スピヲ科(多毛類)の一新種Nerinides yamaguchi n.sp. の記載とその発生について (英文)
12
0
0
全文
(2) Vol. 10. No. 1 Journal of Hokkaido Gakugei University July, 1959. A Description of a New Species of the Spionidae (Polychaeta), Nerinides yamaguchii n. sp., with Notes on its Development. Minoru IMA.TLMA Shirikishinai M-arine Biological Laboratory Hokkaido Gakugei University. 4-r^ /%: ^k°5rl:4 (^r6?ti) ®—tr@l Nerinides yamagtidni n. sp. ®i5l?^^®^^?c:-o^T (With 3 Plates and 1 Table). In April, 1955 and 1958, the writer collected many specimens of a spioniform species referable to the genus Nerinidss which has not yet been recorded from the adjacent waters. of Japan, except N. papillosus described by Qkuda (1937) from Korea. From observation on the specimens, they do not seem to be identical with Okuda's species, and the former has been reported as Nerinides sp. in the writer's preliminary note (1958). Afterward, it appeared to the writer from further survey of literature on the animal group that the writer's species was new to science. In the present paper, will be found the description of the new species and notes on its larval development to metamorphosis, observed in the laboratory. Before proceeding further the writer wishes to tender his cordial thanks to Prof. Dr.. Hideji Yamaguchi of the Hakodate Branch, Hokkaido Gakugei University for his kind guidance during the course of the investigation and preparation of the paper. Nerinides yamaguchii n. sp. (PI. I, Fig. 1-13 ; Table 1) Body measures 50 mm long by 4.5 mm in specimens with 87-102 setigerous segments. The prostomium is spindle-shaped, acutely pointed at the terminal portion and separated from the buccal segment by deep grooves in both the lateral sides. There is a short erect median occipital tentacle on the prostomial ridge posterior to the eyes and between tentacles (Pl. I, Fig. D. The tentacles are very long, taper regularly to their apices and when elon gated backward extend as far as the tenth chaetiger. The dorsal side of basal portion of the tentacles (Pl. I, Fig. 3) is clothed in a granulated membranaceous sheath. These coatings bear about 8-14 short triangular cirriform papillae on the free margin. The ventral bases of tentacles are not completely covered by the membranaceous sheath (Pl. I, Fig. 4). There are four small black eyes betweeen the bases of the tentacles. The first setigerous segment (Pl. I, Fig. 5). has a distinct neuropodium and notopodium, the setigerous lobe of the latter being slightly prominent than that of the former. Each bears two kinds of capillary setae, short and long, both being quite similar in form. —155.
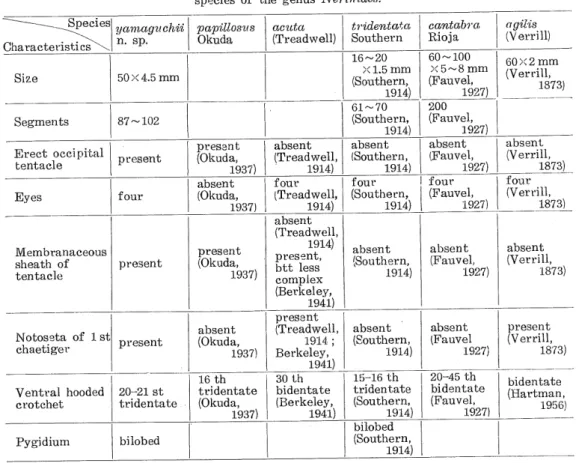
