贋黙蔑,第繍,薦骨)
〔特別講演〕
レンサ球菌の遺伝的変異と臨床との
関連についての考察
東京女子医科大学微生物学教室教授 吉 岡 守 正
ヨシ オカ モリ マサ (受付 昭和49年9月30日)Genetic Changes of Streptococci and Their R.elation
to Streptococcal Infections
Morimasa YOSH10KA
DepartInent of Microbiology, Tokyo Women,s Medical College
Streptococci are class三星cd into groups disregarding of hemolytic properties on the blood agar medium.
Three scries of experimellts are described, in which distinct genet五cal methods were applied on the O「gamsms・
1,Group A strain T l 2 cells were treated with mitomンcin C and nitrosoguanidine, and some of the drug−resistant mutants showed changes in many aspects including loss ofβhemolysis.
2.Group H stra三n Challis cells were separated into Challisβand Challis㏄substrains. The latter, in contrast to the R)rmer, could not transfbrm and also did not produce competence provoking魚ctor. The hemolytic marker was transmitted with DNA independent of the drug resistant marker.
3.Group D strains, which produced bacterioc三n, was co功ugated with bacter三〇cin−negative strains, and transmitted the bacteriocin−producing as well asβhemolytic propertics.
Care血l bacterial examination on the specimens obtained飾om various streptococcal i11飴ctions
recommended.
A群レンサ球菌8彦吻伽。側∫.妙ρg8η6∫1ま申すまでもなく,溶血レンサ球菌(溶レン菌)の代表的
な菌である.病原細菌学の歴史の当初に発見され
たレンサ球菌は,近年まで,血液寒天培地上の集
落の周囲に生じる血液の変化によって,β溶血,
α溶血,非溶血などと区別され,菌名としてε〃β・
ρ’0‘066雛加祝0り‘伽5,3〃ψ‘0‘0‘σπ50癖4απ5などと呼ばれていた.しかし多数の所見が積み重ねられる
に従って,レンサ球菌属の種がふえるとともに,
抽出抗原による沈降反応を用いた群分類法が導
入され12),同じβ溶血を示すレンサ球菌にもA群
のほかB,C, D, G群などが次つぎと確立され
た.また緑レン菌と称されたα溶血菌も,順次群
のなかに加えられてきている.
A群レンサ球菌3彦rψ’o‘o蜘5砂088脇はヒトをお
もな宿主とす.る病原菌であるが,本菌が原因で
あるとされる疾病の種類は,他のグラム陽性球菌
だけでなく,グラム陰性菌にも見られない程多彩
である.その第1はブドウ球菌や肺炎球菌などと
同様に化膿性または滲出炎性疾病を起こす.扁桃
炎,フルンケル,カルブンケル,イソペチゴその
他の化膿症はこの分類に含まれる.第2は,本管
の産生する外毒素である発赤毒によって全身に発
疹を生ずる狸紅熱である.狸紅熱が未だに法定伝
染病として取扱われていることは時代遅れの観が
あるが,狸紅熱そのものが伝染するのではなく
て,A群レンサ球菌による扁桃炎などが伝染し,
その第二次症として狸紅熱が起きるものである.
第3は丹毒で,上記滲出炎性ならび毒素性の混合
型と理解される.第4は,急性腎炎とリウマチ熱
である.両疾病とも本署の感染,とくに扁桃炎を
前駆症として,暫時の潜伏期を経て発病するもの
で,ストレプトリジン0抗体価など本菌の産生物
に対する抗体価の上昇と密接な関係があるにも拘
らず,腎にも心弁膜にも本菌を証明でぎない.こ
れら両疾病の発病機序は未だに確立されていない
が,生化学的活性が多様で,且つ大である半平が
病巣の組織細胞に種々の変性を生ぜしめ,宿主
生体に免疫学的応答を呼び喚こすとの考え方があ
る.この際の抗原としては菌自身,組織の自己抗
原,あるいは両者の結合物,さらに抗原抗体結合
物が可能性として挙げられるが,定説はない現状
である.A群レンサ球菌感染症が多彩であり,その代謝
産生物が多様であるという特徴のほかに,本菌の
化学療法剤に対する感受性スペクトルの安定性が
ある。ドマックのサルファ剤,フレミングのペニ
シリン(PC)発見は化学療法剤開拓のきっかけ
となったわけであるが,嘗胆感染症から分離され
る菌は,未だにこれら薬剤に対して高い感受性を
示し,耐性菌は見付かつていない。同じグラム陽
性化膿球菌であるブドウ球菌の単剤および多剤耐
性菌が医療上の障害となっている事実,また赤痢
菌,結核菌が次つぎと新らしい抗生物質に耐性を
獲得したといった傾向とは趣きを異にするA群レ
ンサ球菌のサルファ剤,PC感受性の安定性は,
いかなる理由によるのであろうか.やはりグラム
陽性化膿球菌である肺炎球菌も,A群レンサ球菌
と同じ様にこれらの薬剤感受性パターンを変えて
いない.私達はレンサ球菌のうちA,HおよびD群菌につ
いて,それぞれ相異つた遺伝学的手法を用いて変
異を起こさせ,レンサ球菌変異に関与する因子を
追究している.まずその内容を紹介し,終りに考
察を加えたい. 1.A群レンサ球菌の薬剤耐性変異5)24)∼27)現在ヒトから分離されるA群レンサ球菌のうち
薬剤耐性菌は,テトラサイクリン(TC)耐性と
エリスロマイシン耐性13)が知られている.エリス
ロマイシン耐性菌は同じマクロライド系薬剤であ
るリンコマイシンとオレアンドマイシンにも耐性
を示すほか,ときにTCおよびクロルアソフェニ
コール(CM)にも交叉耐性のあることが報告さ
れている16)21).私達は約10年前にA群レンサ球菌を用いて,試
験管内で薬剤耐性菌を採取する研究に着手した.
新鮮分離株および研究室保存株を,数種の薬剤を
含む寒天培地を継代し,順次薬剤の濃度を高める
方法を採ったが,頻回植継いでも耐性変異株をう
ることは他の細菌に比べて容易ではなかった.ス
トレプトマイシン(SM)およびTCに対しては
親株の10倍またはそれ以下の耐性獲得に過ぎず,
耐性株の諸性状は親株のそれらとほとんど変りな
く,僅かにマウスに対する病原性の低下を認めた
程度であった.バシトラシンに対しては耐性菌を
えられなかった.ところが,PCとマイトマイシ
ンC(MC)に対しては100倍から1000倍の耐性
変異株をうることができた.PC耐性変異が試験
管内で行われることは松井14),Rosenda120), Mic−haelら15)の報告がすでにあるが,彼らの耐性菌の
性状と私達のえた薗の性状とは若干異っていた.
