Transactions of the JSME (in Japanese)
日本機械学会論文集
離接地信号と
CPG
を用いた脚の離散的な運動周期調節による歩容創発
前田 一成
∗1,前岡 俊
∗2,徳田 献一
∗1,中嶋 秀朗
∗1Gait emergence by discrete leg motion period control using contact signals and CPGs
Issei MAEDA
∗1, Shun MAEOKA
∗2, Kenichi TOKUDA
∗1and Shuro NAKAJIMA
∗1∗1,∗2Wakayama University
930 Sakaedani, Wakayama-shi, Wakayama 640-8510, Japan
Abstract
Most living organisms need locomotion to achieve a series of goals, such as eating, escaping, and breeding. In short, locomotion refers to the periodic activities of various parts of the body and their coordination mechanisms, such as walking, jumping, etc. Each locomotion is characterized by the animal’s body and its surroundings. The common of almost all locomotion is motion of the body center such as the trunk. Therefore, we believe that the periodic movement of the central part of the body is important for achieving movement. In this study, we focused on walking with limbs. We believe that the trunk movement using limbs is a combination of periodic movements of each limb. A central pattern generator (CPG) is used for generating periodic motion. We make robot generate gait to show that walk is the motion of a center part of a body. We propose a discrete control method of leg periodic motion using a ground signal to generate gait. In the periodic adjustment of the proposed method, the leg’s motion period is changed according to the CPG phase when the grounding state of the leg is switched. We use a modular legged robot to verify the proposed control method. We did experiments and simulations using a real robot and a physics engine. It is confirmed that although the initial stage of each leg movement is random, specific gait will be generated over time. Some gaits are gaits that are shown by animals, such as Tripod and Direct Wave.
Keywords : Central Pattern Generator, Locomotion, Interlimb coordination, Gait emergence, Modular robot
1. 緒 言 多くの動物が摂食,逃避,繁殖などの目的を達成するためにロコモーションを行なっている.ロコモーション は身体の周期運動によって行われ,その基本は身体各部の周期運動とその協調機構にある.各ロコモーションは 動物の身体や周囲環境によって特徴づけられるが,全てのロコモーションにおいて共通しているのは身体の中心 部の運動である.身体の中心部は脊椎動物の体幹やクモヒトデの盤,アメーバでは身体全体などである.我々は 身体の中心部の周期運動がロコモーションの実現において重要であると考える.我々は身体の中心部を体幹とす る動物のロコモーションを対象に研究を行なう.
動物の周期運動の根底にあるのは CPG(Central Pattern Generator)である(Grillner,1975).CPG はシンプ ルかつ低次元の入力信号から,多自由度の周期性を有する時空間的パターンの出力を生成できる神経回路である (Ijspeert,2008).CPG は振動子として扱えるため,CPG モデルとして様々な振動子モデルが用いられている.CPG