(3) Minoru Imajima. The branchiae begin at the 2 nd chaetiger (Pl. I, Fig. 6); out of them long ones are present on the 10 th-20 th segments. They are flattened, lobate and densely ciliated on the lower margin, extending to the dorsal surface. Unlike Nerinides acuta (Treadwell, 1914), there are rudimentary presetal lobes on the parapodium anterior to the dorsal setae bundles. The ventral lobes which are situated posterior to the ventral setae bundles, are rounded in segments anterior to the seventh, and then they (Pl. I, Figs. 8-10) become flattened and expanded transversely, the lobes being gradually reduced toward the posterior in the posterior region (Pl. I, Fig. 11). The dorsal setae (Pl. I, Fig. 13), which has a small wing along its middle half, and one, similar to those of the first setigerous segment, are longer and more numerous than the ventral ones. The ventral hooded crotchets (Pl. I, Fig. 12) first appear on the 20 th-21 st chaetiger, showing a row with 4-5 setae. They are tridentate, with two sharp spines on a curved shaft and covered by a hood. The ventral lobe has a number of slender capillary setae and ventral crotchets. There are 20 crotchets in the 30 th chaetiger and 30 crotchets in the 70 th chaetiger. From about 70 th segment, crotchets decrease gradually in number toward the posterior. Pygidium bears a ventral flattened expansion, which is more or less bilobed (Pl. I, Fig. 2). This feature agrees with N. tride.nt.ata Southern fl914). Remarks : As is shown in Table 1, the characteristics of the present species are similar. to N. acuta (Treadwell) reported by Berkeley (1941), in the presence of notosetae of the first chaetiger, in possession of eyes and triangular thickening at the base of tentacles; but the Table 1. Comparison with several characteristics in species of the genus Nerimdes.. Specie Characteristics^ Size. yamagzichi' n. sp.. papilloszts Okuda. aczita. (Treadwell). tridentata Southern 16~20 X 1.5 mm. 50X4.5 mm. Erect occipital tentacle. Eyes. 87-102. ><5~8mm (Fauvel,. 61~70. 200. present. four. (Southern, present. (Okuda,. 1937). absent. (Okuda,. 1937). 60-100. (Southern,. 1914). Segments. cantabra. Rio j a. 1914). agil'is. (Verrill). 60X2 mm (Verrill,. 1873). 1927). (Fauvel,. 1927). (Treadwell,. absent (Southern,. absent (Fauvel,. absent. four. four. four. four. absent. 1914). (Treadwell,. 1914). absent. 1914). (Southern,. 1914). 1927). (Fauvel,. 1927). (Verrill,. 1873). (Verrill,. 1873). (Treadwell, Membranaceous. sheath of tentacle. present. present. (Okuda,. 1937). 1914). present,. btt less. complex. absent (Southern,. absent. (Fauvel,. (Verrill,. absent. absent. present. 1914). 1927). absent. 1873). (Berkeley,. 1941). Notoseta of 1 si. chaetiger. Ventral hooded. present. 20-21 st. crotchet. tridentate. Pygidium. bilobed. absent. (Okuda,. 1937). 16th. tridentate (Okuda,. 1937). present. (Treadwell, 1914 ;. Berkeley,. 1941). 30 th bidentate (Berkeley,. 1941). (Southern,. 1914). 15-16 th. tridentate (Southern,. 1914). bilobed. (Southern,. 1914). - 156 -. (Fauvel. 1927). 20-45 th bidentate (Fauvel,. 1927). (Verrill,. 1873). bidentate. (Havtman,. 1956).
