近畿大学学術情報リポジトリ
103
0
0
全文
図
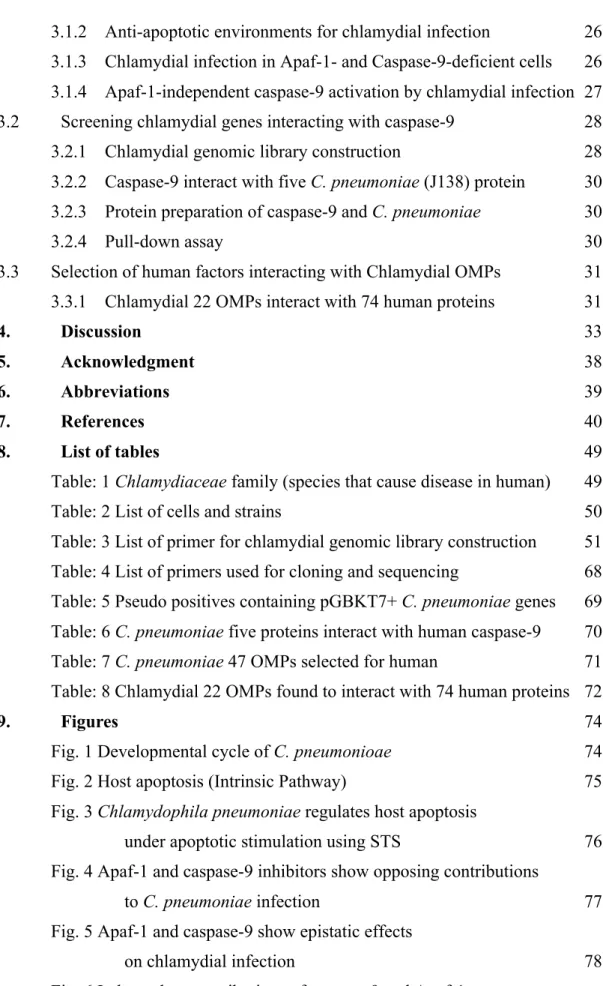


+7




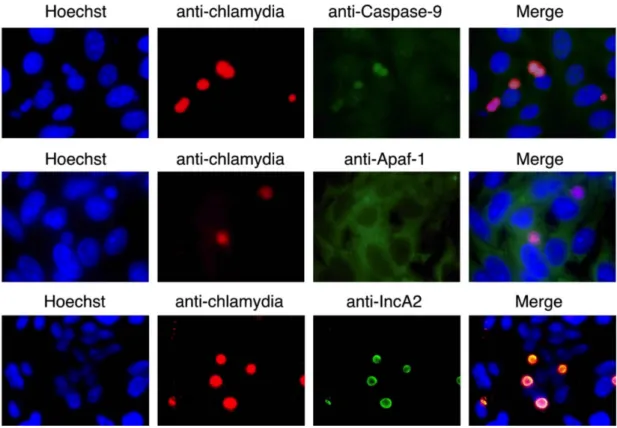

関連したドキュメント
To investigate whether defects in the SPATA17 gene are associated with azoospermia due to meiotic arrest, a mutational analysis was conducted, in which the SPATA17 coding regions
Relative expression levels of RaSTM (open) and RaYAB2 (shaded) in phylloclade primordia (PP), shoot apices and bud scales subtending them (S), floral buds (F), scale leaves (L)
Treatment with ONO-1301 increased hepatic HGF mRNA expression, but decreased the expressions of TGF-β1, connective tissue growth factor, α-smooth muscle actin, and type-I and
Transporter adaptor protein PDZK1 regulates several influx transporters (PEPT1 and OCTN2) in small intestine, and their expression on the apical membrane is diminished in pdzk1
東京大学 大学院情報理工学系研究科 数理情報学専攻. [email protected]
情報理工学研究科 情報・通信工学専攻. 2012/7/12
In [1, 2, 17], following the same strategy of [12], the authors showed a direct Carleman estimate for the backward adjoint system of the population model (1.1) and deduced its
理工学部・情報理工学部・生命科学部・薬学部 AO 英語基準入学試験【4 月入学】 国際関係学部・グローバル教養学部・情報理工学部 AO