私達のえたPC耐性菌とMC耐性菌の性状は表
1,2に示すように,本来のA群レンサ球菌のそ
れとは極めて異っているぽかりでなく,両薬剤に
共通耐性を示す。ウマ血液寒天培地上の溶血はβ
溶血でなくα溶血に変り,溶血素であるStrepto。
Iysin−S(S正S)も一〇(SLO)も産生せず,抗原性およびマウス病原性も著しく減弱している.PC耐性
菌はpenicillinaseのような不活化物質を見出せな
かったが,MC耐性はつねにMC不活化作用を伴
一56一
Table 1. Biological and biochemical characteristics of group A
stτeptococci and their antibiotic resistant mutants
Hemolysis on blood aga「
MinimaUethal
dosea) 10n Antigenicity GroupType
Streptokinase 1ヨ【yaluronidaseH202
0−Tolidine method Iodine method Streptolysin O Streptolysin S Supernatant of broth culture By washed O「ganlsmsT12
β一8
什 十 十 mg/μgN O.720 HU/μgNb) 0.49 13.47 0.97S231K・
β β一9
十 冊 十 0.763 8.33 15.36 1.80一7
? 十 十 2.800 4.13 23.63 1.52 SM−Ka β一2
十 0.840 0.33 5.38 1.20 TC.Ka β一2
四 十 0.275 0.25 2.48 1.09 PC−T12 α 0 十 ± 4.261 0 0 0 PC−Ka α 0 十 4.971 0 0 0MC−T12
α 0 十 ± 4.900 0 0 0 MC−S23 α 0 十 3.115 0 0 0MC−Ka
α 0 4.381 0 0 0a)Minimal lethal dose is expressed as the dilution rate of the beef heart infusion・glucose
broth culture,
b)Fifty per cent hemolytic unit is expressed perμg nitrogen of bacteria contained in a sample of broth culture.
つていることがわかった.ただし,今までのとこ
ろMC不活化物質を菌から遊離せしめることには
成功していない.上述のように,PC高度耐性株はもはやA群レ
ンサ球菌の特徴を完全に失っていて,その性状は
むしろD群レンサ球菌のそれに類似している.そ
こでこのような大幅な変異の過程において,中間
の性状を有する変異が存在するのではないかと考
えて,PCおよびMCを用いて継代毎に慎重な観
察を行なった.PCでは所期の目的を達しえなか
ちたが,MCを用いて次のような結果がえられ
た、菌は国際標準株A群12型のT12株を使用した。
本菌の増殖を抑えない程度の低濃度(0.01μg/ml)
のMCを加えたTodd−Hewittブイヨン中で37℃18
時間培養したT12株を, M Cを含まないTodd・
Hewitt寒天平板に植え,生じた集落を些細に観察
し,親株の菌集落と外観の等しいものを拾って再
び新しいMCブイヨンに継代した.継代を反復す
るに際しては,MCの濃度を菌増殖を抑制しない
範囲で徐々に増加せしめた。また平板上に外観の
異る集落を見出したとき,その菌から出発して別
の継代シリーズとして,同じようにMCブイヨン
継代を反復した.図1はこの実験でえられた8株
の採取順序を示す。これら8株の薬剤感受性は表3に見るように,
Mcの最小阻止濃度(MIc)はQ.05−10μg/ml,
PCは0.01∼10u/ml,パシトラシンは0.25−5u/ml,TL2−Tし3一「一TL3−2
嘩ぐ一一一一
MCTI2−1【→MCTI2一皿Fig・1。 Isolating Process of variants during serial transfers in Todd−Hewitt broth containing incre.
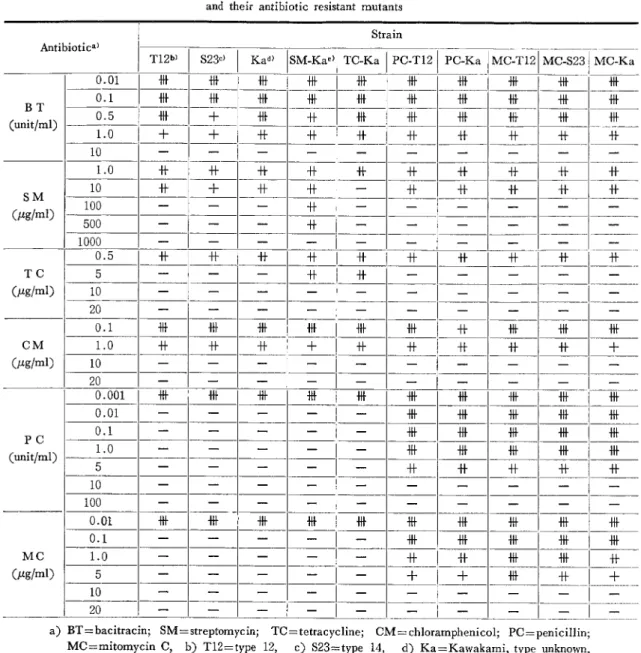
Table 2。 Antibiotic sensitivity patterns of group A streptococci
and their antibiotic resistant mutants
Strain Antibiotica)
T12b) S23・} Kad) SM.Ka・)
TC.Ka ・σT12
P
PC−KaMC−T12
MC−S23 MC−KaBT
(un五t/ml)SM
(μ9/ml)TC
(μ9/m1)CM
(μ9/ml)PC
(unit/ml)MC
(μ9/ml) 0.01 帯 帯 惜 0.1 0.5 十 帯 什LO
十 十 井 什 什 什 10LO
井 什 山 川 什 10 什 十 什 什 卦 什 什 100 500 十 1000 0.5 十 什 掛 帯 5 升 10 20 0.1 柵 冊 柵 十 帯 1.0 十 十 10 20 0.001 帯 冊 冊 帯 下 0.01 0.1 冊 帯 帯LO
惜 5 什 10 100 0.01 帯 帯 帯 柵 帯 0.1 柵 1.0 凹 凹 5 十 十 四 十 10 20 a)BT=bacitracin;SM=streptomycin;TC=tetracycline;CM=chloramphenicol;PC=penicillin; MC=mitomycin C, b)T12=type 12, c)S23=type 14, d)Ka=Kawakami, type unknown, e)SM−Ka=SM resistant Ka mutant strain.SMは50−300μg/mlに分布しているが, T Cお
よびCMについては株間に差を認め難い.また,
一般性状は表4に見るようにTL−2およびTL
5b−3株は6.5%NaClおよびpH g.6の液体培
地で増殖する点を除いて,胆汁酸および温度感受
性,SLSおよびS正0,それにDNase産生に
ついて親株T12と同じである.ただTL5b−3
株は1%ブドウ糖加ウマ血液寒天培地および5劣
ヒツジ血液加寒天培地上すべてβ溶血を呈すた
め,小林分類は皿型に属する.因みに小林1型は
