protein tyrosine phosphataseβ and its ligand pleiotrophin in the submandibular gland of mice
著者 アタパンヤワニット カンニカ
著者別表示 Adthapanyawanich Kannika journal or
publication title
博士論文本文Full 学位授与番号 13301甲第3927号
学位名 博士(医学)
学位授与年月日 2013‑09‑26
URL http://hdl.handle.net/2297/39508
doi: 10.1016/j.archoralbio.2012.09.005
Creative Commons : 表示 ‑ 非営利 ‑ 改変禁止 http://creativecommons.org/licenses/by‑nc‑nd/3.0/deed.ja
Expression and localization of receptor protein tyrosine phosphatase b and its ligand pleiotrophin in the
submandibular gland of mice
Kannika Adthapanyawanich, Miyuki Yamamoto, Tomohiko Wakayama, Hiroki Nakata, Sunisa Keattikunpairoj, Shoichi Iseki *
DepartmentofHistologyandEmbryology,GraduateSchoolofMedicalScience,KanazawaUniversity,Kanazawa,Ishikawa920-8640,Japan
1. Introduction
Thesubmandibulargland(SMG)ofrodentsiscomposedofthe acinus and the duct system, the latter composed of the intercalatedduct(ID),striatedduct(SD),granularconvoluted tubule(GCT),andexcretoryduct.1Extensivedevelopmentof
the GCT from the SD takes place postnatally around the puberty in an androgen-dependent manner, resulting in a markedsexualdimorphisminthemorphologyandfunctionof theductsystem,withtheGCTdevelopedpreferentiallyinthe male gland.2–4 The epithelial cells of GCT have abundant secretorygranulesthatcontainavarietyofbiologicallyactive peptides suchas nerve growthfactor (NGF) and epidermal article info
Articlehistory:
Accepted20September2012
Keywords:
RPTPb Pleiotrophin Salivarygland Ductsystem Sexualdimorphism Androgens Mouse
abstract
Objectives: Thefamilyofreceptorproteintyrosinephosphataseb(RPTPb)iscomposedof4 splicevariantsandthoughttoplayrolesintheneuralmigrationandoutgrowth.Several ligands including the growth factor pleiotrophin (PTN) bind to RPTPb and inhibit its phosphataseactivity,therebyactivatingcellularsignalling pathways.Weexaminedthe expressionandlocalizationofRPTPbanditsligandsinthesubmandibulargland(SMG)of mice,whichisknownforaprominentsexualdimorphismintheductsystem.
Design: ThehomogenatesandtissuesectionsofmaleandfemalemouseSMGwereanalysed withRT-PCR,Westernblotting,andimmunohistochemistry.
Results: TheshortreceptortypeofRPTPb(RPTPb-S)wasdominantlyexpressedintheSMG, andthemaleglandhadsignificantlyhigherlevelsofRPTPb-Sexpressionthanthefemale gland.Inthemale,RPTPb-Swaslocalizedpredominantlyinintercalatedduct(ID)cells,but wasnot foundingranularconvoluted tubule(GCT)cellsoracinar cells.Inthefemale, weakerreactivitywasdemonstratedinbothIDandstriatedduct(SD)cells.Oftheknown ligandsforRPTPb,PTNwasexpressedintheSMG,withoutsexualdifferenceinlevels.Inthe male,PTNwaslocalizedinIDcellsaswellasincellslocatedinthedistalendsofGCTthatare in close vicinity tothe ID, whereasin the female PTNwas colocalized with RPTPb-S throughoutIDandSDcells.
Conclusions: TheseresultsindicatedthatthedistributionofRPTPb-SanditsligandPTNhasa closerelationtothesexualdimorphismintheductsystemofmouseSMG.
#2012ElsevierLtd.Allrightsreserved.
*Correspondingauthor.Tel.:+81762652150;fax:+81762344220.
E-mailaddress:[email protected](S.Iseki).
Abbreviations: SMG,submandibulargland;SLG,sublingualgland;ID,intercalatedduct;SD,striatedduct;GCT,granularconvoluted tubule;RPTPb,receptorproteintyrosinephosphataseb;PTN,pleiotrophin;NGF,nervegrowthfactor;IHC,immunohistochemistry.
Availableonlineatwww.sciencedirect.com
journalhomepage:http://www.elsevier.com/locate/aob
0003–9969/$–seefrontmatter#2012ElsevierLtd.Allrightsreserved.
http://dx.doi.org/10.1016/j.archoralbio.2012.09.005
growthfactor(EGF).5Castrationofmalescausesinvolutionof theGCTaccompaniedbyconversionofthephenotypeofGCT cells into that of SD cells, whereas administration of androgenstofemalesorcastratedmalescausestheopposite phenomenon.6,7 Suchandrogen-dependentGCTdifferentia- tionisaccompaniedbyupregulationofthegeneexpressionfor GCT-specificproducts,8butthemolecularmechanismunder- lyingthisdifferentiationislargelyunknown.
Thereceptorproteintyrosinephosphataseb(RPTPb),also knownasproteintyrosinephosphatasez(PTPz),wasoriginally foundinthecentralnervoussystem.9,10RPTPbformsafamily consisting of 4 splice variants, i.e., the long and short transmembrane receptor proteins(RPTPb-L and -S, respec- tively),andthelongandshortsecretoryproteins(phosphacan and phosphacan short isoform, respectively).11–13 In the receptor types of RPTPb, the extracellular proteoglycan domains bind to multiple ligands, and the intracellular tyrosine-phosphatase domains de-phosphorylate various substrates,resulting in theregulation ofcellular signalling pathways.10 Themembers of theRPTPb familyare mainly expressedin neurons and gliaofthe embryonicand adult central nervous system and thoughtto be involved inthe neural differentiation and migration, neurite outgrowth, synaptic formationandregulation.10,14–16Severalmolecules including the growth factor pleiotrophin (PTN) have been knownasligandsforRPTPb.17–20Uponbindingoftheligands, RPTPbformshomodimersandloosesitstyrosinephosphatase activity,resultinginactivationbyphosphorylationofcellular signallingmolecules.21,22
Inapreliminarystudyanalysingtranscriptsfromthemale and femalemouse SMGwith aDNA microarray,we found RPTPb to be expressedmore abundantly inthe male than femalegland(datanotshown).Therefore,inthepresentstudy weexaminedtheexpressionandlocalizationofthemembers oftheRPTPbfamilyanditsligandsinthemouseSMG,with specialreferencetoitssexualdimorphism.
2. Materials and methods
2.1. AnimalsandorgansMaleandfemaleSlc:ddYmiceattheageof8weeks(W),some ofwhichunderwenttestectomyat6W,werepurchasedfrom NipponSLC(Hamamatsu,Japan),rearedunderstandard12h light/12h dark laboratory conditions with free access to standard food and water, and used at the ages of 8–10W (adults).Miceattheagesof2Wand5Wwerealsoused.All subsequentexperimentswereconductedinaccordancewith theGuidelinesfortheCareandUseofLaboratoryAnimalsat KanazawaUniversity.Somefemaleandtestectomizedmale adultsweresubjectedtosubcutaneousinjectionsof25mg/kg testosterone(WakoPureChemicalIndustries,Osaka,Japan) dissolvedin0.1mlofoliveoilevery24h,andusedeitherat6h afterasingleinjectionorat6hafter5consecutiveinjections.
