駿河湾におけるソラスズメダイPomacenrus coelesisの生活
史
誌名
誌名
東海大学紀要. 海洋学部
ISSN
ISSN
13487620
著者
著者
鈴木, 克美
日置, 勝三
柏原, 正尚
巻/号
巻/号
21号
掲載ページ
掲載ページ
p. 99-114
発行年月
発行年月
1985年10月
農林水産省 農林水産技術会議事務局筑波産学連携支援センターTsukuba Business-Academia Cooperation Support Center, Agriculture, Forestry and Fisheries Research Council Secretariat
東 海 大 学 紀 要 海 洋 学 部 第21号 99-114頁(1985)
駿 河 湾 に お け る ソ ラ ス ズ メ ダ イ
P
o
m
a
c
e
n
t
r
u
s
c
o
e
l
e
s
t
i
sの生活史ネ
鈴 木 克 美 料 ・ 日 置 勝 三 料 ・ 柏 原 正 尚 * 料Life History of the Damselfish
P
o
m
a
c
e
n
t
r
u
s
c
o
e
l
e
s
t
i
s
in Suruga Bay,
]apanKatsumi SuzU!o, Syozo HIOKI and Masanao KASHIHARA
Abstract
The behavior and life cycle of the blue damselfish, Pomacentrus coelestis JORDAN et ST ARKS, was五rstlystudied at th巴coastof Uchiura, Suruga Bay, central Japan, about 350
N. P. coelestis inhabited close to rocky bottoms 1-20m deep and resided this coast throughout the year. They actively behaved from spring to fall but kept a stand still close to the shelters in winter.
Reproductive season began in June and falIed in September. The territorial males deposited their nests each under miscelIan巴oussmall shelters, viz., small boulders, empty cans, broad leaved sea-weeds, etc. The single male courted the plural f巴malesone aft巴r another for their mutual r巴productionand showed his brilliant nuptial coloration.
According to the present study of age determination by annual ring reading on lat -eral body scales, the population of P. coelestis was composed two and/or three year classes. Juveniles firstly appeared in June. They, then, reach吋 their sexual maturation at about one year after being bom. Their life span was presumed about two years of age or less.
The intersexuality was discovered as the testis-ovum. 1t was found in 15 smalI males, 35.0-51.7mm in fork length, in which the yolkless oocytes reduced gradualIy in shape and in number from March to June. 1t suggested a presence of the rudimentary her -maphroditism in P. coelestis.
Early life history was described in detail from the fertilized eggs, 1. 00-1. 15XO. 45-0.55mm in diameter, to the post-larval stage, 6. 72mm in total length, 19 days after hatching.
Stomach cOntents showed a broad feeding habit of P. coelestis containing from cope -podan zoo-planktons to diatomous algae.
* 東海大学海洋学部業績A
第316号.受理1985年4月18日.東海大学海洋科学博物館研究業績 No.85. 料東海大学海洋科学博物館100 鈴木克美・日置勝三・柏原正尚
緒
τ=-Eヨ
日本泊岸産スズメダイ科魚類は約
9
0
種が知られている(日本魚類学会,1
9
8
1
;
RANDALL et a,.l1
9
8
1
;
ほか〉が,その生態的知見はまだ極めて少ない. うちソラスズメダイ PomacentruscoelestisJORDAN
etSTARKSは新潟県佐渡
(
3
8
0N)
を分布北限とし (HONMAa
n
d
KITAMI,1
9
7
8
)
,千葉県以南の日本本土太平洋沿岸では普通種であるのに,生態についてはまだまとまった報告がない.著者らは
1
9
7
7
,....,1
9
8
0
年に駿河湾泊岸で本種の生態と生活史に関する研究を行い,興味ある知見を得たので報告す る.本報告の一部は昭和54年度目本魚類学会年会で、発表された. 研究方法 本報告は月例調査による潜水観察と標本採集,水族館における水槽飼育などの研究結果に基くも のである.沼津市江梨地先に潜水調査の定点を設け (Fig.1
)
, SCUBAを使用して1
9
7
7
年には毎 月1
回の野外観察と標本採集を行い,1
9
7
8
年以降は随時の調査を実施した.調査日数は各国とも1
"""4日である.標本採集には自製巻網を使用し,採集標本は外部形態を観察計測後生殖腺と消化管 Nト
l
l
T
l
Omae-saki Iro -saki 30' 13S0 E 15' 30' 45' 139" Ose-sakio
2km Fig.1. Outline map of the Uchiura coast in Suruga Bay showing the station (Enashi) where the investigation were done.駿河湾におけるソラスズメダイの生活史 101 を摘出して,生殖腺はブアン氏液固定後常法による厚さ7μ のパラフィン包埋切片標本としてエオ シン・ヘマトキシリン二重染色を施し,消化管及び全体標本は10%ホルマリン液浸として研究に供 した. 上記とは別に親魚を採集し水槽(1.25 X O.7 X O.9Hm, O.79mつに飼育して繁殖生態の観察と初 期生活史の記載を行い併せて稚仔育成を試みた.本研究使用の標本は東海大学海洋科学博物館に保 存されている. 結 果 生活環境 調査定点の海底地形及び本種の生息環境について概略を述べる:水深3 m以浅の海 底は勾配約 100 ,径 1m 以上の露岩が多くその間隙を径約50cm 以下の転石が埋める.水深 3~12m の海底は勾配約 400 で径約50cm 以下の転石が重積し,水深 12~20m の海底は勾配約 300 で砂泥底 に径約1 m以上の岩石が散在する.水深20m以深は勾配約400の一様な砂泥底となる .3~6 月に は水深12m以浅にフクロノリ Calρomania sinuosa(ROTH) DERBES et SOLIER及びホンダワラ類 Sargassum spp. が繁茂する. ソラスズメダイは水深 3~12m の海底付近に個体数が最も多く,水
深 15~20m で稀となり,水深20m 以深には見出されなかった.
