別紙様式1
博 士 学 位 論 文
The Effects of Immune Cytokines on Bone Mineral Density and
Osteometabolism
骨代謝・骨密度における 免疫系サイトカインの
作用に関する研究
平成 24 年 7 月 12 日 提出
首都大学東京 大学院
人間健康科学研究科 博士後期課程 人間健康科学専攻
ヘルスプロモーションサイエンス 系 学修番号:08999601
氏 名:大石 芳江
( 指導教員名: 北 一郎 )
THESIS
The Effects of Immune Cytokines on Bone Mineral Density and Osteometabolism
Yoshie Oishi
Department of Health Promotion Sciences, Graduate School of Human Health Sciences,
Tokyo Metropolitan University
SUMMARY
4 Summary
The immune and skeletal systems have the role of host defense and serving as levers for muscle action respectively; however, immune cells are formed and haematopoietic stem cells are maintained in the bone marrow where they interact with bone cells. The immune and skeletal systems share a number of regulatory molecules including cytokines, receptor, signaling molecules and transcription factors.
Although several environmental factors, including diet, smoking, and physical exercise influence BMD, a genetic contribution to this parameter has been recognized, and genes account for 50-60% of the total variation in bone mineral density (BMD). Recent studies have clarified the interaction between bone and immune cells, and found that cytokines are involved with osteometabolism. In study 1, I focused on the possible association between BMD and polymorphism in the
interleukin (IL) 6 -634C>G (rs 1800796), tumor necrosis factor (TNF)α -308G>A (rs 1800629), IL17F 7488T>C (rs 763780), transforming growth factor (TGF)β 869T>C (rs 1800470), osteoprotegerin (OPG) 163A>G (rs 3102735) and methylenetetrahydrofolate reductase (MTHFR) 677C>T (rs 1801133) genes in one hundred young andone hundred elderly Japanese women. It was confirmed that there is a significant difference between each polymorphism of the
IL6 and IL17F genes and BMD in young women. The significance of IL17F genepolymorphism was also indicated in elderly women, therefore polymorphism in the
IL17F 7488T>C (rs763780) may be useful as a new candidate gene for preventing primary osteoporosis. This study of the
IL17F gene is the first report of the results of an analysis of 200 Japanese women.Healthy bone is always restructured that is dependent on the dynamic balance of bone formation and
resorption, which are mediated by osteoblasts and osteoclasts, respectively. In bone,
receptor activator of NF-κB ligand(RANKL) is expressed by osteoclastogenesis-supporting cells, including osteoblasts, in
5
response to osteoclastogenic factors, such as parathyroid hormone
(PTH), TNF-α, IL-6 and IL-17. In
study 2, the bone metabolism was revealed
in vivo by IL-6 and IL-17F which confirmed thepolymorphism of those cytokines related with BMD. Evaluation of the cytokine’s influence was carried
out by the detection of RANKL production using western blot analysis, alkaline phosphatase activity
(ALP) and the mineralization on human featal osteoblast (hFOB) 1.19. As a result, IL-6 promoted
RANKL expression and IL-17F also tended to promote that; furthermore, treatment of both cytokines
tended to suppresses ALP activity such as the marker of bone formation. These results indicated these
possibly promote bone resorpion. Interferon (IFN)γ suppressed bone resorption caused by IL-6, but in the
case of coexistence with IL-17F tended to promote RANKL expression compared to IL-17F alone. These
results implied that, whereas IFNγ has been reported as an antiosteoclastgenic cytokine, it may possible to
promote RANKL expression on osteoblasts in the case of co-existence with other cytokines. On the other
hand the condition of IL-17F with IFNγ suppressed the differentiation of osteoclast precursor cells under
RANKL, this seems to involve an interaction between osteoblasts and osteoclasts with some cytokines for
osteometabolism. My results partially clarify the complex interactions between osteometabolic cells and
single cytokines or the combination of some cytokines for the balance of osteometabolisms. It has
become clear that the complex interactions underlie the relationship between immune and bone cells for
RANKL inhibition including therapeutic strategy and, moreover, shows the clinical relevance of newly
recognized cytokine combinations such as found in these results shows.
INTRODUCTION
6 Introduction
Osteoporosis is widely recognized as an important public health problem because of the significant morbidity and even mortality associated with its complications, including fractures (1, 2). The annual business reports of community and geriatric health care (2007) by the Ministry of Health, Labour and Welfare stated that for people over the age 60 years in Japan 20.7% of the patients who are examined for osteoporosis need to undergo detailed examination. The public health burden of this disease is 1.3 billion yen every year in direct expenditure in Japan – and this is expected to increase dramatically as the population ages (3). Osteoporosis is mainly characterized by low bone mass and microarchitectual deterioration of bone tissue. Peak bone mass is a major factor determining the risk of osteoporotic fracture.
The bone mass attained early in life is perhaps the most important determinant of lifelong skeletal health.
Many factors influence the risk of osteoporosis including diet, physical activity, medication use, coexisting disease, aging, and diminished sex-steroid production. One of the most important clinical risk factors is a positive family history of osteoporosis, emphasizing the importance of genetics in the pathogenesis of osteoporosis. Many studies suggest that genetic factors contribute significantly to the variance in peak bone mass.
Epidemiological studies also support the heritability of bone mineral density (BMD) (4), including the
contribution of the genetic factors to the pathogenesis of osteoporosis. Therefore, the elucidation of
genetic factors for this disease has been awaited. Recently, genetic factors for osteoporosis have been
investigated with polymorphism of genes. Quite a few associated studies have been done with so-called
candidate gene approaches (5) and genome-wide approaches (6). Polymorphism of many genes was
shown to have a significant association with BMD. However, the contribution of each gene in
determining BMD is small and the result is not always reproducible (7, 8). One of the ways to prevent the
7
problems and to make the outcome biologically and clinically relevant would be to use polymorphism in a way that functional variations can be studied. Genetic research and genetic results (including polymorphism) regarding osteoporosis are essential for the prediction, prognosis as well as treatment of the disease.
The bony skeleton enables weight-bearing locomotive activities, the storage of calcium, and the harboring of the hematopoietic stem cells from which blood and immune cells are derived. Although bone appears to be metabolically inert, it is actually a dynamic organ under the active control of osteoblasts and osteoclasts. This process, called the bone remodeling cycle, is essential for the maintenance of a healthy skeleton. It is dependent on the dynamic balance of bone formation and resorption. The novel functions of bone cells in the regulation of cells outside the bone will, therefore, have an impact on many fields of biomedical investigation. In recent years, inflammation has been also implicated. Experimental studies in animals have provided substantial evidence suggesting that certain inflammatory cytokines, including IL-1, IL-6, and tumor necrosis factor (TNF)-α, play an important role in the pathogenesis of osteoporosis (9-13). However, evidence that these inflammatory cytokines play a similar role in human studies is limited. Preliminary studies on the associations between the circulation cytokine levels and bone mineral density (BMD) (14-17) or bone loss (18, 19) in postmenopausal or elderly women have provided inconsistent or even contradictory results. Since immune cells infiltrate into any tissues and organs, bone is not the only tissue linked to the immune system. However, bone has a distinct status as a central immunological organ because hematopoietic stem cells are maintained in the bone marrow (20) and immune cells are derived from that. Thus, immune cells develop in the same microenvironments as osteoclasts (21, 22).