Groupsof6–9maleandfemaleanimalsinvariousagesand experimentalconditionsweresacrificedunderpentobarbital anaesthesia by transcardial perfusion with physiological saline. For RT-PCR and Western blot analyses, the SMG, sublingual gland (SLG), parotid gland, extraorbital lacrimal
gland,andcerebrum(subsequentlyreferredtoasbrain)were dissected out, frozen immediately in liquid nitrogen, and stored at 808C until use.For immunohistochemical(IHC) analysis, the animals were fixed by perfusion with 4%
paraformaldehyde in 0.1M phosphate buffer, pH 7.2, the SMGwasdissectedout,andfurtherfixedbyimmersioninthe same fixative overnight at 48C. Thespecimens were then either rinsed overnight at 48C with 30% sucrose in 0.1M phosphatebuffer,frozen,andcutinto8mmsectionsusinga cryostat,ordehydratedinagradedethanolseries,embedded inparaffin,andcutinto4mmsectionsusingmicrotome.The cryostat and paraffin sections were mounted on silanized glassslides(DAKO,Glostrup,Denmark).
2.2. RNApreparationandconventionalRT-PCR
TotalRNAwasisolatedfromthefrozenspecimenswiththe guanidinine-phenol-chloroformmethodusingacommercial solution(TRIreagent;Sigma–AldrichCo.,St.Louis,MO).The first-strandcDNAwas synthesizedfrom2mg-aliquotofthe totalRNAsamplesusingtheoligo(dT)20primerandMoloney murineleukaemiavirusreversetranscriptase(Toyobo,Osaka, Japan).FromtheseRTproducts,cDNAfragmentsforRPTPb family members and other proteins were amplified with conventional and quantitative PCR. The sequences of the primerpairsusedinthepresentstudyarelistedinTable1.The primerpairsfortheRPTPbfamilyareeitherthosecommonto all the members or those capable of distinguishing each member specifically. The sequences of the specific primer pairs weredesigned according tothegenomic sequence of RPTPb (NCBI Reference, NC_000072) using the SpliceMiner tool,takingadvantageofthatthefamilymembersarederived fromdifferentialsplicingintheExon12.TheconventionalRT- PCR was first performed for 30 cycles using TaqDNA polymerase (ExTaq; TakaraBiomedicals, Kusatsu, Japan)in aDNAthermalcycler(MJResearch,Watertown,MA),andthe amplified products were analysed with an agarose-gel electrophoresis.
2.3. Real-timequantitativeRT-PCR
The real-time quantitative RT-PCR was performed in a StratageneMx-3005PThermocycler(Stratagene,LaJolla,CA) accordingtotheproceduresrecommendedbythemanufac- turer.LightCyclerDNAMasterSYBRGreenI(RocheDiagnos- tics,Indianapolis,IN)wasusedtodetecttheamplificationof cDNAinatotalvolumeof20mlwiththeabsolutequantitative, DCtmethod.23Eachreactionconsistedof10mlofSYBRgreenI, 1mlofthecDNAsample,0.5mlofeachprimerpair(10pmol/
ml), and 8ml ofdistilled water. Thermal cycling conditions were10minat958Cfollowedby45cyclesat958Cfor40s, 608Cfor30s,and728Cfor30s.GAPDHwasemployedasthe endogenouscontroltonormalizethedata.Eachsamplewas analysedintriplicates,andsamplesfrom3differentanimals wereanalysedtodetermineeachvalue.
2.4. Preparationofprimaryantibodies
Rabbit polyclonalanti-PTN antibodyand anti-NGFantibody werepurchasedfromSantaCruzBiotechnology(SantaCruz,
CA), and mouse monoclonal anti-a-tubulin antibody was purchasedfromSigma(St.Louis,MO).Ratpolyclonalantisera againstthe cytoplasmicdomainofRPTPb-S(common with thatofRPTPb-L)wereproducedinourlaboratoryaccordingto themethoddescribedpreviously.24,25Briefly,arecombinant oligopeptide41aminoacidsinlengthcorrespondingtotheC- terminusofRPTPb-S,thatwasfusedwiththecarrierprotein glutathione-s-transferase, was produced in bacteria BL21 (Novagen,Madison,WI)byintroducingtheexpressionvector pGEX-6p-1(AmershamPharmaciaBiotech,Uppsala,Sweden).
TheoligopeptidewasthenemulsifiedwithFreund’scomplete adjuvantandinjectedasantigenintofootpadsofadultfemale Wistarrats.Aboosterimmunizationwasmade2weekslater, andtheantiserawerecollected1weekafterthebooster.
2.5. Westernblotting
Frozen SMG tissues were homogenized in a lysis buffer containing50mMTris–HCl(pH7.5),150mMNaCl,1%NP-40, 0.5% sodium deoxycholate, 0.1%SDS, protease inhibitor cocktail (CompleteTM; RocheDiagnostics) and phosphatase inhibitorcocktail(PhosStopTM;RocheDiagnostics).Thetissue homogenates were then separated on 12% SDS-polyacryl- amide gels and transferred to PVDF membranes (BioRad Laboratories,Hercules,CA).Afterbeingblockedwith5%non- fatskimmedmilkinPBS,themembraneswereincubatedwith rat anti-RPTPb-S antisera (1:500 dilution), rabbit anti-PTN antibody(1:2000),ormouseanti-a-tubulinantibody(1:10,000 dilution) overnight at 48C. After being washed, the mem- braneswere incubated with horseradishperoxidase-conju- gatedsecondary antibody againstrat,rabbit, or mouseIgG
(1:2000)(Dako)for1h.Theimmunoreactionwasdetectedand its intensity was quantified in ImageQuant LAS-4000 mini (FujifilmMedical,Tokyo,Japan)aftertreatmentofthe blots with the chemiluminescent reagent ECL-plus (Amersham PharmaciaBiotech).Samplesfrom3differentanimalswere analysedtodetermineeachvalue,andtherelativeintensityof RPTPb-S bands against the corresponding a-tubulin bands werecalculated.
2.6. IHC
Paraffin sections of SMG, 5mm thick, were made with a microtomeandmountedonsilanizedglassslides(Dako).After being deparaffinized in xylen and rehydrated in a graded ethanolseries,thesectionswerepre-treatedwith5%normal swine sera for 30min. For IHC by the enzyme-detection method,thesectionswereincubatedovernightat48Cwithrat anti-RPTPb-Santisera (1:200 dilution),rabbit anti-PTN anti- body (1:1000 dilution), or rabbit anti-NGF antibody (1:400 dilution).Toconfirmspecificityoftheimmunoreaction,the primary antibodieswere replacedwith non-immunerator rabbitserawiththecorrespondingdilutions.Afterawashwith PBS,thesitesofimmunoreactionwerevisualizedbyincubat- ingthesectionssuccessivelywithbiotinylatedanti-ratoranti- rabbitIgGantibody(VectorLaboratories,Burlingame,CA)at 1:200for1h,horseradishperoxidase-conjugatedstreptavidin (Dako) at 1:300 for 1h, and a peroxidase substrate (ImmPACTTMDAB,VectorLaboratories)forabout5min.The sections were subjected toobservation under an Olympus BX50 microscope. Thepre-embedding immunoelectronmi- croscopy was performed as described previously.25 The Table1–SpecificprimersusedforRT-PCRanalysis.