1%ブドウ糖加血液寒天上ではα溶血を示すもの
である.T正3−2株は上記2株とほぼ等しい一
一58一
Table 3。 Minimum inhibitory concentrations of antibiotics for the MC.treated variants Antibiotica) MC (μ9/ml) pc (unit/ml) BT (unit/m1) TC (μ9/ml) CM (μ9/1nl) SM (μ9/ml) Strain 1st Groupb) TI.一2 TI.3−2 0.05 0.05 0.05 0.05 0.25 0.25 1。0 1.0
5 5
50 50
TL
5b.3 0.1 0.01 0.5 5 5 50 2nd Groupb》TL
5b.1 0.5 0.05 2.5 5 5 50,脚譜、
0.5 0.1 2.5 5 5 50 0.5 0.01 0.25 5 10 50 3rd Groupb)MCT
12−H 5 0.1 5 1.0 10 100MCT
12一皿 10 10 5 1.0 10 300T12
0.05 0.005 0.25 1.0 5 5PCT
12c) 10 100 25 10 5 50 GroupC
9542 0.05 0。01 2.5 1.0 5 50 GroupD
4532 0.5 5 5 5 5 50 GroupD
2025 0.5 5 10 10 20 200 a)MC=m五tomycin C;PC=penicill三n G;BT=bacitracin;TC=tetracylcine hydrochloride;CM=chloramphenicol;and SM=dihydrostreptomycin.
b)Variants were originated from 5’r8μ0606‘π5」妙og6η8∫strain T12(type 12). c)PCT12 is anα.hemolytic PC.resistant mutant obtained from strain T12.
Table 4. Physical, hemolytic and metabolic properties of MC−treated variants
Growth in or ai:… P0%bile
40%bile
6.5%NaCl
pH 9.6
10C
45C
Hemolysis on blood agar
Kobayashi’s criterion Production of: S工Sc) SLOd) DNasee) Strain TL−2 1st Group 驚 ぢ 帯 β 1 帯 帯 翁 ど く 十 β 1 TL3.2 も 自自 十 冊 帯 β 1 冊 ど く 回 雪 帯 β 1 叩 露 井 β H 2nd Group の 貞ゐ 臼ゆ 研 磨 柵 十 十 β 1 き
下
帯 β 1 の 日ゐ トゆ 十 β H 3rd Group 臼目OI
芝鯉 帯 帯 旧 訓 β 豆 井 E−1国Ol
Σ臼 十 β 皿 臼 β 1 田 。 掛 帯 帯 帯 αo
号 90
晶 帯 帯 β H 帯 十 ∩ 含 90
帯 柵 β 皿 ∩ 号 90
α a)Bef.==before subcultivations,b)Aft.=after 10 serial subcultivations on Todd・Hewitt agar. c) SI・S=streptolysin S。 d) SI・O=streptolysin O, e)DNase=deoxyribonuclease。
般性状であるが,10%胆汁酸には抵抗性を示し
た.新鮮分離A群レソサ球菌のなかには10%胆汁
酸抗抗性の株はしぼしぼ見られるところである.
TL5b−1,TL5b−WおよびT工5b−Sの
3株は,上記の培養諸条件のいずれに対しても抵
抗性を示し,またSLS, S正0, DNaseは非産生であ
った.MCT12一皿およびMCT12一皿株もこれ
ら3株と似た性状であるが,MCT12一皿株には
弱いながらSLSとSLOの産生を認めた.MC
T12一皿株はヒツジ血液寒天上α溶血を呈する小
林皿型であった.以上の結果から8株を3群に分けてみた.第1
群はTL−2, TL3−2およびTL5b−3の
3株;第2群はTL5b−1,TL5b・Wおよび
4Table 5. Precipitin reactions of formamide extracts of MC−treated variant cell walls Antigen
Antiserum
≠№@ハnst:
1st Group 2nd Group 3rd Gr・up
㌣目 『雪臼 ○う Pゐ日LΩ 一坙レE唄LQ 津日ゐi一司ゆ の 冾薰d−6LQ ト目Q l
駐c
卜司目Q πー臼
巽臼 目』o店 N昌Σ○ 国自・.雪 ゚き。日o自3等δ8 qq=5cqoeo』 ば)o寸 q旨き鵠6自
竃舘
T12
柵 冊 柵 一 惜
一 一 一 十 一 十 一 一 一 冊 帯冊 帯
一 一 一 惜 一 一 一 一 一 一PCT12
CMT12
Group B I・ewis 一 一 Group C 9542 一 一 一 一 一 一 帯 一 一 一 一 Group D 4532 一 一 一 一 一 一 一 帯 一 升卜 一 Group D 2025 一 一 一 十 一 卦 一 一 一 一 帯 一 一NDa)一 冊 一ND
Group G G16一 一1一
一 一 一 什 一ND 一 一 一 冊
a)Nat doneTL5b−Sの3株;第3群はMCT12−Hおよび
MCT12一一皿の2株である,これらの菌から抽出
した沈降原を各種群抗血清と合わせてみると,表
5のように,A群抗血清と反応するのは第1群す
べてと第2群のTL5b・W株のみであって,同群
の他の2株はA,B, C, DおよびG群抗血清の
いずれとも反応せず,高度PC耐性変異株である
PCT12抗血清と弱い反応を示すのみであった.
第3群の2株はCM耐性変異株CMT12抗血清と
強く反応すると共に,MCT12一∬株はC群抗血
清と,そしてMCT12一皿株はD群のうちの3〃6一
μo‘o卿5!砒α」6∫var・1ゆψ碗η5抗血清とも強く反応した.これらの事実を総合すると,A群レン
サ球菌の薬剤耐性変異は必ずしもβ溶血の消失を
伴うものではないこと;その際のβ溶血はSLS
およびS■0の産生がなくても起こること;変異
に伴って恐らく溶体表面の抗原決定基が変り,既
知の他の群の抗原と共通する場合と,変異菌独自
の抗原が生ずる場合のあることなどが分つた.