The interaction between the immune and skeletal systems has been recognized for more than three
8
decades. However, almost all investigations were related to rheumatoid arthritis (RA). This has led to the recent emergence and subsequent rapid evolution of the field of osteoimmunology. The scope of osteoimmunology has grown to encompass a wide range of molecular and cellular interactions, an elucidation of which will provide a scientific basis for future therapeutic approaches to diseases of both the immune and skeletal systems. Osteoclasts and immune cells share a number of regulatory molecules, including cytokines, receptors, signaling molecules, and transcription factors, which mutually influence each other. In fact, the bone and immune systems are so closely intermingled that all factors regulate immune cells should be investigated for their effect on bone and vice versa. The mechanism of osteoporosis which occurs postmenopausally is concerned with some cytokines such as IL-1, IL-6, TNF-α and some immune cells such as lymphocytes and macrophages. TGF-β is autocline, paracline factor which exists abundantly in bone matrix like an insulin like growth factor (IGF)-1. Its function is to regulate osteoblast differentiation and promote synthesis bone matrix protein such as collagenⅠ.
I aim to analysis the association of polymorphisms in genes encoding immune cytokines, such as IL-6, TNF-α, IL-17F, TGF-β, OPG and MTHFR, with the risk of developing osteoporosis, as measured by BMD, in young and elderly Japanese women.
I used another method to attain the aim such as using cell culture with human osteoblasts and osteoclasts.
Bone resorption is regulated by various cytokines. In postmenopausal osteoporosis, bone loss due to
estrogen deficiency is closely related to the production of bone-resorbing cytokine. Especially, the
increased production of IL-1, IL-6 and TNF-α could induce the expression of RANKL in bone tissues to
enhance osteoclastogenesis. Relationship between various cytokines and osteoclastgenesis is important to
clarify on osteometabolism. Therefore I examined the effect of IL-6, IL-17F (promoting RANKL
expression), IL-4 and IFNγ (suppressing osteoclastgenesis) in cell culture can be evaluated by their
9
concentration and their combination. In bone tissue, some cytokines stimulate both osteoblasts and
osteoclasts, therefore the combination of some cytokines play a role in both cells to regulate
osteometabolism. It is possible that some cytokines could synergize to enhance or suppress the bone
remodeling.
CHAPTER Ⅰ
10
Chapter Ⅰ Literature Review
1. Genetic influence on osteoporosis
Genetic factors have long been recognized as playing an important role in the pathogenesis of osteoporosis. Evidence from twin and family studies suggests that between 50% and 85% of the variance in peak bone mass is genetically determined, depending on skeletal site and the age of the subjects (4, 22, 23, 24). Heritability studies have also shown evidence of significant genetic effects on the other key determinants of osteoporotic fracture risk including quantitative ultrasound properties of bone (25), femoral neck geometry (25), muscle strength (26), bone turnover markers (27), and body mass index (28).
The role of genetic factors in the pathogenesis of osteoporosis is less clear. Many studies on the genetic contribution to age-related bone loss have yielded mixed results. However, it is currently believed that in the general population, genetic influences on BMD and the other phenotypes mentioned above are polygenic in nature and are mediated by the influence of several genetic variants, each of modest effect size, and their interaction with environmental factors. These results suggested that variations in other genes involved in the immune system (described in the next section) could also affect BMD and influence susceptibility to osteoporosis. Some of the factors that concerned with bone metabolism, reported as polymorphism influencing, are described below.
1) Vitamin D Receptor
Vitamin D, through its principal bioactive form 1, 25-dihydroxyvitamine D
3(1, 25-(OH)
2D
3) plays a
crucial role in bone metabolism. The action of 1, 25-(OH)
2D
3is mediated through a specific
hormone-receptor (VDR) that regulates gene expression by forming a heterodimer with retinoic X
receptor (RXR) that binds to vitamin D response elements in target genes. The VDR is the first candidate
11
gene to be studied in relation to osteoporosis. One polymorphism affecting exon 2 of
VDR has beendescribed that creates an alternative translational start site, resulting in the production of two isoforms of the VDR protein, which differ in length by three amino acid (29).
2) Estrogen Receptor (ER)-α
The estrogen receptor-α, encoded by the
ESR1 gene, is another important functional candidate for theregulation of bone mass. A large number of investigators have looked for evidence of an association between
ESR1 alleles and BMD, mostly focusing on two polymorphisms within intron 1 recognized by XbaⅠand PvuⅡ restriction enzymes, and on a TA repeat in the promoter. Studies of ESR1 alleles inrelation to BMD have yielded inconsistent results, possibly because most studies have been of small sample size and involved subjects of different ages, menopausal status, and ethnic backgrounds (30).
3) Interleukin (IL)-6
IL-6 is a multifunctional cytokine essential to regulation of the immune response, hematopoiesis, and bone resorption. It exerts its actions through binding to its cell-surface receptor, IL-6 receptor. IL-6 and its receptor stimulate osteoclast development, and thereby, the process of bone resorption (9).
Polymorphisms of the IL6 gene have been identified in Japanese concerning the promoter region at nt-634 associated with radial BMD (31), and at nt-174 which was related to previous fractures, particularly osteoporotic fractures, in elderly postmenopausal women.
4) Collagen typeⅠα1 (COLIA1)
The gene encoding the αⅠchain of typeⅠcollagen (COLIA1) is also an important functional candidate
for the pathogenesis of osteoporosis, because typeⅠcollagen is the major protein of bone and since
mutations in this gene cause the syndrome of osteogenesis imperfecta - a rare disease characterized by
increased bone fragility and reduced BMD (32). Previous research has focused on a polymorphism within
12
intron 1, which is situated at a binding site for the transcription factor Sp1. The COLIA1 Sp-1binding site polymorphism has been associated with various osteoporosis-related phenotypes, including bone density (33, 34), fragility fractures (33, 34), postmenopausal bone loss (35, 36), and bone mineralization (37).
5) Lipoprotein receptor-related protein (LRP)5
The Lipoprotein receptor-related protein (LRP)5 and LRP6 receptors are transmembrane proteins that function as coreceptors for canonical Wnt signaling. Wnt signaling plays an important role in several key developmental processes, including cell fate decisions, limb patterning, and osteoblast and chondrocyte differentiation, and the development of the central nervous system and other organs (38). The LRP pathway was discovered to be a key regulator of bone mass following linkage studies in two rare human diseases: osteoporosis-pseudoglioma syndrome (OPPS), which is a recessively inherited condition characterized by severe, early onset osteoporosis and congenital blindness due to vitreous opacity (39), and the high bone mass (HBM) syndrome that is an asymptomatic autosomal dominant disorder characterized by increased bone mineral density (40). There is evidence that more subtle variations in LRP5 could underlie variations of BMD in the normal population. Many variants have been studied, the most likely functional candidate being an alanin to valine amino acid substitution at position 1330 (A1330V). The data indicate that genetic variation in LRP5 and possibly LRP6 plays an important role in the regulation of bone mass and osteoporotic fractures in humans.
