金谷 弦・鈴木孝男・牧 秀明・中村泰男・宮島祐一・菊地永祐
2011 年巨大津波が宮城県蒲生潟の地形,植生および底生動物相に及ぼした影響
Effects of the 2011 tsunami on the topography, vegetation, and macrobenthic
fauna in Gamo Lagoon, Japan
金谷 弦
1),*・鈴木孝男
2)・牧 秀明
1)・中村泰男
1)・宮島祐一
3)・菊地永祐
4) 1) (独)国立環境研究所.〒305–8506 茨城県つくば市小野川 16–22) 東北大学大学院生命科学研究科.〒980–8578 仙台市青葉区荒巻字青葉 3) 東北大学理学部生物学科.〒980–8578 仙台市青葉区荒巻字青葉
4) 宮城教育大学環境教育実践研究センター.〒980–0845 仙台市青葉区荒巻字青葉 149
Gen KANAYA, Takao SUZUKI, Hideaki MAKI, Yasuo NAKAMURA, Yuichi MIYAJIMA and
Eisuke KIKUCHI
1) National Institute for Environmental Studies. 16–2 Onogawa, Tsukuba 305–8506, Japan
2) Graduate School of Life Sciences, Tohoku University. Aramaki-Aoba, Aoba-ku, Sendai 980–8578, Japan 3) Biological Institute, Faculty of Science, Tohoku University. Aramaki-Aoba, Aoba-ku, Sendai 980–8578, Japan
4) Environmental Education Center, Miyagi Universiy of Education. 149 Aramaki-Aoba, Aoba-ku, Sendai 980–0845, Japan
Abstract: Gamo Lagoon (Sendai, Miyagi Prefecture, Japan), a shallow brackish lagoon, was struck by a 7.2-m-high tsunami
on 11 March 2011. In order to assess the tsunami-induced changes in the ecosystem of Gamo Lagoon, we examined the to-pography and distribution of plants and macrobenthos between April and August, 2011, and compared them with available pre-tsunami data. The submerged area decreased from 17.3 to 11.8 ha, sand dune vegetation from 8.6 to 0.1 ha, pine forest from 4.2 to 2.0 ha, and macroalgal patches from 7.8 to 1.8 ha. Conversely, the area of bare intertidal flats increased from 4.7 to 5.3 ha, mainly owing to the near disappearance of reed marshes (7.8 to 1.2 ha). Muddy sediment was flushed out and the sediment became sandier. The population of the marsh-associated gastropod Cerithidea rhizophorarum in the lagoon de-clined sharply from about 40,000 (estimated from mean density and habitat area) to just 26 individuals. Of 79 species of macrobenthos found before the tsunami, 47 species (especially bivalves) were absent or nearly extirpated after the tsunami. The temporary immigration of 12 marine species was noted shortly after the tsunami, but most disappeared within 5 months. The populations of dotillid crabs (i.e., Scopimera globosa and Ilyoplax pusilla), the nereidid polychaete Hediste diadroma, and opportunistic species including polychaetes (e.g., Pseudopolydora cf. kempi, Heteromastus cf. similis, and Capitella sp.) and amphipods (Grandidierella japonica and Corophiidae sp.) recovered rapidly within the same 5-month period.
Key Words: community composition, disturbance, intertidal flat, macrobenthic fauna, succession, topography, tsunami,
veg-etation
は じ め に
津波・貧酸素水塊・油汚染などの攪乱は生物群集構造の 改変を引き起こす.撹乱後に群集構造が新たな状態に遷移 してゆく過程を追跡することは,群集構造の決定機構や多 種共存メカニズムを理解するために重要である (Levin etal. 1996; Perkol-Finkel & Airoldi 2010; Botter-Carvalho et al.
2011; Joydas et al. 2012).
津波は,2010 年のチリ地震津波にみられるように,沿 岸域の地形や生物群集を大規模かつ広域的に攪乱して生態
系に非常に大きな影響を与える (Hayasaka et al. 2012; Jara-millo et al. 2012).なかでも底生動物群集は,津波の強度 や生息場所のタイプ−植生や護岸の有無,岩礁帯と砂浜海 岸など−により受ける影響が異なることや,攪乱後の回復 速度が分類群や機能群によって異なることが示されている (Whanpetch et al. 2010; Lomovasky et al. 2011; Jaramillo et al.
2012). 2011 年 3 月 11 日に起きた東日本大震災により,仙台市 の沿岸部は 7 m を超える津波に襲われ,七北田川河口に 位置する蒲生潟一帯も甚大な攪乱を受けた.潟の陸側に設 置されていた防潮堤は数カ所で破断し,海側の砂嘴がほぼ 完全に流失した.そのため,潟は一時的に海に開口した前 浜干潟の様相を呈していた(鈴木 2012; 鈴木・金谷 2012). 干潟やヨシ原も広範囲で流失し,漂砂の流入・堆積により 底質も著しく変化した.こうした津波に伴う物理的攪乱と 日本ベントス学会誌 67: 20–32 (2012)
Japanese Journal of
Benthology
Received 6 June 2012 Accepted 18 July 2012 *Corresponding author
National Institute for Environmental Studies, 16–2 Onogawa, Tsukuba 305–8506, Japan.
環境変化は,蒲生潟に生息する底生動物の組成や出現頻度 にも大きな影響を及ぼしたと予想される. 蒲生潟と周囲のヨシ原や海浜植物群落は,希少な底生動 物や鳥類の生息場所として重要であり,長年にわたり調査 研究が行われてきた(栗原 1975; 栗原ら 2001; 蒲生干潟自 然再生協議会 2008).2004 年から 2009 年の間にも,環境 と底生動物の調査(例えば環境省 2007; Kanaya et al., 2007; 鈴木未発表)が行われており,津波前後での詳細な比較が 可能である.本研究では,震災後の 5 ヶ月間に蒲生潟で 5 回の現地調査を行い,地形,植生(ヨシ原,マツ林,海浜 植物群落,オゴノリ群落),底質の粒度組成および底生動 物相の変化を調べた.調査結果を基に,蒲生潟一帯の汽水 域生態系が津波により受けた攪乱の程度と,今後の回復可 能性について検討した.また,底生動物の個体群が津波に より受けた攪乱の程度と,それぞれの種の生態的特性との 関連について考察した.