∗1 正員,和歌山大学(〒 640-8510 和歌山県和歌山市栄谷 930)
∗2 和歌山大学
E-mail of corresponding author: [email protected]
Received: 28 September 2020; Revised: 21 February 2021; Accepted: 8 April 2021
モデルに用いられる代表的な振動子モデルには松岡振動子と位相振動子が挙げられる.松岡振動子はニューロン同 士の相互抑制による振動のメカニズムである half-center 仮説を数学的に模擬した振動子である(Matsuoka,1985). 位相振動子は振動子の状態を位相のみで記述する振動子である(蔵本,2005).位相振動子は振動子同士の相互作 用による振動子集団のダイナミクスの研究で多く使用されている.我々は周期運動の生成のために CPG を利用し て研究を行なう. 体肢を活用するロコモーションの一つとして陸上の脚式動物の歩行が挙げられる.歩行の様式は歩容と呼ばれ, 一定速度で移動を行なう場合の各脚の運動計画と捉えることができる.連続した移動において速度が一定の場合, 無秩序な脚の運動は転倒の可能性やエネルギー効率の面から適切ではない.4 脚動物では Walk,Trot,Pace,Gallop などが歩容として知られている.昆虫などの 6 脚動物では Tripod などが知られている.動物は歩容を移動速度に 応じてエネルギー消費量が最小になる様に変化させていると考えられている(Hoyt and Taylor, 1981).力学系的 観点では歩容はその系におけるアトラクタ状態を表していると考えられる(Pfeifer and Bongard,2010).アトラ クタ状態は多少の外乱を受けても,元の状態を維持する特徴がある.このアトラクタ状態の特徴によって動物は 環境に対して順応的な運動を実現していると考えられる.また,歩行は身体-神経系-環境の相互作用によって創発 されると考えられている(Taga,1995).動物は CPG によって基本となる周期運動を生成し,身体と環境からの 感覚信号によるフィードバックを受けて身体と環境に対して適切な歩行運動を実現し,アトラクタ状態であると 捉えられる歩容を生成していると考える. 歩容生成について整理する.歩容生成を脚の位相差をあらかじめ設定し歩容を生成する方法と脚の位相差を設定 せずに各脚の協調の結果として歩容が創発される方法に分ける.歩容は各脚の運動周期調節と脚間協調によって 生成されると考える.運動周期調節には連続的な調節と離散的な調節があると考える.連続的な調節では支持脚期 および遊脚期において,脚の運動周期を常に調節する.連続的な調節として Owaki らの研究が挙げられる.Owaki らは脚に働く床反力を感覚フィードバックとして用い,連続的な調節により歩容創発を実現している(Owaki et al.,2017).脚に働く床反力により脚全体の離接地状態を推定することが可能である.Owaki らの手法では,脚 全体の離接地状態を推定することが可能であるが,離接地している脚がどの脚であるかは推定できない.そのた め,隣接脚の情報を利用することで各脚の運動順序を決定させると考えられる.また,Owaki らは脚に働く床反 力の大きさに応じて支持脚期間を延長させることで,脚の運動速度を連続的に調節した.一方,離散的な調節で は支持脚と遊脚の切り替え時に脚の運動周期を調節する.離散的な調節の例として Ambe らの研究が挙げられる (Ambe et al., 2013).Ambe らは各脚の CPG を結合し,脚の接地タイミングに応じて脚の運動周期を調節させる ことで,歩容生成を実現した.杉本らは 4∼20 脚の受動歩行ロボットによる歩容生成をシミュレーションにより 確認した(杉本他,2010).左右 2 脚を 1 対とする胴体を組み合わせることで脚数を変化させることができ,その 胴体間の結合部の自由度構成や胴体数などの身体構造によって歩容が変化することが示された.杉本らの研究か ら他の脚との情報伝達を行なわない能動的な歩容創発手法が考えられる. 本研究では他の脚との情報伝達を用いない能動的な歩容創発手法の実現を目指す.他の脚の CPG と結合しない 独立した CPG によって各脚を運動させ,離接地のタイミングによって脚の運動周期を調節させる.本論文では, 我々は歩容創発させる制御系を解析が容易となるように分割され単純な構造となる自律分散制御として設計する. 制御系による歩容創発を実機ロボットおよびシミュレーションを用いて検証を行なう.ロボットは脚数による体幹 への脚の影響を考慮するために脚数を調節可能なモジュール型を採用する. 本研究では,はじめに多脚の受動歩行の研究例から,各脚が独立であるにも関わらず歩容を創発させる制御則 があるのではないかと考え,他の脚の CPG と接続しない脚単位の CPG と離接地信号を用いた歩容創発の制御則 を発案した.次に松岡振動子をその制御則に組み込んで,実機による歩容創発を実現した.制御則による歩容創 発を解析していくために,まず松岡振動子を用いた歩行シミュレーションを行なった.シミュレーションでも実機 と同様に歩容創発の結果を得られたため,次に歩行中の制御則の挙動を理解しやすくするために簡易な構造であ る位相振動子を用いた歩行シミュレーションを行なった.