6) Vitamin-K-dependent gamma-glutamyl carboxylase
Vitamin-K-dependent gamma-glutamyl carboxylase (GGCX) is necessary for post-translational
modification of vitamin-K-dependent proteins to exert their functions. The genomic structure of GGCX
was elucidated in 1997 (41). Concurrent with conversion of glutamate residue (Glu) to gamma-carboxyl
glutamate residue (Gla), vitamin K hydroquinone is oxidized to vitamin K 2, 3-epoxide by GGCX, then
13
vitamin K 2, 3-epoxide is reduced to vitamin K hydroquinone by 2, 3-epoxide reductase (VKOR) (42).
Osteocalcin and matrix Gla protein (MGP) are also vitamin-K-dependent proteins (43). Osteocalcin and MGP are abundant in bones (44) and assumed to play important roles in bone metabolism. The rare mutations of the GGCX gene with amino acid substitution cause consequential abnormal enzyme activity (45, 46). The different activities of GGCX between the common genotypes which were associated with BMD in elderly Japanese women have also been reported (47).
7) Carbonic anhydrase
Carbonic anhydrase (CA) is a family of zinc metal enzymes that catalyze the reversible hydration of CO
2in the reaction CO
2+H
2O
⇔ HCO3-+H
+(48). Fourteen isozymes in the human CA family have been reported, including 11 functional CAs (49). CAs are essential for pH regulation by the osteoclasts.
Clinically, use of carbonic anhydrase inhibitors has been known to be associated with a bone-sparing effect as judged by spinal BMD. Single nucleotide polymorphisms (SNPs) in carbonic anhydrase genes that are expressed in bone tissues, i.e., CA8 and 10, have been studied for possible association with femoral neck and lumbar spine BMD (50).
2. Physiological bone regeneration
Bone is composed of collagen type
Ⅰ(90%), noncollagenous proteins (10%) and minerals. It isstructurally classified into two types: cortical or compact bone and cancellous or trabecular bone. Bone can exchange H
+for Ca
2+and K
+(ion exchange) or release HCO
3-
, CO
3-
or HPO
4-2
. In acute metabolic acidosis, uptake of H
+by bone in exchange for Na
+and K
+release is involved in buffering and this can occur rapidly without any bone loss (51).
Bone is a highly specialized and dynamic organ that undergoes continuous regeneration. It consists of
14
highly specialized connective tissue matrix, and spaces that include the bone marrow cavity, vascular canals, canaliculi, and lacunae (52). During development and growth, the bone is sculpted to achieve its shape and size by the removal of bone from one site and deposition at a different one; this process is called modeling. Once the skeleton has reached maturity, regeneration continues in the form of a periodic replacement of old bone with new at the same location (53). This process is called remodeling with two main types of cells being responsible for bone renewal: osteoblasts and osteoclasts. Removal of bone (resorption) is the task of osteoclasts. Formation of new bone is the task of osteoblasts. Bone resorption and bone formation, however, are not separate, independently regulated processes. In the uninjured adult skeleton, all osteoclasts and osteoblasts belong to a unique temporary structure, known as a basic multicellular unit or basic multicellular unit (BMU) (54).
1) Features of osteoblasts and their role in osteometabolism
An osteoblast is a bone lining cell which originates from the mesenchymal stem cell (bone marrow stromal cell or connective tissue mesenchymal stem cell) (55), which also gives rise to bone marrow stromal cells, chondrocytes, muscle cells, and adipocytes (56, 57, 58). Hormones, cytokine and growth factors such as PTH, bone morphogenetic protein (BMP), fibroblast growth factor (FGF), insulin-like growth factor (IGF) and Wnt protein act on osteoblasts and control their functions, proliferation and differentiation, and they also have receptors for PTH, 1, 25-(OH)
2D
3and estrogen.
Osteoblasts produce collagen typeⅠ(Fig.2), which accounts for 90% of the total bone proteins and
ground substances. The other 10% of proteins include osteonectin, osteocalcin and cytokine such as
TGF-β, OPG, RANKL, and macrophage colony-stimulating factor (M-CSF), the last two of which are
necessary for growth and differentiation of osteoclasts. Under the light microscope, osteoblasts are seen
with a round nucleus at the base of the cell which is opposite to the bone surface. The cytoplasm is
15
strongly basophilic, and is seen with prominent golgi complex (Fig.1). The plasma membrane of the osteoblast is rich in alkaline phosphatase, which is used as a bone formation marker.
2) Features of osteoclasts and their role in osteometabolism
Osteoclasts are the only cells that play the role of resorption. Osteoclasts originate from the hematopoietic stem cells and are giant multinucleated cells, formed by the fusion of up to twenty cells. A mature osteoclast is about 50 to 100 μm in diameter, with abundant mitochondria, numerous lysosomes and free ribosomes. Multinucleated osteoclasts, after being recruited by M-CSF and RANKL, adhere to bone and undergo differentiation into mature osteoclasts. RANKL stimulates osteoclast activation by inducing the secretion of protons and lytic enzymes into a sealed resorption vacuole formed between the basal surface of the osteoclast and the bone surface (Fig.3).
The most remarkable morphological feature of an active osteoclast is the ruffle border, a complicated system of finger-shaped projections of the membrane, which is completely surrounded by another specialized, clear zone. The ruffle border contains bundles of actin-like filaments, which can attach and seal off a distinct area lying resorbing environment from the surrounding effective capillary flow (ECF) (Fig.3). One possibility is that osteoclast precursors recognize some changes (mechanical stress, microdamage or hypermineralization, decreasing calcium concentration in peripheral blood) in the bone surface and sends signals to activate osteoclasts. The final activation of the osteoclast may occur in the bone matrix that contains arginine-glycine-asparagine (RGD) amino acid sequences such as osteopontin, that is because the interactions occur between integrins on osteoclast cell membranes with protein (59).
Moreover, the lining cells can be stimulated by endocrine signals such as PTH, PTH-related peptide (PTHrP), and 1, 25-(OH)
2D
3to generate local paracrine signals for promoting osteoclast differentiation.
Furthermore, a calcium signal is essential for osteoclast differentiation.
16
3) Remodeling by the basic multicellular unit (BMU)
During childhood and the beginning of adulthood, bone becomes larger, heavier and denser by the process of bone modeling, in which formation is at a higher rate than resorption. Even after bone has attained the adult shape and size in a fully-grown body, renewal of bone is essential for maintenance of bone strength throughout life. Old bone is removed (resorption) and new bone is created (formation) all the time, and the process is known as bone remodeling. This process occurs by dissolving a pit in the bone surface and then filling it with new matrix. The activation is an event in the remodeling sequence in which the bone surface is converted from the original stage to one in which circulating mononuclear cells of the hematopoietic lineage begin to congregate and fuse together to form differentiated osteoclasts. The exact mechanism and specific site of activation are still not understood. (Illustrated in Fig. 1)
①
Bone formation
Osteoblasts begin a two-phase process for Ca
+deposition. The first stage of the mineralization process starts with the generation of 100 nm matrix vesicles. These are cytoplasmic membrane-bound vesicles that are created near the osteoid-osteoblast interface. These matrix vesicles contain a variety of substances, including annexinⅤ(on the inner membrane surface), alkaline phosphatase (glycosylphosphatidylinositol;
GPI-linked), calbindin D9k, pyrophosphatases, carbonic anhydrase, adenosine mono phosphate (AMP) ase, bone sialoprotrein-1 (BSP-1), osteonectin, and osteocalcin. The matrix vesicles are extruded from the osteoblasts in annexin, from channels that transmit Ca
2+-dependent a process where they attach to the underlying matrix (60). Both alkaline phosphatase and annexin are reported to bind to typeⅠcollagen, and these molecules, if expressed on the outer surface of the matrix vesicle, may provide the anchor for osteoblast-derived matrix vesicles (61). Once externalized, the matrix vesicle undergoes physical changes.
