STUDIES ON NUTRITIONAL NITROGEN
FROM RED CLOVER SILAGE*
Mitsuaki
OHSHIMA
Contents
Preface 1
Background of the study 2
Chapter 1 The nitrogen composition of red clover herbage and silage 4 Chapter 2 Daily change of free amino acids during ensilage 15 Chapter 3 The nutritive value of proteins in red clover herbage and silage 38
Conclusion 60
Acknowledgement 6 1
References 62
Japanese summary 67
Preface
Recently, resulting from a n improvement in eating habits, the Japanese requirement for animal foods has yearly grown in amount This has been accompanied by a n increase in the number of farm animals, making it necessary to obtain more feed. Since the feed supply could not keep up with the demand for feedstuffs harvested in our country, a large amount of concentrated feeds have been imported from foreign countries a t a high and variable cost T o establish the stabilization of the management of animal husbandry in Japan, self-supplying of feed is most important, and research to make use of forage crops as feeds of both omnivorous and carnivorous animals is required Moreover, in spite of being short, a large part of the protein foods which are eatable for man are used as animal feeds One of the solutions to the problem is to preserve protein rich herbages such as legumes for winter use. For this purpose, hay or silage is made all over the world But in Japan, hay making is very difficult because of the moist and rainy weather Therefore, the preservation of a green crop as silage is a very important matter When herbage is ensiled, a rapid breakdown of leaf protein to lower molecular non-protein nitrogenous
*
This work was performed at the Laboratory of Animal Nutrition, Nagoya University, the Laboratory ofAnimal Husbandry, Kagawa University, and the Isotope Research Center of Kagawa University Parts of the study were published in "The Japanese Journal of Zootechnical Science", 36: 488 (1965), 37: 1 (1966), 39: 402,462 (1968), 41: 97 (1970).
2
compounds occurs The efficient utilization of non-protein sources of nitrogen by rumi- nant animals depends greatly upon the nature of these sources Furthermore, there has been speculation that the unpalatability of certain silages might be related to changes in the components of the soluble non-protein nitrogen fraction74) Compared to ruminant animals, non-rumina.nt animals have fewer microorganisms in their digestive tract and consequently can use far less non-protein nitrogens as their dietary nitlogen source Fur- thermore, if amino acids exist in quantities in seepage, a large loss of nutrient occurs But, there have been very few detailed investigations on the nature of the non-protein nitrogen fraction in silage Therefore, studies on the course and extent of the proteolysis during the ensilage and on the nutritive value of residual protein are required to minimize nut- rient loss The purpose of this investigation is to explore these problems.
The author also wishes to establish the value of herbage as a dietary protein source which, to date, has been lightly treated in animal nutrition studies As most commercial poultry feed contain about 5% alfalfa meal containing 17% or more protein, poultry feeding on these rations are supplied with about 5% of their day's protein requirement from alfalfa meal The author previously reported on the nutritive values of the proteins from various herbages for chicks78) It is well known that pigs consume a considerable amount of herbage when they are given it71*93*lll) Therefore, we can not neglect the study of the nutritive value of nitrogen in herbages for non-ruminant animals
Background of the Study
As there are many reviews56~101~106) on the history of silage investigation, the author wishes to mention here only those studies concerned with nitrogens in silage
Harvesting of a forage crop is followed by rapid and extensive proteolysis which is only terminated either by the attainment of a high dry matter or a low pH Even in well preserved silage, about 50 - 60% of the protein is broken down to non-protein nitrogenous compoundsg, 33,44.49,50,83) Main components of the non-protein nitrogens are amino acids and volatile basic nitrogens The greatest protein breakdown is due to slow lactic fermentation and the slower pH fall in early stages of the ensilage processg). Since the fresh crop generally contains about 80 - 90% of its nitrogen in the form of protein, large amounts of amino acids are released by autolysis or bacterial digestion44). I n high pH
silages the ammonia may reach an extremely high figure accounting for 50% or more of the total nitrogen44) The ammonia is formed by the action of clostridial bacteria which attack the amino acidsl06) Deamination by lactic acid forming bacteria appears to be restricted to sefine and argininel06) No free ammonia appeared when microorganism free grass was ensiled, suggesting that the formation of ammonia from amino acids during ensilage is a function of the anaerobic bacteria present44) A great number of free amino acids were partially degraded during ensilage but alanine and aminobutyric acid gradual- ly increased44.102) As far as the author knows, there are very few workers investigating the details of deamination which occurs during ensilage The decarboxylation products of the amino acids are the amines and the other amino acids The amines, histamine,
tryptamine, tyramine, cadaverine and putrescine are the decarboxylation products of histidine, tryptophan, tyrosine, lysine and arginine, respectively, have been detected in silages30?52-54,72-74) The conditions in silage affecting amine formation have been stud- ied and explored and the most notable condition perhaps effecting the production of these amines has been the rate of p H fall during ensilage rather than the final p H value which has been known as a condition affecting clostridial activity54) But, on the contrary, tyramine was found in clover silage after 24 hours of incubation at 39 OC and it was specu- lated that this amine was formed by enzymic decarboxylation of tyrosine rather than as a result of bacterial activity73) Ethanolamine22,30), 5-hydroxytr yptamine72) and
P-
phenylethylamine30) were also found in some silages Thirteen different volatile amines were found in 14 kinds of silage# Among 13 amines, trymethylamine and p-pheny- lethylamine were present in larger amounts and the other amines were in smaller amounts When microbe-free grass was conserved anaerobically, the formation of a considerable amount of r-aminobutyric acid was found51) Beta-alanine and 8-aminovaleric acid were found in some silages, but not in good silages30~41~42) O n the contrary, P-alanine was detected as a usual component of herbage cropslg) Alpha-aminobutyric acid was found in anaerobic slurries of kale after 147 days conservation89) and was also found in bad silage 8 weeks after ensiling44) The protein remaining after silage fermentation was little different in mono-amino acid composition from the original grass proteins43)Much work has been done on the nitrogen metabolism of microorganisms, but, as far as the author knows, the process of ensilage and the manner by which the above men- tioned changes in the nitrogenous compounds occur, remain unsolved. T h e present experiment was carried out to clarify the above mentioned unsolved problem, especially the amino acid metabolism in silage
I t goes without saying that there are many workers studying the nutritive value of silage for ruminants But, as far as the author knows, very few nutritional studies on si- lage for monogastric animals have been made Thirty per cent of a growing pigs rations could be replaced with corn silage62). About 40% of a growing pigs rations could be replaced with ladino clover silage by supplying sufficient energy from oilsll0) When mature and growing pigs were given minced corn silage, they ate 5-6 kg and 2 5-3kg of the silage per day, respectively42) The growing pigs given 0 5-1 Okg of minced grass silage and nearly the same amount of fresh grass showed better growth than those on the control ration99) The growing pigs given a protein-rich silage in addition to the basic ration showed better growth than those given the basic ration without the silage55) The palat- ability of fresh and ensiled Russian comfrey was fairly good70). When 50 breeding White Leghorn hens were given 25-100 g of silage, they showed good egg production and high hatchability and consumed carotene from the silage85) The same good results on egg pro- duction were also obtained when 240 hens were given a diet containing 50 g of silage4) T h e opposite results were also reported when the addition of silage to rations of laying hens did not improve egg production, and the consumption of laying mash only slightly reduced68) I t has been shown that the optimum consumption levels of silage for various types of poultry is as follows: 50-60 g for layers, 3-5 g for 20-30 days old chicks, 20-30 g
for 2-3 months old chicks, 200-250 g for ducks, 150-200 g for turkeys and 200-300 g for geese26) But it is considered that there are some nutrient losses of nitrogens when silage is given to monogastric animals because of nutritionally unavailable nitrogens in silage which increase during ensilage as mentioned above I t was reported that the digestibility f11r fowl of crude- and pure-protein of sweet potato leaf silage was about one half and one fifth, respectively, when compared to that of sweet potato leaf69). I t remains uncertain whether these lowerings of digestibility were caused by a physiological abnor- mality produced by some silage components or by the indigestibility of silage protein I n order to know the real value of silage as a nitrogen source for monogastric animals, it is necessary to study in detail the water-soluble non-protein nitrogenous compounds because some of them may have toxic activity for animals and may be easily absorbed from intes- tine, and to study the nutritive value of residual protein There is a report on the phys- iological effect of silage When rats were fed dried silage, they showed an increased basal metabolic rate and a heightening of the follicular epithelial cells of the thyroids*).