本調査定点で周年普通に観察される魚類にネンブツダイ Apogon semilineatus TE~IMINK et SCHLEGEL, クロホシイシモチ A. notata(HOUTTUYN),キンギョハナ夕、、イ Franziasquam争innis (PETERS),スズメダイ Chromisnotatus(TEMMINCK et SCHLEGEL)ナガサキスズメダイ Pomacent-rus nagasakiensisT ANAKA,ホンベラ Halichoerestennuis戸nnisGUNTHER, ニシキベラ Thalas -soma cuρido(TEMMINCK et SCHLEGEL)などがある.
本沿岸の年聞の月平均表面水温は 13~280C である(阿井ほか. 1969). 生活様式 本種は本定点に周年にわたって生息する.12~ 3月は個体数も少なく動作不活発で 1~数尾ずつ海底の転石付近で静止するか転石下や岩石間隙内にかくれる. 4~ 1l月は個体数が著 しく増加し行動も活発化する.この季節には海底を離れて大群を形成するものと海底付近に群がり または散在するものとにほぼ 2 分される.海底に散在するものの多くは雄であって 6~9 月には海 底のくぼみや岩石の間障の周辺になわばりを形成し,活発ななわばり防衛行動を行う.なわばりの 中心には産卵床があって,雌に対する求愛から卵保護に至る一連の繁殖行動を行う.産卵床には雑 多な材料が利用され,径約30cm以下の転石の下,側面,廃棄スコップの匙部, 空缶の内壁,ウミ
ウチワ PodinaarborescensHOLMES (褐藻〉の葉柄部,フクロノリ Cal
ρ
omeniasinuosa(ROTH) DERBES et SOLIER (同〉の葉体内側などが用いられているのが観察された.海藻を産卵床に利用するスズメダイ科魚類には ChromismultilineataGUICHENOT (see MYRBERG et a,.l 1967), C. cae -ruleusCUVIER <=C. caerulea(CUVIER)) (see S
、
NERDLOFF,1970), C. insolatus(CUVIER)<
二
C. insolatα(CUVIER)) (see LONGLEY and HILDERBRAND, , 1941)などが知られ, Swerdloff (1970)はより多くのスズメダイ類が海藻を産卵床に利用する可能性を示唆した. しかし日本産のスズメダイ 科では本種以外にこのような産卵習性を有する種は未知のようである.上記外国産3種では卵はい ずれも産卵床に疎らに産卵付着され,親がこれを保護しないという.この点は密集卵塊を産卵床に 付費させ,仔魚騨化まで親魚が卵保護を行う本種並びにスズメダイ科一般の繁殖習性とは相違する. 産卵床相E聞の最短距離は 1977年 6~8 月の同日観察例で約 50cm であった.本種の産卵床は水 深 2~6m に集中する. 高水温期には成群性が顕著となり海中の至るところで大群を形成する.群は活発に移動し,海面
102 鈴木克美・日程勝三・柏原正尚
F
.
...r...-,_ 岡 崎・h卓 n
Fig. 2. Photomicrographs of gonadal development and testis-ova inP. coelestisfrom
Suruga Bay.
A. Ovary, yolkless stage, 42.6mm FL, April.B.Ovary, tertiary-yolk globule stage, 59.9mm FL, July. C.Young testis, numerous spermatogonia and a few spermatocytesareseencloseto thick walls of the seminiferoustubule, 55.5mm FL, April. D. Mature testis, walls oftheseminiferoustubulethin down and numerous spermatozoa are seen, 54.0mm FL, July. E. Testis-ova found ina young testis of a rathersmallmale, 37.5mm FL, March. F. Re -ducing testis-ova inan oldertestis, 4l.6mm FL, April.