Gene GenBank Fractionlength(bp) Primerdirection Sequence
RPTPba NM_001081306 150 Forward 50-ACCAGCCTTCTGGTCACATG-30
Reverse 50-GAGAATGGCACCCAAGTCCT-30
RPTPb-L BC151071 153 Forward 50-ACCATCTGTTACTAGTGGGC-30
Reverse 50-AAAGTCAGGGCAGACACGAT-30
RPTPb-S BC157965 155 Forward 50-ACCGAAGTGACACCACAGGC-30
Reverse 50-TCCAACCCCTCAGCTAGACC-30
Phosphacanb AJ133130 154 Forward 50-GGTCCTCCACACCATCTGTT-30
Reverse 50-GACCGGAATTCCTTTCTTCC-30
GAPDH NM_008084 579 Forward 50-AAGGGCTCATGACCACAGTC-30
Reverse 50-AGGGAGATGCTCAGTGTTGG-30
151 Forward 50-CATGGCCTTCCGTGTTCCTA-30
Reverse 50-CTGGTCCTCAGTGTAGCCCAA-30
PTNc NM_008973 470 Forward 50-ATGTCGTCCCAGCAATATCA-30
Reverse 50-TTCTTCTTCTTTGACTCCGC-30
Midkine NM_010784 153 Forward 50-ACCCACCAGTGCCTTTTGTC-30
Reverse 50-TAACAAGTATCAGGGTGGGG-30
Tenastin NM_011607 530 Forward 50-TGCAACGAGCCCCTTTGCCT-30
Reverse 50-ATGCACTTGCCCTGTACGCA-30
Contactin NM_001159647 501 Forward 50-CATTGCTGGTCAGCCATCTC-30
Reverse 50-GGTAGCTAAGATCATCTGGG-30
N-CAM NM_001081445 350 Forward 50-ACCGGAAATCAGGCTCCCAT-30
Reverse 50-CCTCCATGTCTTTGCCCTTG-30
NFPd NM_010904 151 Forward 50-TGCCCTCACCAAACAGGAAT-30
Reverse 50-GCGTAGCGTTCAGCATACAT-30
a RPTP-b:theprimerpairiscommontoall4typesoftheRPTP-bfamily.
b Phosphacan:theprimerpairiscommontophosphacanandphosphacanshortisoform.
c PTN:pleiotrophin.
d NFP:theheavychainofneurophilamentprotein.
cryostatsectionsofSMGimmunostained withanti-RPTPb-S antisera or anti-PTN antibody were fixed with 1% OsO4, stainedwith1%uranylacetateandembeddedinGlicidether (SelvaFeinbiochemica,Heidelberg,Germany).Ultrathinsec- tionswerepreparedusinganultramicrotomeandsubjectedto observationwithaJEM-1210electronmicroscope(JEOL,Tokyo, Japan). For fluorescent double-immunostaining, the pre- treatedsectionswereincubatedwithratanti-RPTPb-Santi- sera (1:200) mixed with rabbit anti-PTN antibody (1:500) overnight at 48C. After being washed, the sections were incubatedwithamixtureofanti-ratIgGantibodyconjugated withAlexaFluor594andanti-rabbitIgGantibodyconjugated withAlexaFluor488(MolecularProbes,Eugene,OR)for1h.
They were then mounted in glycerol and subjected to observationwith anOlympusBX50/BX-FLA fluorescent mi- croscope.TheIHCresultswereconfirmedintheSMGfrom3 differentanimals.
2.7. Statisticalanalysis
Thestatisticaldifferencewas analysedbetweentwovalues withStudent’st-test,andamongmultiple valueswithone- factoranalysisofvariance(ANOVA)followedbyBonferroni’s posthoctest.ThedifferencewithaPvaluelessthan0.05was consideredsignificant.
3. Results
3.1. ExpressionoftheRPTPbfamilyinsalivaryand lacrimalglands
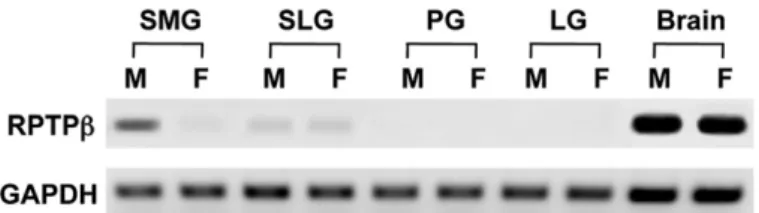
IntheRT-PCRanalysis(30cycles)usingtheprimerscommon toall4typesofRPTPb,expressionofthemRNAforRPTPbwas detectedabundantlyintheSMGaswellasinthebrain,and slightlyinSLG,butwasnotdetectedintheparotidorlacrimal gland. In the SMG, the expression appeared substantially strongerinthemaleglandthanthefemalegland(Fig.1).Such sexualdifferencewasnotrecognizedinthebrainorSLG.
3.2. ExpressionofdifferenttypesofRPTPbinthemaleand femaleSMG
Ofthe4typesofRPTPb,onlytheshortreceptortype(RPTPb-S) was abundantly expressed in the SMG with the RT-PCR analysis(30 cycles)using thespecificprimer pairs, butthe
transcriptsforlongreceptortype(RPTPb-L)orlongandshort secretorytypes(phosphacanandphosphacanshortisoform) were not or scarcely detected (Fig. 2A). The expression of RPTPb-SintheSMGappeared muchmoreabundantinthe malethanfemale.Theseresultswerefurtherconfirmedinthe real-timequantitative RT-PCR(Fig. 2B).Whencomparedfor the copy numbers normalized with those of GAPDH, the abundanceofRPTPb-SmRNAinthemaleSMGwas45%thatin Fig.1–RT-PCRanalysisfortheexpressionofRPTPbinthesubmandibulargland(SMG),sublingualgland(SLG),parotidgland (PG),extraorbitarylacrimalgland(LG)andcerebrum(Brain)ofmale(M)andfemale(F)adultmice.Theproductsof
amplificationusingtheprimerpaircommontoall4typesoftheRPTPbfamilywereelectrophoresedandstainedwith ethidiumbromide.ExpressionofGAPDHisshownaspositivecontrol.
Fig.2–(A)ConventionalRT-PCRanalysisfortheexpression ofRPTPb-L,RPTPb-S,andphosphacanintheSMG,SLG, andbrainofmale(M)andfemale(F)adultmice.The productsofamplificationusingprimerpairsspecificto eachtypeofRPTPbwereelectrophoresedandstainedwith ethidiumbromide.Theprimerpairforphosphacanis commontophosphacanandphosphacanshortisoform.
ExpressionofGAPDHisshownaspositivecontrol.(B) Real-timequantitativeRT-PCRanalysisfortheexpression ofRPTPb-SinthemaleandfemaleSMGandmalebrain.
ThelevelsofmRNAforeachtypeofRPTPbwere
normalizedbydividingwiththoseforGAPDH.Thevalue inthebrainwassetas1.0andtherelativevaluesinthe maleandfemaleSMGareindicated.Eachvaluerepresents meanWSDof3animals.*Significantlydifferent(P<0.05).
thebrainand2.2-foldthatinthefemaleSMG(significantly different,P<0.05).