Chapter 1
The nitrogen composition of red clover herbage and silage
While it has been known for some time that only about 30% of the total nitrogen in grass silage could be accounted for as protein nitrogen there have been very few detailed investigations on the nature of the non-protein nitrogen fraction The efficient utilization of non-protein sources of nitrogen depends largely on the nature of these sources. Fur- thermore, there has been speculation that the unpalatability of certain silages might be related to changes in the components of the soluble non-protein nitrogen fraction Com- pared to graminivorous animals, omnivorous animals have fewer microorganisms in their digestive tract and consequently can use far less non-protein nitrogens as a nitrogen source Furthermore, if amino acids exist in quantities in seepage, a large loss of nl~trient may occur Therefore, studies on the course and extent of proteolysis during ensilage are required to minimize nutrient loss and to make silage useful feed for omnivorous animals. The purpose of this investigation was to explore these problems.
Besides the silage, the nitrogen composition of fresh red clover was also studied This work has been made not only to compare the nitrogenous compounds of herbage to those of silage, but also to establish their values in practical use
The reason why red clover was used as the experimental material was to obtain pro- tein-rich self-sufficing feeds
Materials and Methods
1 Herbage and silage.
T h e herbage used in this experiment was red clover in the full bloom stage harvested on May 10 and 12, 1964, a t Aichi Prefectural Livestock Breeding Farm (Okazaki)
Twenty three point five kilograms of fresh red clover was wilted on the field with oc- casional turnings for 3 hours The weight of the wilted clover was reduced to 20.3 kg.
I t was chopped into about lcm lengths and mixed well with 2% glucose and then packed into four bags of polyethylene film (0 08 mm thick, 40 cm in diameter, and 60 cm in height) weighing 4 4 kg each The air in the packed bag was removed by sucking with a water aspirator for 5 minutes, and as a result, the intracellular liquid of red clover was squeezed through the cell membranes by the surrounding pressure The aspirated bags were tied with linen thread and then buried under the ground until July, 28, 1964
2 Fractionation of herbage and silage into the three parts of plant structure
The fresh red clover was divided into the three parts of plant structure using the foi- lowing method. Using a blender, l OOg of chopped fresh red clover was ground in the pres- ence of 300 ml of a 0 9% NaCl solution containing 0 1
%
ascorbic acid as an antioxidant. During grinding, the cup of the blender was immersed in a cold water bath so as to main- tain the contents a t a low temperature The macerate was separated into a solid and a liquid fraction through a hemp cloth by a hand squeezing operation The liquid frac- tion was stored in a freezer a t -20°C until it was used Prior to analysis, it was melted by running tap water and filtered through No 6 Toyo filter paper using a Buchner funnel attached to a filtering flask The flask was sucked continuously with a water aspirator. The resultant precipitate was referred to as the concentrated part, and the filtrate as the water .soluble part The fibrous and the concentrated parts were dried a t 60 O C in a forced draft oven, then weighed and finely ground The water-soluble part was stored in a freez- er immediately after filtrationThe silage was also divided into the three paits of plant structure using the methods mentioned above, the only exception was that water was used instead of saline.
3 Preparation of samples for amino acid analysis
The hydrolysis of the concentrated parts of the herbage and silage were carried out under the conditions described below About 0 lg of the concentrated part was placed in a 2ml ampule with lml of 6N-HC1, then the ampule was attached to an aspirator and closed up under vacuum The enclosed umpule was heated to 1 lO&O 1 "C for 24 hours. The cooled tube was opened and placed in a vacuum descicator containing NaOH pel- lets. Most of the excess HC1 was removed overnight The pH of the hydrolysate was adjusted to about pH 2 by adding N-NaOH. Then the solution was transferred into a 50 ml volumetric flask and made up to volume with pH 2 2 citrate buffer
The water-soluble non-protein nitrogen fractions of the herbage and silage were made by the method described by Stein and MOO RE^^) Ten milliliters of the water-soluble part was placed in a flask and 50 mI of 1
%
picric acid was added The content was then mixed thoroughly by vigorous stirring, and the suspension was centrifuged for 10 minutes a t 4,000 r p m Fifty milliliters of the supernatant liquid was passed through a column of Dowex 2 x 8 in hydrogen form (column size 2-by-20-cm, resin height 2 cm) to remove the excess of picric acid The column was washed with five 3 ml portions of 0 02N-HC1 The effluent and washings were collected in a 100 ml Petri-dish and then the dish was placed in a desicator containing conc-H2S04, with an incandescent bulb attached to the inner part of the lid The desicator was closed and evacuated continuously with a n efficient aspirator until the contents of the dish was reduced to 2 ml or less The concen-trated solution was adjusted to p H 2 by adding N-NaOH, and then transferred to a 20 ml volumetric flask and made up to volume with p H 2 2 citrate buffer
The hydrolysis of the low molecular peptides in the deproteinized water-soluble part was made in the same way as that of the concentrated part One milliliter of water-sol- uble part was used instead of 0 1 g of concentrated part
4 Analysis of amino acids
The quantitative analysis of the amino acids of hydrolysates was made by the method of MOORE et a1 66) using a Beckman-Spinco automatic amino acid analyzer Model-120 The resin used was Amberlite IR-120 in sodium form A 1
x
150 cm column was used to analyze the acidic and neutral amino acids, and a 1x
15 cm column of resin was used in the analysis of the basic amino acidsThe contents of free amino acids from the water-soluble non-protein nitrogen frac- tion was measured by the method of SPACKMAN et a1 97) using the automatic amino acid analyzer A 1
x
150 cm and a 1 x 50 cm column of Amberlite IR-120 were used to ana- lyze the acidic and neutral amino acids and the basic amino acids, respectively5 The analysis of ammonia, amide, and nitrate nitrogen in the water-soluble part For the quantitative determination of ammonia, amide, and nitrate nitrogen in the water-soluble part, the method of VARNER et a1 103) was used with minor modification One portion of the water-soluble part containing 5 mg nitrogen was mixed with two por- tions of ethanol and then centrifuged The supernatant liquid was transferred into a Kjel- dahl distilling flask through the funnel The part of the sample which adherred to the side of the funnel was washed in with 5 ml of a saturated sodium borate solution. The mixed solution was then steam-distilled a t 100°C for 3 minutes exactly, the distillate being collected in a few milliliters of 2% boric acid solution was titrated with 0 01N-H2S04 This fraction was regarded as ammonia Then 15 ml of 40% NaOH was added to the distilling flask and the distillation was continued for 10 minutes The nitrogen which was distillated was referred to as an amide nitrogen Then, 5 ml of 20% FeS04 and 5ml of saturated AgS04 solution were added to the distilling flask, and the distillation continued for 20 minutes to remove nitrite and nitrate nitrogen