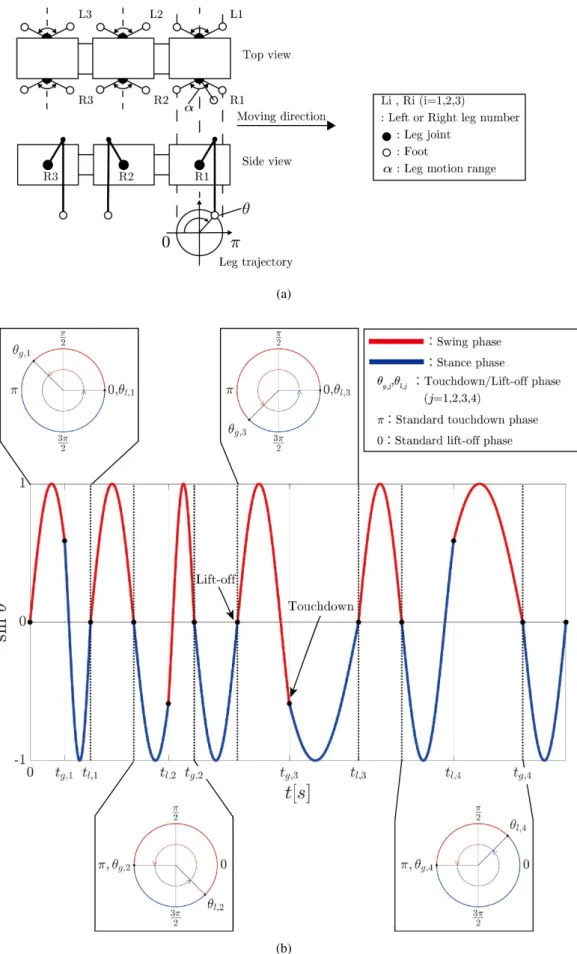
2. 提 案 手 法 各脚が他の脚と情報の伝達を行なわない独立した状態による歩容創発について説明する.我々は各脚が独立した 状態による歩容創発を実現するために離接地信号と CPG を用いた脚の離散的な運動周期調節を提案する(図 1). 脚の離接地の切り替えごとに離散的に CPG の運動周期を調節する(図 1b).運動周期調節は離接地時に基準と なる CPG 位相と実際に離接地したときの CPG 位相を比較し,脚の運動周期を調節する.脚の運動周期調節は脚 毎に独立して行なわれる.実際の接地位相が基準接地位相よりも小さい場合,脚の運動速度は速くなる(図 1b 中 tg,1-tl,1).実際の接地位相が基準接地位相よりも大きい場合,脚の運動速度は遅くなる(図 1b 中 tg,3-tl,3).実際 の離地位相が基準離地位相よりも小さい場合,脚の運動速度は速くなる(図 1b 中 tl,2-tg,2)).実際の離地位相が基 準離地位相よりも大きい場合,脚の運動速度は遅くなる(図 1b 中 tl,4-tg,4). 松岡振動子と位相振動子を CPG として提案手法に組み込む.松岡振動子は CPG として歩行ロボットなどに用 いられている(福岡,木村,2003).松岡振動子は神経回路を模した振動子モデルであるため,実験結果の議論に おいて動物の知見を生かしやすくなると考えられる.しかし,松岡振動子では後述するように決定すべきパラメー タが多いため,歩容創発における CPG の周期と運動周期調節の調節量の影響を解析するには多くの労力が必要と なる.そこで,CPG の周期と運動周期調節の調節量を明確に表すことができる位相振動子も提案手法に組み込む. また,2 種の振動子を用いることによって,提案手法が振動子の性質に依存しないことを示す.本論文では提案手 法により歩容が創発可能であることを示す. 2·1 松岡振動子の組み込み 本節では提案手法への松岡振動子の組み込みについて説明を行なう.松岡振動子は,神経回路によるリズム生 成モデルの中で数学的扱いが容易であり,振動が発生する条件が詳しく解析されている(Matsuoka, 2011).松岡 振動子は式(1)∼(4)のように非線形一階微分方程式で表され,周期的な出力を生成する.
τuU˙e, f i=−Ue, f i− AVe, f i+ u−Cf ey(Uf ,ei) (1)
τvV˙e, f i=−Ve, f i+ y(Ue, f i) (2)
y(Uf ,ei)= max(Ue, f i, 0) (3)
yi= y(Uf i)− y(Uei) (4)
ここで,i は CPG の番号,e, f は伸筋,屈筋ニューロン,また,U,V はニューロンの内部・疲労状態,u は駆 動入力,A は疲労係数,τu,τvは運動周期に関係する時定数,Cf eは拮抗ニューロン間の結合係数,y(Uf ,ei)は一定
の振幅で振動する CPG の出力を表し,本研究では振幅は 1 とする.yiは-1∼1 の範囲で振動する.
我々は脚軌道として単純な軌道である円軌道(図 1a)を採用した.脚軌道の生成には 2 つの CPG を用い,θpi,
θyiの軸関節角度を生成する.θpiは脚を上下に運動させる関節の関節角度であり,θyiは脚を左右に運動させる軸関 節の関節角度である.θpi,θyiは式(5),(6)から生成する.θpi,θyiでは歩行を実現する際に式(1)∼(3)の各 パラメータ(Ue, f,Ve, f, A,Cf e,τu,v)がそれぞれ異なる可能性がある.yp(Uf i),yp(Uei)はθpiにおける式(3)の CPG
の出力を表す.同様に yy(Uf i),yy(Uei)はθyiにおける式(3)の CPG の出力を表す.α は各関節の最大動作角であ
る.図 1 のθを各脚における運動周期調節に用いる CPG とし,本研究ではθi=θpiとする.
θpi=α(yp(U
f i)− yp(Uei)) (5)
θyi=α(yy(Uf i)− yy(Uei)) (6)
CPG に基づいた脚の離接地時の自律的な運動周期調節について説明する.基準とする接地位相をθi=π,基準
(a)
(b)
Fig. 1 Behavior of proposed model in walking. (a) Legged robot and its leg trajectory. (b) Timeseries of CPG phaseθ and touchdown/lift-off event.