3.3. AndrogendependencyoftheexpressionofRPTPb-Sin theSMG
ThelevelsofexpressionofRPTPb-SintheSMGhadnosexual differenceat2weekspostpartum,butwereelevatedcontinu- ouslyinthemaleat5and8weeks,coincidentwithknown postnataldevelopmentoftheGCTinthemalegland(Fig.3).
Upontestectomy, the levels of RPTPb-S in the male gland decreased to those in the normal female gland. Single administration of testosteroneto testectomized males had noeffectafter6h,butrepeatedadministrationoftestosterone for 5 days restored the levels of the normal male gland.
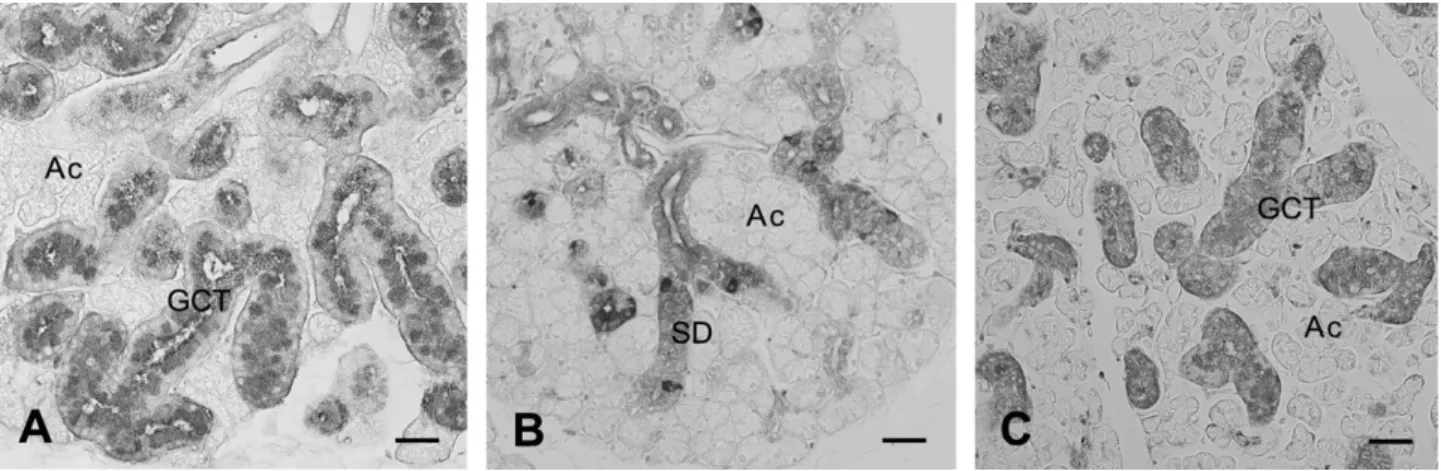
Administrationoftestosteronetofemalescausedthesame effectasintestectomizedmales(datanotshown).Androgen- dependentdifferentiationoftheductsystem,asrepresented bythepreferentialdevelopmentofGCTcellsinthemalegland andtestosterone-treatedfemaleortestectomizedmalegland, wasverifiedwithimmunostainingforNGF,aspecificmarker ofGCTcells(Fig.4).
3.4. ExpressionofRPTPb-SproteinintheSMG
WithWesternblottingusingthespecificratantiseraagainst the cytoplasmic domaincommonto the receptortypesof RPTPb,thebandof220kDacorrespondingtoRPTPb-S was detected in the SMG, with1.8-fold higherintensity in the malethanfemale(significantlydifferent,P<0.05)(Fig.5).In contrast,thebandof380kDacorrespondingtoRPTPb-Lwas detectedinthebrainbutnotintheSMG.Thereasonwhythe band for RPTPb-S was scarcely detected in the brain is unclear,butthebrain(cerebrum)mayindeedproducemuch smaller amount of RPTPb-S than RPTPb-L at the protein level.
3.5. ImmunohistochemicallocalizationofRPTPb-Sprotein intheSMG
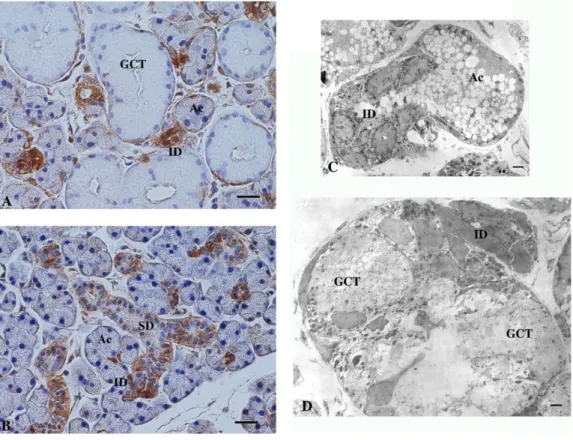
ThemaleandfemaleSMGwereexaminedwithIHCforRPTPb- S.Inthemalegland,intenseimmunoreactivitywaslocalized predominantlyinID,whereastherestoftheintralobularduct portions,composedmostlyofGCT,werecompletelyimmu- nonegative(Fig.6AandB).Largeextralobularducts(excretory ducts)wereweaklyimmunostained.Incontrast,inthefemale gland, where GCT cells are scarce, immunoreactivity of moderateintensitywasdistributedthroughouttheintralob- ularductportionscomposedofIDandSD.Theacinishowed no immunostaining in either sex. IHC with the primary antibody replaced by non-immune rat sera showed no immunostaining(datanotshown).Withelectronmicroscopic IHC,intense immuoreactivityinthemale glandwas recog- nizedthroughoutthemembraneandcytoplasmicportionsof IDcells,thusitwasdifficulttodetermineifthereactivitywas strongerintheluminalorbasalportionsofthecellmembrane (Fig.6CandD).ThepatternsofthedistributionofRPTPb-Sin the SMG of testectomized mice and testosterone-treated female ortestectomizedmicewereessentially thesameas in those of female and male mice, respectively (data not shown).
Fig.3–RT-PCRanalysisfortheeffectsofsex,postnatal daysandhormonalconditionsontheexpressionof RPTPb-SintheSMG.TheproductsofamplificationinSMG frommale(M)andfemale(F)miceat2W,5Wand8W (adult)postpartum,andfromtestectomizedadultmale mice(Tx),Txat6hand5daysaftersingleandrepeated administrationsoftestosterone(T),respectively,were electrophoresedandstainedwithethydiumbromide.
Fig.4–ThesexualdimorphismandtheeffectoftestosteroneintheductsystemofmouseSMG.SMGfromadultmale(A), female(B)andfemalemicetreatedwithtestosteronefor5consecutivedays(C)wereimmunostainedwithanti-NGF antibody.(A)and(C)TheductsystemislargelyoccupiedwithGCTcells(GCT)thathaveabundantNGF-positivesecretory granules.(B)OnlyasmallnumberofNGF-positivecellsarescatteredamongSDcells(SD)thathavelowornoNGF- immunoreactivity.TheresultsintheSMGoftestectomizedmiceandtestosterone-treatedtestectomizedmiceare essentiallythesameasBandC,respectively(datanotshown).Ac,acinus.Bar=40mm.