ところでKrauseとMcCartylo)11)によれば,
A群およびC群レソサ球菌の群抗原決定基は,細
胞壁構造中のN−acetylhexosamineにあり, A群
ではそれがglucosamine, C群ではgalactosamine
とされている.一方,D群抗原はElliott3), JonesとShattockg)らによってglycerol teichoic acid
であり,細胞壁とプロトプラスト膜の問に存在す
ると考えられている22).teichoic acidはribitol
phosphateのポリマーである.表6はガスクロマト
グラフィーによる糖および脂肪酸の分析結果であ
る.第1群の2株を除く6株にgalactosamineを
認め,とくにC群抗血清に反応したMCT12一皿
株はC群標準株と同様に60%に達する高い含量を
示した.脂肪酸については,第2および第3群で
は炭素数18以上の幾つかの成分が余分に見られ
た.親株には見出されないgalactosamineが変異株
に現われる点について,枯草菌やサルモネラで証
明されているuridine diphosphate−N−acetylgluco− samine−4・epimerase,すなわちUDP−N−acetylglucos−amineに作用してUDP−N−acetylgalactosamineに
変える酵素の有無を調べたのが表7である.変異
株には本酵素があり,A群標準株には認められな
かった.私達が初めに予想したA群レンサ球菌の段階的
変異は存在しうると考えられ,変異の方向として
はA群がらCまたはD群に類似の性状を備えるよ
うな代謝の営みや細胞壁の構造へ変るために,菌
の環境に対する抵抗性が変ってくるのではないで
あろうか.しかしどのくらいの単位の遺伝子が変
異に関与しているのか不明であり,また前述の
一.U0一
Table 6. Sugar and fatty acid compositions of MC.treated variant cell walls 皆 8 b 巴 持 曽 の お ゆ臼 上 巳 膨 器 わ 罵 隣
Rhamnose
Glucose Galactose Glucosamine Galactosamine 12:0 14:0 16:0 16:1 18 0 18:1 18:2X1
X2
X3
X4
Strain 1st Group TL−2 56.4 43.5 2 9 正00 62 20 82 1 T工3.2 32.2 21.3 4.4 20.2 21.8 ! 2 100 62 24 101 1X5
1
一i_
T工 5b−3 58.0 2.3 39.6 1 3 100 62 21 63 1 2nd Group TI. 5b−1 TI. 5b.WTL
5b−S 3rd Group 25.2 17.5 7.9 29.9 19.2 10 100 39 22 203 2 trace 57 47 5 26.7 23.6 11.1 22.2 15.2 12 100 38 17 166 10 trace 55 48 11 7MCT
12一皿 27.8 35.8 22.2 一 20.3 一 16.1 3.7 13.5 60.4 8 100 28 26 148 4 trace 17 31 4 9 1 6 100 11 30 65 1 15 17 3 1MCT
12一皿T12
13.5 61.2 18.2 一 28.3 一 !2.7 38.7 27.0 9 100 17 39 66 2 5 5 2 4 100 78 19 111 1PCT
12 32.6 43.5 21.6 17 100 29 6 130 7 trace 34 35 6CMT
12 27.2 22.2 29.7 12.2 8.8ND
ND
ND
ND
ND
ND
ND
ND
ND
ND
ND
ND
GroupC
9542 39.7 trace 3.2 56.5 2 5 100 54 32 111 1 GroupD
4532 25.5 27.8 27.8 5.6 13.2 9 100 17 31 90 4 4 GroupD
2025ND
ND
ND
ND
ND
11 100 13 43 77 trace !5 11 2一:Not detected. Neutral and amino sugars were detected by gas chromatography after treating the cell wall samples with triHuoroacetic anhydrate. Fatty acids extracted from cell waU samples were determined as methyl esters by gas chromatography too。 Relative quantities
to the C16.compound content are presented。
Table 7. Uridine diphosphate.ハ乙acetylglucosamine・4−epimerase activity of crude ce11 extracts
Hexosamine
detected Glucosamine Galactosamine Extract of : T五5b.1 14.5μg 15。0 MCT12一皿 15.1 14.4 MCT12.皿 18.4 11.2T12
30.0 0 Richards 30.0 0The substrate,30μg of UDP−N」acetylglucosamine, was reacted with the crude extract prepared from bacter且al cells for 60 min at 37 C. Hexosamines were detected by gas chromatography after acid hydrolysis.
ep量merase活性は新らしく生じたものか,それとも
構造遺伝子は元来有しているが,調節遺伝子の抑
制が解けて作られるようになったものかなどの点
について,多くの課題を提供.している.
MCはPCと違って変異誘導性があるので,私
達の実験で自然突然変異でなく誘導変異を行なっ
ている可能性が大である.そこで別の変異誘導剤
としてnitrosoguanidineを選び, T 12株に作用させMC, SMおよびPC耐性菌の出現を調べたとこ
ろ,表8に示すように,MCには100倍, SMに
ば10倍以上耐性株がえられた.現在までのとこ
ろ,菌分裂が行われずに核の改造が行われる成績
をえていないので,今後nitrosoguanidine, MC,その他の変異誘導剤を用いての核ゆさぶりの実験か
Table 8. MC, SM, and PC sensitivlties of nitrosoguanidine(NG).treated variants Growth on antibiot五。−agar Strain NG sreat− 高?獅狽 mo. of Rubcu− 撃狽浮窒? At iμ9/ml)
Medium
唐?撃?モ狽?п iμ9/ml) MC(μ9/ml) SM(μ9/ml) . 一 一 幽 一 一 一 『 L 」 oC(u/ml) 0.01 0,1 1.0 5.0 10 50 100 1000 0.001 0.0正 0.1 1,0 T12*1 1
25 2 23 2
25MC
504 2
50 (0・1)冊
5 2 100
6 2
257 3
SM
50 帯
(30) 25 帯8 2
9 3
50 帯 10 2 100 冊 惜 十 十 一 ± 一 十 一 十 一 十 一 十 一 十 一 帯 一 帯 一 辮 一 冊 一 耕 什 一 冊 一 面 一 三 一 一 一 帯 一 帯 一 帯 筋 宰 相 冊 柵 惜 什 什 什*T12 is the original strain, after NG−treatment the variants were selected on MC−or SM−agar plates.
ら,表現型変異と関連する遺伝子型の存在を明ら
かにしたいと考えている.