節される.τ0u,vは固有時定数である.CPG のパラメータτu,vは基準離接地位相と実際の離接地位相に応じて変化 する.
Touchdown :τu,v= π
2π−θiτ0u,v (0≤θi< 2π) (7)
Li f t− o f f :τu,v= π
3π−θiτ0u,v (π≤θi< 2π) (8a)
π
π−θiτ0u,v (0≤θi<π) (8b)
2·2 位相振動子の組み込み 本節では提案手法への位相振動子の組み込みについて説明を行なう.前節と同様に脚軌道は円軌道(図 1a)と する.脚の軌道は円軌道であるため,i 番目の脚の円軌道上の位相と CPG 位相θiは同一と捉えることができる. 脚 i の角速度をωiとすると単純な振動子は式 (9) になる. dθi dt =ωi (9) 位相振動子を用いた提案手法の組み込みにおいても基準離接地位相と実際の離接地位相の差に応じて脚の運動 周期を変化させる.脚の離接地の切り替えが発生したとき,脚の角速度ωiは離接地のフィードバックと固有角速 度ωsによって以下のように変化する. ωi=ωs+ B(θsg,sl−θi) (10) ここで,θi は離接地の切り替え時の CPG 位相である.θsgは基準接地位相,θsl は基準離地位相を表す.脚が 支持脚となる場合はθsg=πである.脚が遊脚,かつ (0≤θi<π) の場合はθsl= 0 である.また脚が遊脚,かつ (π≤θi< 2π) の場合はθsl= 2πである.B は基準離接地位相と実際の離接地位相の差に対するフィードバックゲ インである.ωiは脚の離接地状態の切り替えごとに計算される.式 (10) から脚の運動速度は図 1b のように変化 する.
2·1 節と同様に脚軌道が円軌道となるように軸関節角度θpi,θyiを生成する.軸関節角度θpi,θyiは CPG 位相θi
を基に式(11),(12)から生成される.
θpi=αsinθi (11)
θyi=αcosθi (12)
歩容創発を対象とする Owaki らの制御則(Owaki et al.,2017)と比較を行なう.Owaki らの制御則を式(13) に示す.
˙
ϕi=ω−σ1NiVcosϕi+σ2(
1 nL nL
∑
j∈L(i) kjNVj) cosϕi (13)ここで,ϕ は位相振動子の位相,i は脚の番号(i = 1, 2, ...6),ωは固有角速度,σ1,σ2は正の定数を表してい る.NV i は i 番目の脚の床反力の大きさ,L(i) は i 番目に隣接する脚の集合,nLは L(i) の要素数,kjは隣接脚の床 反力の大きさの重みを表している.式(13)の 2 項目は当該脚による制御量,3 項目は隣接脚による制御量を表し ている.Owaki らの制御則では位相振動子の位相と脚にかかる床反力の大きさによって脚の運動速度が決定され る.脚にかかる床反力は歩行中に変動していくため,脚の運動制御は連続的に行なわれる. 我々の制御則では脚の離接地信号の切り替え時にのみ脚の運動を制御するため離散的な運動制御となり,その制 御量は予め設定したフィードバックゲインと基準離接地位相と実際の離接地位相の差によって決定される.Owaki らの制御則は連続的に脚の運動を制御するが,我々の制御則では離散的に脚の運動を制御する.また,我々の制 御則は隣接脚の情報を必要としていない. 2·3 ロボットへの組み込み 提案手法をロボットへ組み込む方法について説明する.脚毎に他の脚と情報の伝達を行なわない独立した振動 子を組み込む.組み込むプログラムは,離接地判定,脚の運動周期調節,脚の運動の一連の流れを繰り返す.実機 ロボットとシミュレーションへの組み込みは基本的に同じである. 2·3·1 松岡振動子のパラメータ決定 松岡振動子は振動する条件や周波数などが解析されている.パラメータ決定の方法としてはまず CPG の振動周 期を設定する.本研究では CPG の振動周期と歩行周期は同じである.設定した振動周期を満たすように式(1), (2)の各パラメータ(Ue, f,Ve, f, A,Cf e,τu,v)を設定する.式(7)∼(8b)の離接地による時定数調節の定数,式