3.6. ExpressionoftheligandsofRPTPbintheSMG
Of the known ligands for the RPTPb family tested (PTN, midkine, tenastin, contactin and N-CAM), onlythe growth factorPTNwasabundantlyexpressedintheSMG,asdetected withRT-PCT(30cycles),withoutsexualdifferenceinlevels (Fig.7).NomRNAforneurofilamentprotein,aspecificmarker of neurons, was detected in the SMG with the same amplificationcondition.Thisresult,togetherwiththeabsence ofneuronalganglioncellsinthehistologicalsectionsofSMG, suggeststhatthecontributionofneuron-derivedtranscripts in SMG is negligible in the present study. With Western blotting using thespecificrabbit antibodyagainstPTN,the specific band ofapproximately 15kDa was detected inthe SMG,withoutsexualdifferenceinintensity(Fig.8).
3.7. ImmunohistochemicallocalizationofPTNproteinin theSMG
WithIHC,thedistributionofPTNinthemaleandfemaleSMG wasalsorestrictedtotheductsystem(Fig.9AandB).Inthe malegland,theimmunoreactivitywasprimarilypresentinID cellsbutwasalsofoundincellslocatedinthedistalendsof GCTthatareadjacenttoID(Inthispaper,‘‘distal’’referstoa partoftheductsystemthatisclosertotheacinus).Incontrast, in the female gland the immunoreactivity was present
Fig.6–ImmunohistochemicallocalizationofRPTPb-S.Paraffinsectionsofthemale(A)andfemale(B)SMGwere immunostainedwithanti-RPTPb-Santiseraandvisualizedwiththeenzyme-histochemicalmethod.Ac:acinus;ID:
intercalatedduct;SD:striatedduct;GCT:granularconvolutedtubule.Bar=20mm.(C)and(D)Acryostatsectionofthemale SMGwasimmunostainedwithanti-RPTPb-Santiseraandvisualizedwithelectronmicroscopywiththepre-embedding, enzyme-histochemicalmethod.NotethedarkimmunoreactionproductspresentintheentireportionsofIDcells(ID)but absentinacinarcells(Ac)orGCTcells(GCT).Bars=2mm.
Fig.5–WesternblotanalysisfortheexpressionofRPTPb-S inthemale(M)andfemale(F)SMGandmalebrain.The proteinsampleswereelectrophoresed,blottedand immunostainedwithratanti-RPTPb-Santisera.This antibodyrecognizesbothRPTPb-Sand-L.Theexpression ofa-tubulinisshownasloadingcontrol.Themolecular weights(kDa)oftheimmunoreactivebandsareindicated.
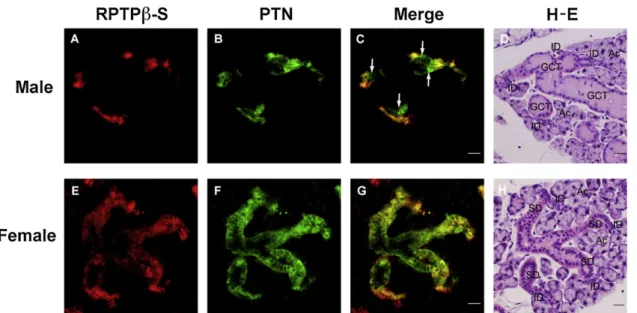
diffuselyinmostIDandSDcells.Withelectronmicroscopic IHC,theintenseimmuoreactivityforPTNinthefemalegland wasrecognizedinbothmembraneandcytoplasmicportions ofSDcells,thusitwasdifficulttodetermineifthereactivity wasstrongerintheluminalorbasalportionsofthecytoplasm (Fig.9C). Double-immunostainingconfirmedthatPTN-posi- tivecellsmostlyoverlapwithRPTPb-S-positivecellsintheID ofbothsexesandSDofthefemale,althoughPTNappeared weakerintheIDthanSD,andRPTPb-Sappearedweakerinthe proximalportionsthanthedistalportionsofSD(Fig.10).In
addition, cells in the distal ends ofGCT ofthe male were positiveforPTNbutnegativeforRPTPb-S.
4. Discussion
Thecellularkineticsresponsibleforformationandmainte- nanceofthestructureofrodentSMGarenotfullyunderstood.
Inthefirstfewpostnatalweeks,cellsconstitutingthedistal ends of intralobular ducts (terminal tubules) give rise to acinar cellsandundergoextensiveproliferation,aprocess promotedbyb-adrenergicmechanisms.26–28Thereafterthe ductsystemcontinuestodeveloptoformdifferentcompart- ments,ofwhichtheGCTisdifferentiatedfromtheSDafter puberty dependenton the action of hormones, mainlyof androgens.4,6,7Intheadulthood,theacinarcellsaswellas cellsofthe differentcompartmentsoftheduct systemall retainproliferativepotentialofvaryingdegrees.Forexample, singleadministrationofb-adrenergicagonistslikeisoproter- enolcausesaremarkableincreaseintheproliferationrateof entireacinarcells.29,30Nevertheless,thereisevidencethat theIDcompartmentofadultrodentSMGcontainsstemcells, whichproliferateandeventuallyreplenishallparenchymal celltypesoftheSMG.31–34Kineticstudiesusingradiolabeled DNAprecursorshaverevealedthatIDcellsdifferentiateinto bothacinarandGCTcellsthatarelocatedattheoppositeends oftheID.31,33,35Thus,GCTcellsinadultSMGarisefromself- proliferation and also from differentiation of ID cells.
AlthoughinthefemaleglandGCTcellsaremostlyreplaced bySDcells,cellslocatedattheendsoftheSDandadjacentto theIDoftenhavesecretorygranulesandareconsideredto representtheintermediatebetweenSDandGCTcells.3,35,36 Furthermore, administration of androgens into females causes extensive conversion ofSD cells into GCT cells.6,7 Thesephenomenaleadustoaconceptthat thereisacell lineageintheductsystemoriginatingfromstemcellsinthe ID compartment and terminating in GCT cells through thestageofSDcells.32,37Higherlevelsofplasmaandrogens inthemalemaypromoteprogressionofthiscelllineageatthe stepfromSDtoGCTcells.
Inthepresentstudy,werevealedthatRPTPb-S,theshort receptortypeofRPTPbwiththemolecularweight220kDa,is abundantlyexpressedintheSMGofmice,athigherlevelsin themalethanfemale.Thisdifferencewasabout2.2-foldat themRNAleveland1.8-foldattheproteinlevel.IHCrevealed thatRPTPb-Sisprimarilylocalizedinthe IDinbothsexes, contrarytotheexpectationthatitmightbelocalizedinthe GCTthatismuchmoredevelopedinthemale.Furthermore, whereasinthefemaleRPTPb-SwasalsoexpressedintheSD, inthemaleitwascompletelynegativeintheGCT.Takinginto accountthemale-dominanceofthetotallevelsofRPTPb-S expression, the levels of RPTPb-S expression in the ID is considered to be much higher in the male than female.
Testosterone-inducedconversionofSDintoGCTcellsinthe SMGoffemalesortestectomizedmaleswasaccompaniedby anincreasedexpressionofRPTPb-S,presumablyinIDcells.