2.H群レンサ球菌の(形質)転換transfor・
mation7)8)細菌から抽出したDNAを他の生菌にとりこま
せて表現型を一部変える遺伝学的手法を転換(形
質転換)と呼ぶ.1928年GriHith4)が肺炎球菌で
本現象発見のきっかけを作ってから,髄膜炎菌,
インフルエンザ菌,枯草菌などでも観察された.
レンサ球菌に関しては,1957年Braccoらi)が
3∫7ψ∫06066鋸5α伽〃歪鋸∫のDNAを用いて緑レン
菌と肺炎球菌にSM耐性を転換ぜしめた.また
Pakulaら18)19)もSM耐性を肺炎球菌から緑レン
菌,H群レンサ球菌,肺炎球菌に,ブドウ球菌か
ら3彦砂伽α,6〃∫∫加およびH群レンサ球菌に伝達
した.彼らが見出したH群レンサ球菌Challis株
は転換率が大きいので,各国の研究者が使用して
いる.私達もChallis株の分与を受けてウマ血液寒天
平板上にβ溶血を起こす集落のほかに,α溶血を
示す集落を見出した.おもにヒッジー血液を用い
る外国では気付かれなかったものと思われる。
Challis一βおよびChallis一α株の一般性状はほとんど差はないが,抽出DNAの受容菌recipientとし
て用いた場合,Challis一β株は従来の報告通り受
容菌となりうるが,Challis一α株はなりえないと
いう差のあることがわかった、表9にChallis・α
株のSM, PC,MCおよびTC耐性菌からそれ
ぞれDNAを抽出し,三原株とそれらの薬剤耐性
株を受容菌とした時の転換率を示す.β原株は最
も高率に耐性遺伝子をとりこみ,とくにSM耐性
に至っては約1/80の割合であった.α原株とその
PC耐性, MC耐性株はいずれも転換を起こさな
かったが,そのSM耐性株は低率ながらPC, T
C,MC耐性変異を生じ,α原株とは異なって受
容菌となりえた.三原株から2剤耐性株を作って
受容菌として用いると,α系ではSM耐性のある
場合と,SM単独耐性の時と同様の率で転換し,
その率はまたβ系のSM耐性株の場合とも近似
していた.このことはChallis・βならびに一α二丁
がSM耐性を獲得することによって,受容菌とし
ては類似の態度をとるようにそれぞれ変ったもの
と考えられる.またPC−SM, MC−SMのような2剤耐性菌の
DNAを用いるとき,単独耐性の転換率よりは低
いながら両剤耐性が伝達されることを示してい
一62一
TaUe 9. Transformation rate in per cent of ChaHisα, Challisβ and their antibiotic−resistant mutants
DNA
?窒盾
Transformants, ■ reSlstant tO l Recipient straina)
@Challisα . . . .
「晦
・σ副
Mα・1・σS晦 . 「 } lC・SM一α 一 1 SM一αSM
0 _b) 0 0 PC一αPC
一〇 MC一αMC
0 TC一αTC
0 PC−SM.αPC
0SM
0 MC−SM一αMC
0 0.00097 0 0.00066 0 0.00052 0 0 0.00005 0 0 0 0.00026 0SM
q
0 0 0.00025 0.00516 0.00051 0.00022 N.D.c) N.D. Recipient straind) Challisβ SM一α SM PC一αPC
MC一αMC
TC一αTC
PC−SM一αPC
SM
PC−SM
MC・SM一αMC
SM
MC−SM
3.35 0.0014 0.033!8 0.00617 1.13 3.38 0.02 0.12 4.87 0,0005 SM一β 0.00017 0.0002 0.00037 0.00018 0.00004 PC一β 0.959 0.00096 0.0123 1.93 0.52 1.62 N.D. MC一β 0.533 0.00005 0.00005 0.013 0.62LO3
PC−SM一β 0.00065 0.00084 ND。 MC−SM一β 0.00016 0.00007 N.D.a)CPF was added to the transformation system. b)一lImperformable.
d)ET2 medium was used to the transformation system.
c)N.D.:Not done.
る.
ところで,H群レンサ球1菌の転換系にはcompe−
tence provok量ng factor(CPF)がないとDNAの
取りこみは行われない.Challis一β株が受容菌にな
りうるのは培養液中にCPFが作られるためであ
る.Wicky株はそれ自身CPFを産生しないが,
他からCPFが与えられれば受容菌になりうるこ
とが知られている.そこでWicky株を受容菌と
して,.種々のCha11is二二の菌の培養液から菌を
除いた濾液を加えてSM耐性の転換を行なった.
表10の如く,Challis一β系諸株はすべてCPF陽性
であるのに対して,SM耐性のChallis一α株は二
二と同様にCPF陰性であった.したがってこの
点でSM耐性のChallis一β株とは異なるわけであ
る.受容菌となりうるもの,およびなりえないも
Teble 10・CPF activity of Challisα, Cbailisβ and their antibiotic.resistant mutant for Wicky strain
CPF from Challisα SM一α Challisβ SM一β PC一β MC一β TC一β PC−SM一β MC−SM一β Without Transformantsa) (%) 0 0 4。60
L70
0.264 0.26 0.51 0.127 0.33 0 a)SM・αDNA donor applied to Wicky recipient cells.Table 11. Transformation ofα一andβ一hemolysis DNA from α一Hem・lytic β一Hemolytic Challis α SM一α PC−SM一α
wicky
SM−Wicky
Challisβ SM一β Challisβ SM一β Rec1Plents Transformants Cha1正sβ Challisβ Challisβ Chall三sβChaUisβ
Cpallis ab) Challis αWicky
Wicky
α 0.428.i%) α O.154SM
0.251 α 0.642SM
0.562 周目 0.0075PC−SMO.0001
α 0.530 α 0.810SM
0.607 β 0.272 β 0.667SM
0 3 0.299 β 0.247SM
0.438 Transformants replicated on BAa) (%)α0.00278
α 0.008α0。00016
α0.000001
α0.00236
β N.D.c) a)BA:horse blood agar plate, on which SM−resistant transformants were imprinted by areplica−plating method. b)CPF was added. c)N.D.=Not done.
ののDNAとりこみに差があるかどうかを,32Pで
標識したDNAを用いてカウントしてみたが,
Challis一α株が一β株の約3/4と幾分低値ではあるが,転換しない原因としてDNAのとりこみがないと
は言えない結果であった.
