は じ め に
イスカ Loxia curvirostra は,スズメ目アトリ科の小鳥である.日本においては,極東の亜種 L. c. japonicaが主に冬鳥として渡来し,一部地域では繁殖している(日本鳥学会 2012).青森県の下北半 島に ₁ つ繁殖個体群(本稿では下北個体群と呼ぶ)があり,そこでは例外もあるがほぼ毎年イスカが 繁殖している(蛯名・三上 2012,2017,蛯名ら 2015).付近には大きな個体群はなく,孤立した場所 であるため,一見,局所個体群として隔離されているかのように見える. 冬鳥として日本各地に訪れる本種の越冬数は年によって大きく変動するが,2017/2018年の冬から翌 春までの間,東北地方や北海道の一部地域には非常に多くのイスカが渡来した.たとえば北海道の七 飯町では例年 ₃ ~ ₅ 月に訪れるイスカが ₁ 月から ₅ 月にかけて滞在し,群れサイズも60を超えること があった(三上かつら 個人観察)し,下北半島も例外ではなかった.繁殖個体群である下北個体群が 生息している林で多くのイスカが見られ,しかもその一部の群れは在来の個体群とは異なる鳴き声で あった.ヨーロッパのイスカでは“フライトコール”と呼ばれる地鳴きについて,局所個体群間で変 異がみられることが知られている(Förschler & Kalko 2009).このように,音声によって識別できた 由来の異なる個体群が同所的に越冬することがはっきりわかる例は興味深い.今回この下北個体群の いる林において,鳴き声の異なるイスカの群れの音声と形態のデータを得ることができた.一時的に 同居した ₂ タイプの群れの間にどのような違いがあったのかをここに報告する.青森県下北半島の松林に一時的に同居した
鳴き声の異なるイスカの形態比較
蛯名純一・三上かつら*
青森県下北半島において,2017/2018年の冬から春にかけて, ₂ タイプの声をもつイスカが一つ の林に同居していた.片方は通年生息し,繁殖もしている下北個体群のイスカ,もう片方は越冬 に訪れた繁殖地不明のイスカである.在来集団については2006年以降標識調査を行っており,計 測値を蓄積してきた.今回越冬集団と在来集団について計測値を比較したところ,翼長,尾長, ふしょ長は両者で重複が大きかったが,嘴峰長と体重は偏りがみられた.判別分析でも嘴峰長と 体重が寄与しており,在来集団は相対的に嘴が短く体重が軽い,越冬集団は嘴が長く体重が重い という特徴があった.嘴峰長の違いはおそらく繁殖地で多く利用している餌資源の違いを反映し ているだろう.越冬集団の重い体重は渡り前に体重を増加させていたためかもしれない. キーワード:青森,イスカ,形態,集団間変異,鳴き声. * 責任著者 Corresponding author2 蛯名純一・三上かつら
調査地と方法