6 The determination of the p H value and the organic acid content of silage.
The p H of silage was determined by a Hitachi-Horiba p H meter HM-5 The deter- mination of the organic acid content of the silage was carried out by the method of SHIBATA and K I K U C H I ~ ~ ) using a Yanagimoto gaschromatograph GCG-220
Results
The chemical composition of the red clover herbage and silage are shown in Table
1 The fresh herbage contained more crude protein and less crude fat than the silage. The organic acid content and the p H value of the silage are shown in Table 2 The p H value of these silages ranged from 4 00 to 4 30 T h e main constituent organic acids of the silage were lactic acid followed by acetic acid I n the silages of lower p H butyric acid existed in only small amounts but large amounts of butyric acid were found in the si- lage of p H 4 30, and all were evaluated as being excellent or good according to their or-
Table 1 Chemical composition of the fresh red clover and of the ensiled one calculated on dty matter basis
-- --A
~ i t r o ~ ~ n
Crude Crude Crude Crude
free
protein fat fiber ash
extract
Yo
% % % YoFresh herbage 174 7 3 43 9 21 8 9 7
Silage 15 4 8 9 43 3 22 9 9 5
ganic acid composition92). A subjective evaluation of silages in appearance and odor
Table 2 The pH value and organic acid content of the juice from the red clover silage
--
-sample
I
pH
I
Organic acid/
pointI
Gradewas also fairly high
T h e distribution of nitrogen in the three parts of plant structure of both herbage and silage were studied and the results are shown in Table 3. I n the herbage, 27% of the ni- trogen remained in the fibrous part, 44% in the concentrated part and 29% in the water-
Excellent
Excellent
Good
Table 3 Distribution of nitrogen in the three parts of plant structure of red clover herbage and silage exhibited in mg per 100 g of wet materials
- Herbage Silage Fibrous part 1392 (269)* 1644 (29 4)* Concentrated part 229 2 (44 4) 81 2 (14 5) Water-soluble part 1484 (28 7) 313 2 (56 1) Non-protein nitrogen 124 0 (24 0) 313 2 (56 1) Protein nitrogen 24 4 ( 4 7) 0 0 ( 0 0 ) Total 516 8(100 0) 558 8(100 0) 25 - 25 - 4 5 9 5 25 - 25 -- 45 9 5 2 5 - 25
-
30 80 1 2 3 pp - -*
Figure in parentheses indicates a percentage 4 00 23 30 mg/dl Yo Lactic 3315 6 76 2 Succinic 286 7 6 6 Acetic 719 4 16 6 Propionic 8 9 0 2 Butyric 18 5 0 4 Total 4349 1 100 0 Lactic 2794 5 70 5 Succinic 424 8 10 7 Acetic 718 8 18 1 ~ropionic 4 4 O 1 Butyric 24 6 0 6 Total 3967 1 100 0 Lactic 2369 7 71 7 Succinic 345 7 10 4 Acetic 404 7 12 1 ~ropionic 25 9 0 8 Butyric 188 3 5 6 Total 3334 0 100 0soluble part Therefore, about 70% of the nitrogen was solid in form and was considered to be protein nitrogen P I R I E ~ ~ ) mentioned in his review of leaf proteins that most of the nitrogen in the fibrous part comes from entrapped chloroplasts Of the water-soluble nitrogen, only 16 5% was protein nitrogen which was precipitable with 1% picric acid and the remainder was considered to be non-protein nitrogen The distribution of nitro- gen in the fibrous part of the silage corresponded closely to that same part in the fresh herb- age But, the nitrogen distribution in the concentrated part of the silage was only one third that of the fresh herbage, and this lost nitrogen appeared in the water-soluble part The most distinguished difference between the herbage and the silage was observed in the water-soluble protein nitrogen content.
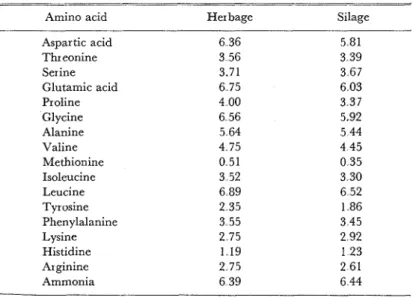
In spite of these facts, there was very little difference in the amino acid composition between the concentrated parts of the herbage and of the silage as shown in Table 4
Table 4 Amino acid composition of proteins in the concentrated parts of red clover herbage and silage exhibited as percent of a-amino acid nitrogen in
total nitrogen
-
Amino acid Herbage Silage
Aspar tic acid 6 36 5 81
Thr eonine 3 56 3 39 Ser ine 3.71 3 67 Glutamic acid 6 75 6 03 Proline 4 00 3 37 Glycine 6 56 5.92 Alanine 5 64 5 44 Valine 4 75 4 45 Methionine 0 51 0 35 Isoleucine 3 52 3 30 Leucine 6 89 6 52 Tyrvsine 2 35 1 86 Phenylalanine 3 55 3 45 Lysine 2 75 2 92 Histidine 1 19 1 2 3 Arginine 2 75 2 61 Ammonia 6 39 6 44
Ammonia-, amide-, nitrate-, amino-, and peptide-nitrogen of the water-soluble parts of the red clover herbage and silage were determined and the results are shown in Table
5 The water-soluble part of the silage contained fourteen times as much ammonia-nitro- gen and six times as much amino-nitrogen as the fresh herbage I n contrast, amide- and nitrate-nitrogen were decreased during ensilage and no peptide was measurable in the si- lage
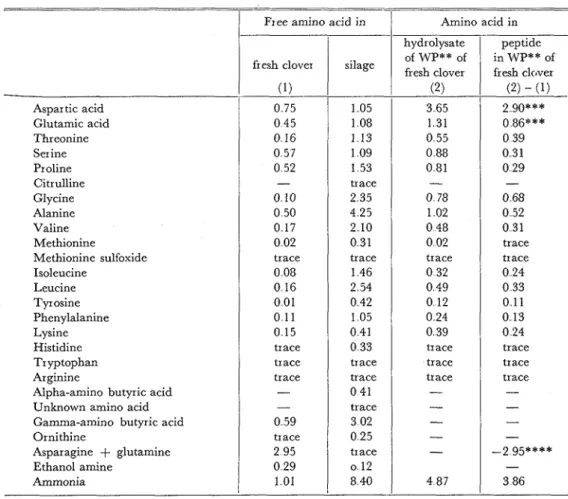
The composition of free amino acids and the amino acid composition of the lower molecular peptide in the water-soluble parts of the red clover herbage and silage were determined and the results are presented in Table 6 The amino acid composition of pep- tide was determined by subtracting the value of free amino acids from that of the hydro- lysate of the deproteinized water-soluble part. I n the silage, since the amino acid corn-
Total
1
9 5 7/
8 4 31
8 2 3/
718*
Non-protein nitrogenTable 5 Nitrogen composition of red clover herbage and silage
Table 6. Free amino acids and amino acid copmosition of peptide* in the water-soluble parts of red clover herbage and silage, exhibited as percent of a-amino N in total N
Free amino acid in
I
Amino acid inI
1
I
hydrolysate Ammonia - N Amide - N Nitrate - N Amino - N Peptide - N Protein - N1
freshclover1
silageI
of WP** of fresh clover Per total NPN* Herbage % 4 2 31 3 10 4 18 1 18 3-
Per total N Herbage1
Silage peptide in WP** of Silage Yo 25 6 trace 1 7 44 5 0 0 - % 1 0 7 5 2 5 4 3 4 4 76 0Aspar tic acid Glutamic acid Threonine Ser ine Proline Citrulline Glycine Alanine Valine Methionine Methionine sulfoxide Isoleucine Leucine Tyrosine Phenylalanine Lysine Histidine Tryptophan Arginine
Alpha-amino butyric acid Unknown amino acid Gamma-amino butyric acid Ornithine Asparagine
+
glutamine Ethanol amine Ammonia fresh clover (2) - (1) 2 go*** 0 86""" 0 39 0 31 0 29 % 14 4 trace 1 0 25 0 0 0 43 9 - 0 68 0 52 0 31 trace*
No peptide was found in the silage** The water-soluble part
* * *
Containing corresponding free amide (possibly)****
These amides may be converted to corresponding amino acids. 0 75 0 45 0 16 0 57 0 52 - 0 10 0 50 0 17 0 02 trace 0 08 0 16 0 01 0 11 0 15 trace trace trace - - 0 59 trace 2 95 0 29 101 trace 0 24 0 33 0 11 0 13 0 24 trace 105 1 08 1 13 1 09 1 53 trace 2 35 4 25 2 10 0 31 trace 146 2 54 0 42 105 0 41 0 33 trace trace 0 41 trace 3 02 0 25 trace o 12 8 40 trace trace 3 65 131 0 55 0 88 0 81-
0 78 1 02 0 48 0 02 trace 0 32 0 49 0 12 0 24 0 39 trace trace trace-
-
-
-
-
4 87position of the water-soluble part and its hydrolysate was exactly the same, it seemed that there was no peptide in the water-soluble part of the silage The water-soluble part of the fresh herbage contained a high concentration of aspartic acid in free form, and contain- ed a large quantity of amides such as asparagine and glutamine Since the amides con- verted to the corresponding monoamino dicarboxylic acids, the hydrolysate of the water
-
soluble part of the fresh herbage contained no amide but did contain a large amount of aspartic and glutamic acids From the data shown in Table 6, more than 2 9% of the total nitrogen is aspartic acid nitrogen in water-soluble peptide But it is easily consider- ed that a large percentage of it may have been derived from asparagine in the free form O n the other hand, glutamic acid which was detected in thc peptides was only 0 9% of the total nitrogen Although asparagine and glutamine in the water-soluble part could not be separated by the present procedure, it is supposed from the above fact that the great- er part of the amides in the fresh red clover consisted of asparagine There was no relation- ship between the amino acid composition among the protein, the lower molecular peptide, and the free amino acids of the fresh red clover. O n the other hand, in the silage, the com- position of the free amino acids of the water-soluble part paralleled the amino acid com- position of the protein in the concentrated part except for a few amino acids Of the free amino acids in the silage, alanine was the largest component, followed by y-aminobutyric acid, leucine, glycine, and valine The silage contained a-aminobutyric acid, citrulline and an unknown amino acid (perhaps /3-aminoisobutyric acid) which were not present in the fresh red cloverDiscussion