(4) New Species of the Spionidae (Polychaeta), Nerinides yamagzich.ii n. sp.. present species differs from the latter in the following respects : 1) A erect median occipital tentade lying between the tentacles; 2) The triangular membranaceous sheath is more complex ; 3) Ventral hooded crotchets are tridentate. On the other hand, the present species seems to be more or less related to N. capillosus Okuda (1937) by agreement in the presence of the membranaceous sheath of tentacles and a erect median occipital tentacle, and the. feature of the ventral hooded crotchets. But both species are easily distingushable by the absence of eyes in the latter, and by the presence of the dorsal capillary setae of the first setigerous segment in the former. Though similar tridentate ventral crotchets are found in N. iridentata, this species does not accord with the present species in many other characteristics. The species was found embedded in a sandy bottom of the littoral zone. The name of this new species has been chosen as an expression of my gratitude to Prof. H. Yamaguchi. Notes on the development (Pl. II, Fig. 1-8; Pl. Ill, Fig. 1-6) The material for this investigation was obtained from natural spawn-masses attached to the tube of the parent, and was brought to the laboratory and transferred into culture dishes filled with a culture medium which was natural sea-water filtered through fine bolting silk. Though the dishes were usually kept at natural room temperature, they were cooled by running tap water when the temperature of the culture medium become conspicuously higher than that of natural habitats, Breeding habit; The adult worms were found in sand between low and high tide marks, living in soft sandy tubes. In spring, from April to May, spawn-masses (Pl. II, Fig. 1) which are more or less clubshaped and jelly-like, are found attached by their narrow stalk to the mother tube and can move to and fro in the waves. Their jelly-masses contain a large number of minute eggs which are aggregated within swollen portion of the jelly-mass, forming a long elipsoidal egg-mass, The spawn-mass measures about 23 mm in length and 5 mm in breadth at its gratest width in the swollen portion. Though such features of the jelly-mass. resemble those of Spio filicornis observed by Okuda (1947), but they differ in size. Development within spawn-mass. The material obtained from a newly formed spawn-mass have been developed to the stage of gastrula (?) (Pl, II, Fig. 2). Then, the early development have not been observed. The embryos are ellipsoidal and pale green in reflected light, measuring about 0.24 mm long and 0.18 mm wide, showing some variation in size and shape. About two days after, it (Pl. II, Fig. 3) begins to rotate slowly. It is about 0.27 mm long and 0.21 mm broad at the prototroch, with a darker vesicle near the centre. The prototroch is seen as a small area with short cilia. The telotroch is also seen as a similar area. The base of the prototroch and the telotroch are slightly transparent. In the lateral side of the upper part of the prototroch lies a black chromatophore with indistinct contour, which is a rudiment of a eye.. About three days later, the prototrochal cilia lengthen, forming a ciliated bundle extending toward both the dorsal and ventral sides (Pl. II, Fig. 4). On the other hand, the telotroch is present as six groups of cilia. The first bristle bundles have appeared. The embryo is well rounded dorsally, less so ventrally, The paired black chromatophores on the dorsal surface have translocated toward the median line, growing as paired dorsal eyes. About four days later the embryo (Pl. II, Fig. 5) developes a trochophore-like form. It is about 0.35 mm long — 157 —.
(5) Miuoru Imajima and 0.23 mm broad at the prototroch. A few sensory cilia are found at the anterior end of the body. In addition to the dorsal eyes a pair of ventral eyes has developed on the ventral surface, being widely separated. The telotrochal six groups of cilia have developed further. The part bearing the prototroch becomes rounded in elevation, forming a lateral cheek. The first bristle bundles are well developed and located in the upper part of the segment. Three. segments are distinguishable in the embryo at this stage. The pygidium has developed as an ellipsoidal projection with a notch. About seven days after, it is about 0.48 mm long and 0.29 mm broad at the prototroch (Pl. II, Fig. 6). The head is oval in front with a small round elevation at the extremity. The gut which has developed from the dark central mass in the early stages, is marked by a darker area, which extends from the first setiger to the posterior end. The two pairs of eyes are fully developed, one pair being dorsal and the other ventral. On the midway between the apical end and the prototroch a pair of finely. ciliated cubic swellings appears on the lateral walls. During the further development similar structures are also found just behind bristle bundles in successive segments. These swellings seem to be rudiments of ventral parapodia. The