調査地は,青森県下北半島にある約 1.8 km₂ のクロマツ Pinus thunbergii を主体とした松林である (生息地保護のため詳細は伏せる).この調査地では,イスカが通年生息しており,繁殖も確認されて いる(蛯名・三上 2012,2017,蛯名ら 2015)が,移出入がどのくらいあるかはまだわかっていない. この林に生息するイスカの個体数は50~100羽程度だが,数羽しかみられなかった2012/2013年のよう なシーズンもあった(蛯名・三上 2017).この地では鳥類標識調査を2006年以降毎年実施した.捕獲 の時期は ₄ ~11月の間で,12~ ₃ 月は繁殖状況を確認するための目視調査や撮影を行い,月に ₀ ~ ₃ 回の頻度で任意に行った.捕獲には36メッシュ 12 m のかすみ網 ₂ ~ ₄ 枚,もしくは片無双網(網の ₅ m×2 m)を用いた.捕獲した全個体に対し,通し番号が入った環境省の鳥類標識調査用の金属足輪 を装着したほか,2011年からは目視観察でも個体識別ができるよう,金属足輪に加え,プラスチック の色足輪を右足に ₁ 個,左足に ₂ 個,装着した.形態計測と写真撮影は捕獲した全ての個体に行った. 計測箇所は次の ₆ つである;自然翼長,尾長,全頭長,ふしょ長,全嘴峰長,体重.計測方法は鳥類 標識マニュアル改訂第11版(山階鳥類研究所 2009)にしたがった.計測にはデジタルノギス(Mitu-toyo CD-15PS),デジタル体重計(CUSTOM CS-240)を用い,すべて第一著者が計測した.捕獲時に は年齢,性別を次のように記録した;本種は換羽が部分的に行われ,その換羽の法則性が見出しにく い(Ginn & Melville 1983, Demongin 2016)ことから,全身が幼羽に覆われている個体に限って幼鳥 として記録し,それ以外は成鳥として記録した.今回の解析には成鳥のみを用いた.その際,幼羽の 残り方から第 ₁ 回冬羽の可能性が高いと判断された個体については解析対象から除外した.性別につ いては,本調査地では黄色い雄成鳥個体と,黄色い雌成鳥個体がみられる(蛯名・三上 2012)ため, 抱卵斑が確認され,かつ黄色い個体は雌と判定したが,抱卵斑がなく,かつ黄色い個体については主 に頭部の色が青っぽく斑があるもの(蛯名・三上 2018)を暫定的に雌として記録した.そのため,参 考として性を分けて整理したが,本論文では,性別を確定することは主題ではないので,性別に関す る考察は行わないこととした. 鳴き声の違いによって,2018年に捕獲した31羽を飛翔の際に出す地鳴きを基準(図 ₁ )に捕獲個体 の声を調査者が耳で聴き分けて「在来イスカ」,「ピンピンイスカ」,「不明」の ₃ タイプに分けた(図 ₁ ).ピンピンイスカは鳴き声の聞こえかたから名付けた本稿における便宜的な名称である.在来イス カはシラブルの前半にアクセントがあり,ピンピンイスカはシラブルの後半にアクセントがある.ア クセントが強いピークにおける最も高い周波数も両者で異なる(図1a, b).2017年以前に同地で捕獲 された個体はすべて「在来イスカ」とみなした.