Cha11is・βおよび一α株のマーカーである溶血
性がDNAによって伝達されるか調べた成績を
表11に示す.ChalHs・β株を受容菌とするとき,
Challis一αおよびWichky株のDNAはα溶血性を
0.4∼0.8%の頻度で伝達し,そのlocusはSM
耐性のIocusと異なるものと思われる.逆にCha−
11is一αおよびWicky株を受容菌とし, S M耐性の
Challisβ株のDNAを用いるとき, S M耐性は
伝達されないのにβ溶血は0.2∼0.6%の頻度で
伝達された.なお転換菌がα溶血を獲得すると同
時にCPF非産生性を伴なったのに対して,β溶
血を獲得した転換菌にはCPF非産生性のままの
ものがみられ,溶血性マーカーとCPF産生マー
カーが必ずしも平行するものではないことを示唆
している.またChallis一α株のDNA伝達による
α溶血転換菌は受容菌となりえない (nontransfo一rmable)のに対して, Wicky株のDNA伝達によ
るα溶.血転換菌はCPFを加えれば転換を起こ
す(transformable)ことから, transformabilityのlocusはα溶血の10cusと極めて近いか,あるい
は等しいのではないかと考えられる.
転換実験において,Cha11is・β株のように受容
菌として適当な菌であっても,適当な培地(ET
3培地など)で2時間ぐらい培養した菌の転換
率が最大で,それ以上培養を続けた菌の転換率は
急速に低下し,4時間培養菌ではほとんど転換し
なくなる.すなわち,competentな状態とincom−
petentな状態のあることがわかっている(図2).
そこでCPFを産生しないWicky株を受容菌とし
て,Challis一β株の種々の培養時間の無菌濾液を
CPF材料とし, SM耐性の伝達頻度をとってみ
ると,図3のように,やはり2時間培養時にCP
F活性が最大である.incompetentな状態にある
Challis一β株6時間培養濾液,もともとincompetent
であるChallis一α株,およびWicky株の培養濾
液を,competentな状態にあるChallis一β株およ
びWicky株に加えて, D N AによるSM耐性の
一64一
ゆ
96
≡ コ 、ミ5 巽 竃 Σ4 ぎ あ …i3 年 … く2 お 砺 臣 さ σ) 1 2 3 4 5 6HOURS Hg.2。 Time factor of transformation. Donor: SM resistant ChailisαDNA. Recipient=OCha− 11isβ(CPF+.Trans+)in ET3 medium,●Cha・ llisβin autoclaved ET3medium,×Challisα (CPF冒Trans雪)五n ET3 medium,△Wicky(CPF’Trans+)三n ET3 medium. DNA was added to the recipient culture incubated for the indicated time periods. Spont耽neous SM resistant mutants apPea− red at up to approximately 100 per mL
伝達を行なってみると,表12のようにcompetency
は強く,あるいは全く抑制されることがわかっ
た.これら培養濾液にはDNase活性はなく,ま
た抑制効果はDNase, RNase,60℃30分,90℃15
分の作用に安定であるが,抑制物質の化学的解明
はされていない.表13はChallis一β株の2時間および6時間培養
96
≡ 謹5 お 旨蓬4
臣署3
年皇2
痒 曲 部 畔 Σ ω 1 2 3 4 5 6HOURS 聡g・3・Competence provoking factor(CPF)acti− vity in Challisβculture丘ltrate. Donor=SM resistant ChallisαDNA. Recipient:Wicky. ○:Challisβculture丘ltrate grown in ET3 mediuln,●:fresh ET3 medium was substituted at 2 hr’in−
cubation of Challisβculture and丘ltrate was
obtained further 2 and 4 hr incubation later。
菌,すなわちcompetentおよびincompetentな時
の菌から抽出したDNAを用いて転換実験を行な
うと,本来転換可能な菌にも拘らず,incompetent
時の菌のDNAはSM耐性を伝達できないことを
示している.このことはCPFを合成する速度を
調節する遺伝子の活性がそのまま受容.菌に伝達さ
れるのか,CPFを不活化する物質の産生情報が
伝達されるなどの可能性を示しているが,その詳
細は不明である.丁呂ble l2. Inhibition of transformation by the addition of culture filtrate
Recipient CQmpe−
狽?獅モ
Nulnber of SM−resistant transfQ㎜ants per ml @ after the addition of: (103×)
6hr・丘1t.a) HE丘1t.b) α一丘It.・) Wicky丘1亡.
None
Challisβ grown ln=
Wicky
growe inET3 medium
十 0 0 0 0 5,480HE medium
0 0 0 0 0.29 HE mediumd) 十 0 0 0 0 8,250Brain heart infusion
(趾n)medium
0.32 0.34 0 0。2 0.04BHI mediumd) 十 0.27 0.33 0.09 0.35
1.
31
a)6−hour culture丘正trate of Challisβ, b)2−bour culture創trate of ChaHisβgrown in autoclaved
ET3(HE)medium, c)2−hour culture且Itrate of Challisαgrown in ET3 medium, and d)supple. mented with competence−provokiug factor.
Table 13・Transfo㎜ation by and prepared from strain challisβat incompetent state
SM・resistance transformability of二 DNA from Incompetent Cha11isβ Grown至n: HE medium for 2 hr ET3 medium fQr 6 hr Competent Challisβ Without Cells grown in ET3 十 十 十 十 Cells with
CPF
十 十 十 十 十 十 十 十 Wicky with spna) 十 十 十 Number of colonies exarnined 3 47 3 47 0 30 0 十 30 CPF冒 6% 6 0 0Recipient cells are all Challisβ. a)spn:ce11−free supematant fluid of 2−hr the primarily selected cells grown in ET3 medium.
culture of
3.D群レンサ球菌のバクテリオシン産生と遺
伝子の組み換え23) 抗生物質を産生する微生物はおもに浸漉加吻,68∫,8〃ψ彦。吻脇および真菌とされているが,細菌のな
かにも大腸菌が耳リシンを,緑膿菌がピオシンを
作るように,抗生物質(バクテリオシソ)を産生
するものが少なくない.レンサ球菌の出すバクテ
リオシン(ストレプトシン)についても報告はあ
るが,とくにD群の3’7ψ’06066〃5勲86α薦var.ろ卿。−8θη6∫についてBrockとDavie2)はそのバクテリ
オシンと溶血素は同一物質ではないかと述べてい
る.私達は,ヒトおよび動物由来のD群レンサ球菌
L5 缶 旨9LO
塞 窪 毘 0.5 垂α3 § ζ 慧α2 ≦ §o・「/\
\
52ハ
16註8ぎ
是4出
謡 1:2HOURS
Fig.4. Time course of hemolysin and bacteriQ. cin production by strain 4532.×Hemolytic titer, ● Optical density, O Bacteriocin titer.