震災前における調査地の概要
蒲生潟は,仙台市北部を流れる七北田川の河口北側に発 達した奥行き 850 m,幅約 250 m の浅く富栄養な汽水性潟 湖であり (Fig. 1),一帯は国の鳥獣保護区(蒲生特別保護 地区)および宮城県の自然環境保全地域に指定されてい る.潟入口には導流堤が設置され,水交換は 3 基の水門を 介して行われており (Fig. 2a, 2c),干潮時には導流堤付近 を中心に砂泥質の干潟が出現した.潟の陸側には,津波を 防ぐための防潮堤が建設されていた.隣接した養魚池から 淡水が流入し,塩分は近年では概ね 25∼30 の範囲にあっ た(蒲生干潟自然再生協議会 2008).潟は富栄養化が進行 しており,夏期には水のクロロフィル a 濃度が 100 μg L−1 にも達した (Kanaya et al. 2007).潟奥部には還元的な軟泥 が堆積し,底生動物の斃死も生じていた(金谷・菊地 2011). 埋在性底生動物の個体数では,多毛類のヒメヤマトカワ ゴカイHediste atoka やドロオニスピオ Pseudopolydora cf. kempi が優占し,導流堤付近では二枚貝のイソシジミ Nut-tallia japonica が優占した(金谷・菊地 2011).護岸や石積 にはマガキCrassostrea gigas などの付着生物が生息し,潟 中央の軟泥域にはカキ礁が点在した.潟辺縁にはヨシ Phragmites australis が優占する塩性湿地が発達し,カワザ ンショウガイ類や仙台湾を北限とするフトヘナタリ Ceri-thidea rhizophorarum が多数生息した(環境省 2007; 蒲生干 潟自然再生協議会 2008).潟奥部にはオゴノリ Gracilaria vermiculophylla が密生し,ヨコエビ類の生息基質となっていた (Aikins & Kikuchi 2002).
方 法
地形と植生 2011 年 6 月 20 日(大潮)に,現場で GPS (GPSmap 60CSx, Garmin) を用いて最満潮時と最干潮時に汀線をトレースし た.画像解析ソフト ImageJ により,潟湖の水面積と潮間 帯の面積を推定した.潟湖の水面積と潮間帯面積から,ヨ シ原を除いたものを開放水面および裸地干潟の面積とし た.震災前の潟湖と干潟の面積は,2008 年 9 月 1 日およ び2010 年 4 月 4 日 の Google Earth 画 像 か ら 推 定 し た. 2011 年 7 月 6 日の満潮時に潟内の 63 地点 (Fig. 6b)で測 深を行い,各地点の相対高度(標高換算)を求めた.震災 前の相対高度分布は,1997 年 8 月に潟内の 45 地点 (Fig. 6b)で行った測深の結果から作成した. 2011 年 6 月 21 日に,現場で GPS によりヨシ原と海浜 植物群落をトレースし,面積を推定した.マツ林の面積 は,2011 年 4 月 6 日 の Google Earth 画 像 か ら 算 出 し た. オゴノリは,2011 年 6 月と 7 月には一時的に絶滅状態に あったが,7 月後半に回復の兆しが見られたため 8 月 2 日 に分布調査を行った.潟内を踏査し,オゴノリ藻体の発見 時に場所と被度を記録した.震災前における各植生の分布 面積は,蒲生干潟自然再生協議会 (2008) の植生図から算 出した.なお,同植生図の調査時期は,湿地・陸上植生は 2003 年,オゴノリについては 2004 年 8 月である. 底質と塩分 震災後の調査では,調査時に底質(砂質または泥質)を 定性的に記録した.2011 年 7 月 31 日と 8 月 2 日には,潟 全域に設けた 63 地点 (Fig. 6b) において,粒度分析用の底 土コア(内径 5 cm, 深さ 10 cm まで)を採取した.実験室 において,深さ 1∼4 cm までの底土を 60℃で 48 時間乾燥 させた後,0.063 mm 目のふるいを用い,湿式法により底 土のシルトクレイ含量を求めた.震災前のシルトクレイ含 量については,1997 年 8 月に潟内の 45 地点 (Fig. 6b) で 測定されたデータを使用した(金谷・菊地 2011).底土コ アの採取時に,現場で底土中に電気伝導度計 (CM-21P, TOA-DKK) を差し込み(深さ 10 cm),間隙水の塩分を測 定した.潟内と養魚池の表層水の塩分についても,各調査 日に同機器を用いて測定した.間隙水と表層水の塩分は, いずれも干潮時に測定した. 震災前後における底生動物相と出現頻度の比較 底生動物の定性調査を 2011 年 4 月 21 日,6 月 15 日,6 月 20 日∼21 日,7 月 5 日∼7 日,7 月 31 日∼8 月 4 日 に 行った.干潮時と満潮時に 1∼2 名で潟全域を汀線に沿っ て踏査し,出現した表在性・付着性の底生動物の種を,肉 眼による目視によって判別し記録した.また,残存したヨ シ原やマツ林内,護岸壁の上や転石の下も確認した.潟内金谷 弦・鈴木孝男・牧 秀明・中村泰男・宮島祐一・菊地永祐 の干潟や潮下帯の数カ所において,底土をスコップで深さ 30 cm まで複数回掘り起こし,出現した埋在生物の種を, 肉眼による目視によって判別し記録した.8 月の調査時に は,1 mm 目のふるい上でオゴノリを洗い出し,ヨコエビ 類を確認した.確認された種について,出現頻度を 5 段階 で評価した.(1) −:未発見,(2) +:1 日の調査(2∼4 時間)で 1 個体しか見つからない,(3) L: 1 日の調査(2 ∼4 時間)で数個体が見つかる,(4) M: 数個体をより短時 間で発見できる,(5) H: 数十∼数百個体を容易に発見でき る.埋在性底生動物については,以下の方法で行った定量 採集の結果も考慮し,出現頻度を評価した.2011 年 8 月 上旬に,63 地点 (Fig. 6b)でコアサンプラー(内径 15 cm) を用いて深さ 30 cm までの底土を採取し,1 mm 目のふる い上に残ったものを全て 5%中性ホルマリンで固定し,実 験室に持ち帰った.サンプル中の底生動物を,実体顕微鏡 下で同定・計数した.震災前の生物相と出現頻度は,2004 年(環境省 2007),2007 年と 2008 年(鈴木未発表),2005 年と 2009 年(Kanaya et al. 2007, 金谷未発表)の調査結果 に基づいて評価した.なお,2007 年と 2008 年の調査は, 本研究と同じ手法による定量採集(32 地点)であり,そ の他は定性的な調査である. 底生動物各種の出現頻度を震災前後で比較し,津波の影 響 を 6 段 階 で 評 価 し た.(1) 絶 滅(●),(2) 絶 滅 寸 前 (○):激減し新規加入も確認出来ない,(3) 減少(△), (4) 耐性・回復(→):影響が軽微もしくは回復,(5) 増加 (↑),および(6) 移入 (Temp):海域からの一時的移入. 津波以前に絶滅していた可能性があるハマガニ Chasmag-nathus convexus については情報不足 (?) とした.また,各 種の生息場所を以下のように類別した.(1) Alga:大型藻 類,(2) Hard: 護 岸 や 転 石 上,(3) Marsh: ヨ シ 原,(4) Land: マ ツ 林 や 砂 浜, 草 原, お よ び(5) Soft-bottom (Sb):底土中やその表面. 震災前後におけるフトヘナタリの個体群サイズ フトヘナタリは表在性であり,分布域と生息個体数を目 視で比較的容易に確認できることから,震災後に発見され た全個体の位置を GPS で記録した.重複カウントを避け るために,位置を記録した個体は隔離されたヨシのパッチ
Fig. 1. Location and topography of Gamo Lagoon before the tsunami. The surrounding area immersed by the tsunami on 11 March 2011 is
内に移植した.2011 年 6∼8 月の調査で確認された全個体 数を,震災後の個体群サイズとした. 蒲生潟のフトヘナタリはヨシ原内の潮間帯上部に限定し て生息し,震災前の生息場所(A∼E の 5 エリア,Fig. 4) がわかっている.また,2002 年 5 月には潟内の 13 地点 (n =112) で 0.25 m2または1 m2方形枠を用いた生息密度の 調査が行われている(宮島未発表).そこで,画像解析ソ フトImageJ により各エリア毎に分布面積を算出し,これ と各エリアの平均生息密度を用いて震災前の個体群サイズ を推定した.