(5),(6)の動作角度αは実験的に求める. 2·3·2 位相振動子のパラメータ決定 位相振動子の各パラメータは前節の松岡振動子の挙動と一致するようにパラメータを求めた.前節の松岡振動 子を離接地の感覚フィードバックによる時定数変更を行なわない状態のまま動作させ,松岡振動子の出力周期か ら固有角速度ωsを求める.次に基準離接地位相と実際に離接地したとする位相の差とその場合における松岡振動 子の出力周期から,フィードバックゲイン B を求める.動作角度αは実験的に求める. 3. 実験およびシミュレーション 本章では提案手法の歩行実験およびシミュレーション環境について説明する.また,松岡振動子を用いた多脚 ロボットの歩行実験と歩行シミュレーション,位相振動子を用いた歩行シミュレーションの結果を示す.歩行実験 では実機ロボットを,シミュレーションではロボットモデルを使用する.本論文では提案手法が CPG に依存しな いことおよびシミュレーションが実機と同様に歩容創発をシミュレートできていることを示すために,実機実験 とシミュレーションの二つを用いている.今後の発展ではシミュレーションを用いて制御則による歩容創発の解 析を行なう.本稿では歩容創発のみを報告するが,今後の課題としては位相振動子の簡易さとシミュレーションに よる歩行実験の自動化を利用して歩容創発の条件・変化を明らかにしていく.また,ロボットの初期状態や位相振 動子の各パラメータによる歩容の変化についても明らかにする. 3·1 実験およびシミュレーション環境の共通点 実験およびシミュレーション環境の共通点を説明する.実験およびシミュレーションでは歩行を行ない,定常 歩行を対象とする.定常歩行とは一定速度の歩行である.歩行環境は水平な整地とする.ロボットの脚数は 6 脚 と 8 脚である.ロボットの自由度構成を図 2 に示す.図 2 に示したものは 1 モジュールである.モジュールを組 み合わせることで任意の脚数のロボットを実現させる.1 モジュールに 1 対の脚を有する.実機ロボットとシミュ レーションモデルは同一の自由度構成である.脚毎に振動子による CPG を設定する.松岡振動子を組み込んだ提 案手法と位相振動子を組み込んだ提案手法では共通して,脚軌道を表す CPG が設定されている.松岡振動子を用 いる場合では,脚軌道の位相θiの CPG と実際に脚軌道を実現するための軸関節角度θpi,θyiをそれぞれ生成する CPG が用いられる.脚軌道の位相θiと軸関節角度θpiは同一であるため,CPG の数は省略可能である.松岡振動 子を用いる場合では 1 脚に軸関節角度θpi,θyiを生成する 2 つの CPG が設定される.一方,位相振動子では脚軌道
の位相θiから軸関節角度θpi,θyiを生成するため,1 脚に 1 つの CPG が設定される.CPG は他の CPG と結合をし ない独立した状態であり,各 CPG は当該脚の離接地位相のみで出力を調節する.各脚の動作開始時間はランダム に設定し,足並みが乱れた状態から歩行が開始される.
Robot Torso
Fig. 2 DoF structure of the modular robot.
3·2 松岡振動子を用いた歩行実験 3·2·1 実験環境 本項では松岡振動子を用いた提案手法の検証を行なうための実験環境について説明する.実機ロボットを用い て歩行実験を行なう. 実機ロボットは同じ構造のモジュールを複数組み合わせ構成する(図 3).モジュールを 2 つ接続すると 4 脚移 動ロボット(図 3b)となり,モジュールを 3 つ接続すると 6 脚移動ロボット(図 3c),モジュールを 4 つ接続する と 8 脚移動ロボット(図 3d)となる.このように,脚数を容易に変更可能な単純な構造である.モジュール 1 個 の寸法は,体長 0.08 m,横幅 0.20 m,高さ 0.10 m,質量 0.38 kg である.モジュールの前後に搭載された凹型と凸 型のマグネットジョイントが,モジュール間の接続を可能にしている(図 4).マグネットジョイントは十字に配 置されている.各磁石の吸着力は 2.74 N であり,大きな衝撃をロボットに加えると接続が解除されることがある. 歩行実験は接続が解除されない条件下で行なう.各脚は 2 自由度(Pitch,Yaw 方向)を有し,そのアクチュエー タはサーボモータ(KONDO 社製: KRS-2572HV ICS)である.各モジュールにつき 1 台の Arduino UNO がそれ らのアクチュエータを制御する.脚先の接地センサが離接地状態を検知する.接地センサにはタクトスイッチを 用いており,スイッチの ON/OFF で離接地を検出する.接地センサのサンプリング周波数はマイコンのループ速 度に依存し,本稿では約 10 ms である.
歩行実験での松岡振動子θi,θpi,θyiの共通するパラメータを表 1 に, 伸筋/屈筋ニューロンの内部・疲労状態の 初期状態を表 2 のように設定した.松岡振動子の振動周期の初期値は 1.3 s である.