A silage made from red clover was compared with a corresponding fresh herbage in its chemical composition. As shown in Table 1, the silage contained 2% less crude protein than the green herbage This may be due to the evaporation of volatile nitrogenous com- pounds of the silage in which is contained a large amount of ammonia and perhaps amines. The silage contained a little more crude fat than the herbage This result may be attribu- table to the organic acid synthesis of the silage I n spite of adding 2% glucose per fresh weight before ensiling, the silage showed a lower content of nitrogen free extract than the herbage. This can be explained from the well known fact that microorganisms convert nitrogen free extract to organic acids during fermentation in silage
A comparative study was made on the nitrogenous compounds of the silage and herba- ge I n the herbage 76% of the total nitrogen was protein nitrogen (27% in the fibrous part, 44
%
in the concentrated part, and 5% in the water-soluble part) and 24% was non- protein nitrogen These results' showed good agreement with the previous data77) obtain- ed on red clover which were cultivated at a different place and harvested at the same stage of maturity But in the silage only 44% remained as protein nitrogen in form and 56% of the total nitrogen was water-soluble non-protein nitrogen O H Y A M A ~ ~ ) reported that the most dominant form of nitrogen in grass silages was water-soluble non-protein nitrogen the amount of which ranged from 50 to 60%, and protein nitrogen was 30 to 40% of the total nitrogen This figure of the protein nitrogen appeared much lower than that of fresh229
-
81dried herbage From the data in Table 3, it is calculated that about 70%
(----)
229 the protein in the concentrated part was degraded during ensilage while the protein in the fibrous part was not degraded because of firm fixation in the structural material P I R I E ~ ~ ) mentioned in his review of leaf proteins that the greatest component of the leaf protein concentrate (the concentrated part) is chloroplastic protein and most of the nitro- gen in the fibrous part comes from entrapped chloroplasts I n the fresh red clover, 47%
of the total nitrogen was found as the water-soluble protein nitrogen but in the silage no protein nitrogen was found in the water-soluble part This may have been due to degrada- tion of the water-soluble protein to lower molecular nitrogenous compounds during ensilage or the coagulation of the protein by the organic acids and its shifting to the concentrated part SINGER el a1 96) reported that 50% of cytoplasmic protein which is known to be wa-ter-soluble was coagulated when this protein solution was acidified to pH 5 5. O H Y A M A ~ ~ ) also reported that water-soluble protein diminished remarkably when the grass was extract- ed with a p H 1 2 citrate buffer According to BRADY~), the increase in the water-soluble non-protein nitrogen during wilting was accompanied by a decrease in the soluble protein nitrogen
HORIGOME
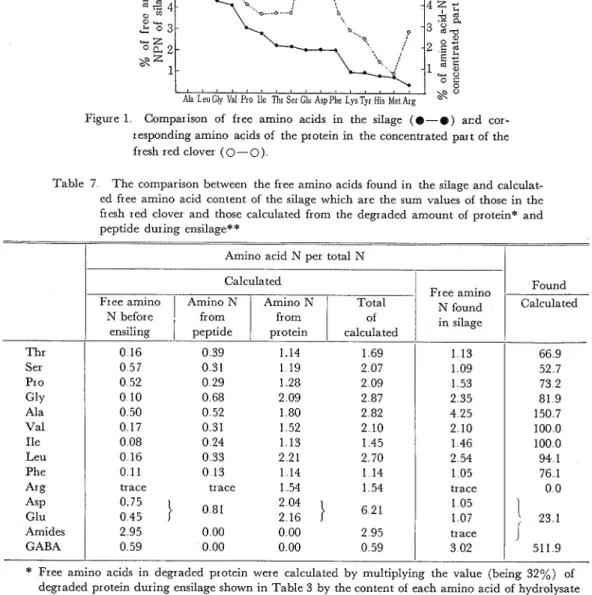
and K A N D A T S U ~ ~ ) found some proteolitic enzymes in the cvtoplasmic and the chloroplastic protein fractions of white clover leaf, and they reported that during incubation of white clover leaf, 32% of the cytoplasmic and 16% of the chloroplastic pro- teins were autodigested. These reports suggest that cytoplasmic protein which is water- soluble may be more easily degraded than chloroplastic protein As shown in Table 5, the green herbage contained Iower molecular peptides at a level of 4 4% of the total nitro- gen in as water-soluble form. while the silage contained no peptide This result suggests that peptide also decomposed during ensilage But BRADY~) reported that changes in the peptide nitrogen fraction during ensilage were not considerable I t is considered that the lower molecular water-soluble peptide may be more easily degraded than the water- soluble protein. Therefore, the above difference can be attributable to the difference in the incubation period which was about 80 days in this experiment and 10 days or less in Brady's experimentAs shown in Table 4, proteins from the concentrated parts of the silage and the herbage show almost the same pattern in amino acid composition This fact suggests that each amino acid from proteins of the concentrated part of red clover was liberated in the same degree during fermentation, even though 70% of the protein was degraded The same result was also reportrd by KEMBLE and M A C P H E R S O N ~ ~ ) on 10 amino acids This fact reflects that the free amino acid composition of the water-soluble part of the silage is similar to the amino acid composition from protein of the concentrated part with some exceptions as shown in Figure 1 As an exception, a lesser quantity of glutamic acid, aspartic acid and arginine and a larger quantity of alanine exist in the water-soluble part of silage than in the concentrated part But when the determined free amino acid compositions of the silage are compared with the maximum values possible which were calculated from the expression, F + P + P r ((Ns-Nf)/100), where F is the free amino acid content of the fresh herbage, P is the amino acid content of the lower peptide of the fresh herbage, Pr is that of the concentrated part and Ns and Nf are the water-soluble non-protein nitrogen con-
I
Ala L e u Gly Val Pro Ile Thr Ser Glu AspPhe LysTyr His Met Arg
Figure 1 Comparison of free amino acids in the silage ( 0 - @ ) acd cor- responding amino acids of the protein in the concentrated part of the fresh red clover (0-0)
Table 7 The comparison between the free amino acids found in the silage and calculat- ed free amino acid content of the silage which are the sum values of those in the fresh red clover and those calculated from the degraded amount of protein* and peptide during ensilage**
I
Amino acid N per total N-
tents
(%
of the total nitrogen) of the silage and herbage, respectively, there is observed a large difference between them as shown in Table 7 The larger differences are observed in the amino acids mentioned above as an exception followed by serine But when com- parison is made between the amino acids derived from degraded protein and the deter- mined value, there exists good agreement between them except for the three amino acidsThr Ser Pro G ~ Y Ala Val Ile Leu Phe '4% A ~ P Glu Amides GABA
*
Free amino acids in degraded protein were calculated by multiplying the value (being 32%) of degraded protein during ensilage shown in Table 3 by the content of each amino acid of hydrolysate of the concentrated part**
Such small amino acids were omitted as the calculated values being less than 1% may reflect large errorsCalculated
Free amino Amino N Total
N before of 0 16 0 39 1.14 1 69 0 57 0 31 1 19 2 07 0 52 0 29 1 28 2 09 0 10 0 68 2 09 2 87 0 50 0 52 1 80 2 82 0 17 0 31 1 52 2 10 0 08 0 24 1 13 1 4 5 0 16 0 33 2 21 2 70 0 11 0 13 1 14 1 14 trace trace 1 54 1 54 0'75
}
0 81 2 04 0 45 2 1 6}
6 2 1 2 95 0 00 0 00 2 95 0 59 0 00 0 00 0 59 Free amino N found in silage 1 13 1 09 1 53 2 35 4 25 2 10 1 46 2 54 1 05 trace 1 05 107 trace 3 02 Found Calculated 66 9 52 7 73 2 81 9 150 7 100 0 100 0 94 1 76 1 0 0 ' 2 3 11
511 9This fact suggests that the degradation of amino acids to other substances may take place largely before the protein degradation occurs I n this calculation, the amino acids which were determined to be less than 0 5% of the total nitrogen were omitted because even if calculated there may be large errors As is evident from Table 6, fresh clover contains large quantities of asparagine and glutamine in the water-soluble part, but the silage con- tains them only in trace amounts From the results obtained in the hydrolysate of the water-soluble part shown in Table 6, it is clear that the greatest constituent of the amide fraction is asparagine But in this experiement, the amides were not divided and shown their sum value I t is well known that these amides converted easily to the corresponding amino acids under acidic conditions Therefore, it is considered that the amides in the herbage may have been converted to the amino acids during ensilage by the organic acids which appeared in the silage And in Table7, the sum of amides and corresponding amino acids are used in calculation and it is shown that only 23% of the maximum value possible was contained in the silage If the amides are omitted from the calculation, it is only 34%.