結 果
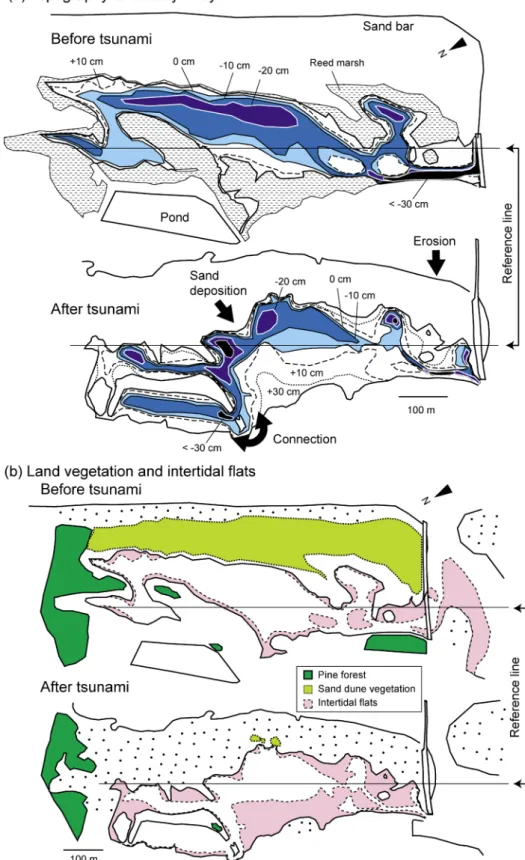
津波による潟湖地形の変化 震災前後の蒲生潟の形状と水深分布を Fig. 3a に示す. 震災直後には砂嘴が流失し,蒲生潟は前浜干潟の様相を呈 していたが(鈴木 2011),6 月 20 日になると漂砂が堆積し 砂嘴は復旧していた (Fig. 2b).海岸線は最大で 100 m ほ ど陸側に移動し,潟奥部の海側は砂が流入し陸化した.堤 が破断したため淡水池と潟が繋がり,陸側のヨシ原は流失 し砂干潟となった.導流堤の石積は崩壊し (Fig. 2d),防 潮堤も 4 カ所で破断した (Fig. 2e).しかし,潟の深度分 布に顕著な変化は見られず,ほぼ全域が−30∼+30 cm T.P. の範囲にあった. 潟奥部の陸化に伴い,ヨシ原を含む水面積は 5.5 ha (32%) 減少した (Table 1).しかし,ヨシ原の裸地化や淡 水池との連結により開放水面の面積は 0.3 ha 増加し,導流 堤内側の干潟も 2.5 ha 増加した.一方,七北田川河口部に あった干潟 (1.9 ha) は消失したため,七北田川河口部と蒲 生潟内の干潟は全体として 0.7 ha (15%) 増加した. 津波による植生面積の変化 津波により海浜植物群落の 99%が流失した (Fig. 3b お よびTable 1).残存したパッチ内では,ハマヒルガオCa-lystegia soldanella やハマナス Rosa rugosa が確認された.
マツ林も 52%が流失し,残存個体にも立ち枯れが認めら れた.ヨシ原も 7.8 ha のうち 6.6 ha (84%) が流失し (Fig. 4),残存パッチ内では稈密度の減少や草丈の小型化(震災 前の半分以下)が確認された.震災前には開放水面積の 75% (7.8 ha) にオゴノリが密生していたが(Fig. 5 および Table 1),津波でほぼ全てが流失し,8 月になると潟奥部 や旧淡水池で再び生育が確認された(面積 1.8 ha,被度
Fig. 2. Photographs of Gamo Lagoon. A panoramic view of the lagoon before (a) and after (b) the tsunami, a levee before (c) and after (d) the
金谷 弦・鈴木孝男・牧 秀明・中村泰男・宮島祐一・菊地永祐
Fig. 3. Changes in lagoon topography and bathymetry (a) and in extents of land vegetation and bare intertidal flats (b) following the tsunami.
Relative height (cm T.P.) is indicated with lines and different colors. The dotted area indicates the reed marshes before the tsunami. A reference line is placed in the same location in each map. Land vegetation data before the tsunami are from literature (蒲生干潟自然再生協議会 2008).