」
塁 §o,3 ご 舘 OO,2 ぎ § 0,1/×
/
×一X
。/
/
/
/
1 2 3 4 5 6HOURSFig.5. Effect of bacteriocin on the strain 2025
culture.●Optical density and viabiHty measu.
red after the addition of supernatant Huid of the
strain 4532 culture, O Those parameters after
the additioo of supernatant fluid of the strain
2025,as controL
81株に紫外線を照射して,その77株からパクテリ
オシンが誘発されることを知った.図4はε’7ψ∫o・ 60卿∫ノ砒α」∫∫varJゴg麗ψ6‘θη54532株の培養時間の経過に伴ってバクテリオシンおよび溶血活性が
ほぼ平行して増減していることを示している.こ
の株にさらに紫外線を照射してバクテリナシン非
産生(Bact一)の変異菌を採取したところβ溶血
性も失っていた(Hemつ.
Bact+4532株の培養上清を受容菌2025株に加え
,
一66一
Table 14. Recombination between Bact+and Bact−cellsa〕 PCs. SMr. TCr. Bact㌔He:n+liq.A PCr. SMs. TCs. Bact噛. Hem幽2025 PC・. SM・. TCr. Bact㌔Hem+liq−A PCr. SMs. TCs. Bact一. Hem冒2025 Incubation time (hr) 0 2 4 0 2 4 0 2 4 CFU/mlb) 1.0×108 2.4x108 4。3×108 1.0×106 4.3x107 4.8×108 7.2XIO7 9.0×108 2.5×109 Recombinants(%) PCr. SMs. TCg, Bact+. Hem+ 0 0.66 5.80 0 0 0 0 0 0
Both cells were mixed at a ratio of 1119, and inoculated Qn horse bloQd BHI agar Plates contained 10 u/ml of penicillin at O, 2nd and 4th hr of the incubation。 a)Bact+:Bacteriocin producer, Bact一:non−producer.
b)CFU/mll Colony−fo㎜ing units per ml in the inoculum.
翌 § .、 、 〉ご ●一’ ヒ。.
岩 ’
二 し
蚤 暮。・1唾
1 /
2’遡BLE C岬 ,。/ 、o” 、 OPTICAと邸ぼY 、 ロ ●、一_ V「ABLE COUNT ヘヨヘへ へ OPTICAL’摧J
ヨ8箋
ゴ ミ7ξ
雲 差 卑6ぎ
§ 5 1 2 3HOURS
Fig.6. Growth curve of the mixed culture of
bacterioc三n−producing strain liq鼻A and nonprodu− cing strain 2025。×Streptomycin’「esistant st「ain
2025,0Strain Iiq−A,●Mixed culture of both
stralns.
37℃で作用させると,図5のように0・D・(光学
密度)が1時間で急速に低下するとともに,生菌
単位も比例して減少した.3’ψ’0‘00螂勲θ6α傭var・Z蜘吻ゴ6η∫A(liq−A)株をBact+の供与菌とし,
SM耐性の2025株を受容菌として混合培養したと
きの0・D・を,それぞれの単独培養と比べてみる
と,増殖速度が著しく遅れることがわかった(図
6).この混合培養から分離された菌のなかにSM
耐性で且つBact+という両親株の形質を共有する
いわゆる組み換え型recombinantが見出された.
そこで種々のマーカーをつけた変異菌を作って組
み換えrecOmbinationの実験を行なった1例が表
14である.多くの検討の結果,組み換え型の採取
はもちろん混合培養のときに限り,供与菌と受容
菌の五二の比は1:9∼19が最適であり,組み換
えの起こる形質はBact+とHem+のみで薬剤耐
性は伝達されなかった.しかもBact+とHem+
の組み換えはつねに同時に起こった.組み換えは
供与菌と受容野が接合するときに何らかの機序で
遺伝子の一部の授受が行われるために生ずるもの
であるが,混合培養系に供与菌の抗血清あるいは
トリプシンを加えると組み換え率は著しく低下
し,また供与菌の生菌の代りに加熱死菌,培養
無菌濾液あるいは抽出DNAを用いたのでは全く
組み換えが起こらなかった,これらの結果から,
D群のバクテリオシンは少なくとも活性のある細
菌ウイルス(バクテリオファージ)ではないと思
われ,パクテリオシン産生因子は,大腸菌などで
知られている性因子fertility(F)factorと同様の
ものであるか,F因子に密接に結合したものでは
ないかと考えられる.そしてパクテリオシソは同
時にβ溶血性を有するもののようである.
4.考察と結語
以上私はレンサ球菌のA,DおよびH群におけ
る相異なる変異型式について述べてきた.その何
れの場合においても溶血性の変異が見られるが,
溶.血性そのものは菌の有する一表現型に過ぎない
のであって,それも他の表現型を別の面から見て
いる可能性の少なくないことを指摘した積りであ
る.レンサ球菌とく.にヒトに関連するレンサ球菌
各群のなかで,AおよびD群は種々の環境あるい
は薬剤に対する感受性において対称的な性状を有
し,A群は感受性が大であり, D群は小である.
そして両群とも遺伝学的に見て可成り安定してい
るように思われる.といって遺伝的変異を免かれ
るものではないことは上述した通りである.
このような実験的な事実を直ちに臨床医学に適
用することは飛躍しすぎるとの回りを免か激ない
が,若干の示唆を空き出すことはでぎると考え
る.レンサ球菌の検索に際して,β溶血を指標とす
ることは多くの場合正しいが,β溶血陰性である
からといって病原性なしとして捨てるわけにはい
かないことがあるであろう.臨床的には明らかに
溶レン菌感染症であり,ASL−0など抗体が陽性
であるにも拘らず,いかに努力しても菌を分離す
ることができないということを往々にして耳にす
る.もちろん化学療法の有無を考慮に入れて判断
した上での発言である.この際,いわゆる緑レン
菌が培養されても常在菌として無視されるであろ
うし,また弱いβ溶血を示すレンサ球菌を調べて
バシトラシン感受性や血清学的にA群でないと判
定されると,これを原因菌としては判断し難いこ
ともあろう.私達はin vitroで観察したような変
異が,in vivoでも起こるという確証はまだえて
いないが,A群レンサ球菌が非定型的な姿で潜ん
でいる可能性を捨てることはでぎないと考える.