(a) (b) (c) (d)
Fig. 3 Multi-legged module robot. (a) One module robot. (b) Four-legged robot composed of two modules. (c) Six-legged robot composed of three modules. (d) Eight-legged robot composed of four modules.
3·2·2 歩行実験結果
図 5 に歩行実験のゲイトダイアグラムを示す.ゲイトダイアグラム内の黒の領域は支持脚期,白の領域は遊脚 期を表す.L は左脚,R は右脚を示している.数字 (1,2,・・・) は前方から数えた脚の番号である.歩容が創発さ れるタイミングを予測することは困難なため,ゲイトダイアグラムの時間幅は実験毎に異なる.
(a) (b)
Fig. 4 Magnet joints of real robot. (a) Top view. (b) Side view.
Table 1 Common CPG parameters ofθi,θpi,θyi. Drive Input u 3.6 Fatigue Coefficient A 6.0 Antagonism Coefficient Cf e 1.2
Time Constantτu 2.0 Time Constantτv 12.0 Max operating Angle [deg]α 7.5
Table 2 The initial state of extensor/flexor neuron for
θi,θpi,θyi.
θi θpi θyi
Initial internal state of extensor neuron Uei 2.0 2.0 1.0
Initial internal state of flexor neuron Ufi 0.0 0.0 0.0
Initial fatigue state of extensor neuron Vei 2.0 2.0 1.0
Initial fatigue state of flexor neuron Vfi 0.0 0.0 0.0
図 5a は 6 脚定常歩行実験のゲイトダイアグラムを示す.歩行開始時の 6 脚ロボットの足並みは不規則であるが, 時間の経過につれて,同側の前脚と後脚および反対側の中脚が同期して接地するように変化している.この歩容 は昆虫に見られる Tripod である.平均歩行速度は約 3.5 cm/s である.
図 5b は 8 脚定常歩行実験のゲイトダイアグラムを示す.歩行開始時の 8 脚ロボットの足並みは不規則であるが, 時間の経過につれて,左脚,右脚が後から前へと順に接地するように変化している.この歩容はムカデなどの多足 類が示す Direct Wave である.また,サソリも同様の歩容を示す(Bowerman,1975).平均歩行速度は約 4.2 cm/s である. 松岡振動子を用いた実機歩行実験では,歩行開始時に乱れていた足並みが揃いはじめ,最終的には生物に見ら れる歩容を確認することができた. 3·3 松岡振動子を用いた歩行シミュレーション 3·3·1 シミュレーション環境 本項では松岡振動子を用いた歩行実験と同様の結果をシミュレーションで得られることを確かめるために松岡 振動子を用いた歩行シミュレーションを行なう.3 次元動力学シミュレータ ODE(Open Dynamics Engine)を用 いる.シミュレーションの時間刻み幅は 10 ms である.歩行時の地面モデルは剛体とした.本研究では力学的制 御は対象としていないため,脚に働く力を単純化するために滑りは発生しないようにする.地面との摩擦係数は 1.0×105と設定した.ロボットモデル(図 6)は実機ロボットと同様の寸法,自由度構成である.重心位置や慣性 モーメントなどはロボットモデルの形状および質量から ODE によって計算される.実機のモジュール間の接続は 磁石により,多少の柔軟性を持っているが,シミュレーションでは単純化のためにモジュール間の接続は剛体とし て設定した. 3·3·2 松岡振動子を用いた歩行シミュレーションの結果 松岡振動子を用いた歩行シミュレーションの結果を図 7 に示す.図 7a は,6 脚定常歩行シミュレーションのゲ イトダイアグラムを示す.歩行実験と同様に Tripod が創発した.平均歩行速度は約 3.3 cm/s である.図 7b は,8 脚定常歩行シミュレーションのゲイトダイアグラムを示す.歩行実験と異なり左脚側は前脚から順に接地し,右脚 側は後脚から順に接地する歩容が創発した.平均歩行速度は約 4.3 cm/s である.
Time [s] L1 L2 R1 R2 0 15 L3 R3 (a) Time [s] L1 L2 R1 R2 0 13 L3 R3 L4 R4 (b)
Fig. 5 Result of the experiment involving steady walking locomotion on flat terrain using the multi-legged robot system and Matsuoka oscillators. (a) The gait diagram of steady walking using the six-legged robot. (b) The gait diagram of steady walking using the eight-legged robot.