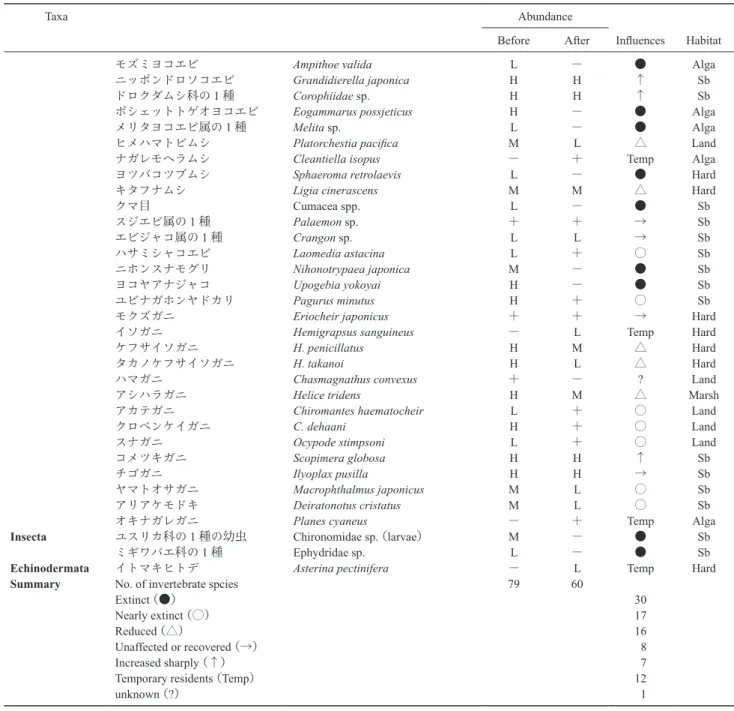
0∼30%). 底土のシルトクレイ含量と表層水・間隙水の塩分 潟の表層水の塩分は,6 月 20 日∼21 日には 25.1∼28.2, 7 月 5 日∼7 日 に は 7.0∼24.2,7 月 31 日∼8 月 4 日 に は 24.2∼27.2 の範囲にあった.津波が流入した養魚池の塩分 は,6 月 20 日に 12.1∼13.7,7 月 6 日に 6.9∼8.6 と徐々に 低下した.間隙水の塩分 (Fig. 6a) は 6.6∼27.4 の範囲にあ り,潟中央で高く潟縁辺部で低い傾向を示した.間隙水塩 分は,養魚池と隣接した潟陸側の干潟部で 6.6∼10 と最も 低かった. 津波により,潟奥部に堆積していた軟泥は流失し,底質 は著しく砂質化した (Fig. 6b).津波前には,潟奥部の広 範囲にシルトクレイ含量が 50%を超える軟泥が堆積して いたが,津波後にはシルトクレイ含量が 10%を超える地 点はわずかに 3 地点 (19.9∼25.6%) のみとなった.震災前 (1997 年 8 月)におけるシルトクレイ含量の潟平均値は 30.5±31.6% (SD, n=45) であったが,震災後(2011 年 8 月)の潟平均値は 4.1±4.6% (SD, n=63) へと大きく減少 した.ほとんどの地点で,堆積した砂の厚さは 30 cm を超 えており,底土を深く掘り返しても軟泥の層は確認できな かった. 震災前の底生動物相 震災前(2004 年∼2009 年)には 79 種が確認され,軟体 動物が 23 種,環形動物が 20 種(うち多毛類が 19 種),甲 殻類が 29 種を占めた (Table 2).多毛類のカワゴカイ属 (ヒメヤマトカワゴカイ),ドロオニスピオ,ホソイトゴカ イHeteromastus cf. similis,ヨコエビ類のニッポンドロソコ
エビGrandidierella japonica とドロクダムシ科の 1 種
Co-rophiidae sp. が優占し,埋在性二枚貝ではイソシジミ,ア サリRuditapes philippinarum,サビシラトリガイ Macoma
Table 1. Areas (ha) of the submerged zone, bare tidal flat, and vegetated zones of Gamo Lagoon before and after the tsunami. Areas (m2) inhab-ited by the gastropod Cerithidea rhizophorarum and their density are also given. A–E are the same as in Fig. 4.
Before After Difference (%)
Submerged area at high tide (ha)
Open water + marsh 17.3 11.8 − 5.5 (− 32)
Open water 10.4 10.7 0.3 (+3)
Bare tidal flat (ha)
Inside of levee 2.8 5.3 2.6 (+92)
Outside of levee 1.9 0.0 − 1.9 (− 100)
Total 4.7 5.3 0.7 (+15)
Vegetation (ha)
Sand dune vegetation 8.6 0.1 − 8.5 (− 99) Pine forest (Pinus spp.) 4.2 2.0 − 2.2 (− 52) Reed marsh (Phragmites australis) 7.8 1.2 − 6.6 (− 84) Macroalgal patch (Gracilaria vermiculophylla)
Total 7.8 1.8 − 6.0 (− 76)
>50% coverage 5.9 0.0 − 5.9 (− 100) 30–50% coverage 0.6 0.0 − 0.6 (− 100)
0–30% coverage 1.3 1.8 0.6 (+45)
Gastropod Cerithidea rhizophorarum population a Area (m2) A 178 B 3106 C 1679 D 835 E 1244 Total 7041
Mean density (ind. m−2)
A 4.9 B no data C 6.5 D 3.1 E no data Lagoonal average 5.8 Total population 39580b 26 − 39554 (− 99.93) Areas were calculated from GPS data (22 June 2011), Google Earth images (1 September 2008, 4 April 2010, and 6 April 2011), field observations, and vegetation maps (蒲生干潟自然再生協議会 2008).
a Mean density before the tsunami was obtained in 2002 (Miyajima, unpubl. data, n=112 in total) for areas A, C, and D (see Fig. 5). The lagoonal averaged density was used for areas B and E where density data before the tsunami were not available.
金谷 弦・鈴木孝男・牧 秀明・中村泰男・宮島祐一・菊地永祐
contabulata,ソトオリガイ Laternula marilina が多く見られ
た.地高の高い砂質干潟にはコメツキガニ Scopimera
glo-bosa が,砂泥質干潟にはチゴガニ Ilyoplax pusilla が群生
し,低潮線付近にはアリアケモドキ Paracleistostoma cris-tatum が分布した.ヨシ原にはヒラドカワザンショウ As-siminea hiradoensis をはじめとするカワザンショウガイ類, フトヘナタリやアシハラガニHelice tridens が生息し,マ ツ林にはクロベンケイガニ Chiromantes dehaani が群生し た.護岸上や転石帯にはタマキビ Littorina brevicula やマ ガキ,ケフサイソガニ属の 2 種が見られた.軟泥域には多 毛類の Capitella 属の 1 種,ユスリカ科の 1 種 Chironomi-dae sp. やミギワバエ科の 1 種 EphydriChironomi-dae sp. が生息し,オ ゴノリ群落内にはポシェットトゲオヨコエビ Eogammarus possjeticus やメリタヨコエビ属の 1 種 Melita sp.,モズミヨ コエビAmpithoe valida が生息した. 津波による底生動物への影響 2011 年の 4 月∼8 月に 60 種の大型底生動物が確認され
Fig. 4. Changes in the reed marsh vegetation and spatial distribution of the marsh-associated gastropod Cerithidea rhizophorarum (areas A to E)
following the tsunami. Individuals of C. rhizophorarum found after the tsunami (April–August, 2011) are indicated with arrows. A reference line is inserted at the same location in each map. Marsh vegetation data before the tsunami are from literature (蒲生干潟自然再生協議会 2008).
Fig. 5. Changes in macroalgal coverage (Gracilaria vermiculophylla) following the tsunami. Coverage (%) is indicated with different colors.