ThecontinuouspresenceofRPTPb-SintheIDthroughtoSDin thefemale,incontrasttothecleargapbetweenthepresence inthe IDandabsenceinthe GCTofRPTPb-Sinthe male, suggeststhatexpressionofRPTPb-Shasacloserelationto Fig.7–RT-PCRanalysisfortheexpressionofvarious
ligandsforRPTPb-SintheSMG,SLGandbrain.The productsamplifiedwiththeprimerpairsforpleiotrophin (PTN),midkine,tenastin,contactinandN-CAMwere electrophoresedandstainedwithethidiumbromide.
Expressionoftheheavychainofneurofilamentprotein (NFP)isalsoshownasaspecificmarkerofneurons.
Fig.8–WesternblotanalysisfortheexpressionofPTNin themale(M)andfemale(F)SMGandmalebrain.The proteinsampleswereelectrophoresed,blottedand immunostainedwithratanti-PTNantibody.The
expressionofa-tubulinisshownasloadingcontrol.The molecularweights(kDa)oftheimmunoreactivebandsare indicated.
androgen-dependentdifferentiationoftheductcomponents leadingtothesexualdimorphism.
ThefamilyofRPTPbareexpressedprimarilyinthecentral nervoussystemincludingtheretinafromtheearlyembryonic ages through to adulthood, in both neurons and glia, suggestingpleiotrophicrolesofRPTPbintheneuraldifferen- tiationandmigration,neuriteoutgrowth,synapticformation andregulation.10,14–16AlthoughmicedeficientforRPTPbwere initiallydescribedtodisplaynoobviousabnormalityinthe nervous system,38 subsequent studies reported defects in myelination of oligodendrocytes39 and abnormal motor coordination and nociception.40In addition tothe nervous system,RPTPbisknowntobeexpressedinotherorgansand tissues,such as boneand stomach,and mice deficientfor RPTPb arereportedtodisplay abnormal boneremodeling41 andresistancetogastriculcerinductionbyHelicobacterpylori infection.42 The biological significance of the presence of differenttypesofRPTPbislargelyunknown.Inthedevelop- ment ofthe central nervous system, including that ofthe retina, all types of RPTPb seem to have similar biological roles.43Interestingly,similartothepresentcase,osteoblasts specificallyexpresstheshortreceptortypeofRPTPb(RPTPb-S), thedeficiencyofwhichcausesabnormalityinboneremodel- linginmice.41
SeveralligandsareknowntobindtoRPTPb,includingthe extracellular matrix protein tenastin, the cell adhesion moleculescontactinandN-CAM,andthegrowthfactorPTN and midkine.17–20PTN isa18kDasecretedheparin-binding cytokineoriginallyfoundintheuterusandbrain.44–46PTNis expressedprimarilyintheneuronsandgliaintheembryonic andadultcentralnervoussystem,47–49inwhichitisthoughtto signaldiversefunctions,includingneuralstemcelldifferenti- ation,glialcelldifferentiatiation,neuronalmigration,neurite outgrowth,andangiogenesis.45,46,50,51PTNisalsoexpressedin avarietyofdevelopingnon-neuraltissues47andmitogenicfor fibroblasts, endothelial,epithelial, and tumour cells.50 Mice deficient for PTN display thickning of cerebral cortex and behavioural abnormality.52,53 PTN was the first soluble moleculetobeidentifiedasligandforRPTPb.19Uponbinding tothereceptor,PTNinactivatesthephosphataseactivity of RPTPb and therebycauses tyrosine phosphorylation ofthe substrates ofRPTPb suchas b-catenin.21 This mayleadto activationofavarietyofdownstreamsignallingpathways.For example,PTN-inducedlossofassociationofb-cateninwithN- cadherindisruptsthecell-celladhesionandcausesepithelial- mesenchymaltransitioninculturedcells.54
Inthepresentstudy,werevealedPTNtobeexpressedin the SMG without sexual difference in levels. PTN was Fig.9–ImmunohistochemicallocalizationofPTN.Paraffinsectionsofthemale(A)andfemale(B)SMGwereimmunostained withanti-PTNantibodyandvisualizedwiththeenzyme-histochemicalmethod.Ac:acinus;ID:intercalatedduct;SD:
striatedduct;GCT:granularconvolutedtubule.NotethepositiveimmunoreactioninthedistalendportionsofGCT(arrow).
Bar=20mm.(C)AcryostatsectionofthefemaleSMGwasimmunostainedwithanti-PTNantibodyandvisualizedin electronmicroscopywiththepre-embedding,enzyme-histochemicalmethod.Notethedarkimmunoreactionproducts presentintheentireportionsofSDcells(SD)butabsentinacinarcells(Ac).Bar=2mm.
colocalizedwithRPTPb-SinIDcellsofbothsexesandSDcells ofthefemale.Inaddition,inthemalePTNwaspresentincells inthedistalendsofGCTthatarenegativeforRPTPb-S.Since theultrastructurallocalizationofRPTPb-Stoapicalorbasal membranecouldnotbedeterminedinthisstudy,thelocation
of the ligand-receptor interaction is unknown. However, consideringthatthemajorityofIDandSDcellsarelacking inapicalsecretorygranules,theautocrine/paracrinemodeof PTN binding to RPTPb-S is likely operating at the basal membraneofthesecellpopulations.Ontheotherhand,there isevidencethatRPTPb-Sofgastricepithelialcellscanbindto Helicobacter pylorivacuolatingcytotoxinthatissecretedinto thegastriclumen.42Therefore,apossibilityisnotruledout thatRPTPb-SintheSMGislocatedintheapicalmembraneof duct epithelial cells and binds to some unknown ligand secretedintosaliva.
The present study has revealed distinct patterns of expression ofthereceptorandligandintheductsystemof mouseSMG,i.e.,presenceofbothRPTPb-SandPTNinIDcells inbothsexesandSDcellsinthefemale,presenceofPTNbut absenceofRPTPb-SinasubpopulationofGCTcellsadjacentto IDinthemale, andabsence ofbothPTNand RPTPb-Sina majority of GCT cells in the male. These suggest that combinationsofthepresenceorabsenceofRPTPb-SorPTN arecloselyrelatedtoandrogen-dependentprogressionofthe celllineagefromIDtoGCTcells(schematicallypresentedin Fig.11).Whetherornotthisisacausativerelationshipremains to be clarified in future study in terms of the molecular mechanismsunderlyingtheductcelldifferentiationinmouse SMG.
Funding
ThisstudywassupportedinpartbyGrant-in-AidforScientific Research(No.20590187and23590231)toSIfromtheMinistry Fig.10–DoubleimmunostainingforRPTPb-SandPTNintheSMG.Paraffinsectionsofthemale(A),(B)and(C)andfemale(E), (F)and(G)SMGwereimmunostainedsimultaneouslywithratanti-RPTPb-Santiseraandrabbitanti-PTNantibodyand observedwithfluorescentmicroscopy.Afterwashing,thesamemale(D)andfemale(H)SMGsectionswerestainedwithH- E.ThefluorescenceforRPTPb-S(red;(A),(E)andPTN(green;(B),(F)overlapstovariousextentsindifferentportionsofthe ductsystemwhenmerged(yellow;(C),(G).NotethesinglefluorescenceforPTNinthedistalendportionsofGCT(arrows).
Ac:acinus;ID:intercalatedduct;SD:striatedduct.Bar=40mm.