なお,ピンピンイスカはすべて2018年 ₄ ~ ₅ 月に捕 獲され,2018年秋以降は同地で確認されていない(2019年 ₃ 月までの観察において). 今回は両者の形態形質を比較するため,2009年~2018年の間に捕獲された成鳥個体の計測値を整理 した.各計測箇所について平均値と標準偏差は算出したが,両者を比較する検定は行わず,「在来イス カ」と「ピンピンイスカ」の違いに寄与の大きい変数を知るための判別分析を行った.これは,厳密 には両者を統計的に比較できるのが2018年度のみであり,その組み合わせでは充分なサンプル数がな いためである.判別分析のデータセットは欠測値のある ₃ 個体を除外して, ₆ 変数の値を標準化し, Rの MASS パッケージ中の lda 関数を用いて,判別関数の係数,判別率,誤判別率,判別スコアを算 出した.結 果
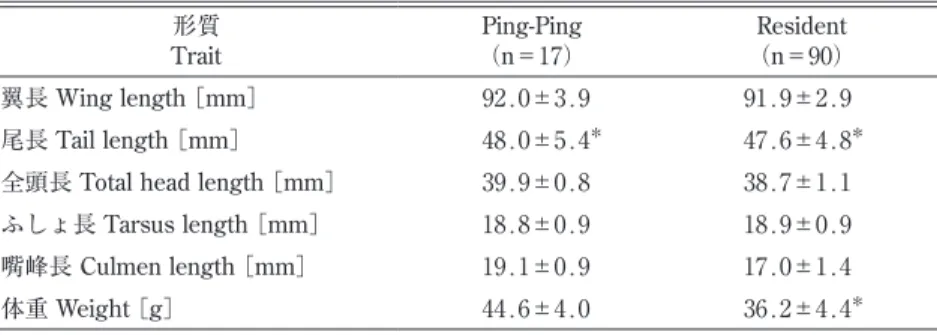
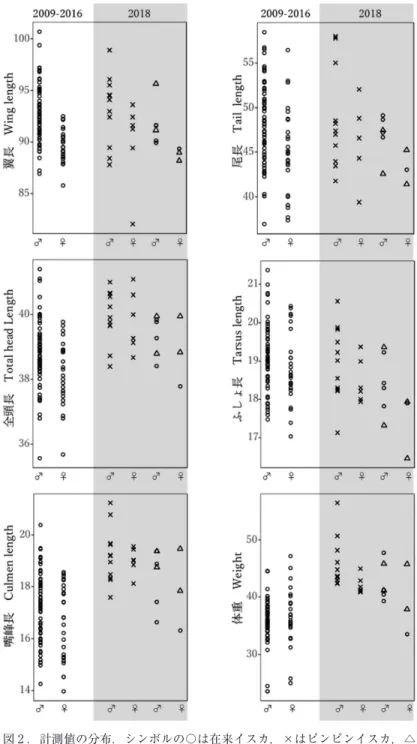
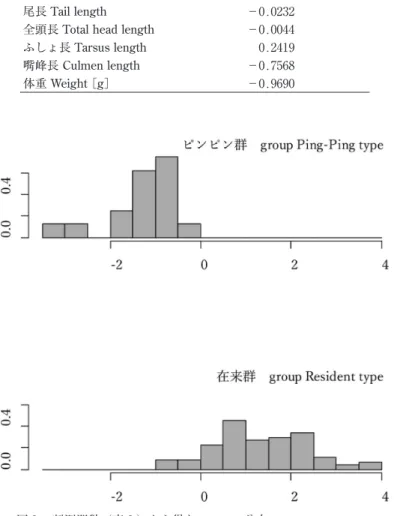
全111羽の計測値を比較した.翼長,尾長,ふしょ長では在来イスカとピンピンイスカで大きな違い はみられなかったが,全頭長,嘴峰長,体重については計測値の範囲に重複があるもののサイズ分布 にタイプごとの偏りが見られた(表 ₁ ,図 ₂ ). 欠測値を含まない104羽(在来 n=88,ピンピン n=16)のデータセットを用い,判別分析を行った (表 ₂ ).得られた線形判別係数(LD1)によると,判別寄与率の高い変数は体重と嘴峰長であった. この判別式による判別率は94.2%,誤判別率は5.8%であった.これらの係数をそれぞれの計測値に乗 じて得られた判別スコアの分布を図 ₃ に示した.考 察
翼長や尾長において在来イスカとピンピンイスカで重複が見られた(図 ₂ )一方で,嘴峰長や体重 においてはタイプによる偏りがみられた.全頭長にも偏りがみられるが,これは明らかに嘴峰長が反 図 ₁ .イスカの音声タイプ別スペクトログラム.a)在来イスカ,b)ピ ンピンイスカ.横軸は時間を,縦軸が周波数を示す.いずれも ₂ 秒間を 抽出.Fig. 1. Spectrograms of different two types Common Crossbills observed at Shimokita Peninshla, Japan. a) Resudent type, b) Ping-Ping type derived from unknown population. The horizontal axis and the vertical axis repre-sents time and frequency, respectively. Both panels show sound for ₂ seconds.