103 直下まで上昇する場合も認められる.とくに 6~9 月には海底に散在する大型個体の周辺にこのよ うな群が接近し,群中の個体が群を離脱しで海底に下りたり海底から上昇して群に再加入する行動 が頻繁に認められる.それらは主として雄の求愛に応じて産卵床に出入りする雌である. しかし同 様な行動は繁殖期以外にも観察され,この場合の個体閣の相
E
関係ははっきりしない. 9月以降海 底には新たに着底した稚魚多数が出現し,海底でなわばりを形成する大型個体の数は著しく減少す る.なお日没後は全個体が海底に下りて転石の周辺や間隙内に密集して静止する. 生殖腺の季節変化と繁殖期 1977年の月例標本中,雌260尾(尾文長29.8~68. 3mm),雄108 尾(同35.0~70. 7mm)の生殖腺組織標本と同じく雌267尾(尾叉長22.8~68. 3mm),雄108尾(同 35.0~70. 7mm)の生殖腺指数各月平均値の変動傾向をもとに本麓の繁殖期について検討を行った. その結果を以下に述べる. 駿河湾におけるソラスズメダイの生活史 る 達 も の る す に な 種 わ 達 期 熟 本 ゅ に 動 未 ' い 期 移 と お 球 胞 後 な し 黄 旺 卵 ・ 存 卵 放 た 共 し 期 ' れ が い 球 り さ 胞 な 黄 な 出 細 期 卵 と 見 母 胞 次 け も 卵 黄 三 だ 巣 の 卵 第 ﹀ 卵 階 に は 期 る 段 月 に 仁 れ 多 6 月 辺 さ ・ 8 周 た で わ ( 満 管 U 7 期 で 器 ジ ・ 黄 胞 状 斗 る 卵 細 嚢 ひ あ 無 母 の る で ど 卵 対 あ 数 ん 期 一 で 少 と 球 右 期 だ ほ 黄 左 黄 ま は 卵 る に 数 る 細 は 質 8 る 壁 存 し 卵 は 本 の す 壁 少 れ 母 に 実 す 管 が 著 無 胞 標 中 達 管 か ま 精 月 巣 7 埼 精 胞 が 討 て 細 の 壊 発 精 ほ 含 に 6 精 ・ 汁 細 空 向 検 ベ 母 月 崩 の ・ 細 る が 般 ・ が る 支 は の 傾 的 す 卵 日 ・ 腔 る は れ 本 一 る 子 す が に 数 行 学 は た む 巣 す に さ 標 は あ 精 成 子 月 多 退 織 月 し 9 含 卵 当 月 出 る に で ' 形 精 日 に に 組 5 始 ・ を と 該 4 見 め 月 著 胞 を に 内 体 の 閉 め 巣 膜 に が 認 5 顕 細 嚢 内 9 巣 総 腺 3 を 幻 卵 被 型 3 数 を ・ が 精 包 管 ・ 精 殖 成 予 の 巣 生 多 胞 刀 加 ' で 精 巧 ' ど 生 形 l 一 様 卵 発 胞 細 引 増 胞 所 輸 引 し な 巣 黄 ( 両 は 時 巣 細 母 F F の 細 随 は 巴 厚 る 日 卵 卵 る と 巣 向 精 原 精 日 数 母 の に 町 肥 す が す の 卵 非 精 の ( 胞 精 部 月C
が 在 い n=362 15 10 ︾ ︽ 白 ℃ E 一 方 四 ECO 5Mar. Apr. May June July Aug. Sep. Qct. Nov. Fig.3. Monthly change of average gonad indexGI=
(GW jFL3) X 104 • (GW, gonad weight; FL, fork length) inP. coelestis. Solid circles, female; open circles, male.
。
2) 生殖腺指数平均値の変動 生殖腺指数GI二 (GW/
F
L3)X 104 (GW,生殖腺重量 g;FL,尾叉長 mm)の各月平均値の変動傾向を見 ると,卵巣,精巣ともに 3~5 月に 低く 6月より増加を始め 7月に急増 して8月に最高値(13.30)を示すが, 9 月に急減して以降は上記 3~5 月 と同様低指数値を示す.雌雄の生殖 腺指数各月平均値は互いに似た変動104 鈴木克美・日置勝三・柏原正尚 傾向を辿るが, 6月だけは精巣の同指数平均値が卵巣のそれより高く,雄性機能の成熟が先行する 示唆を受ける (Fig.3).生殖腺指数平均値の年間変動傾向は,前述の組織標本による検討結果とよ く一致する. すなわち, 本沿岸における本種の繁殖期は6~8月でその盛期は7~8 月とみなされ た.機能的な生殖腺を有する最小の雌は尾叉長 44.1mm,同じく雄は47.9mmで,いずれも 6月に 採集された. 雌雄同体現象 1977年 3~6 月に採集された比較的小型の雄15尾(尾叉長 35. O~51. 7m m)の 精巣には小型の無卵黄期卵母細胞が見出された.これは YOSHIKAWAand OOURI (1978) が Tilゆ
-ia zilli(GERVARIS)で報告したのと同様なもので,3~4 月の小型標本で、は卵母細胞数も多いが, 5月の標本には少ない (Fig.2 E, F). 体色 海中で観察される本種成魚の体背面から体側にかけての体色には個体差があり,金属光 沢を有する青緑色(輝青緑色), 無光沢の紫褐色, 無光沢のj農藍青色に大別することができる.う ち輝青色の個体は主として海底を離れて泳ぐ群中に存在し,無光沢紫褐色の個体は一般に海底で散 在する.群を離脱して海底に下りた個体が輝青色から紫褐色へ短時間中に変わり,海底を離れて群 に加入した個体がこの逆の体色変化を示す例が珍しくない.無光沢の波藍青色は繁殖期に海底に散 在する個体にのみ現れ,水槽飼育によってこれが求愛行動中の雄の婚姻色であることが確認された. これを詳細に見ると,体後半部がとくに濃く,垂直鰭縁辺が明るい輝青色で,頭部に明瞭な白い1 縦条,体背面に不明瞭な白い 3~4 横帯を認める (Fig. 4).幼稚魚の体色には個体差はほとんどな し金属光沢のある鮮明なコバルト青色を呈する. 中村 (1935) は全長17~20mm の本種稚魚が無 色透明で,次いで腹部に黄色が発現するとしたが,著者らの知見です主体長15m mで上述の体色を呈 する.但し,採集直後に急速にコバルト青色を失って全体無色透明となり,まもなく原体色を回復 したl尾(全長 20mm)があった. 卵,仔稚魚の形態 本種の産卵習性については松岡 (1962),岡本 (1973a,b)の報告があり, 著者らの観察結果も既報とほぼ同様であったので、省略する.初期生活史についても松岡(1962) が 卵内発生と府:化直後の仔魚の形態を報告したが,ごく簡単な記述に止まっているので,以下に鮮化 Fig. 4. Nuptial coloration appearedintheterritorial male of P. coelestis under his courtship and reproductive behavior in the aquarium.