Fig.11–ThedistributionofRPTPb-SandPTNandpossible androgen-dependentprogressionofcelllineageinthe maleandfemalemouseSMG.A:acinus;ID:intercalated duct;GCT:granularconvolutedtubule;SD:striatedduct.
Darknessofthemaskineachductportionsrepresents intensityoftheimmunostaining.
of Education, Culture, Sports, Science and Technology of Japan.
Competing interests
Nonedeclared.
Ethical approval
Ethicalapprovalwasnotnecessaryforthiswork.Theapproval fortheanimalexperimentswasgivenbyKanazawaUniversity (AP-081042andAP-081043).
references
1. PinkstaffCA.Thecytologyofsalivaryglands.International ReviewofCytology1980;63:141–261.
2. JacobyF,LeesonCR.Thepostnataldevelopmentoftherat submaxillarygland.JournalofAnatomy1959;93(2):201–16.
3. CaramiaF.Ultrastructureofmousesubmaxillarygland.I.
Sexualdifferences.JournalofUltrastructureResearch 1966;16(5):505–23.
4. GresikEW.Postnataldevelopmentalchangesin submandibularglandsofratsandmice.Journalof HistochemistryandCytochemistry1980;28(8):860–70.
5. BarkaT.Biologicallyactivepolypeptidesinsubmandibular glands.JournalofHistochemistryandCytochemistry
1980;28(8):836–59.
6. CaramiaF.Ultrastructureofmousesubmaxillarygland.II.
Effectofcastrationinthemale.JournalofUltrastructure Research1966;16(5):524–36.
7. Chre´tienM.Actionoftestosteroneonthedifferentiation andsecretoryactivityofatargetorgan:thesubmaxillary glandofthemouse.InternationalReviewofCytology 1977;50:333–96.
8. GubitsRM,ShawPA,GresikEW,Onetti-MudaA,BarkaT.
Epidermalgrowthfactorgeneexpressionisregulated differentlyinmousekidneyandsubmandibulargland.
Endocrinology1986;119(3):1382–7.
9. KruegerNX,SaitoH.Ahumantransmembraneprotein- tyrosine-phosphatase.PTPzeta,isexpressedinbrainand hasanN-terminalreceptordomainhomologoustocarbonic anhydrases.ProceedingsoftheNationalAcademyofSciencesof theUnitedStatesofAmerica1992;89(16):7417–21.
10.LevyJB,CanollPD,SilvennoinenO,BarneaG,MorseB, HoneggerAM,etal.Thecloningofareceptor-typeprotein tyrosinephosphataseexpressedinthecentralnervous system.JournalofBiologicalChemistry1993;268(14):10573–81.
11.MaurelP,RauchU,FladM,MargolisRK,MargolisRU.
Phosphacan,achondroitinsulfateproteoglycanofbrain thatinteractswithneuronsandneuralcell-adhesion molecules,isanextracellularvariantofareceptor-type proteintyrosinephosphatase.ProceedingsoftheNational AcademyofSciencesoftheUnitedStatesofAmerica 1994;91(7):2512–6.
12.NishiwakiT,MaedaN,NodaM.Characterizationand developmentalregulationofproteoglycan-typeprotein tyrosinephosphatasezeta/RPTPbetaisoforms.Journalof Biochemistry1998;123(3):458–67.
13.GarwoodJ,HeckN,ReichardtF,FaissnerA.Phosphacan shortisoform,anovelnon-proteoglycanvariantof phosphacan/receptorproteintyrosinephosphatase-beta,
interactswithneuronalreceptorsandpromotesneurite outgrowth.JournalofBiologicalChemistry2003;278(26):24164–
73.
14.CanollPD,BarneaG,LevyJB,SapJ,EhrlichM,Silvennoinen O,etal.Theexpressionofanovelreceptor-typetyrosine phosphatasesuggestsaroleinmorphogenesisand plasticityofthenervoussystem.BrainResearch DevelopmentalBrainResearch1993;75(2):293–8.
15.EngelM,MaurelP,MargolisRU,MargolisRK.Chondroitin sulfateproteoglycansinthedevelopingcentralnervous system.I.cellularsitesofsynthesisofneurocanand phosphacan.JournalofComparativeNeurology1996;366(1):34–
43.
16.MaedaN,NodaM.Involvementofreceptor-likeprotein tyrosinephosphatasezeta/RPTPbetaanditsligand pleiotrophin/heparin-bindinggrowth-associatedmolecule (HB-GAM)inneuronalmigration.JournalofCellBiology 1998;142(1):203–16.
17.MilevP,FriedlanderDR,SakuraiT,KarthikeyanL,FladM, MargolisRK,etal.Interactionsofthechondroitinsulfate proteoglycanphosphacan,theextracellulardomainofa receptor-typeproteintyrosinephosphatase,withneurons, glia,andneuralcelladhesionmolecules.JournalofCell Biology1994;127(6Pt1):1703–15.
18.PelesE,NativM,CampbellPL,SakuraiT,MartinezR,LevS, etal.Thecarbonicanhydrasedomainofreceptortyrosine phosphatasebetaisafunctionalligandfortheaxonalcell recognitionmoleculecontactin.Cell1995;82(2):251–60.
19.MaedaN,NishiwakiT,ShintaniT,HamanakaH,NodaM.
6B4proteoglycan/phosphacan,anextracellularvariantof receptor-likeprotein-tyrosinephosphatasezeta/RPTPbeta, bindspleiotrophin/heparin-bindinggrowth-associated molecule(HB-GAM).JournalofBiologicalChemistry 1996;271(35):21446–52.
20.MaedaN,Ichihara-TanakaK,KimuraT,KadomatsuK, MuramatsuT,NodaM.Areceptor-likeprotein-tyrosine phosphatasePTPzeta/RPTPbetabindsaheparin-binding growthfactormidkine.Involvementofarginine78of midkineinthehighaffinitybindingtoPTPzeta.Journalof BiologicalChemistry1999;274(18):12474–9.
21.MengK,Rodriguez-Pen˜aA,DimitrovT,ChenW,YaminM, NodaM,etal.Pleiotrophinsignalsincreasedtyrosine phosphorylationofbetabeta-cateninthroughinactivation oftheintrinsiccatalyticactivityofthereceptor-typeprotein tyrosinephosphatasebeta/zeta.ProceedingsoftheNational AcademyofSciencesoftheUnitedStatesofAmerica
2000;97(6):2603–8.
22.PariserH,Perez-PineraP,EzquerraL,HerradonG,DeuelTF.
Pleiotrophinstimulatestyrosinephosphorylationofb- adductinthroughinactivationofthetransmembrane receptorproteintyrosinephosphataseb/z.Biochemicaland BiophysicalResearchCommunications2005;335(1):232–9.
23.SchmittgenTD,LivakKJ.Analyzingreal-timePCRdataby thecomparativeC(T)method.NatureProtocols2008;3:1101–8.
24.WakayamaT,KatoY,UtsumiR,TsujiA,IsekiS.Atime-and cost-savingmethodofproducingratpolyclonalantibodies.
ActaHistochemicaetCytochemica2006;39(3):79–87.
25.WakayamaT,NakataH,KuroboM,SaiY,IsekiS.