prototroch increases m length, forming a continuous ciliary band on the lateral cheeks which are due to become the lateral tentacles.. The pygidium is ovate, with a slight notch and bears a pair of telotrochal ciliary bands which originate from the cilia-groups already mentioned. A pair of small cilia-groups is observable. near the notch on the surface of the pygidium. The first provisional bristle bundles are longest and reach backwards as far as the end of the third chaetiger. The bristles on the second and succeeding setigers are always slightly shorter than thoes of the first setiger. Three days after this, the embryos have been liberated from the jellymass into the medium as larvae. Development in pelagic life. About five days after liberation from jelly-mass, the larvae swim very actively in the culture dishes and have grown to about 0.77 mm long and 0.25 mm wide. A dorsal view of the larva at this period is shown in Pl. II, Fig. 7. The cheeks are elongated forming tentacles; and the prototroch located on the tentacle is observable on the ventral side as prominent cilia-bundles extended toward the mouth. On the ventral surface of the head a vestibule is recognisable, bordered by fine cilia. Between the. mouth and the middle part of the first setiger a short row of fine cilia is present along the mid-ventral line. A short swelling appears ventrally at the anterior end of the head. The gastrotroch is present in. pairs on the third and fourth setiger and that of the latter more separated than that of the former. The provisional bristles and telotroch become larger. About ten days after liberation, the larvae reach the stage showed on Pl. HI, Fig. 1. The larva is now 1.4 mm long with 17 setigerous segments. One pair of tentacles become more elongated. A short median tentacle, originating from the short swellings of the earlier period, appear on the ventral side near the anterior end of the head. The extremity of the tentacle is observable from the dorsal side. The body becomes slender, and dorsal and ventral parapodia are well developed. The ventral parapodia of the tenth and succeeding segments are provided with hooded crotchets which are not present in anterior segments. The ventral hooded crotchets (Pl. Ill, Fig. 5) are tridentate, with two sharp spines, resembling that of the adult. Nototrochs are developed on the dorsal surface of all segments from the second to the eleventh setigers. The first provisional bristle bundles are borne on the dorsal surface at the level of the developing lateral tentacles. 158—.
(6) New Species of the Spionidae (Polychaeta), Nerinides yamaguchii n. sp. Setting-down and metamorphosis. About thirteen days after liberation, some larvae began to settle down to the bottom of the glass dishes and all larvae finished settling down in few days. The larvae settled down,. hid themselves in the clear sand which lay at the bottom of the culture dishes, exposing tentacles which stretched obliquely forward and outward, moving to and fro in the medium. The larva drawn in Pl. Ill, Fig. 2 is one recently settled down. The lateral tentacles and the short median tentacle have lengthened further, but the prototroch and the provisional setae and the cubic-shaped ciliated swellings shown in the previous stage are still present. The telotroch becomes powerful and forms a ring of ciliated bundles. Eighteen days after,. a larva (Pl. Ill, Fig. 4) is 2.8 mm long with 25 ehaetiger. The relative length of the tentacles is great in comparison with the adult. The long provisional bristles which were present in the preceeding stages have been lost, but the dorsal setae on the first notopodium and the erect occipital tentacle have still not appeared. The body-wall was very transparent and almost colourless, except for the darker par fc of the gut which is visible from the tenth segment to the seventeenth segment. I have unfortunately been unable to culture these larvae for observation of later development. Literature cited Berkeley, E. and G.: 1941. On a collection of polychaeta from Southern California. Bull. So. Calif. Acad. Sci. Vol. XL. Part 1. Fauvel, P. : 1927. Faune de France. Vol. 16. Polychetes Sedentaires. Hartman, 0.: 1956. Bull. Amer. Mus. Nat. Hist. Vol. 109, p. 291. Imajima, M.: 1958. Note on a Spioniform polychaeta, Nerinzdes sp. (preliminary) (in Japanese). Jour. Hokkaido Gakugei Univ., Sec. B. Vol. 9, No. 1. Okuda, 8. ; 1937. Spioniform polychaetes from Japan. Jour. Fac. Sci. Hokkaido Imp. Univ., Ser. Vol. 6, No. 3.. Okuda, S.: 1947. Studies on the development of Annelida polychaeta I. Jour. Fac. Sci. Hokkaido Imp. Univ., ger. VI, Vol. IX, No. 2. Southern, R. : 1914. Archiannelida and Polychaeta. Clare Ireland Survey Proc. R. Irish. Acad., Vol. 31.. Treadwell, A.: 191^. Polychaetous annelids of the Pacific Coast in the collections of the Zoologicl Museum of the University of California. Univ. Calif. Publ. Zool. Vol. 13, No. 8. Verrill, A. and Smith, S. : 1873. Inverfcebrate Animals of Vineyard Sound. Annelids. Report U. S.. Fish Commission for 1873,. 159—.