Both Ca
2+and Pi are transported into the vesicle.
17
This is accompanied by phosphate uptake, which is tied to external Pi generation via alkaline phosphatase activity (60, 62, 63). Uptake of Pi occurs via Na-dependent Pi transport system Type 3 (NPT3)/Pit1 Na/Pi cotransporters (53, 54, 55, 56, 64, 65). Once internalized, Ca
2+and Pi form rudimentary Cap crystal (amorphous Cap). Early formation is predicated upon initial Ca
2+and Pi interaction with phosphatidylserin (63, 66, 67, 68, 69). The first crystal formed is referred to as an octa-CaPi-like mineral and does not yet qualify as organized hydroxyapatite (Ca
10(PO
4)6(OH)
2) (57, 58). In the presence of elevated intra-vesicle Ca
2+and Pi, crystal growth occurs internally on the Octa-Capi-like complex, generating early hydroxyapatite. Undoubtedly, some of the gamma-carboxy-expressing molecules regulate these initial phases of the process. Events to this point constitute the first phase of hydroxyapatite formation (70). In the second phase, the enlarging CaPi complex actually begins to penetrate the matrix vesicle membrane. Complete matrix vesicle rupture does not seem to occur until later. Rather, phospholipase punch holes in the plasma membrane, exposing the nascent hydroxyapatite to the extracellular fluid. Factors that regulate the continuing growth of the hydroxyapatite include Ca
2+and Pi concentration, local pH, and the presence of non-collagenous matrix proteins (70). At a minimum, these proteins include BSP-1, osteocalcin, and osteonectin. They also include osteopontin, which would seem to play a major, if not crucial, role in large-scale hydroxyapatite formation. Osteopontin is a secreted phosphoglycoprotein that is induced by 1, 25-(OH)
2D
3(66, 71, 72, 73). This molecule has an RGD sequence that may provide anchorage for cells. It is also variably phosphorylated, the degree to which apparently impacts its function. When 40% of its phosphorylation sites are filled, it blocks mineralization.
When 95% of its sites are filled, it appears to promote hydroxyapatite formation (73). Whether 95%
phosphorylated osteopontin occurs regularly in bone is unknown (71, 73). It has been suggested that
osteonectin and osteocalcin interact and expose Ca
2+-binding nucleation sites on osteopontin necessary
18
for hydroxyapatite formation (59). (Illustrated in Fig.2)
② Bone resorption
The process of bone resorption is regulated by the osteoclasts, a multi-nucleated, monocyte-derived cells that respond to both pH and PTH/low extracellular Ca
2+. Although cytokine-induction of osteoclasts has been a favorite topic in experimental biology over the last decade, acidosis and the response to acidosis by osteoclasts is perhaps just as important. Osteoclasts express a proton-sensing G protein-coupled receptor termed ovarian cancer G protein-coupled receptor (OGR) 1/ G protein-coupled receptor (GPCR)1. In the presence of low pH, this receptor is activated and increases intracellular Ca
2+. This begins the process of bone resorption with demineralization. Hydroxyapatite contains a considerable amount of stored OH
-ions. During metabolic acidosis, where renal and respiratory homeostatic mechanisms are inadequate, bone is broken down to provide buffering capacity to the blood/extracellular fluid (74). The osteoclasts first generate a sub-osteoclastic space between the osteoclasts and underlying bone. Carbonic anhydraseⅡin cytoplasm of osteoclasts catalyses H
2O and CO
2to proton and hydrogen carbonate. A proton pump/transporter (V-ATPase and/or Na
+/H
+) linked to K
+/Cl
-co-transporters/channels (KCC) and chloride channel-7 (CLC7) facilitates the release of H
+into sealed space or lacuna (75). This dissolves the underlying mineral and generates high local levels of Ca
2+and Pi. Tartrate-resistant acid phosphatase (TRAP) and cathepsin K are also secreted, and these enzymes digest bone mineral and collagen matrices together with metalloproteinase 9 and collagenase (76, 77). The high amount of the phosphohydolase enzyme, TRAP, is commonly used for the morphological detection of osteoclasts in bone specimens (78). The residues from this extracellular digestion are either internalized or transported across the cell, possibly induced by a calcium-sensing receptor, to the bone-resorbing compartment (79).
The residual collagen fibers are digested either by collagenase or cathepsins at low pH. Since collagen
19
typeⅠis enriched in hydroxyproline in the urine and in N-terminal collagen peptides in serum, they can be used as indirect markers of bone resorption in humans.
There appear to be at least three negative-feedback mechanisms associated with osteoclast activity. The first involves released Ca
2+, which it has been suggested enter the osteoclasts via transient receptor potential vanilloid (TRPV) channels (80). What opens the channels is not clear. Osteoclasts have Ca receptors associated with ryanodine receptor 2 (RyR2), and this complex may initiate Ca
2+transit through the osteoclasts into the extracellular fluid. Regardless, Ca
2+-influx is reported to induce apoptosis in osteoclasts, potentially limiting the time the bone is exposed to osteolytic agents. Clearly, intracellular Ca
2+-binding agents such as calbindin may modulate the effects of any increased cytoplasmic Ca
2+levels (80-84). A second potential negative-feedback mechanism involves platelet-derived growth factor -BB (PDGF-BB). In this scenario, RANKL on osteoblasts will bind to RANK on osteoclast precursors, generating functional osteoclasts. Osetoclasts now secrete a PDGF-BB homodimer that has a down-modulation effect on osteoblast activity. This includes a reduction in typeⅠcollagen production and osteocalcin secretion. Perhaps most importantly, PDGF-BB induces osteoblasts to secrete OPG, a natural soluble receptor for RANKL (85). This blocks RANKL activity on osteoclast precursors and limits osteoclast development (80). The effect is amplified by the presence of 1, 25-(OH)
2D
3(a bone anabolic agent), which also promotes OPG secretion by osteoblasts (86). A final mechanism for osteoclast down-regulation involves osteoclast production of IFNβ, an autocrin inhibitor of osteoblast development.