その意味で臨床家ならび検査担当者の徹底的な繭
検索を期待したい.α溶血を示す緑レン菌などをin vitroでA群の
特徴を備えたβ溶血菌に変える現象について,細
谷ら6)および著者ら25)が報告している.これは病
原性の観点から見ると,亜急性心内膜炎を起こし
うる菌が,急性腎炎やリウマチ熱を起こしうる菌
に変わったともいえるであろう.細菌の病原性
は,その菌の代謝,化学構造,産生物質などの生
物活性の総合であり,代謝が一一部変わればそれに
伴って化学構造も産生物質も,薬剤感受性も変わ
りうる.とすれぽA群レンサ球菌が変異によって
C群あるいはD群レンサ球菌と同定されるような
性状を示すようになること,あるいはその逆の現
象が起こることも不可能ではないかも知れない.
分類は便宜的な手段であって不変のものではな
い,私達は捉われずに謙虚に自然界の現象を見つ
める態度を忘れずにいたいものである.
これらの研究は本学微生物学教室ならびに北里研究所 の多くの同僚の協力によって行われセ.最後に本特別講演の機会を与えられた東京女子医科
大学学会長および会員に感謝する.文 献
1)Bracco, R.M., M.R Kraus, A.S. Roe and C。M. MacLeod= Trans飴rmation reaction between pneumococcus and three strains of streptococci. J Exp Med 106247∼259(1957)
2)Brock, T.D. and J.M.1)avie= Probable identity of a group D hemolysin with a bacteriocine. J Bacteriol 86 708∼712 (1963)
3)EUiott, S.D。= Type and group polysac− charides of group D streptococci. J Exp Med
111621∼630(1960)
4)Grimth, F.: Signi丘cance of pneumococcaI types. J Hyg 27113∼159 (1928)
5)H董rano, T., T.1toh, T. Tomura and M.
Yosh董oka 3 Characteristics of group A strepto.
coccal variants treated with Initomycin C. Appearance of cell waH galactosamine and uridine diphosphate−N−acetylglucosamine−4− ePime「ase activity。 Japan J Microbiol 1753∼ 62(1973)
6)細谷省吾・中沢昭三・杉原久義:溶血性連鎖球
菌と緑色連鎖球菌との間の可逆的変異につい
て.皿.綜合医学 12537∼546(1955)
7)Ito, T., T. Hirano, T. Tomura and M. Yos11董oka 3 Trans負)rmability of group Hstreptococcus Challis.1. D三minution alユd ac.
quisition of transR)rmability inα一hemolytic variants of group H streptococcus Challls. Japan J Microbiol 17431∼437(1973) 8)Ito, T., T. Hirano, T. Tomum and M. Yoshioka 3 Trans仮)rmab量lity of group H
streptococcus Challis. II. Transformation of hemolytic activity and competence-provoking factor nonproducibility. Japan J Microbiol 17 439tNi444 (1973)
9) Jones, D. and P.M.F. Shattock: The
tion of the group antigen of grlup D tococcus. J Gen Microbiol 23 335,w343 (1960)
10) Krause, R.M. and M. McCarty: Studies on
the chemical structure of the streptococcal cell wall. I. The identification of a mucopeptide in the cell walls of group A and A-variant streptococci.J Exp Med 114 127N140 (l961)11) Krause, R.M. and M. McCarty: Studies
on the chemical structure of the streptococcal cell wall. II. The composition of group C cell walls and chemical basts for serologic specificity of the carbohydrate moiety. J Exp Med 115 49---v62 (1962)
12) Lancefield, R.C.: The antigenic complex of Stroptococcus hemolyticus. I. Demonstration of type-specific substnace in extracts of coccus hemolyticus.J Exp Med 47 91 ew103 (1928)
13) Lowbury, E.J.L. and L. Hurst: The
sitivity of staphylococci and other wound bacteria to erythromycin, oleandomycin, and spiramycin. J CIin Pathol 12 163N169 (1959) 14) za#ifi iAp : rsM・hSl me enXts o na = v v nt rc
-g-6maeethgl E v・< ; i>i y yaxttesdwD,hgi)Iftet -[) LN-C. J Antibiotics 3 158-165 (1949)
15) Michael, J.G., B.F. Massen and R.E.
Perkins: Effbct of sublethal concentrations of penicillin on the virulence and antigenic composition of group A streptococci. J teriol 85 128eN1287 (1963)
l6) asStwZ}O<RB ・ spl.Ll ut・ :Mre Ii: : fii[EXN pt.,mz h" 6 s) ag L 1: lj pexo rsnma ec tgl , ag ec
= v JiL pydyyftNttegoMEe:Db(. ptva
fi¥mess 46 80・-82 (1972)
17) Pakula, R., Z. Fluder, E. Hulanicka and
W. Walczak: Studies on transformation of
streptococci. Bull Acad Polon Sci,. Cl II 6 319ev323 (1958)
18) Pakula, R., E. Hulanicka and W. Walczak: Transfbrmation reactions between streptococci, pneumococci, and staphylococci. Bull Acad Polon Sci, Cl II 6 325N328 (1958)
l9) Pakula, R., E. Hulanicka ancl W. Walczak: Transfbrmation reactions between different species of streptococci and between cocci, pneumococei and staphylococci. Schweiz Z AII Pathol 22 202N214 (l959)
20) Rosenda1, K.: Investigations of resistant streptococci belonging to group A streptococcus. Acta Pathol Microbiol Scand 42 181rNv188 (1958)
21) Sanders, E., T.M. Fester and D. Scott:
Group A beta-hemolytic streptococci resistant to erythromycin and lincomycin. New England J Med 278 538ty540 (1968)22) Shockman, G.D. and H.D. Slade: The
cellular location of the streptococcal group D antigen. J Gen Microbiol 37 297t-v305 (1964)
23) Tomura, T., T. Hirano, T. Ito and M.
Yoshioka: Transmission of
genicity by coajugation in group D cocci. Japan J Microbiol 17 445N452 (!973)
24) SmaTE : tsMmesuXvaoptrwM`rc-R B - y
LR¥eYmeta¥RYXeeS. J Antibiotics Ser B
17 126・--129 (1964)
25) Yoshioka, M. T. Konuma and T. Kunii:
Antibiotic resistant group A strepto¢occi. II. General characteristics of P-hemolysis restored organisms derived from ct-hemolytic mutants.
JapanJ Microbiol 10 33tw42 (1966)
26) Yoshieka, M. and T. Kunii: Antibiotic
resistant group A streptococci. I. Acquired in vitro resistance to penicillin, mitomycin C, tetracycline and streptomycin. Japan J giol 9 87N99 (1965)