(7) Mmoru Imajima. Explanation of Plate I Nerinides yamagzichi.i n. sp. Fig. 1. Dorsal view of anterior body, tentacles being removed. x 15. Fig. 2. Dorsal view of posterior end. X 20. Fig'. 3. Base of tentaele, dorsal view. X 40. Fig. 4. Ventral view of base of tentacle. X 40. Fig. 5. First left parapodium. X 20. Fig. 6. Second left parapodium. ><20. Fig. 7. Tenth left parapodium. X 20.. Fig. 8. Thirtieth right parapodium. X 20. Fig. 9. Sixtieth left parapodium. X 20. Fig. 10. Eightieth left parapodium. X 20. Fig. 11. Right parapodium from posterior part of the body. ><30. Fig. 12. Grotchet of Seventieth neuropod. X440. Fig. 13. Dorsal seta. x 120.. —160 -.
(8) New Species of the Spionidae (Polychaeta), Nerinides yamagztchii n. sp.. 161.
(9) Minoru Imajima. Explanation of Plate II Development of Nerinides yamagnchii n. sp. Fig. 1. Spawn-mass. X 1.4.. Fig. 2. Fertilized egg. X 140. Actual diameter approx. 0.24mm. Fig. 3. Early embryo about two days after. Dorsal view. x 140. Actual length approx. 0.27mm. Fig. 4. Embryo about three days after. Dorsal view. X 140. Fig. 5. Trochophore-like embryo about four days after. Dorsal view. X 140. Actual length approx. 0.35 mm. Fig'. 6. Later embryo about seven days afte>\ Dorsal view. X 140' Actual length approx. 0.48mm. Fig. 7. Larva about five days after liberation from jelly-mass. Dorsal view. x 140. Actual length approx. 0.77mm. Fig. 8. Ventral view of a larva at the same stage as shown in Fig. 7. ><140.. - 162.
(10) Ne» S^ie, „. „ ^^ ^^ ^^ ^^ ^ ^. —163-.
(11) M:inoru Imajima. Explanation of Plate III Development of Nerinides yamaguohii n. sp. Fig. 1. Larva about ten days after liberation. Dorsal view. x 95. Actual length approx. 1.4mm. Fig. 2. Larva recently settled down. Dorsal view. X 95. Actual length approx. 1.7mm. Fig. 3. Lateral view of anterior body of the same larva as shown in Fig. 10. x 95. Fig. 4. Anterior body of recently metamorpbosed larva. Lateral view. X 95. Fig. 5. Crotchets from the eleventh neuropod of sixteen setiger larva. X1300. Fig. 6. Dorsal seta of recently metamorphosed larva. X 560.. —164.
(12) New Species of the Spionidae (Polychaeta), Nerinides yamagiwhii n, sp.. 165 - -.
(13)
図
関連したドキュメント
We use these to show that a segmentation approach to the EIT inverse problem has a unique solution in a suitable space using a fixed point
Since one of the most promising approach for an exact solution of a hard combinatorial optimization problem is the cutting plane method, (see [9] or [13] for the symmetric TSP, [4]
Kilbas; Conditions of the existence of a classical solution of a Cauchy type problem for the diffusion equation with the Riemann-Liouville partial derivative, Differential Equations,
We present sufficient conditions for the existence of solutions to Neu- mann and periodic boundary-value problems for some class of quasilinear ordinary differential equations.. We
Analogs of this theorem were proved by Roitberg for nonregular elliptic boundary- value problems and for general elliptic systems of differential equations, the mod- ified scale of
Then it follows immediately from a suitable version of “Hensel’s Lemma” [cf., e.g., the argument of [4], Lemma 2.1] that S may be obtained, as the notation suggests, as the m A
In analogy with Aubin’s theorem for manifolds with quasi-positive Ricci curvature one can use the Ricci flow to show that any manifold with quasi-positive scalar curvature or
Correspondingly, the limiting sequence of metric spaces has a surpris- ingly simple description as a collection of random real trees (given below) in which certain pairs of