IFNβ, acting through the typeⅠinterferon receptor, reduces c-fos, a transcription factor needed for
continued osteoclast activity (75). (Illustrated fig. 3)
20
Blood vessel
Bone
Osteoclast precursor
Osteoblasts
Low extracellular Ca
2+PTH 1,25-(OH)
2D
3RANKL RANK
Osteometabolism
Osteoclasts Multi nucleated
Osteoblasts
Osteoclasts
Fig. 1 Osteometabolism
21
Type Ⅰ collagen
Hydroxyapatite formationMatrix vesicles
Extrude
Ca2+
HPO42-
Cap crystal
Proteoglycan
Crystal growth Carbonic anhydrase
Annexin V Carbindin D9K
protease
Bone formation
Osteoblast
Penetrate Matrix vesicle
Fig. 2 Bone formation
Calbindin D9K
22
Bone surface
Capillary
Integrin αVβ3 RANKL
RANK
Ruffle border Clear zone
Lysosomes
Cathepsin K MMP 9 collagenase
Cl-transporter/
channels
CO
2H
2O
H
2CO
3H
+Lucuna
carbonic anhydrase Ⅱ
HCO
3-ATP
HCO
3-Cl
-Calcium phosphate
Ca
2, Pi
Bone resorption
Cl
-H
+Proton pump
Osteoclast
Ca
2,Pi
Macrophage
M-CSF
Clear zone Osteoclast
activation
Fig.3 Bone resorption
Ca
2+, Pi
Ca
2+, Pi
23 3. Cytokine effects on osteometabolism
The cytokines and growth factors that are involved in origination and the bone-derived attract osteoblasts.
Osteoblasts are generated from the actions of M-CSF, RANKL, and likely, IL-1 on osteoclast precursors.
M-CSF is secreted by immature osteoblasts and promotes the formation and proliferation of osteoclast precursors from colony forming unit-megakaryocyte (CFU-M) (75, 87). In the presence of elevated 1,25-(OH)
2D
3, IL-11 or hypocalcemic condition that induces PTH release, osteoblasts will be stimulated to express RANKL on the surface (75, 76). RANKL is a 45kDa, typeⅡtransmembrane glycoprotein that belongs to the TNF superfamily and contains a C-terminal receptor-binding domain and transmembrane domain. It is expressed by osteoclastogenesis-supporting cells, including osteoblasts, in response to osteoclastogenic factors, such as 1, 25-(OH)
2D
3, prostaglandin E
2(PGE
2) and PTH, and is a crucial determinant of the level of resorption in vivo (88).
When expressed, it binds to either constitutively expressed, or M-CSF-induced, RANK on the surface of osteoclast precursors (79). In synergy with M-CSF, RANKL induces differentiation to “pre-fusion”
osteoclasts that ultimately fuse under the influence of M-CSF, IL-1, and RANKL. Fusion forms a mature, multi-nucleated osteoclast (75, 76). Finally, RANKL and IL-1 activate the osteoclast, initiating bone resorption. It should be noted that pro-inflammatory cytokines such as IL-1β and TNF-α will also induce RANKL expression. Although the expression is on a T cell rather than osteoblasts, the effect is the same:
osteoclast development (89, 90). Osteoclast-differentiation factor expressed by osteoblasts would be that
same molecule expressed by T cells to stimulate dendritic cells (91-94). RANKs is a typeⅠmembrane
protein sharing high homology with CD40, and the binding of RANKL to RANK is inhibited by the
decoy receptor OPG (95, 96). On the other hand, the decreasing levels of estrogen during menopause
have implications for the functions of many organs in the body. Although it is likely that different
24
pathogenesis mechanisms are involved, at least partly, in the effects of estrogen on bone, the cardiovascular system, and adipose tissues, there is much interest in the role that cytokines may play in the effects of estrogen deficiency in all these organs. Estrogen is a potent inhibitor of IL-1β and TNF-α production in bone marrow cells and monocytes (97). It also inhibits the production of IL-6 in stromal cells and osteoblasts (98). Inflammatory cytokines, such as IL-1, IL-6 and TNF, which have a potent capacity to induce RANKL expression on osteoblasts and accelerate RANKL signaling, thus directly contributing to the bone destruction process (99). On the other hand, TGF-β, which is one of the most abundant cytokines in bone matrix and, it is enhancing the production by estrogen. The expression of TGF-β by estrogen decrees bone resorption (100). In addition to that, here are some cytokines concerned with osteometabolism: e.g. the IL-17-producing Th-cell subset (Th 17cells), and it represents the long-sought-after osteoclastgenic Th-cell subset (101). The infiltration of Th 17 cells into the inflammatory lesion links the abnormal T-cell response to bone damage, and the pathogenesis of autoimmune arthritis should be reconsidered in the context of a Th 17-type disease. Clearly, this subset will be an auspicious target for future therapy, and cytokines related to Th 17-cell differentiation and expansion, such as IL-6, TGF-β and IL-23, will be of great clinical importance. It has been reported that IL-11 is essentially involved in the early onset of osteoporosis, where enhanced adipogenesis due to decreased production of IL-11 impairs bone metabolism, resulting in osteoporosis (102). Other information concerned with IL-11 is that mechanical unloading suppresses and reloading enhances
IL11gene expression in the hindlimb of mice
in vivo. Mechanical stress to osteoblastogenesis and suppressionof adipogenesis suggests the requirement of the ΔFosB/JunD complex (transcript factor) to the activator protein (AP)-1 site of the IL-11 for the stimulation of osteoblast differentiation by mechanical stress.
IL-11 may play a critical, hierarchical role in osteoclast development and activation (103). In cultures
25
employing mouse bone marrow, IL-11 has been observed to stimulate osteoclast formation.
The nuclear factor of activated T-cells, cytoplasmic 1 (NFATc1), is another important target controlling excessive osteoclastogenesis. The activation of NFATc1 is mediated by the phosphatase calcineurin, which is activated by calcium-calmodulin signaling in osteoclasts (104, 105, 106).
The features of some cytokines that affect bone metabolism are described below.
1) Function of interleukin (IL)-6 and its role in osteometabolism
IL-6, a multipotent cytokine with a wide variety of activities, is produced by osteoblastic cells (107) and other tissues (98, 108). Although IL-6-mediated bone resorption varies depending on the
in vitro assaysystem (109, 110), it has been shown to regulate the development of matured osteoclasts (9), and to directly stimulate the production of RANKL and OPG mRNA as well as prostaglandins in bone (111, 112). IL-6 appears to mediate the increase in bone resorption and pathologies that characterize various clinical syndromes, including Paget’s disease (113), hypercalcemia associated with malignancy (114), fibrous dysplasia (115), giant cell tumors of bone (116), and Gorham-Stout disease (117).
When experiments were conducted on mice, bone loss was associated with estrogen depletion (98). The production of IL-6 is suppressed by both estrogen and androgen, as well as the expression of its receptor (118). The level of IL-6 is elevated in bone marrow and in the peripheral blood of estrogen-deficient mice, rats and humans (119, 120). The roles of the IL-6 cytokine family in osteoclast formation should be examined based on data demonstrating that mice lacking the gp130 activator protein have increased osteoclast numbers (121). Because gp130 transduces signaling for all of the IL-6 family members, this result suggests that at least some of these factors inhibit osteoclast formation and bone resorption.