た(Table 2,タマシキゴカイについては糞塊のみ).ここ から,海域からの一時的移入種(12 種)と情報不足であ るハマガニを除外すると 47 種となり,震災前に生息して いた79 種のうち 30 種が絶滅(−)したことになる.ま た,残る 47 種のうち 17 種が絶滅寸前と評価された. 震災前の出現頻度が高い(H または M)種のうち,絶 滅または絶滅寸前と評価された種は 19 種である.内訳は, フトヘナタリ,タマキビ,アラムシロ Nassarius festivus, ムラサキイガイ,マガキ,サビシラトリガイ,ヒメシラト リMacoma incongrua,アサリ,オオノガイ Mya arenaria oonogai,ソトオリガイ,シロスジフジツボ Fistulobalanus albicostatus,ポシェットトゲオヨコエビ,ニホンスナモグ
リNihonotrypaea japonica,ヨコヤアナジャコ Upogebia yo-koyai,ユビナガホンヤドカリ Pagurus minutus,クロベン
ケイガニ,ヤマトオサガニ,アリアケモドキ,ユスリカ科 の1 種である.震災前の出現頻度が高い(H または M) 種のうちで減少した種 (△) は 12 種であり,その内訳はタ テジマイソギンチャクHaliplanella lineata,ヒモムシ類 Nemertinea spp.,クリイロカワザンショウ,ヒラドカワザ ンショウ,イソシジミ,ヒメヤマトカワゴカイ,ヤマトス ピ オPrionospio japonica, ヒ メ ハ マ ト ビ ム シ Platorchestia pacifica, キタフナムシ Ligia cinerascens, ケフサイソガニ属
の2 種,アシハラガニであった.イソシジミについては生 息密度が減少したものの,二枚貝の中で唯一,殻長 4 cm 以上の生残個体が潟内の広範囲で確認された. チゴガニは新たに形成された干潟への新規加入がみられ たため,個体群サイズは震災前と同等(→)と推定され た.震災後に著しく増加した種(↑)として,多毛類とヨ コエビ類の 6 種(ヤマトカワゴカイ Hediste diadroma, ドロ オニスピオ,Capitella 属の 1 種,ホソイトゴカイ,ニッポ ンドロソコエビ,ドロクダムシ科の 1 種)が挙げられた. 彼らの生息密度は,場所によっては震災前の数倍に達し た.また,コメツキガニも津波で形成された地高の高い砂 干潟へ多数が加入したため,生息域が著しく拡大し個体数 も増加した. フトヘナタリの個体群サイズ 震災前に蒲生潟のフトヘナタリは A∼E の 5 エリアに分 布し (Fig. 4),その面積はそれぞれ 178 m2, 3106 m2, 1679 m2, 835 m2, 1244 m2であった (Table 1).エリア A, C およ びD における震災前の生息密度はそれぞれ 4.9, 6.5 および 3.1 ind. m−2であった.エリア B と E については震災前の デ ー タ が無いため,エリア A, C, D における平均密度 (5.8 ind. m−2) を用いて個体群サイズを推定した.5 エリ アを合算すると,震災前の個体群サイズは 39,580 個体と 推定された.震災後に発見された個体は 26 個体であり, 個体群サイズは千分の一以下となった.彼らはいずれも震 災前の生息エリア近傍で発見されたが,ヨシ原が流失した ためほとんどが裸地干潟上で確認された. 一時的移入種 津波で運ばれてきた,もしくは破断した砂嘴から一時的 に潟内に移入したと考えられた種 (Temp) は,アラレタマ キビNodilittorina radiata, ナミノコガイ Latona cuneata, シ
ズクガイTheora lubrica, コタマガイ Gomphina melanegis,
チョウセンハマグリMeretrix lamarckii, タマシキゴカイ Ar-enicola basiliensis と思われる多数の糞塊,ユムシ Urechis unicinctus, ナガレモヘラムシ Idotea metallica, オキナガレガ
ニPlanes cyaneus, イ ト マ キ ヒ ト デ Asterina pectinifera で
あった.震災後にはがれきが潟内に漂着したため,これら に付着したエボシガイ Lepas anatifera も確認された.これ らの種は,時間が経つにつれ減少し,8 月以降も確認され たのはナミノコガイとコタマガイのみであった.
考 察
蒲生潟は津波により導流堤と防潮堤が破壊され,砂嘴や ヨシ原が流失するなど大きな物理的攪乱を受けた.潟奥部 が陸化し,七北田川河口部の干潟が流失するなど,地形改 変は非常に大きかったが,震災後 3 ヶ月で砂嘴が復旧し, 2011 年 6 月の時点で潟は旧来の形状を取り戻しつつあっ た.水深分布も震災前とほぼ同様で,干潮時には干潟が出 現した.しかし,潟湖地形が比較的短期間で回復したのに 対し,植生の回復には未だ時間を要すると考えられる. 潟周辺のマツ林は 52%が流失し,海側砂嘴の海浜植物 群 落 は ほ ぼ 全 て が 流 失 し た (Table 1).Hayasaka et al. (2012) は,2011 年 8 月に岩手県沿岸の海浜植生を調査し, 群集の多様性や均等度が低下すると共に,攪乱耐性の高い 少数の陸生種が優占したことを報告している.蒲生潟で は,2011 年 8 月の時点においても砂嘴上に植物がほとん ど見られなかった.これは,砂嘴のほぼ全てが津波で流失 するなど,攪乱の規模が非常に大きかったことによると推 測される.ヨシ原も,84%が流失し密度や成長の低下が認 められた.ヨシは本来淡水性であり,塩分 10 以上で発芽 に負の影響が認められ (Mauchamp & Mésleard 2001),稈や 葉の生育不良はさらに低塩分でも生じる (Lissner & Schi-erup 1997).今後,ヨシ原が回復するためには低塩分環境 が維持される必要がある.幸い,底土間隙水の塩分は陸側 の干潟で低く (<10),地下を通じて淡水の供給があるこ とを示唆している.蒲生潟のヨシ原を保全する上では,潟 陸側の地域を淡水供給源として維持していくことが重要と 考えられる. 震災により,多くの底生動物が絶滅または絶滅寸前とな り,その影響は生息場所のタイプを問わず大きかった (Table 2).分類群毎に比較すると,巻貝では震災前に確認 された8 種のうちクリイロカワザンショウとヒラドカワザ ンショウを除く 6 種,二枚貝では 15 種のうちホトトギス ガイとイソシジミを除く 13 種が絶滅または絶滅寸前であ り,津波の影響が大きかった.一方,多毛類は,震災前に金谷 弦・鈴木孝男・牧 秀明・中村泰男・宮島祐一・菊地永祐
Table 2. Abundance of macrobenthic fauna in Gamo Lagoon before (2004–2009) and after (April–August 2011) the tsunami. The influence of
the tsunami on the population of each species was evaluated. Habitat type is shown for each species.