About 1 5 times as much alanine and 5 times of 7-aminobutyric acid were found in the water-soluble part of the silage as the calculated values From those fact, it is considered that a portion of glutamic acid and aspartic acid were decarboxylated to 7-aminobutyric adid and alanine, respectively, and the existence of these metabolic pathways is well known34). As shown in Table 6, during the ensilage a decrease of arginine was observ- ed, which was accompanied by a n increase in ornithine and the appearance of citrulline I t is known that the ornithine cycle exists in the amino acid metabolism of some micro- organisms such as lactic acid bacteriagd), which is the dominant microorganisms of silage. Further, it is also known that in microorganisms, arginine is converted to citrulline by arginine desimidase, and the citrulline is converted to ornithine by citrulline phos- phor ylase94)
Alpha-aminobutyric acid is a common constituent of some proteins32), but this amino acid was not found in the fresh red clover, while it appeared in the silage as a free form as shown in Table 6 And the appearance of this amino acid in bad silage has been reported by K E M B L E ~ ~ ) in which it appeared after 8 weeks of ensiling ROBERTSON and BAR NETT*^)
also detected this amino acid in fermented marrow stem kale and supposed it was formed perhaps from threonine I t is considered that this amino acid can be produced by mi- croorganisms from a-ketobutyric acid which is present in plants or formed from several amino acids The unknown acid shown in Table 6 agreed well with the peak of p-amino- isobutyric acid in the chromatographic analysis This amino acid was found in the roots of Irzs tzngznata by ASEN ~f a1 2), but there is no report which proves the presence of this acid in any other plant species And the unknown acid was not found in the fresh clover but was found in the silage Some microorganisms catabolyze valine and produce meth- ylmalonate semialdehyde and the latter is transaminated from glutamic acid and converted to p-aminoisobutyric acid34) KIRCHMEIER and K I E R M E I E R ~ ~ ~ found p-alanine and 6-
aminovaleric acid in silages of bad quality, but these acids were not found in silage of good quality
the nitrate was degraded during the ensilage There are many wor kers35,65,10*) reporting on the disappearance of nitrate during ensilage JACOBSON and WISE MAN^^) reported that the nitrate content of silage was largely affected by the pH of silage: I n a silage of pH 5 2 about 20% of the nitrate was degraded, while in a silage of p H 4 7 about 54% of the nitrate disappeared In the present experiment, the pH of the silage ranged from 4.0 to 4.3, and the percent of nitrate which disappeared was in agreement with the report of JACOBSON and WISEMAN.
From Table 5, it is seen that 18 and 28% of the total non-protein nitrogen (4 3 and 15.7% of the total nitrogen) were undetermined in the herbage and in the silage, respec- tively. BRADY~) also reported that about from 20 to 50% of the total non-protein nitro- gens in various herbages and the corresponding silages (from 3 to 16% and about 20% of the total nitrogen, respectively) were unaccounted for as amino-, steam volatile-, amide-, and peptide-nitrogen, and that the unaccounted nitrogen fractions were increased by ensilage O H Y A M A ~ ~ ) reported that the unaccounted for water-soluble nitrogen in various silages ranged from 16 to 37% of the water-soluble nitrogen (1 1 to 25% of the total nitrogen) Many workers reported on the other nitrogenous compounds in herbage or silage FERGUSON and T E R R Y ~ ~ ) determined the content of various nitrogenous bases in grass, clover and lucerne to be : Purine nitrogen 1.3 - 2 2%, pyrimidine nitrogen 0 7 - 3 5%, betaine nitrogen 0 3 - 1.1
%,
and choline nitrogen 0 2 - 0 5%, of the total nitro- gen MACPHERSON~~) and MACPHERSON and V I O L A N I E ~ ~ ) determined histamine, ty- ramine, tr yptamine, putr escine and cadaver ine in grass silages and reported that they were contained only in trace amounts in good silages but in considerable amounts in bad silages.I n the present experiment, no systematic determination of amine was performed but a small amount of ethanolamine was detected in both the herbage and the silage. Eth- anolamine is formed from serine by decarboxylation and is a precursor of choline The silage contained smaller amounts of ethanolamine than did the green herbage This fact shows that a part of it was destroyed during the ensilage.
