms of tet genes in CWs. Further research is needed to explore the fate and removal mechanism of ARGs in CWs.
TC and tet genes removal by CWs in the long-term experiments
To examine a stable long-term removal of TC and tet genes from wastewater by CWs, sequencing batch experiments to treat wastewater containing TC (250 μg/L) were conducted. The concentrations of TC in the effluent samples from the planted and non-planted CWs are shown in Fig. 5. During the 28-days experimental period, the TC concentrations in the effluent of the planted CWs were significantly lower than those of the non-planted CWs. In the planted CWs, TC was completely and repeatedly removed from wastewater during the 28 days. In contrast, the TC concentration in the effluent of
Figure 5. TC concentrations in the effluent of planted CWs during a month of
long-term experiments under sequencing batch mode. The TC concentration in the influent was 230 μg/L. Data are the means of values obtained in duplicate experiments, and error bars indicate the standard deviations. Day 0 = May 23, 2016 to Day 28 = Jun 20, 2016.