105 19日後(後期仔魚期)までを記載する. 27.20Cである. 1)受精卵と卵内発生 受精卵は長卵形の付着沈性卵(径1.00~ 1. 15XO. 48~0. 55mm)で, 受精直後は卵黄内に大油球 l個(径0.15mm)と小泊球数個を有するが発生の進展に伴って油球は 癒合し,目主体形成時は大油球 l個となる.卵の長軸の一端に膜状付着器がある.本種受精卵付着器 の性状は既知の本科魚類 (Dascyllusspp., FrsHELsoN, 1964, ほか ;Amphi.ρr.ion spp., GOHAR, 1984,ほか ;Chromi.s spp., MYRBERG et a,.l 1967,ほか〉が撚糸状の付着糸を有するのとは相違 しむしろ特異的である.卵内発生中,卵の大きさや外形はほとんど変化しない. 受精45分後,付着器側に旺盤が形成される.旺盤形成期以降の卵内発生経過の概要をTable1及 びFig.5にまとめた.受精27時間後,旺体と卵黄上に黒色素胞が出現,同106時間20分後,府化直 前となり眼に黒色素胞が出現 (Fig.5).受精4日後,日没後まもなく鮮化が始まり,その 15分後 に一部を残して鮮化を一旦終了する.残余の卵は翌日のほぼ同時刻に勝化した.gjl手化は日没の 13~ 26分後に始まり,騨化開始時刻は日没時刻の変動に伴って変わる. 2) 前期仔魚期 解化直後,全長2.76~2. 80mm,卵黄長径0.21mm,油球径O.08~0. 12mm, 筋節数5+23二28,口が聞き,JlI門も体前端より全長の1/3の位置に聞く.卵黄前端近くに1油球 がある.胸鰭が形成されている.頼粒状黒色素胞が頭部に1個,消化管上に数個あり,尾部腹面に 沿って十数個並ぶ.背膜鰭起点は頭頂部にある.眼球の黒化はまだ不十分である.体表全面に頼粒 が散在(図では省略)する (Fig.6A). 醇化24時間後,全長2.80~2. 90mm,卵黄長径0.12mm,油球径0.05mm,尾部渡面の黒色素胞 が拡張し,最後端の1個を除いて相互に連結する (Fig.6 B). 3) 後期仔魚期 卵黄は鮮化3日後に消失して後期仔魚期に入る. 癖化5日後,全長3.02~3. llmm,筋節数4+21=25 (Fig. 6 C). 醇化12日後,全長4.56mm,筋節数7+17=24.体高が増し全長の18.2%となる.偲蓋後縁から 第4筋節にわたり鰻が認められる.胸鰭基底上方に背膜鰭起点がある.鱈蓋部に 2 *と 3本の 2小 糠列形成.上顎骨及び顎歯が明瞭となる.偲傑骨出現.頭部及び尾部腹面の黒色素胞消失.体腹面 に多数の赤色素胞出現,ヰ胞下縁に黄色素胞出現(Fig.6 D).鮮化13日後以降,その前日まで容器 表層付近を浮漂していた仔魚が器底付近に下降する傾向が明瞭となった. 仔魚期のそれは 24.6~ 駿河湾におけるソラスズメダイの生活史 卵期の飼育水温は 24.4~25.30C, Developmental processs of eggs ofP. coelestis Table1.
Time after spawning Drawing code hr. min. in Fig. 5. Developmental stage 4-cell stage 8-cell stage 16-cell stage Morula stage Blastula stag巴 Formation of embryo 5-myotome stage; Kupffer's vesicle appears 15-myotome stage; Kupffer's vesicle disappears; lens in eye 28-myotome stage; auditory vesicles appear; tip of tail a -parts from the surface of yolk lmmediately before hatching A B C D E F G nununununumD ハ U ハ リ ハ U のムハ UFU つ dqυ つ 臼 ハ UAunu 1 2 2 3 5 7 0 7 6 - A つ ︼ つ 臼 P D H 20 106
鈴木克美・日置勝三・柏原正尚 106
Development of eggs of P. coelestis.
A. 8-cell stage, 2 hr.aft巴rspawn. B. Morula stage, 3 hr. 30min. Blastula stage, 5 hr. 30min. D. Formation of embryo, 17hr.25min. 5-myotome stage, 20hr. F. 15-myotome stage, 27hr. G. 28-myotome stage, 56hr. H. lmmediately before hatching, 106hr. 20min. C. E. Fig.5. i d -2 型 車 普 通 司 選 宮 剖 週 僻化16日後,全長4.81mm,体高は全長の27.1%,神経椋と血管椋が認められる.脊椎骨末端が 上屈し尾部棒状骨形成.尾鰭僚が上下2群に分かれ合計13本を数える.智鰭篠原基形成,腹鰭原基 出現,筋節の背腹両縁が前方に向かう (Fig.6E). 瞬化19日後,全長6.72mm,体高は全長の34.8%,各鰭の聴と軟僚の識別が可能となりD =沼十 8, A=II+9,
V
二 1を数える (Fig.6 F).鮮化20日後以降の仔魚飼育を継続することができず, 19日間で稚仔育成実験を終了した. 稚魚の出現と群形成 本研究において年間で最も早い稚魚の出現は6月24日 (1977年)に記録 されている.稚魚の個体数は6月にはまだ極めて少ないが, 7""'9月に著しく増加し,成長も顕著 である.稚魚は転石の間隙や海底のくぼみに集まり, 一般に数尾, 稀れに約50尾に達する群がり (assembly)を形成する.最小の稚魚標本(尾叉長13.2mm)は9月に採集された.尾叉長18.0mm の標本で初生鱗の形成と各鰭係数が定数に達しているのが確認された.採集はできなかったが,本 定点に出現する稚魚とみなされる最小個体の尾叉長は約10...,llmmである.発見または採集された, 稚魚が鮮化後何日を経たものかは明らかでないが,本科他種の既知例では瞬化後約1カ月で稚魚が 着底出現するとされる (HELFRICH,1958; MYRBERG et a,l. 1967; SWELDLOFF 1970;塩田, 1977;駿河湾におけるソラスズメダイの生活史
Fig. 6. Larvae ofP. coelestis.