Summary
By using a mechanical separation method, the tissue of fresh red clover and wet red clover silage was divided into three main parts, i e
,
a fibrous, a concentrated and a water- soluble part I n this method, fresh materials were minced in a blender and the resultant pulp squeezed through a fine hemp cloth to give a green extract, leaving the fibrous part behind Then the green exrtact was divided into two fractions by filtration; the super- natant liquid fraction and the green solid fraction. The former was referred to as the water-soluble part and the latter as the concentrated part1. There was a large difference in nitrogen distribution between herbage and silage I n herbage, 27% of the total nitrogen was distributed in the fibrous part, 44% in the con- centrated part and 29% in the water-soluble part O n the other hand, in the silage, 29% of the nitrogen was distributed in the fibrous part, 15% in the concentrated part and 56% in the water-soluble part
2. Seventy percent of the protein from the concentrated part was degraded to the water-soluble non-protein nitrogen during ensilage. The water-soluble protein and lower molecular peptide were completely degraded during ensilage
3. The amino acid composition of the prokin from the concentrated part was deter- mined and there was no difference found between that in the green herbage and that in the silage I n both samples, aspartic acid, glutamic acid and leucine were highest in quantity followed by alanine with the concentration of methionine lowest
4 The water-soluble part of the fresh clover contained 1.0% ammonia nitrogen, 7 5% amide nitrogen, 2 5% nitrate nitrogen, 4 3% amino acid nitrogen and 4 4:/, peptide nitrogen per total nitrogen, and that of the silage contained 14 4% ammonia nitrogen, trace amide nitrogen, 1.0% nitrate nitrogen, 25% amino acid nitrogen and no peptide nitrogen
5 The amino acid composition of the lower molecular peptide of the water-soluble part of the fresh clover was determined, and found to differ from that of the protein of the concentrated part and the free amino acid composition, and it was unexpectedly observed that more glycine existed in the peptide form than in the free form
6. Free amino acids which were found in large quantities in the water-soluble part of the fresh clover were aspartic acid (0 75% of the total nitrogen), 7 -aminobutyric acid (0 597,), serine (0 57%), proline (0 52%), alanine (0 50%), and glutamic acid (0.45%), and those in the silage were alanine (4 25%), 7-aminobutyric acid (3 02%), leucine
(2 54%), glycine (2 35%) and valine (2 10%) Monoaminodicarboxylic acids and ar- ginine were largely decomposed and a large portion of alanine and 7-aminobutyric acid were formed from other substances during ensilage
Alpha-aminobutyric acid, citrulline and an unknown amino acid (perhaps p-amino- isobutyric acid) were found in the water-soluble part of the silage These were not found in the fresh red clover
Chapter
2
Daily change of free amino acids during ensilage
I n Chapter 1, it was made clear that a large part of the protein in red clover was degraded to free amino acids and a great number of these amino acids were partially de- composed during ensilage These decompositions were greater for monoamino dicarboxy- lic acids and serine More alanine was detected in the silage than expected The free basic amino acids were so small in quantity that the changes in these amino acids during ensilage could not be made clear
The object of the present experiment is to make clear the processes and the substances the amino acids are changed to during incubation For this purpose, universally labeled 14C amino acids were added to red clover juice and their daily changes during incubation were followed
The chemical composition of silage is largely affected by microbiological, chemical, and physical factors And it is not a n easy matter to differentiate these three factors be-
cause they are extremely complex But to compare the data obtained in different silages, it is necessary to make the silages as nearly as possible the same in quality. For this pur- pose, the plants were minced, the fibrous fraction filtered off, and the resultant green liquid was incubated with glucose under anaerobic conditions. But in this case, since the microorganisms make more rapid contact with their nutrients than in silage, there may occur some differences in the bacteria phase between the two treatments. Moreover, as a result of macerating, the plant cell walls are broken and the activity and the reaction period of the plant enzymes may differ from those in silage. Therefore, a silage was also made from the intact red clover and the differences between "the juice" and "the silage" were discussed Because in the case of the silage, if the 14C amino acid was dropped on the surface of the plant, the degradation process of the amino acid may largely differ from that in the plant cell, and if the 14C amino acid is injected into the plant cell, a part of it would be metabolized before ensiling.
Materials and Methods
1. Incubated red clover juice
I n this work, two experiments were carried out The first(a) was to compare the fermentation pkocesses of incubated juice with silage and the latter(b) was to determine the daily change of the added 14C amino acids
(a) Two kilograms of red clover harvested on June 13th, 1967 in the full bloom stage from the Kagawa University farm was ground in a mixer (Hitachi JB-10) with 4 l of water. The fibrous fraction was removed by filtration and the dark greenish liquid fraction was poured into a 5 liter bottle containing 40g of glucose. The bottle was stoppered with a rubber stopper to which a glass tube connected with rubber hose was attached. Through this the air in the bottle was sucked out with a water aspirator and then the rubber hose was closed off with a pinch cock The bottle was shaken well and stored in a room regu- lated to 20°C. Sampling was made on juice incubated 0, 1, 2, 3, 4, 5, 6, 7, 10, 14, 21, and 74 days The bottle was shaken well before each sampling and air in the bottle was aspirated again as described above immediately after sampling. The sample taken was centlifuged for 10 minutes at 4,000r p.m. and separated into a supernatant liquid and a greenish precipitate The former was referred to as the water-soluble part and the latter as the concentrated part. The water-soluble part was kept in a freezer until ready for analysis and the concentrated part was oven dried a t 70°C.
(b) Red clover was harvested on Aug 5th, Oct 15th, Oct 25th, 1968, Jan loth, Jan 30th, Apr 24th, May, 7th, Jul 18th, 1969 and Apr 22nd, 1970 Soon after harvest- ing, the fresh red clover was separated by filtration into liquid and fibrous fractions following maceration in a juicer (Hitachi JB - 10) The liquid fraction was mixed with glucose in the ratio of 100 to 2 and the 5 ml of the mixed clover juice was transferred into test tubes(l5
x
160 mm) Five microcuries of a 14C amino acid was added into each test tube containing clover juice To the juices separated from red clover har- vested on Apr 5th and Oct 15th, 1968, 1%-U-glutamic acid was added 14C-U- aspartic acid or -arginine, 14C-U-alanine, -lysine, or -threonine, 14C-U-tyrosine, -valine,-phenylalanine or -histidine, 14C-U-glycine, -1eucine or -isoleucine, and 14C-methionine, -proline, -serine or -glycine were added to the juices separated from the red clover har- vested on Oct 25th, 1968, Jan 30th, Apr 24th, May, 7th and Jul. 18th, respectively 14C-3-tryptophan was added to the juice made on
Apr 22nd, 1970 The test tubes were stoppered with rubber stoppers attached to small glass cups (Fig. 2) in which a 1 m l of aliquot of "Hyamine 10X"
Rubber tube was kept to trap the carbon dioxidel05) After
aspiration through the rubber tube attached on the
top of glass tube, the contents of the test tubes was -Glass tube incubated a t 20°C. After incubation of 4 and 8
hours, 1, 2, 7, 14 and 35 days the fermentation of --Rubber stopper clover juice was terminated by heating for a few
seconds in boiling water Then the tubes containing the incubated juice were stored in a freezer until ready for analysis The 14C-U-glutamic acid treat- ment was preformed on the 3rd day of incubation using the juice separated from the red clover harvest-
ed on Jan loth, 1969, and the termination was done
-
Test tube onl, 2, 4,
7 and 32 days after glutamic acid treatment
2 Test tube silage
A part of the fresh red clover which was harvested on Aug. 5th, 1968 and used for separating juice for 14C-U-glutamic acid treatment was chopped into about 0 5 cm long and mixed well with 2% of glucose Five grams of the chopped clover was packed tightly into each of 5 test tubes (15x 160 mm). Ten microliters (5 pc) of ethanol solution of 14C-U-glutamic acid was injected into the central
Figer 2. ApPratus for
part of the packed clover of each of the 5 test tubes 14C containing materials. The test tubes were stoppered with rubber stoppers
attached to small glass cups in which 1 ml of the Hyamine was kept, aspirated and then stored at 20°C Each test tube was moved into the freezer after 0, 2, 7, 14 and 35 days of ensiling, respectively, for storage until ready for analysis.