4 蛯名純一・三上かつら 映されたものだろう.また,判別分析の結果においても,明瞭に分離したスコアが得られ(図 ₃ ),判 別係数の絶対値および符号(表 ₂ )から,ピンピンイスカは「嘴が長くて体が重い」,在来イスカは 「嘴が短く体は軽い」という相対的な特徴があるといえる. 現時点でピンピンイスカがどこの地域で繁殖している個体群に由来するのかわかっていない.極東 のイスカについては,亜種や個体群の情報が不足しており,多くを考察することは難しいが,今後の ヒントを得るため,波形の似たフライトコールがあるかどうかを音声アーカイブ xeno-canto(https:// www.xeno-canto.org,閲覧日2019年 ₇ 月29日)にある ₅ 地点の音声と比較を試みた(図 ₄ ).ピンピン イスカに最も近いスペクトログラムの波形をもつのは図4c(XC268333)で,これはロシアのウスリー スクで2015年 ₅ 月21日に録音されたものである.一方,下北の在来イスカにやや近いのは図4b (XC268325)だと思われるが,シラブル後半の波形が少し異なる.なお,XC268325の録音地もウス リースクで2015年 ₄ 月28日に録音されたものであるが,XC268333とは 70 km 離れている.また, XC268333と XC268325がウスリースクで繁殖している個体かどうかは情報がないためわからない.音 声の記録を蓄積し続けてくことで極東のイスカへの理解が進むだろう. 翼長や尾長の集団内変異が大きいにもかかわらずこれらは両タイプで重複が多い一方で,嘴,体重, 飛翔の際に出す地鳴きの特徴は下北の在来イスカとピンピンイスカとの間で異なっていたのは興味深 い.嘴の形については北米(Benkman 1993, 2003)やヨーロッパ(Edelaar et al. 2012)のイスカでも 地域間変異がよく知られており,後者のスペインのイスカ(種群)Loxia curvirostra complex の例で は,嘴の形状と音声,mtDNA が違う度合いは地理的距離よりも利用している餌資源と相関が高いこ とが報告されている.これらの例と同様に,両者の主要な餌資源に依存して嘴峰長に違いが生じてい る可能性が考えられる.下北のイスカはクロマツを主に利用するが,ピンピンイスカは繁殖地で大き さや形の異なる球果をつける別のマツ類かトウヒ類を多く利用しているのかもしれない.体重のちが いは,ピンピンイスカはすべて ₄ ~ ₅ 月に捕獲されたことから,渡りをする直前に渡り用の脂肪や筋 肉を多く蓄えた状態だった可能性はあるが,胸の羽毛を除けて脂肪の蓄積量を直接確認しなかったた め,この影響の有無はよくわからない.ただし,ピンピンイスカを手に持った際,胴がまるまると太っ た個体を持っているという感触はあった(蛯名 個人観察). 鳴き声については,今回はタイプの違いの基準として扱ったため,集団内の変異として定量的に検 証できたわけではないが,両者の地鳴きの違いはヒトの耳で聞いてわかる違いである.“ピンピンイス 表 ₁ .計測値の比較.表中には平均値±標準偏差を示した. Table 1. Comparison of six measurements. (Mean ∓ SD)
形質
Trait (n=17)Ping-Ping (n=90)Resident 翼長 Wing length [mm] 92.0±3.9 91.9±2.9 尾長 Tail length [mm] 48.0±5.4* 47.6±4.8* 全頭長 Total head length [mm] 39.9±0.8 38.7±1.1 ふしょ長 Tarsus length [mm] 18.8±0.9 18.9±0.9 嘴峰長 Culmen length [mm] 19.1±0.9 17.0±1.4 体重 Weight [g] 44.6±4.0 36.2±4.4*
図 ₂ .計測値の分布.シンボルの〇は在来イスカ,×はピンピンイスカ,△ は判定不能.2018年は,ピンピンイスカと在来イスカの両方があることに 注意(グレー部分).左からピンピンイスカ雄,雌,在来イスカまたは判定 不能の雄雌.
Fig. 2. The distribution of measurements. Open circles show Resident type, crosses show Ping-Ping type, open triangles show type unknown. Note that, in 2018, PingPing-type and Resident type coexisted together (shaded area). From left to right: PingPing-type male, PingPing-type female, native or unidentified male and female.