A. Newly hatched larva, 2. 80mm in total length. B. Larva, 2. 92mm in total length, 24hr.after hatching. C. Larva, 3. llmm in total length, 5 days after hatching. D. Larva, 4. 56mm in totallength, 12days after hatching. E. Larva, 4. 81mm in total length, 16days after hatching. F. Larva, 6.72mm in total length, 19days after hatching. 107 鈴木ほか, 1985).従って本種でも, 稚魚は鮮化約1カ月後に出現するとみなしてよいものと思わ れる. 8月には成長のよい稚魚は尾叉長約40mmに達して未成魚の小群を形成し,成魚群の下方に接近 して成魚群の移動に追従する様子が観察される.
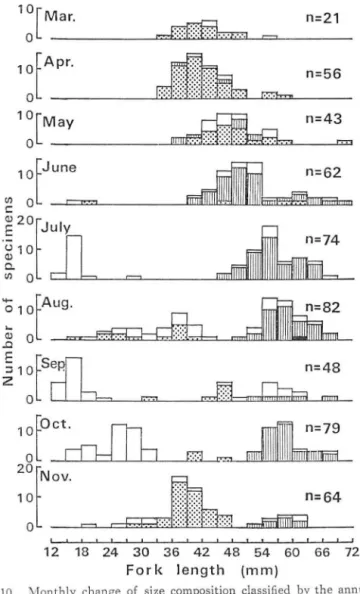
9
月以降,この未成魚群は次第に成魚群に加入混 合して同一群を形成するに至る.この経緯は月例標本の体長組成にも明らかである(Fig.10).本種 では体側鱗の検査によって年齢を査定し得たので,その結果と照合しながら後述する. 年齢査定 スズメダイ科魚類の年齢査定に関する研究例はまだないようである.本種では体側 鱗に見出される輪紋によって年齢査定を試み標本群の年齢構成について知ることができたのでその108 鈴木克美・日産勝三・柏原正尚
Fig.7. Lateral bコdyscales show:ng th巴growthlines and annual ringsof
P.coelestisfrom Suruga Bay
結果を以下に述べる. A. No ring, 38.9mm FL, August. B. One ring, 48.9mm FL, May. C.Two rings, 60.8mm FL, August, rl~r2, annual ring 1977年の月例標本中273尾(尾叉長13.2~70. 7mm) の右体側(第 4~6 側線鱗の下方 5~6 番 目)を検査したところ0,1, 2輪紋の3群に分離することができた.但し2輪紋が見出されたの は8月採集の1尾(尾叉長60.8mm)だけであった.本種体側鱗に現れる輪紋はキンセンイシモチ
Apogon syanosoma BLEEKER (鈴木・上野, 1983)及びクロホシイシモチ(鈴木ほか,印刷中〉の それと同じく成長線間隔が広くなって形成されるもので,いゆわる冬輸ではない(Fig.7 ). しかし, 以下に述べるような理由で,年齢形質として採用できるものと考えた. 標本のうちまず輪紋1~を有する鱗の輪紋縁辺成長率 α =(R ーの/r
(
R
,鱗長;r,輪紋長)につ いて述べる. 5月には縁辺成長率0.00級への顕著な集中性が認められ,6 月の縁辺成長率はO.OO~ 0.50級でモードは0.20級にある.その他の月には縁辺成長率の集中傾向を認めがたい.12~ 2月の 標本を欠くが,本種が低水温期に行動不活発で, 4~5月以降に行動が急に活発化することを考え 合わせると,体側鱗の成長線間隔の伸長による輪紋形成が年l回5月を中心に形成されると見て, 無理はないように思わ九る (Fig.8).鱗長 (R) と尾叉長 (L)はほぼ直線的に相関し, 関係式駿河湾におけるソラスズメダイの生活史 109 n=2
r
n u n u n u q ι 4 a n=18 10[Ju町
n=44 o Jイ
1
円
n
[ ω July n=46E
10。
~ 0 AUgr
-
f
YLJ1
= i " " t ー n=36。
。
一
Sep. n=5E
0 コ octn寸千~
n=34=
コ
Nov. n=8 Eコ Eコr1-『ョ
=
L...L...。
0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0 Growth rate of marginal regionFig. 8. Monthly growth rate frequency of marginal region of one annual ring on the lateral body scale ofP. coelestisfrom Suruga Bay. L二24.89R +8.59が得られた (Fig.9). 体側鱗に 1輪紋を有する 193尾の平均鱗長1.41mmを上 式に代入して得られた L=43. 69は輪紋形成期と考えられる 4~5 月採集の標本の尾叉長平均値43. 50mmに近い. 成長・成熟体長・寿命 海底観察の結果に採集標本の尾叉長組成 (Fig.10)を考えあわせると, 駿河湾泊岸での本種の稚魚期以降の生活史は概略次のように推測される.稚魚は 6~9 月に尾叉長 1O~13mm となって着底出現し,以後成長をつづけて 11月に尾叉長36~42mm 級を中心とする未成 魚群となる.その後は成長が非常に緩慢となり,翌年 3 月の尾叉長33~54mm 級に連続する.精巣 と卵巣はすでに分化しているが,精巣内に小型卵母細胞が見出される例があり,幼期雌雄同体現象 の存在が示唆された. 4~6 月は体側鱗の輪紋形成期である. 6月には新規出現の稚魚 1尾を含む 3尾が無輪紋である 他はすべて1輪紋を有し, 7月には稚魚以外の全個体が 1輪紋を有する. 6~8 月は本種の本泊岸 における繁殖期である (6月出現の稚魚については論議の項で述べる). すなわち本沿岸では本種 は生後約 7~11 カ月で体側鱗に 1 輪紋を形成し,その約 2 カ月後に性成熟に達する.前述のように, 性成熟に達した最小の標本は雌で、尾叉長44.1mm,雄で同47.9mmであった. 8月には 1輪紋を有 する成魚群と新規出現の無輪紋の稚魚群との2群構成が明瞭になる.この月には 2輪紋を有する 1 尾(尾叉長60.8mm) が得られ,少数ながら満約 2歳またはそれ以上の年齢の個体の生残の可能性 が示唆された. 9~11月は l 輸紋と無輪紋の 2 群で、構成される.前者は前年生まれの個体に相違な
鈴木克美・日置勝三・柏原正尚
110
30 40 50 Fork length (mm)
Relationship between length of lateral body scale and fork length inP. coelestisfrom Suruga Bay. 80 70 60 20 10 2.5 2.0 0.5
。
。
Fig.9. ~ 1.5 E E 』.