3 Determination of 14C in carbon dioxide
All of the Hyamine in the glass cup was dissolved into 10 ml of toluene scintillant which contained 4 g of PPO and 0 1 g of POPOP in 1 1 toluene, and the radioactivity of 14C in carbon dioxide was determined with the use of a Horiba Type LS-500 liquid scintillation spectrometer
4. Fractionation of 14C-containing substances in incubated juice
The incubated juice was centrifuged for 10 minutes a t 4,000 r p m
,
and the super- natant was transferred into a flask. The residue was washed with 5 ml of' distilled waterfive times, and the washings were mixed with the supernatant The juice was divided into the three fractions, amino acids, organic acids and neutral fractions by the method of CANVIN and B E E V E R S ~ ~ ) as described below. The centrifuged juice was first passed through a 6
x 1
cm column of Dowex 50 x 8 (H), and the effluent was then passed through a 6x
1 cm column of Dowex 1 x 10 (formate) The effluent from the latter column is the amino free neutral fraction The amino acids were eluted with 50 ml of N-NaOH from the former column, and the organic acids were eluted with 40 ml of 4N-formic acid from the latter column The organic acid fraction was neutralized with 28% aqueous ammonia and concentrated to about 10 ml in a water bath a t 60°C under a vacuum The concentrated organic acid fraction was transferred into a 50 ml volumetric flask and made up to volume with methanol The amino acid and neutral fractions were evaporated to nearly dryness in the water bath at 60°C under a vacuum. The concentrates were redis- solved in 2 ml of water and then transferred into 10 ml volumetric flasks and made up to volume with methanol5 Treatment of 14C-containing silage
The silages containing 14C were treated as follows The contents of the test tube were transferred into a 50 ml centrifuge tube After addition of 10 ml of water, the cen- trifuge tube was attached to a homogenizer (Nihon Seiki Universal homogenizer) and the contents were macerated The macerate was centrifuged for 10 minutes at 4,000 r.p.m and the supernatant was transferred into a flask This treatment was repeated four times 'The supernatants were mixed and then treated in the same manner as the incubated juice
6 Determination of the radioactivities of the amino acid, organic acid and amino free neutral fractions
One hundred microliters of each solution, 0 9 ml of Hyamine and 10 ml of the toluene scintillant were mixed in a liquid scintillation vial and the radioactivity was determined by using a liquid scintillation spectrometer
7 Paper chromatography of 14C-amino acids
The amino acid fraction was further developed chromatographically on Toyo No 50 filter paper (2 ~ 4 0 cm) The solvent systems used in this study were phenol-water (85 : 15, v/v) and collidine-lutidine (1 : 3, v/v) saturated with water In the 14C-trypto- phan experiment, ethyl methyl ketone-acetone-formic acid-water (40 : 2 : 1 : 6, v/v) and ethyl methyl ketone-diethylamine-water (46 : 0 1 : 3 9, v/v) were also used The radio- activity of each amino acid separated on the paper was determined by using a n Aloka radio- paper chromatograph JPC-101
8 Determination of the pH value
The pH values of the water-soluble parts of 14C-free materials were determined elec- trically by using an Hitachi-Horiba pH meter H-5 While the pH values of the "C-con- taining juices were determined with the use of Toyo p H test papers
9 Determination of lactic and volatile acids
Lactic acid contents of the fermented materials were determined colorimetrically on the water-soluble parts according to the method of BARNETT 3) The lactic acid in
which gives a purple-red colour with @-hydroxydiphenyl in the presence of copper ions The developed colour was determined at 570 m p using a Shimazu Bausch and Lomb Spec- tronic 20 calorimeter The determination of the volatile acids was made as follows Steam distillates of 50 ml of the water-soluble parts were titrated with 0 05 N-NaOH to know the total VFA contents and then the titrated solutions were evaporated to dryness by a rotary evaporator at 84OC The dried samples were dissolved in a few drops of formic acid followed by a few milliliters of ethylether Three to five microliters of the dissolved samples were injected into a Hitachi gaschromatograph Model 53 attached with a hydro- gen flame ionization detector The column used was 2 m long and packed with polyethy- lenglycol-adipate-celite The carrier gas used was N2
Results
1 The experiment on the fermentation process of incubated red clover juice
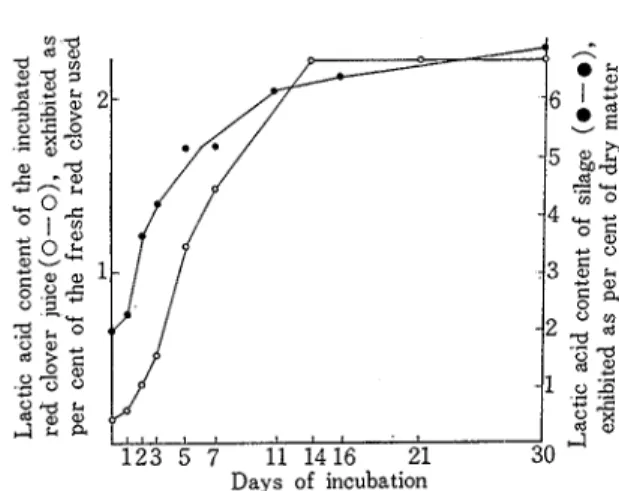
I n most of this work, red clover juice was used instead of the red clover for studying the daily change of free amino acids during fermentation Therefore, a t first, it was nec- essary to compare the fermentation processes of incubated juice with those of silage The daily changes of the organic acid content and the pH values of the incubated juice were determined as shown in Table 8 and the results were compaied with those of silage quoted from a bookloo) From Table 8 and Figure 3, it is clear that the incubated juice had a strong resemblance to the silage in both the lactic acid formation and the pH fall
The protein degradation and ammonia formation during incubation of the juice were also determined and the results are shown in Table 9 The non-protein nitrogen and the volatile basic nitrogen formation were most vigorous within the first period of incuba-
Table 8 Daily change of the pH value and the organic acid content of the incubated red clover juice compared with those of silage
Incubated juice
after Lactic Acetic
ensiling acid acid
0 1 2 3 5 7 10 11 14 16 2 1 30 74 86 Silage* Lactic PH acid 6 15 0 06 0 02 5 86 0 17 0 07 4 62 0 34 0 15 4 50 0 49 0 21 4 10 1 12 0 25 3 98 1 4 5 0 29 3 87 1 72 0 27 -
-
- 3 80 2 22 0 29 - - - 3 80 2 19 0 31 --
- 3.80 2 21 0 46 - - -I
-
0 /o**
0 /o**
I
0 /o***
*
Quoted from a book edited by Suto '"O'**
Calculated on the basis of fresh red clover***
Calculated on the basis of dry matterFigure 3 Comparison in lactic acid formation during incubation between the incubated red clover juice and a practical silage.
tion During 74 days, about 104 mg of the protein nitrogen degraded to water-soluble nitrogen in which about 12 mg (1 1 5%) of volatile basic nitrogen was contained These values are far lower than those obtained on the silage shown in Tables 3 and 5.
2 The experiments on the daily changes of added 14C-amino acids during incubation of red clover or its juice
14C-U-glutamic acid: Three experiments were carried out on the fate of 14C-U- glutamic acid during incubation The first is on the fate of 1%-U-glutamic acid added to fresh juice a t the start of incubation, the second is on the fate of it when added to the juice on the 3rd day of incubation, and the third is on the fate of it when added to the intact red clover at the time of ensiling.
The fate of 14C-U-glutamic acid added to the fresh juice and the daily change of the pH value of the juice are shown in Table 10 The results show that the major part of the free glutamic acid was decarboxylated to 7-aminobutyric acid and only a small part was deaminated to organic acids during the early stage of incubation Gamma-aminobutyric acid was not further degraded during the course of the experiment I t was also observed
Table 9 Daily change of the water-soluble non-protein nitrogen and volatile basic nitrogen content of the in- cubated red clover juice, exhibited as mg in 100 g of fresh red clover material
Days of
I
Water-soluble incubation nitrogenVolatile basic nitrogen
Table 10 Incorporation of 14C from "C-U-glutamic acid into various fractions of the incubated red clover juice which was added with "C-U-glutamic acid at the start of incubation (exhibited as per cent recovery of 14CC: in the various fractions)
I
Incubation periodCompounds
Carbon dioxide Amino acid fraction
Glutamic acid 7 -aminobutyric acid Organic acid fraction N free neutral fraction
Hours Days
1
81
2 7 3 5Recovery of "C in liquid
and gaseous fractions
/
8 0 3 85 9 93 3 99 1 96 1 99 1pH of the juice
1
5,8 .5 8 5 4 4.2 3.6 3.6that about 20% of th., total 14C disappeared from the liquid and gaseous fractions for the first 4 hours of incubation and after then gradually reappeared in those fractions All of the added 14C-glutamic acid was changed to other substances within 2 days.
The fate of 1%-U-glutamic acid which was added to the juice on the 3rd day of incuba- tion and the daily change of the pH value are shown in Table 11. As a result, added 14C- glutamic acid was not degraded to 7-aminobutyric acid till the 7th day after glutamic acid treatment, but 16 6 and 17.8% of the added radioactivity were shifted to ?-amino- butyric acid and organic acids, respectively, after the 32nd day after 14C treatment.