Taxa Abundance
Before After Influences Habitat
Cnidaria タテジマイソギンチャク Haliplanella lineata M L △ Hard
Platyhelminthes 多岐腸目の 1 種 Polycladida sp. L + △ Sb
ウズムシ綱の 1 種 Turbellaria sp. L + △ Sb
Nemertea ヒモムシ類 Nemertinea spp. M L △ Sb
Phoronida ホウキムシ動物門の 1 種 Phoronida sp. L − ● Sb
Gastropoda ウミニナ Batillaria multiformis L − ● Sb
フトヘナタリ Cerithidea rhizophorarum H L ○ Marsh アラレタマキビ Nodilittorina radiata − L Temp Hard
タマキビ Littorina brevicula H + ○ Hard
サザナミツボ Elachisina ziczac L − ● Sb
クリイロカワザンショウ Angustassiminea castanea M L △ Marsh ヒラドカワザンショウ Assiminea hiradoensis H M △ Marsh ムシヤドリカワザンショウ A. parasitologica L − ● Marsh
アラムシロ Nassarius festivus M − ● Sb
Bivalvia ムラサキイガイ Mytilus galloprovincialis H L ○ Hard
ホトトギスガイ Musculus senhousia L + △ Sb
マガキ Crassostrea gigas H L ○ Hard
チリハギガイ Lasaea undulata L − ● Sb
ナミノコガイ Latona cuneata − + Temp Sb
ユウシオガイ Moerella rutila L − ● Sb
サビシラトリガイ Macoma contabulata H L ○ Sb
ヒメシラトリ M. incongrua M L ○ Sb
シズクガイ Theora lubrica − + Temp Sb
イソシジミ Nuttallia japonica H M △ Sb
マテガイ Solen strictus L − ● Sb
ウネナシトマヤガイ Trapezium liratum L − ● Hard
カガミガイ Phacosoma japonicum + − ● Sb
アサリ Ruditapes philippinarum H L ○ Sb
コタマガイ Gomphina melanegis − L Temp Sb チョウセンハマグリ Meretrix lamarckii − + Temp Sb
オキシジミ Cyclina sinensis + − ● Sb
オオノガイ Mya arenaria oonogai M + ○ Sb
ソトオリガイ Laternula marilina H + ○ Sb
Annellida ホソミサシバ Eteone longa + − ● Sb
ハナオカカギゴカイ Sigambra tentaculata + + → Sb
ヤマトカワゴカイ Hediste diadroma L H ↑ Sb
ヒメヤマトカワゴカイ H. atoka H L △ Sb
スナイソゴカイ Perinereis nuntia brevicirris L − ● Sb
イトメ Tylorrhynchus osawai L L → Sb Nephtys 属の 1 種 Nephtys sp. L − ● Sb ナガホコムシ Leitoscoloplos pugettensis L − ● Sb ヤマトスピオ Prionospio japonica H M △ Sb イトエラスピオ P. pulchra L − ● Sb ドロオニスピオ Pseudopolydora cf. kempi H H ↑ Sb Pseudopolydora 属の 1 種 Pseudopolydora sp. L − ● Sb ミズヒキゴカイ Cirriformia comosa L L → Sb ツツオオフェリア Armandia lanceolata + − ● Sb Capitella 属の 1 種 Capitella sp. H H ↑ Sb ホソイトゴカイ Heteromastus cf. similis H H ↑ Sb シダレイトゴカイ属の 1 種 Notomastus sp. L + △ Sb タマシキゴカイ? Arenicola cf. brasiliensis − L (feces) Temp Sb ヒャクメニッポンフサゴカイ Thelepus japonicus + − ● Sb
ケヤリムシ科の 1 種 Sabellidae sp. L − ● Sb
イトミミズ科の 1 種 Tubificidae sp. L L → Sb
ユムシ Urechis unicinctus − + Temp Sb
Crustacea シロスジフジツボ Fistulobalanus albicostatus H L ○ Hard
エボシガイ Lepas anatifera − + Temp Hard
確認された 19 種のうち 10 種の生息が確認され,そのうち 4 種が著しく増加するなど,津波による影響が比較的小さ く回復も早かった.甲殻類は震災前に確認された 29 種の うち16 種が絶滅または絶滅寸前と評価された.特にオゴ ノリに付着して生息する 3 種のヨコエビ類,半陸生のクロ ベンケイガニ,埋在性のニホンスナモグリ,ヨコヤアナ ジャコ,ヤマトオサガニ,アリアケモドキ,表在性のユビ ナガホンヤドカリの減少が顕著であった.軟泥域に特異的 であったユスリカ科の 1 種や,ミギワバエ科の 1 種も確認 されていない.津波により生息場所であるオゴノリやマツ 林,砂泥質の干潟や軟泥が失われたことが主な要因と考え られる.これらの種の多くは,8 月以降も新規加入が確認 出来ておらず,個体群の回復には時間を要すると思われ る. 震災後の 5 ヶ月間に,4 種の多毛類(ヤマトカワゴカ イ,ドロオニスピオ,ホソイトゴカイ,Capitella 属の 1 種)と 2 種のヨコエビ類(ニッポンドロソコエビやドロク ダムシ科の 1 種)が潟内で著しく増加した.このうちドロ Taxa Abundance
Before After Influences Habitat
モズミヨコエビ Ampithoe valida L − ● Alga
ニッポンドロソコエビ Grandidierella japonica H H ↑ Sb ドロクダムシ科の 1 種 Corophiidae sp. H H ↑ Sb ポシェットトゲオヨコエビ Eogammarus possjeticus H − ● Alga
メリタヨコエビ属の 1 種 Melita sp. L − ● Alga
ヒメハマトビムシ Platorchestia pacifica M L △ Land ナガレモヘラムシ Cleantiella isopus − + Temp Alga ヨツバコツブムシ Sphaeroma retrolaevis L − ● Hard
キタフナムシ Ligia cinerascens M M △ Hard
クマ目 Cumacea spp. L − ● Sb スジエビ属の 1 種 Palaemon sp. + + → Sb エビジャコ属の 1 種 Crangon sp. L L → Sb ハサミシャコエビ Laomedia astacina L + ○ Sb ニホンスナモグリ Nihonotrypaea japonica M − ● Sb ヨコヤアナジャコ Upogebia yokoyai H − ● Sb ユビナガホンヤドカリ Pagurus minutus H + ○ Sb
モクズガニ Eriocheir japonicus + + → Hard
イソガニ Hemigrapsus sanguineus − L Temp Hard
ケフサイソガニ H. penicillatus H M △ Hard
タカノケフサイソガニ H. takanoi H L △ Hard
ハマガニ Chasmagnathus convexus + − ? Land
アシハラガニ Helice tridens H M △ Marsh
アカテガニ Chiromantes haematocheir L + ○ Land
クロベンケイガニ C. dehaani H + ○ Land
スナガニ Ocypode stimpsoni L + ○ Land
コメツキガニ Scopimera globosa H H ↑ Sb
チゴガニ Ilyoplax pusilla H H → Sb
ヤマトオサガニ Macrophthalmus japonicus M L ○ Sb アリアケモドキ Deiratonotus cristatus M L ○ Sb オキナガレガニ Planes cyaneus − + Temp Alga
Insecta ユスリカ科の 1 種の幼虫 Chironomidae sp. (larvae) M − ● Sb
ミギワバエ科の 1 種 Ephydridae sp. L − ● Sb
Echinodermata イトマキヒトデ Asterina pectinifera − L Temp Hard
Summary No. of invertebrate spcies 79 60
Extinct (●) 30
Nearly extinct (○) 17
Reduced (△) 16
Unaffected or recovered (→) 8
Increased sharply (↑) 7
Temporary residents (Temp) 12
unknown (?) 1
Abbreviations: −, not found; rare (+) and low (L), only one or a few individuals, respectively, were found per 1-day survey; intermediate (M), several individuals were easily found in a short time; high (H), many individuals were distributed widely in the lagoon; ●, absent; ○, nearly extir-pated; ∆, reduced; →, unaffected or recovered; ↑, increased sharply; temp, temporary residents (i.e., marine species); ?, unknown; Alga, macroal-gae; Hard, hard substrate; Marsh, reed marsh; Land, terrestrial zone; Sb, soft-bottom.