ニ.
.
o z 由。
市 U ω1.0 安 打 倒g 句 4 2 6 b h N u ι 、tz E A 守 ぷ 誕 湾 局 理 ι い.12~2 月に標本が採集されなかったが,上述の年齢組成と体長分布の毎月の変動傾向から,本 種は出生後翌年の繁殖期に性成熟に達l.-,少なくとも1回以上繁殖に加わったのち,生後 2度目の 低水温期に大多数が死亡するものであろう.すなわち本種の寿命は一般に生後約 l年半と考えられ るが,一部の個体が生後翌々年の夏まで生残する可能性もあるC
F
i
g
.
1
0
)
.
食性と摂餌活動 本種の胃は空虚状態でV字型,膨満状態でト型に近い.各月の標本中より雌 雄各5
尾を任意に抽出した合計9
0
尾の胃内容物は主として動物プランクトシで,カイアシ類が最も 多量かつ普遍的に見出され,出現頻度は96%であった.属段階まで査定できたカイアシ類は19属で, Oncaea (出現頻度51.5%), Paracalanus (同17.5%),
Corycaeus (同5.9%),
Claussocalanus (同駿河湾におけるソラスズメ〆イの生活史 111
;
:
1
1
J
ポ 羽 市 {
n=21 n=56 n=43J
幅
一
ニ
=
コ
~
:
1
1
A
u
ン早
ι J
臨
2;10[1h
n=48同関
川
川
口コロ皿
2
;
?
:
E E l -: : -E z-::
:
;
;
:
:
;
;
;
; : : : : ! 目 h v ・ i ・ -J EE -: ・ : : ; ; : : : : z -J J ・ -; -: 一 : F E -一 -J-:
;
;
t
F
1 0
4
v
v
-c
-o
u
-b
﹁
L
同
L │ L 円 u n u n u n u n u 句 E q ' ﹄ 4 2岬
L
:
ニ
n=64 E園 田
12 18 24 30 36 42 48 54 60 66 72For
k
l
e
n
g
t
h
(mm)
Fig. 10. Monthly change ofsize compositionclassifiedby theannual ring group ofP. coelestisfrom Suruga Bay.
Dotted bars, no ring; striped bars, one ring; solidbars, two rings; Open bars, undetermind. 3.5%), Acrocalanus (同2.4%)などが多い方であった.ソラスズメダイ 1尾が摂餌していたカイ プシ類は平均27個体であった.その他の動物プランクトンには魚卵(出現頻度33%), フジツボ幼 生(同28%)が目立ち, 多毛類幼生,尾虫類,偲脚類も比較的普遍的に見出された.ほかに珪藻 (出現頻度20%),藍藻(同29%),その他の植物片(同16%)なども比較的普通に検出されている. 本種の歯は A OYAGI(19-11)が2列型とした通り, 門歯が密に並んで歯帯を形成するなかに先端が 円錘形に尖る犬歯がほぼ等間隔に突出することが確かめられた.歯列の性状と食性の関連について は論議の項で述べる. 月例標本
3
7
2
尾の摂餌率F
R
=(SCWjBW)X100
(SCW , 胃内容物重量 ;BW,体重〉はO.OO~ 2.43の範囲を示し個体差が大きく,性差や体長との相聞は認めがたい.季節との関連については6112 鈴木克美・日置勝三・柏原正尚
"
"
"
'
8
月及び1
0
"
"
"
'
1
1
月には0
.
1
0
,...,0
.