Table 11 Incorporation of "C from "C-U-glutamic acid into various fractions of the incubated red clover juice which was added to "C glutamic acid on the 3rd day of incubation, exhibited as per cent recovery of 14C in the various fractions
Compound
1
Days after 14C glutamic acid treatment
Yo Yo Yo % Yo
Carbon dioxide 2 4 3 1 3 2 4 0
Amino acid fraction Glutamic acid
-aminobutyr ic acid Organic acid fraction N free neutral fraction Recovery of "C in liquid
and gaseous fractions
1
9 6 5 100 0 98 1 96 5 97 3 p H of the juice1
4 2 3 8 3 6 3 6 3 6
The fate of 14C-U-glutamic acid which was added to the intact red clover was studied and the results are shown in Table 12 together with the daily change of the pH values I n this experiment, the organic acids and the neutral fraction were not divided and are
Table 12 Incorporation of "C from "C-U-glutamic acid into various fractions of the silage (exhibited as per cent recovery of "C in the various fractions)
-
Days after ensiling Compounds
7 14 3 5
Carbon dioxide 12 4 15 7 15 7 15 7
Amino acid fraction*
1
6 6 9 71 5 71 2 69 9 N free liquid fraction*" 8 6 5 8 6 1 6 8 Recovery of 14C in liquidand gaseous fractions
1
8 7 8 93 0 93 0 92 4 p H of silage*
Most of this fraction consisted of 7-aminobutyric acid**
This fraction consisted of organic acids and an amino-free neutral fraction.exhibited as the nitrogen free liquid fraction Most of the 14C-glutamic acid was degraded to r-aminobutyric acid by decarboxylation within 2 days of ensiling and only a trace a- mount of 14C-glutamic acid was found throughout the experiment
14C-U-aspartic acid: The fate of 14C-U-aspartic acid in the red clover juice during incubation is tabulated in Table 13 During the first 4 hours of incubation, more than
10% of the added 14C was incorporated into organic acids, and the incorporation rate in- creased till 24 hours after treatment This result indicates that a part of the free aspartic acid is deaminated during the early stage of incubation Within the first 2 days of incuba- tion, a part of the organic acids was incorporated into the neutral fraction and 14% of the added 14C was transferred into carbon dioxide Decarboxylation of aspartic acid to a-alanine was observed during the course of the experiment but most of it was done within
2 weeks
Table 13 Incorporation of "C from 14C-U-aspartic acid into various fractions of the incubated red clover juice (exhibited as per cent recovery of 14C in the various fractions)
- --
1
Incubation periodCompound Hours Days
8 2 7 14 35
Carbon dioxide Amino acid fraction
Aspar tic acid a-alanine Organic acid farction N free neutarl fraction Recovery of "C in liquid
and gaseous fractions 1 9 5 8 9 2 4 8 7 3 8 2 8 8 4 9 8 8 8 8 9 0 p H of the juice
1
5 8 5 8 5 4 4 1 3 8 3 8 3 814C-U-glycine: Two experiments were made on the fate of 14C-U-glycine during the incubation of the red clover juice, and the results are shown in Table 14 together with the pH values of the juices The data in experiment 1 shows the fate of 14C-glycine under bad fermentation, while the result in experiment 2 was obtained under better conditions, as is evident from the pH values A slight decarboxylation was observed daily in both experiments The decarboxylation rate was higher in experiment 2 than in experiment 1 till the 7th day, but after then no obvious difference between them was observed The most distinct difference between the two experiments was the recovery of 14C in the liquid and gaseous fractions More than 90% of the 14C in the liquid and gaseous fractions was found in glycine during the whole incubation period in both experiments as shown in pa- rentheses in Table 14
Table 14 Incorporation of 14C from 14C-U-glycine into various fractions of the incubated red clover juice (exhibited as per cent recovery of "C in the various fractions)
- --
Incubation period
Recovery of "C in liquid
and gaseous fractions
1
96 3 98 2 91 2 94 4 93 3 93 4Compounds Hours Days
8 7 14 3 5
Experiment 1 Carbon dioxide Amino acid fraction*
hT free liquid fraction
Recovery of l 4 C in liquid
and gaseous fractions
1
9 6 7 9 3 8 9 2 3 8 7 0 716 6 8 3 714% % % % % % % 0 0 4 0 0 5 0 0 9 0 18 0 74 1 1 1 0 9 3 4 9 5 5 88 6 90 0 88 3 88 8 (97 0) (97 2) (97 1) (96 1) (94 6) (95 2) 2 9 2 7 2 5 3 7 3 9 3 6 p H of the juice Experiment 2 Carbon dioxide Amino acid fraction* N free liquid fraction
p H of the juice
I
5 5 5 5 5 5 5 0 4 2 3 8 3 7 5 6 5 6 5 6 5 6 5 0 4 1 0 1 1 0 1 1 0 2 2 0 9 0 1 1 1 1 1 4 93 7 91 2 89 7 83 0 73 5 64 0 66 0 (96 9) (97 2) (97 1) (95 4) (94 8) (93 7) (92 5) 2 9 2 5 2 4 3 1 3 0 3 2 4 0*
Glycine was the only 14C constituent of this fraction The data in parentheses show amino acid- 14C per recovered "C in liquid and gaseous fractions1%-U-alanine: The fate of 14C-U-alanine during incubation of the juice was studied and the results are shown in Table 15 Within 4 hours of incubation, more than 11
%
of the 14C of 14C-alanine was found in the organic acids and neutral fractions In the pe- riods from 4 to 8 hours, the 14C in these fractions decreased to about 4 5%, and the total 14C recovered in gaseous and liquid fractions also decreased to 80% The rapid fall of the pH value and the rapid increase of 14C labeled carbon dioxide and 14C in the neutral fraction were observed in the 1 to 2 day periods of incubation The 14C in the organic acid fraction increased again from 2 to 7 day periods of incubation No other 14C aminoTable 15 Incorporation of 14C from 14C-U-alanine into various fractions of the incubated red clover juice (exhibited as per cent recovery of "C in the various fractions)
-- -- - - -
I
Incubation periodRecovery of 14C in liquid
and gaseous fractions
1
9 0 0 80 3 82 2 79 9 82 7 80 4 Compounds - Hours Days 8!
2 7 3 5 I p H of the juice1
5 8 5 8 5 6 4 8 4 1 3 8 Carbon dioxideAmino acid fraction* Organic acid fraction Neutral fraction
--
*
Alanine was the only 14C constituent of this fraction The values in parentheses indicate amino acid- 14C per recovered 14C in the liquid and gaseous fractions% % % % % % 0 25 0 33 0 54 3 0 4 8 5 4 78 6 75 4 75 3 66 5 63 0 59 4 (87 3) (93 3) (91 6) (83 2) (76 2) (73 9) 10 7 4 0 5 7 4 9 9 2 9 1 0 44 0 57 0 67 5 5 5 7 6 5
acid was found in the amino acid fraction than alanine throughout the experiment Dur- ing the course of incubation, 40% of the added alanine disappeared from the liquid and gaseous fractions
1%-U-serine: The 14C-U-serine added to the juice was largely degraded during the course of incubation as shown in Table 16. Nearly 9% of the 14C-serine was deami- nated and shifted to the nitrogen free liquid fraction within 4 hours of incubation. The largest degradation of 14C-serine was observed in the 1 to 2 day periods of incubation when the pH of the juice was falling Within 2 days of incubation, 85% of the 14C of 14C-serine disappeared from the amino acid fraction, 18 and 34% of it was recovered in the carbon dioxide and the nitrogen free liquid fractions, respectively, and only about 68% of it was recovered in the liquid and gaseous fractions After then little change was observed and
Table 16 Incorporation of "C from "C-U-serine into various fractions of the incubated
Carbon dioxide Amino acid fraction*
red clover juice (exhibited as per cent recovery of 14C in the various fractions)
N free liquid fraction Compounds
Recovery of 14C in liquid
and gaseous fractions
1
97 2 95 3 76 0 67 7 64 7 65 2 73 8 Incubation periodHours
1
Daysp H of the juice