2) Function of tumor necrosis factor (TNF)-α and its role in osteometabolism
TNF-α has been shown to stimulate bone formation and bone resorption
in vivo, and enhance the26
formation of osteoclast-like cells (OCLs) in bone marrow cultures (88, 122-125). The ability of TNF-α to stimulate osteoclast formation in mixed stromal cell/ osteoclast precursor cell cultures is shown to be IL-1 dependent (125), whereas TNF-α-induced osteolysis was found to be dependent on M-CSF (126).
Lymphotoxin and TNF are molecules that are related functionally to IL-1. They share the same receptor which is distinct from the IL-1 receptor. Many of their biological properties overlap with those of IL-1;
however, their effects on bone are synergistic with those of IL-1. They cause osteoclast resorption and hypercalcemia by stimulating proliferation of osteoclast progenitors, and cause fusion of committed precursors to form multinucleated osteoclasts (13).
Cultures of cells from RANKL-deficient mice suggested that TNF-α directly stimulates osteoclast formation independent of RANK (127), although TNF-α administration produced only a few osteoclasts in RANK-deficient mice (128); it also inhibits osteoblast differentiation and collagen synthesis (129, 130, 131).
3) Function of transforming growth factor (TGF)-β and its role in osteometabolism
Bone morphogenetic proteins (BMPs), members of the TGF-β superfamily, induce bone formation both in
vitro and in vivo. Signaling cross-talk between BMP and TGF-β pathways plays a crucial role in theregulation of osteoblastic differentiation, and TGF-β inhibitors may be invaluable for the treatment of various bone disease by accelerating BMP-induced osteogenesis. TGF-β, one of the most abundant cytokines in the bone matrix, has positive and negative effects on bone formation, although the molecular mechanisms of these effects are not fully understood. TGF-β inhibits osteoblast differentiation through modulation of the expression and transcriptional activity of runt-related gene (Runxs)2.
TGF-β is related to epidermal growth factor (EGF) and is a powerful stimulator of osteoclastic bone
resorption (132). TGF-β is produced by osteoblast, osteoclast and cancer cells. TGF-β is a potent
27
regulatory cytokine with diverse effects on hemopoietic cells (133). The pivotal function of TGF-β in the immune system is to maintain tolerance via the regulation of lymphocyte proliferation, differentiation, and survival. In addition, TGF-β controls the initiation and resolution of inflammatory responses through the regulation of chemotaxis, activation, and survival of lymphocytes, natural killer cells, dendritic cells, macrophages, mast cells, and granulocytes. The regulatory activity of TGF-β is modulated by the cell differentiation state and by the presence of inflammatory cytokines and costimulatory molecules.
Collectively, TGF-β inhibits the development of immunopathology to self or nonharmful antigens without compromising immune responses to pathogens.
4) Function of interleukin (IL)-17 and its role in osteometabolism
IL-17 is a CD4
+T cell-derived cytokine that promotes inflammatory responses in cell lines and is elevated in rheumatoid arthritis, asthma, multiple sclerosis, psoriasis and transplant rejection. Human IL-17F exists as glycosylated 20-to 30kDa homodimers. IL-17 has been shown to signal through the IL-17 receptor molecule and promote production of TNF-α, IL-1β, IL-6, IL-8, and G-CSF (134-136). Human sequence of IL-17F has been described from lymphocytes and patient with asthma (137, 138), IL-17 increased PGE
2synthesis in cocultures of bone marrow cells and osteoblasts and in single cultures of osteoblasts, but not in single cultures of bone marrow cells. In addition, IL-17 dose-dependently induced expression of RANKL mRNA in osteoblasts. Level of IL-17F in synovial fluids was significantly higher in rheumatoid arthritis (RA) patients than osteoarthritis (OA) patients (184).
5) Function of interleukin (IL)-4 and its role in osteometabolism
IL-4 is an immunoregulatory cytokine found to influence skeletal metabolism (139). Produced and
secreted by activated T lymphocytes and mast cells, this 20kDa glycoprotein is well-recognized growth
and/or differentiation factor for a wide variety of cells of hematopoietic lineage. IL-4 was originally
28
characterized on the basis of its capacity to co-stimulate B cell growth with anti-IgM antibodies (140), but it is known that this cytokine can affect antibody isotype and antibody production, T cell activation, mast cell growth, monocyte activation, and hematopoiesis (141).
The potential for IL-4 to affect bone resorption was initially documented by Watanabe
et al. (139), whodemonstrated that IL-4 inhibited
in vitrobone resorption induced by a wide variety of agents, such as PTH, PTHrP, 1,25 (OH)
2D
3, IL-1α and β, prostaglandin E
2(142).
6) Function of interferon (IFN)γ and its role in osteometabolism
T-cell-derived IFNγ has been shown strongly suppresses osteoclastgenesis by interfering with the RANKL-RANK signaling pathway. IFNγ induces rapid degradation of the RANK adaptor protein, tumor necrosis factor receptor associated factor (TRAF)-6, which results in strong inhibition of the RANKL-induced activation of the transcription factor NF-κB and c-Jun N-terminal kinase (JNK). This inhibition of osteoclastgenesis is rescued by over-expressing TRAF-6 in precursor cells, which indicates that TRAF6 is the target critical for the IFNγ action. It has been reported that the accelerated degradation of TRAF6 requires both its ubiquitination, which is initiated by RANKL, and IFNγ induced activation of the ubiquitin-proteasome system (143). The balance between RANKL and IFNγ action may regulate the osteoclast formation. For example, during acute immune reactions as enhanced production of IFNγ counterbalances the RANKL, the augmentation of RANKL expression and reduces aberrant osteoclast formation.
4. Receptor activator of NF-κB ligand (RANKL) and Osteoprotegerin (OPG) regulate osteometabolism
1) Features of RANKL and OPG protein
RANKL is a member of the TNF ligand family, originally identified as TNF-related activation induced
29
cytokine (TRANCE). It has been recognized in two forms, a 40 to 45kDa cellular membrane-bound form and a 35kDa soluble form caused by cleavage of full molecules. Activation of RANK by its ligand, RANKL, leads to the expression of specific genes which in turn induces differentiation of osteoclast precursors into mature osteoclasts.
OPG is a peptide of 380 amino acids, that is secreted as a soluble protein from osteoblasts. It is a TNF receptor-related protein with mRNA expression in a variety of tissues including kidney, lung, heart, liver, stomach, intestines, thyroid gland, brain and spinal cord, and bone. The potential action of OPG in these other tissues beside bone remains unclear. The major biological role of OPG is to block bone resorption by inhibiting osteoclast differentiation and activity both in vivo and in vitro.
2) Mechanism of action of OPG and RANKL
OPG and RANKL have dominant roles in osteoclast regulation. RANKL is the attachment of osteoclasts to bone (144), their subsequent activation (145, 146), and their survival (147). As expected from its mechanism of action, OPG inhibits the formation (95, 145), attachment to bone (148), activation (146, 149) and survival of osteoclasts (147). OPG rapidly decreases osteoclast numbers (114) and increases osteoclast apoptosis (147). The ratio of RANKL: OPG may be the ultimate determinant of bone resorption.
In many situations, bone resorption is stimulated by both increased RANKL and decreased OPG, which can amplify pro-resorptive signals.