6 蛯名純一・三上かつら カ”は本稿で便宜的に用いた呼称だが,もともと青森県の一部地域のイスカ愛好者ら(かつて青森県 にはイスカを飼育する文化があった)はこの鳴き声のイスカを“ねがっこ”と呼んで在来のイスカと 区別していた.両者はいずれも現在亜種 L. c. japonica とみなされているとはいえ,繁殖地の間で普段 交流がなく方言の違いが顕著となっているのだろう.両者の交流の様子については興味深い観察があっ た.イスカは頻繁に水を飲む習性がある.調査地内において,ピンピンイスカは,複数ある水飲み場 のうちの特定の一部の水飲み場でのみ観察されたが,在来のイスカはいずれの場所も利用していた(蛯 名 個人観察).亜種 L. c. japonica のなかでもピンピンイスカはクラスタの違いを認識して別行動して いた可能性がある.北米の個体群では同じタイプの声の個体同士でつがいになっていた例も知られて 表 ₂ .判別分析の結果.
Table 2. Results of the discriminant analysis. Traits Coefficients of linear discriminants LD1 翼長 Wing length 0.1510 尾長 Tail length -0.0232 全頭長 Total head length -0.0044 ふしょ長 Tarsus length 0.2419 嘴峰長 Culmen length -0.7568 体重 Weight [g] -0.9690
図 ₃ .判別関数(表 ₂ )から得たスコアの分布.
図 ₄ .日本およびロシアで録音されたイスカのフライトコール(音声アーカイブ xeno-canto より <https://www. xeno-canto.org>,閲覧日2019年 ₇ 月29日).いずれも ₂ 秒間を抽出.
Fig. 4. Flight calls of Loxia curvirostra recorded at in and around Japan referring from the bird sound archive xeno-canto (<https://www.xeno-canto.org/> accessed July 29, 2019). All panels show sound for ₂ seconds.
a) XC183756: Bert Jahnke, XC183756. Accessible at www.xeno-canto.org/183756. (Muraviovka Park, Russian Fed-eration)
b) XC268325: Albert Lastukhin, XC268325. Accessible at www.xeno-canto.org/268325. (Mikhaylovskiy rayon, Primor-skiy kray, Russian Federation)
c) XC268333: Albert Lastukhin, XC268333. Accessible at www.xeno-canto.org/268333. (Gornyy Khutor, Chernigov-skiy rayon, PrimorChernigov-skiy kray, Russian Federation)
d) XC286229: Peter Boesman, XC286229. Accessible at www.xeno-canto.org/286229. (Oketo Lake forest, Hokkaido, Japan)
e) XC395664: Anon Torimi, XC395664. Accessible at www.xeno-canto.org/395664. (Near Fukui Nature Conservation Center, Ono city, Fukui Prefecture, Japan)
8 蛯名純一・三上かつら いる(Groth 1993).声の違いが種分化を促してこのまま両者はより分かれていくのか,観察できてい ないだけで接触が起こって遺伝子流動が生じているのかはわからないが,種分化研究のヒントが詰まっ た材料であることは間違いない.今後も機会があるごとに観察を続けていきたい.
謝 辞
本調査地における代々の管理人の方々,また本調査にあたり様々な形でご協力いただいた古川 博, 宮 彰男,今兼四郎,佐々木秀信,高橋雅雄,廣川 茜,三上 修,上田恵介の各氏にこの場を借りてお 礼を申し上げる.引 用 文 献
Benkman, C. W. 1993. Adaptation to single resources and the evolution of crossbill (Loxia) diversity. Ecological monographs 63(₃): 305–325.