9
0
への集中傾向が多少認められるが,有意とみなすほどでなく, 季節的変動傾向を認めることもで、きなかった.潜水観察では本種の摂餌活動が潮流速の大なるとき に集中する傾向が窺われたが,標本採集が潮時や月齢との関連を考慮して行われたものではなかっ たためもあって,明瞭な結果を得ることができなかったと思われる. 論議
1) 雌雄性について スズメダイ科の雌雄性については先に触れたようにMOYERa
n
d
N
A
K
A
Z
O
N
O
(
1
9
7
8
)
がクマノミ属 6種で雄性先熟雌雄同体現象を報告した.すなわち幼若な個体の生殖腺には卵巣・精巣両部分が併 存し,先に精巣が機能的になったのち精巣部分が退縮して卵巣が機能化する,というのである. し かし本研究のソラスズメダイでは,前述の通り3
"
"
"
'
6
月に得られた小型の1
5
尾の精巣内にいわゆる 精巣卵が見出されたものの,機能的雌雄同体現象の証拠またはそれを示唆する結果は得られなかっ た.本種の精巣は細精管の発達したt
u
b
u
l
a
rt
y
p
e
であって,組織学的所見からもこれが機能的卵巣 から変換した二次精巣である見込みは少ない.本種の尾叉長組成では大型の個体に雄がやや多い傾 向はあるが,雌雄の大きさに明瞭な性差を見出しがたく,既知のクマノミ属でl唯が雄よりも明らか に大であるのとはかなり相違する.小型個体に見出された精巣卵は前年生まれの未成熟な個体の精 巣に検出されたもので,この無卵黄期卵母細胞は繁殖期に向かい精巣が発達を始めるに伴い退化消 失する.調査はなお不十分であるが,いゆわる痕跡的雌雄同体現象r
u
d
i
m
e
n
t
a
r
yh
e
r
m
a
p
h
r
o
d
i
t
i
s
m
の存在が考えられる. 2) 食性についてH
r
A
T
T
a
n
d
S
T
R
A
S
B
U
R
G
(
1
9
6
0
)
はM
a
r
s
h
a
l
l
諸島産スズメダイ科のうち Chromis,
Dascyllus 両 属のみが動物プランクトIン食性,
Abudザdザ,
Pomacentrus両属がb
r
o
w
s
e
r
またはg
r
a
z
e
r
の典 型とし,前2属は先端円錘形に尖る歯を上下顎に疎生させ,後 2属は門歯が密に並んで歯帯を形成 すると述べた.F
r
s
H
E
L
s
o
N
e
t
a
l.(
1
9
7
4
)
,BELK (
1
9
7
5
)
,S
A
N
O
e
t
a
l.(
1
9
8
4
)
も上述の食性につい てほぼ同様な研究結果ないし見解を述べ,祖ー(
1
9
7
0
)
はソラスズメダイ亜科が海藻を主食とする 傾向を報告した.A
L
L
E
N
(
1
9
7
5
)
はPomacentrusをb
r
o
w
s
e
r
またはg
r
a
z
e
r
とした上でそのうちの ソラスズメダイがプランクトン食性でしかも Chromisのように中層に群生する Pomαcentrusの うちの例外的な種であると述べた.ALLEN (
1
9
7
5
)
がソラスズメダイを Pomacentrus中の例外的 な食性種であるとした根拠は明らかでないが,上述の本種の歯列の形状はこの種の広食性を裏付け るもので、ある. 3) 再生産と稚魚の出現について 本沿岸で本種が再生産を果していることは確実であり,繁殖期が6
,...,8
月,その盛期が7
"
"
"
'
8
月 とみなされたのは前述の通りである.しかしこの泊岸では稚魚の初出現は6月である. 6月出現の 稚魚の個体数はまだ少ないが, 7月には極めて多数となる.これら稚魚は騨化約 1カ月後と推察さ れるが,するとその発生は早いもので5月とみなければならない.本泊岸で 5月に繁殖の可能性は ほとんどなしそれらの稚魚がどこから由来したのか,その散布機構に興味がもたれる.本種稚魚 が流れ藻について海潮流に搬送される事実はすでに知られ(中村,1
9
3
5
;
千田,1
9
6
5
)
, 内田, 道 津(
1
9
5
8
)
は対馬暖流域の表層プランクトン中から本種稚魚を採集している.鈴木(
1
9
6
6
)
は能登駿河湾におけるソラスズメ夕、イの生活史 113 半島に夏期出現する本種稚魚が若狭湾以南西から搬送されるものと考え,松岡 (1962)は相模湾で 7 月に見出される全長20~30mm の本種稚魚が相模湾以南の海域から流入したものと推測した.駿 河湾内浦泊岸での本種の繁殖期の開始とほぼ同時期に出現する稚魚は,本泊岸以南の海域から搬送 されたものと考えざるを得ない.繁殖盛期以降に本治岸に出現する稚魚群は,他海域よりの搬送流 入個体と本泊岸での再生産による個体の混在した群で、あろう.即ち暖温帯浅海を中心に分布する場 合,その地理的分布の北限に近い場所で南方海域からの搬送と定着再生産の両要因の併存によって 個体群を維持していることが推測され,本泊岸のソラスズメダイをその1例とみなすことができる ように思われる.このような個体群の消長には冬期水温の年変動が重要な意味をもつはずである. 上述の広食性や産卵床材料の選択性の幅広さ,稚魚の流れ藻による長距離搬送の可能性なども,本 種をスズメダイに次いで北方に分布定着させている重要な要因と考えてよいかも知れない.因みに 本種の低温耐忍限界は 12~130C (鈴木, 1966)で,これは本調査定点となった駿河湾内浦泊岸水深 O~50m の冬期の月間平均水温とほぼ一致する.