3) Regulation of RANKL and OPG on bone density, geometry and strength
RANKL and OPG have consistent and opposing influences on bone density, geometry and strength in a
wide variety of animal models and species (150). Preclinical studies have highlighted the skeletal benefits
of RANKL inhibitors in diverse disease models including bone metastasis (151), rheumatoid arthritis
(152), ovariectomy (153) and inflammatory bowel disease (154). OPG also increases bone strength. The
30
focus on RANKL inhibitors in these models might be related to the extremely rapid bone loss associated with skeletal unloading, particularly at cortical sites (155). In contrast, OPG significantly increased the density and strength of cortical bone (156) in rats.
5. The role of calcium signaling on osteometabolism
RANKL activates TRAF6, c-Fos, and calcium-signaling pathways in osteoclasts, all of which are
indispensable for the induction and activation of NFATc1, the master transcription factor for
osteoclastogenesis. The autoamplification of
NFATc1 gene results in the efficient induction ofosteoclast-specific genes. An AP-1 transcription factor complex containing c-Fos plays a crucial role in
these processes. Jun family members have a redundant role. The immunoreceptor tyrosine-based
activation motif (ITAM) is an important signaling component for a number of receptors and Fc receptors,
but its contribution to the skeletal system remains unclarified. The calcium-mobilizing mechanism during
osteoclastogenesis, multiple immunoglobulin-like receptors associated with ITAM-harboring adaptors, Fc
receptor common γchain (FcRγ), and DNAX-activating protein (DAP) 12, are essential for
osteoclastogenesis (157). The phospholipase C (PLC) family is also related to calcium signaling that
regulates PKC activation, and intracellular calcium levels. PLC cleaves the membrane phospholipid
phospatidylinositol-4, 5-biphosphatase (PIP2) into the second messenger molecules inositol-1, 4,
5-triphosphate (IP3) and diacylglycerol (DAG). IP3 directly increases intracellular calcium levels by
inducing the release of endoplasmic reticulum calcium stores, while DAG activates PKC at the plasma
membrane (158).
CHAPTER Ⅱ
31
Chapter Ⅱ Analysis of correlation between bone mineral density and polymorphic variants of immune cytokines in young and elderly Japanese women (Study 1)
1. Aim
Peak bone mass is a major factor determining the risk of osteoporotic fracture. Bone mass attained early
in life is perhaps the most important determinant of lifelong skeletal health. Many studies suggest that
genetic factors contribute significantly to the variance in peak bone mass. I could evaluate the
polymorphism variants that tend to cause osteoporosis. However, many studies on the genetic
contribution to age-related bone loss have yielded mixed results. As a new viewpoint, I focus on the
polymorphism variants of immune cytokine that affect osteometabolism for finding genes associated with
BMD. In addition, recent studies have highlighted the interaction between bone and immune cells, and
many of the soluble mediators of immune cells, including cytokines, chemokines, and growth factors,
regulate the activation of cells that control bone turnover. Inflammatory cytokines, such as IL-1, IL-6 and
TNF-α, have a potent capacity to upregulate RANKL expression on osteoblasts and accelerate RANKL
signaling, thus directly contributing to the bone destruction process (99). Some cytokines concerned with
osteometabolism include the IL-17-producing helper T (Th) cell subset (Th17 cells), and this represents
the long-sought-after osteoclastgenic Th cell subset (101). The infiltration of Th17 cells into the
inflammatory lesion links the abnormal T cell response to bone damage. TGF-β, which is one of the most
abundant cytokines in bone matrix, and enhances the production by estrogen (100). OPG is secreted as a
soluble protein from osteoblasts. It acts by competing with receptor RANK, a surface receptor expressed
on osteoclasts and dendritic cells, for binding to RANKL. The major biological role of OPG is to block
bone resorption by inhibiting osteoclast differentiation and activity, which has been demonstrated in vivo
and
in vitro (95). The MTHFR gene has been identified as the candidate gene for osteoporosis (159). A32
severe deficiency or lack of MTHFR leads to homocysteinuria, including skeletal abnormalities and early onset osteoporosis.
The aims of the present study were to analysis the association of polymorphisms in genes encoding immune cytokines, such as
IL-6, TNFα, IL17F, TGFβ, OPG,and MTHFR, with the risk of developing osteoporosis, as measured by BMD, in young and elderly Japanese women.
2. Methods
1) Participants
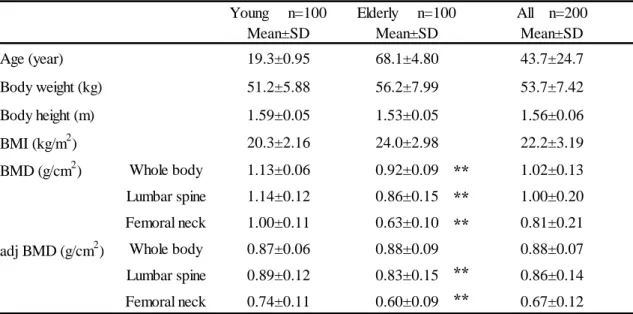
One hundred young women (aged 18–23 years old, mean ± standard deviation 19.3 ± 0.95 years) without
evidence of metabolic disorders (assessed by questionnaire) and one hundred elderly women (aged 60-83
years old, mean ± standard deviation 68.1 ± 4.80 years) participated in this study. The young participants
were enrolled from among students attending Tokyo Metropolitan University (Tokyo, Japan). The elderly
participants were recruited from residents in the city of Hamura, a suburb of Tokyo. The lifestyle of the
residents in this area is typical of that in most regions of Japan. A questionnaire was used to assess
medical history and lifestyle factors (food intake, behavior and physical training history). All of the
participants were considered to be healthy, after excluding women with factors that might affect bone
metabolism, including chronic diseases/conditions affecting vital organs (e.g., heart, lung, liver, kidney,
and brain), women with severe endocrine, metabolic, or nutritional diseases, and women with a known
history of metabolic bone disease or hip fracture, and those using medications known to influence bone
metabolism. This study was approved by the Research Ethics Committee of Tokyo Metropolitan
University and written informed consent was obtained from all participants.
33
2) Measurement of bone mineral density (BMD) and general characteristics
The BMD (g/cm
2) of the whole body, lumbar spine (young; L
1–L
4, elderly; L
2–L
4) and femoral neck were measured by dual-energy X-ray absorptiometry with a Prodigy Advance (GE Healthcare, Piscataway, NJ, USA) for young women, and QDR-A (Hologic Inc., Waltham, MA, USA) for elderly women. For young women, body height (± 0.1cm) and weight (± 0.1kg) were measured using a Tanita body fat analyzer (Model No. BF-220; Tanita Corp., Tokyo, Japan ). For elderly women, body height (± 0.1cm) was measured using a Yagami height meter (Model No. ST-2M; YAGAMI Inc., Nagoya, Japan ) and weight (±
0.1kg) was measured using a Tanita body inner scan (Model No. BC-520; Tanita Corp., Tokyo, Japan).