金谷 弦・鈴木孝男・牧 秀明・中村泰男・宮島祐一・菊地永祐
オニスピオ,ホソイトゴカイ,Capitella 属の 1 種は周年産 卵を行いライフサイクルも短い日和見種 (opportunistic spe-cies) で あ る (Grassle & Grassle 1974; Kanaya et al. 2011). ニッポンドロソコエビとドロクダムシ科の 1 種も,個体数 が春∼夏に急激に増加すること (Aikins & Kikuchi 2002), 周年抱卵個体が観察されることなどから,前に挙げた 3 種 と同様の生活史特性を有すると考えられる.したがって, 不安定な環境下で迅速な増殖を可能とする彼らの生活史特 性が,津波後のいち早い回復をもたらした要因と考えられ る (Grassle & Grassle 1974; Levin et al. 1996; Perkol-Finkel & Airoldi 2010; Joydas et al. 2012).一方,ヤマトカワゴカイ は年 1 回産卵であり,蒲生潟では 4 月の大潮時に繁殖群泳 が観察され,初夏は浮遊幼生の着底時期にあたる (Kikuchi & Yasuda 2004).本種については,津波後にタイミング良 く繁殖・加入期を迎えたため,空白のハビタットに加入で きた可能性が高い.震災後に増加したコメツキガニとチゴ ガニは,それぞれ 6∼8 月と 5∼9 月が繁殖期であり,夏か ら秋が新規加入のピークである(博多湾での事例:Henmi & Kaneto 1989).2011 年 6∼8 月の調査時には,震災前か らの生き残りと思われる比較的大型の個体も確認されてお り,幼生の新規加入だけでなく,生残個体による移動分散 も,震災後の生息域拡大に寄与した可能性がある. 震災前に約 40,000 個体が生息していたフトヘナタリは, ヨシ原の消滅に伴いほぼ絶滅した (Table 1).本種は,仙 台湾では松島湾以南の干潟に生息しているが(環境省 2007),蒲生潟より南の生息地(井土浦,鳥の海,広浦, 鳥の海,松川浦)では分布域が狭く(金谷・鈴木未発表), いずれの干潟も津波で甚大な攪乱を受けている(鈴木 2011, 2012).一方,蒲生潟から 10 km ほど北に位置する松 島湾内は津波の影響が小さく,個体群が健全な状態で残さ れている(鈴木 2012).今後,ヨシ原が再生・復元し他地 域からの幼生供給があれば,蒲生潟の個体群が回復する可 能性がある.しかし,一般に仙台湾南部では北向きの沿岸 流が卓越するため(三浦ら 2008),松島湾から蒲生潟への 幼生供給・定着は起こりにくいかもしれない.一方,同じ くヨシ原に生息するヒラドカワザンショウは,2011 年の 9 月以降急速に個体群サイズを回復しつつあることが確認さ れている.蒲生潟のヒラドカワザンショウの加入時期は 8 ∼9 月であり(Kurata & Kikuchi 1999:ここではカワザン ショウガイAssiminea japonica とされている),現場で小型
Fig. 6. Salinity of the interstitial water (10-cm-deep sediment) after the tsunami (a) and changes in the silt-clay content of the sediment
(1–4 cm deep) following the tsunami (b). A reference line is placed at the same location in each map in (b). Silt-clay data before the tsunami are from literature (金谷・菊地 2011). Sampling stations are indicated with black and white dots.
個体が多く見られていることから,2011 年の秋口に多く の浮遊幼生が加入したと考えられる. オゴノリ群落も津波でほとんどが流失したが,8 月にな ると潟奥部で急速に分布域を拡大しつつあった (Fig. 5). オゴノリ藻体上に生息する 3 種のヨコエビ類も絶滅した が,回復したオゴノリ群落内においても,彼らの生息は確 認されていない.これは,底土中に生息するヨコエビ類 が,震災後に著しく増加したのと対照的である.オゴノリ は汽水環境(塩分 15∼30)で特に高い成長率を示すこと が知られており (Yokoya et al. 1999),蒲生潟では震災前と ほぼ同等の汽水環境が回復していることから,今後徐々に オゴノリ群落が回復していく可能性が高い.オゴノリを生 息基質とするヨコエビ類についても,モニタリングを継続 し,その生息状況を定期的に確認する必要がある. 環境攪乱後の底生動物群集の回復には,攪乱の程度に応 じ数ヶ月から数年を要することが知られている (Levin et
al. 1996; Whanpetch et al. 2010; Joydas et al. 2012).震災後
の蒲生潟では,コタマガイ,タマシキゴカイ(糞塊),ユ ムシやイトマキヒトデなど,一時的に移入した海産種が 12 種確認されたが,彼らの多くは夏以降みられなくなっ た.今後,汽水環境が安定して維持されれば,震災前にみ られたような汽水性の底生動物群集が徐々に回復する可能 性が高い.震災後 5 ヶ月が経過し,一部の多毛類のように 著しく増加した種が存在する一方で,多くの種(特に二枚 貝)で震災後の加入が確認されていない.一般に,攪乱後 一定の時間が経過し,底生動物群集の多様性が回復してい くと,一時的に増加した日和見種は再び減少する (Levin
et al. 1996; Joydas et al. 2012).蒲生潟においても,時間の
経過と共に底生動物群集の多様性が徐々に回復していくと 予想される.底生動物群集の中長期的な遷移過程について は,今後も継続的なモニタリングが必要である.