Expression,localization,andbindingactivityoftheezrin/
radixin/moesinproteinsinthemousetestis.Journalof HistochemistryandCytochemistry2009;57(4):351–62.
26.ChangWW,BarkaT.Stimulationofacinarcellproliferation byisoproterenolinthepostnatalratsubmandibulargland.
AnatomicalRecord1974;178(2):203–9.
27.CutlerLS,ChaudhryAP.Cytodifferentiationoftheacinar cellsoftheratsubmandibulargland.DevelopmentalBiology 1974;41(1):31–41.
28.SrinivasanR,ChangWW,vanderNoenH,BarkaT.The effectofisoproterenolonthepostnataldifferentiationand
growthoftheratsubmandibulargland.AnatomicalRecord 1973;177(2):243–53.
29.BarkaT.Inducedcellproliferation:theeffectof isoproterenol.ExperimentalCellResearch1965;37:662–79.
30.BasergaR,HefflerS.StimulationofDNAsynthesisby isoproterenolanditsinhibitionbyactinomycinD.
ExperimentalCellResearch1967;46(3):571–80.
31.ZajicekG,YagilC,MichaeliY.Thestreaming
submandibulargland.AnatomicalRecord1985;213(2):150–8.
32.DennyPC,DennyPA.Dynamicsofparenchymalcell division,differentiation,andapoptosisintheyoungfemale mousesubmandibulargland.AnatomicalRecord
1999;254:408–17.
33.ManY-G,BallWD,MarchettiL,HandA.Contributionsof intercalatedductcellstothenormalparenchymaof submandibularglandsofadultrats.AnatomicalRecord 2001;263:202–14.
34.KimotoM,YuraY,KishinoM,ToyosawaS,OgawaY.Label- retainingcellsintheratsubmandibulargland.Journalof HistochemistryandCytochemistry2008;56(1):15–24.
35.DennyPC,ChaiY,KlauserDK,DennyPA.Parenchymal cellproliferationandmechanismsformaintenanceof granularductandacinarcellpopulationsinadultmale mousesubmandibulargland.AnatomicalRecord 1993;235:475–85.
36.ChaiY,KlauserDK,DennyPA,DennyPC.Proliferativeand structuraldifferencesbetweenmaleandfemalemouse submandibularglands.AnatomicalRecord1993;235(2):303–11.
37.DennyPC,ChaiY,PimprapaipornW,DennyPA.Three- dimensionalreconstructionofadultfemalemouse submandibularglandsecretorystructures.AnatomicalRecord 1990;226(4):489–500.
38.HarrochS,PalmeriM,RosenbluthJ,CusterA,OkigakiM, ShragerP,etal.Noobviousabnormalityinmicedeficientin receptorproteintyrosinephosphatasebeta.Molecularand CellularBiology2000;20(20):7706–15.
39.HarrochS,FurtadoGC,BrueckW,RosenbluthJ,LafailleJ, ChaoM,etal.Acriticalrolefortheproteintyrosine phosphatasereceptortypeZinfunctionalrecoveryfrom demyelinatinglesions.NatureGenetics2002;32(3):411–4.
40.LafontD,AdageT,Gre´coB,ZaratinP.Anovelrolefor receptorlikeproteintyrosinephosphatasezetain modulationofsensorimotorresponsestonoxiousstimuli:
evidencesfromknockoutmicestudies.BehaviouralBrain Research2009;201(1):29–40.
41.SchinkeT,GebauerM,SchillingAF,LamprianouS,Priemel M,MueldnerC,etal.Theproteintyrosinephosphatase Rptpzetaisexpressedindifferentiatedosteoblastsand affectsboneformationinmice.Bone2008;42(3):524–34.
42.FujikawaA,ShirasakaD,YamamotoS,OtaH,YahiroK, FukadaM,etal.Micedeficientinproteintyrosine phosphatasereceptortypeZareresistanttogastriculcer inductionbyVacAofHelicobacterpylori.NatureGenetics 2003;33(3):375–81.
43.KlausmeyerA,GarwoodJ,FaissnerA.Differential expressionofphosphacan/RPTPbetaisoformsinthe developingmousevisualsystem.JournalofComparative Neurology2007;504(6):659–79.
44.MilnerPG,LiYS,HoffmanRM,KodnerCM,SiegelNR,Deuel TF.Anovel17kDheparin-bindinggrowthfactor(HBGF-8)in bovineuterus:purificationandN-terminalaminoacid sequence.BiochemicalandBiophysicalResearch
Communications1989;165(3):1096–103.
45.RauvalaH.An18-kdheparin-bindingproteinofdeveloping brainthatisdistinctfromfibroblastgrowthfactors.EMBO Journal1989;8(10):2933–41.
46.LiYS,MilnerPG,ChauhanAK,WatsonMA,HoffmanRM, KodnerCM,etal.Cloningandexpressionofa
developmentallyregulatedproteinthatinducesmitogenic andneuriteoutgrowthactivity.Science1990;250(4988):1690–
4.
47.VanderwindenJM,MailleuxP,SchiffmannSN,
VanderhaeghenJJ.Cellulardistributionofthenewgrowth factorpleiotrophin(HB-GAM)mRNAindevelopingand adultrattissues.AnatomyandEmbryology1992;186(4):387–
406.
48.WanakaA,CarrollSL,MilbrandtJ.Developmentally regulatedexpressionofpleiotrophin,anovelheparin bindinggrowthfactor,inthenervoussystemoftherat.
BrainResearchDevelopmentalBrainResearch1993;72(1):133–44.
49.Silos-SantiagoI,YehHJ,GurrieriMA,GuillermanRP,LiYS, WolfJ,etal.LocalizationofpleiotrophinanditsmRNAin subpopulationsofneuronsandtheircorrespondingaxonal tractssuggestsimportantrolesinneural-glialinteractions duringdevelopmentandinmaturity.JournalofNeurobiology 1996;31(3):283–96.
50.DeuelTF,ZhangN,YehHJ,Silos-SantiagoI,WangZY.
Pleiotrophin:acytokinewithdiversefunctionsandanovel signalingpathway.ArchivesofBiochemistryandBiophysics 2002;397(2):162–71.
51.HienolaA,PekkanenM,RauloE,VanttolaP,RauvalaH.HB- GAMinhibitsproliferationandenhancesdifferentiationof neuralstemcells.MolecularandCellularNeurosciences 2004;26(1):75–88.
52.AmetLE,LauriSE,HienolaA,CrollSD,LuY,LevorseJM, etal.Enhancedhippocampallong-termpotentiationin micelackingheparin-bindinggrowth-associatedmolecule.
MolecularandCellularNeurosciences2001;17(6):1014–24.
53.PavlovI,Vo˜ikarV,KaksonenM,LauriSE,HienolaA,TairaT, etal.Roleofheparin-bindinggrowth-associatedmolecule (HB-GAM)inhippocampalLTPandspatiallearningrevealed bystudiesonoverexpressingandknockoutmice.Molecular andCellularNeurosciences2002;20(2):330–42.
54.Perez-PineraP,AlcantaraS,DimitrovT,VegaJA,DeuelTF.
Pleiotrophindisruptscalcium-dependenthomophiliccell–
celladhesionandinitiatesanepithelial-mesenchymal transition.ProceedingsoftheNationalAcademyofSciencesof theUnitedStatesofAmerica2006;103(47):17795–800.