The BMD of elderly women was recorded as adjusted BMD (adj-BMD) in order to correct for differences in age, height, and weight. The formulas used were as follows: body mass index (BMI) = (body weight, kg) / (body height, m)
2; adjusted BMD (adj-BMD) = BMD - 0.0052432908 × (73.1716102-age) + 0.0088382998 × (23.2271299-BMI) (160).
3) Genotyping of polymorphism in target genes
① DNA extraction and polymerase chain reaction (PCR)
The genotypes of IL6 -634C>G (rs 1800796), TNFα -308G>A (rs 1800629), IL17F 7488T>C (rs 763780),
TGFβ 869T>C (rs 1800470), OPG 163A>G (rs 3102735) and MTHFR 677C>T (rs 1801133) (31, 159,161-165) were analyzed by the polymerase chain reaction–restriction fragment length polymorphism
(PCR-RFLP) method. Genomic DNA was isolated from the fingernail using a DNA isolation kit (Nippon
Gene Co., Ltd, Tokyo, Japan). The concentration of purified DNA was determined using a U-008OD
spectrophotometer (Hitachi Ltd., Tokyo, Japan). The extracted genomic DNA was subjected to PCR to
amplify the IL6, TNFα, IL17F, TGFβ, OPG and MTHFR genes. PCR was carried out in a 50 μl reaction
volume containing 0.5–1 μg of genomic DNA, 1.5 mM or 1 mM magnesium chloride, 0.2 mM
34
deoxy-NTP, 0.4μM of each primer, 0.5 μl of DNA polymerase (Ampli Taq Gold: Applied Biosystems, Carlsbad, CA, USA or Prime Star HS: Takara Biotechnology Co., Ltd, Shiga, Japan) and each PCR buffer (Applied Biosystems or Takara Biotechnology).
IL17F gene and TGFβ gene were amplified adding 4%DMSO or 3% formamide when I were unable to obtain PCR products. Each primer set was synthesized by Sigma-Aldrich Japan (Tokyo, Japan), and PCR was performed on a thermal cycler (model 2720, Applied Biosystems), sequence of the primers and PCR programs for each genes shown in Table 1. The PCR products were separated by electrophoresis on a 2.5% agarose gel and stained with ethidium bromide. Gels were visualized on a transilluminator under ultraviolet light and photographed.
② Restriction fragment length polymorphism (RFLP) method
The primers for IL6, TNFα, IL17F, TGFβ, OPG and MTHFR genes were used to amplify 180-, 107-, 412, 500-, 253 and 198-bp PCR products. The products were then digested with BsrB I, Nco I, NlaⅢ, MSPA1 I, Mfe I and Hinf I (New England Biolabs Inc., Hitchin, UK or Promega Corp., WI, USA), respectively, at 37°C for 3h-16h, followed by electrophoresis using e-Pagel AE6000 (Model E-T15L: Atto Corp., Tokyo, Japan) and visualized by ethidium bromide staining. Cleavage of the PCR product of IL-6 with
BsrB Igenerates two fragments of 120 and 60 bp in length. The 60bp fragments represent (-634)G allele (31).
Digestion of the PCR products of TNFα with NcoI generates two fragments of 87 and 20 bp in length; the (–308)A allele remained uncut with a length of 107 bp (161). Digestion of the PCR products of
IL17Fwith
NlaIII generated two fragments of 288 and 124 bp in length. The 412 bp product remainedundigested for the (7488)C allele (162, 163).
TGFβ gene amplification products digested with MspA1Iproduces four constant fragments of 67, 67, 41 and 40bp. The (869)C allele is defined by the presence of polymorphic bands of 273bp and 12bp, and the (869)T allele is defined by a 285 bp fragment (164).
Digestion of the
OPG PCR products by MefI generated two fragments of 232 and 21 bp in length. The35
(163)A allele remained uncut with a length of 253 bp. The (163)G allele was defined by the presence of 232 and 21 bp fragments (165). The amplified fragments of MTHFR were digested with
hinfI into 175and 23bp fragments. The (677)C allele remained undigested, and the 175bp fragment derived from the (677)T allele (159).
4) Statistical analyses
① Genomic analyses
Quantitative associations between genotypes and BMD values (g/cm
2) were analyzed by ANOVA with regression analysis as a post hoc test. Student’s
t-test and the Mann-Whitney’s U test were utilized toexamine the effects on each genotype on BMD for young women, on adj-BMD for the elderly and all women. All tests regarded
P-values<0.05 as significant. χ2tests were used to determine the Hardy–Weinberg equilibrium for each genotype, and P<0.05 was considered statistically significant.
② Anthropometry
Regarding the relationship between the BMD of total body, lumbar spine (young; L
1–L
4, elderly; L
2–L
4and femoral neck and BMIs were assessed by linear regression analysis. A correlation coefficient of r >
0.5 was considered significant.
36
Table 1 Sequence of the forward and reverse primers used to generate each PCR product and PCR programs for each gene Gene Position
substitution
rs ID Primer and restriction condition (Amplitaq Gold/ Prime Star)
IL6–634C>G rs1800796 F: 5’-GAGACGCCTTGAAGTAACTG-3’
R: 5’-AACCAAAGATGTTCTGAACTGA-3’
35 cycles of 94°C for 30 s, 55°C for 30 s, and 72°C 30 s; and 35 cycles of 98°C for 10 s, 55°C for 15 s, and 72°C for 11 s
TNFα–308G>A rs1800629 F: 5’-AGGCAATAGGTTTTGAGGGCCAT-3’
R: 5’-TCCTCCCTGCTCCGATTCCG-3’
35 cycles of 94°C for 30 s, 60°C for 30 s, and 72°C for 30 s; and 35 cycles of 98°C for 10 s, 60°C for 15 s, and 72°C for 10 s
IL17F7488T>C
His 161 Arg
rs763780 F: 5’-GTTCCCATCCAGCAAGAGAC-3’
R: 5’-AGCTGGGAATGCAAACAAAC-3’
40 cycles of 94°C for 30 s, 60°C for 30 s, and 72°C for 30 s; and 35 cycles of 98°C for 10 s, 60°C for 15 s, and 72°C for 14 s
TGFβ869T>C
Leu 10 Pro
rs1800470
F: 5’-TTCAAGACCACCCACCTTCT-3’R: 5’-TCGCGGGTGCTGTTGTACA-3’
35 cycles of 94°C for 30 s, 60°C for 30 s, and 72°C for 30 s; and 35 cycles of 98°C for 10 s, 60°C for 15 s, and 72°C for 30 s
OPG163A>G rs3102735
F: 5’-AGTCTAACTTCTAGACCAGGCAATT-3’R: 5’-AGTTAGAGCCAGAGAGAATCTG-3’
35 cycles of 94°C for 40 s, 53°C for 40 s, and 72°C for 40 s; and 35 cycles of 98°C for 10 s, 53°C for 15 s, and 72°C for 15 s
MTHFR677C>T
Ala 222 Val
rs1801133
F: 5’-TGAAGGAGAAGGTGTCTGCGGGA-3’R: 5’-AGGACGGTGCGGTGAGAGTG-3’
35 cycles of 95°C for 60 s, 62°C for 90 s, and 72°C for 60 s; and 35 cycles of 98°C for 10 s, 62°C for 15 s, and 72°C for 12 s
37 3. Results