補 記
本稿は,仙台市の蒲生潟において,震災から 2011 年 8 月上旬までの 5 ヶ月間に行われた調査結果をまとめたもの である.調査終了時において,蒲生潟の生態系は着実に回 復しつつあると推測された.しかし,8 月中旬になると砂 の堆積により七北田川河口が完全に閉塞し,潟は約 1 ヶ月 間淡水化した.9 月 21 日には,台風 15 号の大雨で潟内の 海側砂嘴が決壊し,潟の南半分が七北田川の河道となっ た.干潟は洗掘され,海浜植物群落や潟海側のヨシ原が流 失した.2012 年 2 月には,閉塞した旧河口部の開削と導 流堤の仮復旧工事が宮城県によって行われた.3 月初めに 旧河口部が開き,潟内に形成されていた河口は漂砂の堆積 によってほどなく閉じられたため,蒲生潟は再び袋状の形 状を回復した. 謝辞:秦浩司氏には野外調査を,木下今日子氏には室内作業を お手伝い頂きました.底土の粒度分析では,(株)日本海洋生物 研究所の協力を得ました.感謝申し上げます.本稿の改訂にあた り,貴重なご助言を頂いた伊谷行博士と 2 名の匿名査読者に,篤 く御礼申し上げます.本研究の一部は,三井物産環境基金 2011 年度東日本大震災復興助成による援助を受けました.引 用 文 献
Aikins, S. and E. Kikuchi 2002. Grazing pressure by amphipods on mi-croalgae in Gamo Lagoon, Japan. Marine Ecology Progress Series, 245: 171–179.
Botter-Carvalho, M. L., P. V. V. C. Carvalho and P. J. P. Santos 2011. Recovery of macrobenthos in defaunated tropical estuarine sediments.
Marine Pollution Bulletin, 62: 1867–1876.
蒲生干潟自然再生協議会 2008. 蒲生干潟自然再生全体構想.宮城 県,52pp. http://www.pref.miyagi.jp/sizenhogo/sizen/saisei-hp/ zentaikousou2/zentaikousou2-zenbu.pdf
Grassle, J. F. and J. P. Grassle 1974. Opportunistic life histories and ge-netic systems in marine benthic polychaetes. Journal of Marine
Re-search, 32: 253–284.
Hayasaka, D., N. Shimada, H. Konno, H. Sudayama, M. Kawanishi, T. Uchida and K. Goka 2012. Floristic variation of beach vegetation caused by the 2011 Tohoku-oki tsunami in northern Tohoku, Japan.
Ecological Engineering, 44: 227–232.
Henmi, Y. and M. Kaneto 1989. Reproductive ecology of three ocypo-did crabs, I. The influence of activity differences on reproductive traits. Ecological Research, 4: 17–29.
Jaramillo, E., J. E. Dugan, D. M. Hubbard, D. Melnick, M. Manzano, C. Duarte, C. Campos and R. Sanchez 2012. Ecological implications of extreme events: footprints of the 2010 earthquake along the Chilean Coast. PLoS ONE, 7: e35348 doi:10137.
Joydas T. V., M. A. Qurban, A. Al-Suwailem, P. K. Krishnakumar, Z. Nazeer and N. A. Cali 2012. Macrobenthic community structure in the northern Saudi waters of the Gulf, 14 years after the 1991 oil spill.
Marine Pollution Bulletin, 64: 325–335.
Kanaya, G., T. Suzuki and E. Kikuchi 2011. Spatio-temporal variations in macrozoobenthic assemblage structures in a river-affected lagoon (Idoura Lagoon, Sendai Bay, Japan): Influences of freshwater inflow.
Estuarine, Coastal and Shelf Science, 92: 169–179.
金谷 弦・菊地永祐 2011. 富栄養化が汽水域の底生生態系に及ぼ す影響について.地球環境 16: 33–44.
Kanaya, G., S. Takagi, E. Nobata and E. Kikuchi 2007. Spatial dietary shift of macrozoobenthos in a brackish lagoon revealed by carbon and nitrogen stable isotope ratios. Marine Ecology Progress Series, 345: 117–127.
環境省自然環境局生物多様性センター 2007. 浅海域生態系調査 (干潟調査)業務報告書.環境省自然環境局生物多様性セン
ター,236 pp.
Kikuchi, E. and K. Yasuda 2006. Comparison of the life cycles of two sympatric estuarine polychaetes, Hediste diadroma and H. atoka (Polychaeta: Nereididae), in the Nanakita River estuary, northeastern
Japan. Limnology 7: 103–115.
Kurata, K. and E. Kikuchi 1999. Life cycle and production of
As-siminea japonica V. Martens and AngustasAs-siminea castanea
(Wester-lund) at a reed marsh in Gamo Lagoon, northern Japan. Ophelia, 50: 191–214.
栗原 康 1975. 蒲生干潟の環境保全に関する基礎的研究.宮城 県,133pp.
栗原 康・菊地永祐・上原忠保 2001. 蒲生干潟の生態学と保全手 法.蒲生干潟環境保全対策基礎調査報告書,宮城県,92pp. Levin, L. A., D. Talley and G. Thayer 1996. Succession of
macroben-金谷 弦・鈴木孝男・牧 秀明・中村泰男・宮島祐一・菊地永祐
thos in a created salt marsh. Marine Ecology Progress Series 141: 67– 82.
Lissner, J. and H.-H. Schierup 1997. Effects of salinity on the growth of Phragmites australis. Aquatic Botany 55: 247–260.
Lomovasky, B. J., F. N. Firstater, A. G. Salazar, J. Mendo and O. O. Irib-arne 2011. Macro benthic community assemblage before and after the 2007 tsunami and earthquake at Paracas Bay, Peru. Journal of Sea
Research, 65: 205–212.
Mauchamp, A. and F. Mésleard 2001. Salt tolerance in Phragmites
aus-tralis populations from coastal Mediterranean marshes. Aquatic Bota-ny, 70: 39–52.
三浦佑輔・南 將人・有働恵子・真野 明 2008. 仙台湾南部海岸 におけるヘッドランド周辺の土砂輸送解析.海岸工学論文集, 55: 571–575.
Perkol-Finkel, S. and L. Airoldi 2010. Loss and recovery potential of marine habitats: an experimental study of factors maintaining resil-ience in subtidal algal forests at the Adriatic Sea. PLoS ONE, 5: e10791. 鈴木孝男 2011. 東日本大震災による干潟環境の変化と底生動物へ の影響.水環境学会誌,34: 395–399. 鈴木孝男 2012. 東日本大震災が南三陸と仙台湾沿岸の干潟と底生 動物に与えた影響.仙台湾の水鳥を守る会(編),東日本大震災 による仙台湾沿岸湿地への影響,仙台湾の水鳥を守る会,宮城 県仙台市,pp. 3–18. 鈴木孝男・金谷 弦 2012. 大津波で撹乱された蒲生干潟は回復す るのか.震災緊急シンポジウム「津波と地震とベントス―大撹 乱のインパクト」,日本ベントス学会誌,66: 120–122.
Whanpetch, N., M. Nakaoka, H. Mukai, T. Suzuki, S. Nojima, T. Kawai and C. Aryuthaka 2010. Temporal changes in benthic communities of seagrass beds impacted by a tsunami in the Andaman Sea, Thailand.
Estuarine, Coastal and Shelf Science, 87: 246–252.
Yokoya, N. S., H. Kakita, H. Obika and T. Kitamura 1999. Effects of environmental factors and plant growth regulators on growth of the red alga Gracilaria vermiculophylla from Shikoku Island, Japan.