Benkman, C. W. 2003. Divergent selection drives the adaptive radiation of crossbills. Evolution 57(₅): 1176–1181. Demongin, L. 2016. Identification Guide to Birds in the Hand. Beauregard-Vendon (自費出版),フランス. 蛯名純一・三上かつら 2012.青森県下北地方におけるイスカ Loxia curvirostra の換羽と体色変化.日本鳥類標識協 会誌 24(₂): 62–71. 蛯名純一・三上かつら 2017.青森県下北地方におけるイスカ Loxia curvirostra の営巣環境と繁殖行動の季節性. Bird Research 13: S11–S17 蛯名純一・三上かつら 2018.バードリサーチ生態図鑑 イスカ.バードリサーチ ニュースレター 2018年 ₁ 月. 蛯名純一・三上かつら・仲村 昇 2015.青森県下北地方で採集されたイスカの巣の構造と巣材.山階鳥類学雑誌 47: 43–47.
Edelaar, P., Alonso, D., Lagerveld, S., Senar, J. C., & Björklund, M. 2012. Population differentiation and restricted gene flow in Spanish crossbills: not isolation-by-distance but isolation-by-ecology. Journal of Evolutionary Biology, 25(₃): 417-430.
Förschler, M. I., & Kalko, E. K. 2009. Vocal types in crossbill populations (Loxia spp.) of southwest Europe. Journal of Ornithology 150(₁): 17.
Ginn, H. B. & Melville, D. S. 1983. Molt in Birds BTO Guide 19, p. 82. Page Bros, Norwich.
Groth, J. G. 1993b. Call matching and positive assortative mating in Red Crossbills. Auk 110: 398–401. Howell, S.N.G. 2010. Molt in North American Birds. Houghton Mifflin Harcourt, Boston.
日本鳥学会 2012.日本鳥類目録改訂第 ₇ 版.日本鳥学会,三田.
山階鳥類研究所 2009.鳥類標識マニュアル改訂第11版 2008年度版.山階鳥類研究所,我孫子. 蛯名純一₁
〒039-2241 青森県八戸市大字市川町字桔梗野上19-137 NPO 法人おおせっからんど Jun-ichi Ebina₁
NPO Oosekka-land, 19-137 Kikyouno-kami, Ichikawa-machi, Hachinohe, Aomori 039-2241, Japan. 三上かつら₂*
〒183-0034 東京都府中市住吉町1-29-9 NPO 法人バードリサーチ Katsura Mikami₂* [email protected]
Japan Bird Research Association, 1-29-9 Sumiyoshi, Fuchu, Tokyo 183-0034, Japan.
ABSTRACT
Comparison of morphological traits of between two types
Common Crossbills with different calls that were observed in the
same pine forest in Shimokita Peninsula, northern Japan
Jun-ichi EBINA₁ and Katsura MIKAMI₂*
In Shimokita Peninsula, Aomori Prefecture, northern Japan, the two types of Common Crossbills with different flight calls that lived temporarily in a pine forest from winter to spring in 2017/2018. One is Crossbills of the Shimokita population that inhabits all year round and reproduces there (we call them 'Resident type'), and the other is visitor from the unknown place ('Ping-Ping type'). The resident type has been observed and ringed since 2006, we are accumulating their morphological data. Compared six morphological traits between 16 Ping-Ping type birds and 88 Residents, culmen length and weight were biased. Linear discriminant analysis also showed contributions of the two traits to separate two types. Resident type crossbills had relatively shorter culmen and light weight, whereas Ping-Ping type had longer culmen and heavy weight. The difference of culmen would be reflect to the difference regular diet, i.e. cone of conifer species in breeding site. Heavy weight of Ping-Ping types might be cause fat accumulation and building muscle in pre-migration.
Key words: Common Crossbill, breeding population, Loxia curvirostra japonica, morphometric variation, variation of call, wintering
受付日:2019年 ₄ 月12日,受理日:2019年 ₇ 月29日,Received: 12 April 2019, Accepted: 29 July 2019 *Corresponding author: E-mail: [email protected]