(a) (b)
Fig. 6 Robot model with the three-dimensional simulator (ODE). (a) Six-legged robot model. (b)Eight-legged robot model. 3·4 位相振動子を用いた歩行シミュレーション 3·4·1 シミュレーション環境 本項では位相振動子モデルを用いた歩行シミュレーションのシミュレーション環境について説明する.シミュ レーション環境は松岡振動子を用いた歩行シミュレーションと同様である.位相振動子モデルの各パラメータは 固有角速度ωs= 8.89 rad/s,フィードバックゲイン B = 2.9,動作角度α=π/24 rad である. 3·4·2 位相振動子を用いた歩行シミュレーションの結果 本項では位相振動子を用いた歩行シミュレーションの結果を示す.6 脚定常歩行シミュレーションのゲイトダイ アグラムを図 8 に示す.3·2·2 項の 6 脚定常歩行実験と同様に Tripod が創発した.平均歩行速度は約 3.4 cm/s で ある. 8 脚定常歩行シミュレーションの結果を図 9 に示す.図中の赤枠は歩容が創発されている箇所を示す.平均歩行 速度は全て約 3.5 cm/s である.図 9a では L1,R2,R3,L4 のグループと R1,L2,L3,R4 のグループが交互に接 地する Tripod を確認できた.図 9b では L1,R2,L3,R4 のグループと R1,L2,R3,L4 のグループが交互に接地 する歩容を確認できた.図 9c では L1,R1,L3,R4 のグループと L2,R2,R3,L4 のグループが交互に接地する
(a)
(b)
Fig. 7 Result of the simulation involving steady walking locomotion on flat terrain using the multi-legged robot system and Matsuoka oscillators. (a) The gait diagram of steady walking using the six-legged robot. (b) The gait diagram of steady walking using the eight-legged robot.
Fig. 8 Result of the simulation involving steady walking locomotion using the six-legged robot and phase oscillators.
歩容となった.この歩容では,左脚側は図 9b の歩容を,右脚側は図 9a の歩容を示している.図 9d では L1,R1, L3,R3 のグループと L2,R2,L4,R4 のグループが交互に接地する歩容となった.
本節においても,歩行開始時では乱れていた足並みが揃いはじめた.しかし,図 8 のように生物に見られる歩 容である Tripod を確認することもできたが,8 脚においては図 9 のように多様な歩容を確認した.
Fig. 9 Result of the simulation involving steady walking locomotion using the eight-legged robot and phase oscillators.
4. 考 察 松岡振動子と位相振動子をそれぞれ用いた提案手法により昆虫に見られる Tripod を創発した.我々は離接地信 号と当該脚の CPG 位相のみを利用して歩容を創発させた.脚の運動周期周期調節は離接地位相によって決定され る.この離接地位相は胴体の傾斜に大きな影響を受ける.シーソーのように胴体が傾いた場合,本来の離接地離と 実際の離接地位相にはズレが生じる.胴体の傾斜は地面の傾斜や凹凸,支持脚の組み合わせによって生じる.各 脚の足並みが乱れている場合,胴体の傾きは大きくなる.胴体の傾きによって実際に離接地したときの位相は基 準離接地位相と大きく異なる.その離接地位相の差に応じて脚の運動周期を調節していく.そして,歩行中の胴 体が傾かなくなるまで脚の運動周期は調節されると考えられる.脚数が 6 脚の場合は,Tripod 歩容が最も胴体が 傾かない安定した歩容だと考えられるため,我々の提案手法では Tripod のみが創発されたのだと考えられる. 5. 結 言 本論文では離接地信号と独立 CPG を用いた脚の離散的な運動周期調節による歩容創発を実現した.提案手法の 運動周期調節では脚の離接地状態の切り替えと同時に,CPG 位相に応じて脚の運動周期を変化させる.検証のた めにモジュール型多脚ロボットを用いた.各脚の動作開始位相はランダムでありながら,時間経過によって特定 の歩容が創発されることを確認した.確認した歩容には生物に見られる Tripod や Direct Wave などが確認された.
8 脚歩行ロボットの場合,松岡振動子と位相振動子をそれぞれ用いた提案手法では結果に相違点が見られた.松 岡振動子の場合では,ムカデなどの多足類やサソリに見られる Direct Wave が創発した.一方,位相振動子を用い
た提案手法の場合では,松岡振動子に見られる歩容と異なる歩容が創発した.松岡振動子を用いた提案手法と位 相振動子を用いた提案手法で創発した歩容の数が異なる点については更に実験を行なって明らかにしていく. 我々は脚の離接地信号のみで独立 CPG 群を協調させ,歩容創発を実現したが,歩容創発のメカニズムとその条 件,創発される歩容については明らかにしていない.今後は歩容創発メカニズムと条件,身体や制御系による歩 容変化について明らかにしていく. 文 献
Ambe, Y., Nachstedt, T., Manoonpong, P., W¨org¨otter, F., Aoi, S. and Matsuno, F., Stability analysis of a hexapod robot driven by distributed nonlinear oscillators with a phase modulation mechanism, 2013 IEEE/RSJ International Conference on Intelligent Robots and Systems (2013), pp. 5087-5092.