謝 辞
本研究は東海大学海洋学部学生 (1977~1978年当時〉多川達郎,高橋明彦両氏の協力によってな されたものである.生殖腺組織の検討には東海大学海洋科学博物館小林弘治氏,卵・仔稚魚の飼育 には岡田中洋一講師,胃内容物査定には東海大学海洋学部魚谷逸朗講師,現地調査には沼津市内浦 漁業協同組合並びに同杉山邦彦氏にそれぞれ懇切な援助を受けた.各氏に深く感謝する.引 用 文 献
阿井敬雄・松原壮六郎・松島又十郎(1969):内浦湾の養殖場環境についてー1.静岡水試研報, (3), 29-37. ALLEN, G.R.(1975): Damsel五shesof the south seas. T.F.H. Publ, N巴ptuneCity, 240pp.AOYAGI, H. (1941): The damselfishes found in the waters of Japan. Trans. Biogeogr.Soc. Japan, 4 ( 1), 157-279.
BELK, M.S. (1975): Habitat partitioning in two tropical reef fishes, Pomacentrus lividus and P. albo -fasciatus. Copeia, 1975( 4), 603-607.
FISHELSON, L.(1964)・Observationsof the biology and behavior of Red Sea coral五shes.Bull. S巴aFish Res. Sta., (37), 11-26.
FISHELSON, L., D. POPPER, and A. AVIDOR (1974): Biosociology and ecology of pomacentrid fishes aro -und th巴SinialPeninsula (north巴rnRed Sea). J. Fish Bio,.l1974 (6), 119-133.
GOHAR, H.A.F. (1984): Commensalism between五sh and anemone (with description of the eggs of Amphiρrion bicinctus RむPPELL).Fouad Univ. Pub.lMar.Bio.lSta. Ghadag巴(RedSea), (6), 282
289.
HELFRICH, P. (1958): Th巴earlylife history and reproductive behavior of the mao mao, Abudefduf
abdominalis (QUAY et GAIMARD). Ph. D. Diss. Univ. Hawaii, 228pp. (not seen).
HlATT, R.羽T,and D.W. STRASBURG (1960): Ecological relationships of th巴 fishfauna on coral reefs of th巴MarshallIslands. Eco.lMonogr., 30 (1), 65-127.
HONMA, Y.and T. KITAMI (1978): Fauna and flora in the waters adjacent to the Sado Marine Biolo -gical Station, Niigata University. Ann. Rep. Sado Mar. Biol.Sta., Niigata Univ., (8), 7-81. LONGLEY, W.H. and S.F. HILDERBRAND (1941): Systematic catalogue of the五shesof Tortugas, Florida.
9 で 1 1 4 1 姐 Carnegie Inst., Wash. Publ,. (535), 1-531. 松岡飛良 (1962):ソラスズメダイの産卵生態について.水産増殖, 10(3), 1-6.
MOYER, ,T. and J A. NAKAZONO (1978): Protandrous hermaphroditism in six species of the anemonefish genus AmphiPrioπin Japan. Japan.,JIchthyol., 25 (2), 101-106.
MYRBERG, A.A., Jr., B.D. BRAHY, and A.R.EMERY (1967) : Field observations on reproduction of the damselfish, Chromis multilineata (Pomacentridae), with additional notes on general behavior. Copeia, 1967 (4), 819-827. 中村秀也 (1935):小湊附近に現われる磯魚の幼期(其八).養殖会誌, 5 (3/4), 35-44. 日本魚類学会(編) (1981) :日本産魚名大辞典.三省堂,東京, ii +vii+834pp. 岡本一志(1973a ) :僕のフィールドノートその1.産卵期のソラスズメ夕、イの分布.マリンパピリオン, 2(9), 54-55. 岡本一志 (1973b) :僕のフィールドノートその 2. 自然環境下における産卵期のソラスズメダイの生態につい て.マリンパピリオン, 2(10), 62-63.
RANDALL, J.E., H. IDA, and J,T. MOYER (1981):A review of the damselfishes of the genus Chromis from Japan and Taiwan, with description of a new species. Japan. J. Ichthyol,. 28 (3), 203-242. SANO, M., M. SHIMIZU, and Y. NOSE (1984): Food habits of teleostean reef五shes in Okinawa Island,
southern Japan. Univ. Mus., Univ. Tokyo, Bull., (25), 1-128. 千田哲資(1965):流れ藻の水産的効用.水産研究叢書13, 日本水産資源保護協会,東京, 56pp. 塩田昭仁(1977):ネッタイスズメダイの産卵,仔魚の飼育.ノアの箱舟(宮島水族館通信), (15), 2-3 祖 一 誠 ( 1970):珊瑚礁に見られるスズメダイ類 Pomacentridae の食住と社会行動に関する研究.鹿児島大 卒業論文, 1-28. 鈴木克美(1966)・九十九湾とその周辺の磯魚の生態 金沢大能登臨海研報, 6, 17-24 鈴木克美・日置勝三・栗田雅紀 (1985)・駿河湾におけるスズメダイ Chro加・snotatus の生活史.東海大紀要 海洋, (21), 115-127. 鈴木克美・上野信平(1983)・駿河湾におけるキンセンイシモチ(テンジクダイ科)の生活史 東海大海洋研報, (5), 49-55.
SWERDLOFF, S.N. (1970): Behavioral observations on Eniwetok damselfishes Pomacentridae:Chromis with special reference to the spawning ofChromis caeruleus. Copeia, 1970 (2), 371-374.
内田恵太郎・道津喜衛 (1958):対馬暖流水域の表層に現われる魚卵・稚魚概説. 対馬暖流開発調報, 第2輯
(卵・稚魚・プランクトン篇), 3-65
YOSHIKAWA, H. and M. OGURI (1978): Effects of steroid hormones on the sex differentiation in chichlid
五sh,Tilapia zilli. Bull. Japan. Soc. Sci. Fish., 44 (10), 1093-1097. 鈴木克美・日置勝三・柏原正尚 114