Bowerman, R. F., The control of walking in the scorpion. I. Leg movements during normal walking, Journal of comparative physiology, Vol. 100 (1975), pp. 183-196.
福岡 泰宏, 木村 浩, 4 足ロボットの生物規範型不整地適応動歩行, 日本ロボット学会誌, Vol. 21, No. 5 (2003), pp. 569-580.
Grillner, S., Locomotion in vertebrates: central mechanisms and reflex interaction, Physiological Reviews, Vol. 55, No. 2 (1975), pp. 247-304.
Hoyt, D. and Taylor, C., Gait and the energetic of locomotion in horses, Nature, Vol. 292 (1981), pp. 239-240.
Ijspeert, A. J., Central pattern generators for locomotion control in animals and robots: A review, Neural Networks, Vol. 21 (2008), pp. 642-653.
蔵本 由紀編, リズム現象の世界 (2005), 東京大学出版会, pp. 137-184.
Matsuoka, K., Sustained oscillations generated by mutually inhibiting neurons with adaptation, Biological Cybernetics, Vol. 52 (1985), pp. 367-376.
Matsuoka, K., Analysis of a neural oscillator, Biological Cybernetics, Vol. 104 (2011), pp. 297-304.
Owaki, D., Goda, M., Miyazawa, S. and Ishiguro, A., A minimal model describing hexapedal interlimb coordination: the tegotae-based approach, Frontiers in neurorobotics, Vol. 11 (2017).
Pfeifer, R. and Bongard, J., 知能の原理 - 身体性に基づく構成論的アプローチ (2010), 共立出版, pp. 101-104. 杉本 靖博, 吉岡 秀隆, 大須賀 公一, 超多脚型受動的動歩行の実現, 日本ロボット学会誌, Vol. 28, No. 8 (2010), pp.
961-969.
Taga, G., A model of the neuro-musculo-skeletal system for human locomotion, Biological Cybernetics, Vol. 73, No. 2 (1995), pp. 97-111.
References
Ambe, Y., Nachstedt, T., Manoonpong, P., W¨org¨otter, F., Aoi, S. and Matsuno, F., Stability analysis of a hexapod robot driven by distributed nonlinear oscillators with a phase modulation mechanism, 2013 IEEE/RSJ International Conference on Intelligent Robots and Systems (2013), pp. 5087-5092.
Bowerman, R. F., The control of walking in the scorpion. I. Leg movements during normal walking, Journal of comparative physiology, Vol. 100 (1975), pp. 183-196.
Fukuoka, Y. and Kimura, H., Biologically inspired adaptive dynamic walking of a quadruped on irregular terrain, Journal of the Robotics Society of Japan, Vol. 21, No. 5 (2003), pp. 569-580 (in Japanese).
Grillner, S., Locomotion in vertebrates: central mechanisms and reflex interaction, Physiological Reviews, Vol. 55, No. 2 (1975), pp. 247-304.
Hoyt, D. and Taylor, C., Gait and the energetic of locomotion in horses, Nature, Vol. 292 (1981), pp. 297-304.
Ijspeert, A. J., Central pattern generators for locomotion control in animals and robots: A review, Neural Networks, Vol. 21 (2008), pp. 642-653.
Matsuoka, K., Sustained oscillations generated by mutually inhibiting neurons with adaptation, Biological Cybernetics, Vol. 52 (1985), pp. 367-376.
Matsuoka, K., Analysis of a neural oscillator, Biological Cybernetics, Vol. 104 (2011), pp. 297-304.
Owaki, D., Goda, M., Miyazawa, S. and Ishiguro, A., A minimal model describing hexapedal interlimb coordination: the tegotae-based approach, Frontiers in neurorobotics, Vol. 11 (2017).
Pfeifer, R. and Bongard, J., How the Body Shapes the Way We Think-A New View of Inteligence (2010), KYORITSUSHUPPAN Co., Ltd, pp. 101-104 (in Japanese).
Sugimoto, Y., Yoshioka, H. and Osuka, K., The realization of the super multilegged passive dynamic walking, Journal of the Robotics Society of Japan, Vol. 28, No. 8 (2010), pp. 961-969 (in Japanese).
Taga, G., A model of the neuro-musculo-skeletal system for human locomotion, Biological Cybernetics, Vol. 73, No. 2 (1995), pp. 97-111.