Nagoya City University Academic Repository
学 位 の 種 類 博士 (薬科学) 報 告 番 号 甲第1550号 学 位 記 番 号 第319号 氏 名 古川 純士 授 与 年 月 日 平成 28 年 3 月 31 日 学位論文の題名 ENBT1 の核酸塩基トランスポーターとしての同定と代謝酵素との機能的協 働の解析及び応用利用 論文審査担当者 主査: 林 秀敏 副査: 湯浅 博昭. 牧野 利明. 山村 壽男
名古屋市立大学学位論文
ENBT1の核酸塩基トランスポーターとしての同定と
代謝酵素との機能的協働の解析及び応用利用
平成
28年(2016年)3月
名古屋市立大学大学院薬学研究科
薬物動態制御学分野
古川 純士
名古屋市立大学学位論文
ENBT1の核酸塩基トランスポーターとしての同定と
代謝酵素との機能的協働の解析及び応用利用
名古屋市立大学大学院薬学研究科
薬物動態制御学分野
古川 純士
Functional Identification of ENBT1 as a Nucleobase Transporter
and Analysis and Application of Its Cooperation with Enzymes
Department of Biopharmaceutics
Graduate School of Pharmaceutical Sciences, Nagoya City University
一.本論文は、2016年 3月、名古屋市立大学大学院薬学研究科において審査されたもので ある。 主査 林 秀敏 教授 副査 湯浅 博昭 教授 牧野 利明 教授 山村 寿男 准教授 二.本論文は、学術雑誌に収載された次の論文を基礎とするものである。 (基礎となる報文)
1. Furukawa J, Inoue K, Maeda J, Yasujima T, Ohta KY, Kanai Y, Takada T, Matsuo H and Yuasa H: Functional identification of SLC43A3 as an equilibrative nucleobase transporter involved in purine salvage in mammals. Sci. Rep., 2015, 5, 15057
2. Furukawa J, Inoue K, Ohta KY, Yasujima T and Yuasa H: Role of ENBT1 in a suicide gene therapy using herpes simplex virus thymidine kinase with ganciclovir. Manuscript in preparation
3. Furukawa J, Inoue K, Ohta KY, Yasujima T and Yuasa H: Validation of decynium-22 as an ENBT1-selective inhibitor. Manuscript in preparation
三.本論文の基礎となる研究は、湯浅 博昭 教授の指導の下に名古屋市立大学大学院薬 学研究科において行われた。
本論文では、以下の略号を用いた。
ATP Adenosine triphosphate
APRT Adenine phosphoribosyltransferase APS Ammonium peroxodisulfate
BCA Bicinchoninic acid BSP Sulfobromophthalein
CNT Concentrative nucleoside transporter DMEM Dulbecco's modified eagle medium DTT Dithiothreitol
EEG1 Embryonic epithelia gene 1
EGFP Enhanced green fluorescence protein ENBT1 Equilibrative nucleobase transporter 1 ENT Equilibrative nucleoside transporter EST Expressed sequence tag
FBS Fetal bovine serum GCV Ganciclovir
GFP Green fluorescence protein
HEK293 Human embryonic kidney 293 cell line
HPRT1 Hypoxanthine-guanine phosphoribosyltransferase 1 HSV-TK Herpes simplex virus thymidine kinase
LAT Large amino acid transporter LDS Lithium dodecyl sulfate LPS Lipopolysaccharide
MDCKII Madin Darby canine kidney II cell line
MTT 3-(4,5-dimethylthial-2-yl)-2,5-diphenyltetralium bromide M.W. Molecular weight
NBMPR Nitrobenzylmercaptopurine riboside PBS Phosphate buffered saline
PCR Polymerase chain reaction P-gp P-glycoprotein
SDS Sodium dodecyl sulfate SE Standard error
SLC Solute carrier
SNBT1 Sodium dependent nucleobase transporter 1 TEMED N,N,N’,N’-tetramethylethylenediamine
目次
序論 1 本論 第1章 ENBT1の核酸塩基トランスポーターとしての同定と核酸塩基代謝酵素との機能 的協働 1 ENBT1の核酸塩基トランスポーターとしての同定 3 2 ENBT1の核酸塩基輸送機能の解析 5 3 ENBT1による核酸塩基輸送の速度論的解析 7 4 核酸塩基の細胞内取り込みにおける ENBT1と核酸塩基代謝酵素との 機能的協働 9 5 ENBT1の臓器分布及び細胞内局在 12 6 まとめ 14 第2章 HSV-TK/GCV 自殺遺伝子治療における ENBT1の役割:ENBT1と代謝酵素との 機能的協働の応用利用 1 ENBT1による GCV 輸送の解析 15 2 GCV の細胞内取り込みにおける ENBT1と HSV-TK との機能的協働 18 3 各種がん由来細胞株における HSV-TK 一過性導入が GCV 取り込み 及び殺細胞効果に及ぼす影響 22 4 まとめ 25 第3章 ENBT1特異的阻害薬としての decynium-22の特性1 ENBT1による adenine 輸送に対する decynium-22の阻害特性
:ENT1及び ENT2による adenine 輸送との比較 26 2 HepG2細胞における adenine 輸送への ENBT1の寄与
:decynium-22の阻害効果の解析 30
3 まとめ 32
実験の部 1 実験材料 1 試薬 34 2 培養細胞 34 2 実験方法 1 各種遺伝子のクローニング 35 2 細胞の培養方法 39 3 トランスフェクション 39 4 取り込み実験 41 5 Western blot 解析 42 6 real-time PCR 解析:hENBT1の臓器発現分布の解析 45 7 二重蛍光免疫染色による ENBT1及び P-gp の細胞内局在の解析 46 8 細胞毒性試験 46 3 データ解析 1 取り込みデータ処理 47 2 統計処理 48 謝辞 49 参考文献 50
1
序論
核酸塩基は、核酸の構成成分であり、細胞の正常な機能や増殖に不可欠な生体内化合物で ある。哺乳類における核酸の生合成経路には、アミノ酸などを出発物質として一から合成 するde novo 経路と、食餌由来あるいは不要となった核酸(核酸塩基及びヌクレオシド)を再利用 するsalvage 経路1) が存在する。特に、後者の経路は、核酸塩基の合成に必要な多量のATP を 節約できるという利点があり、肝細胞を除くほとんどの細胞において主要な核酸合成経路となって いる。ただし、salvage 経路においては、細胞外から核酸塩基やヌクレオシドを取り込む必要があ り、この過程には特異的な核酸輸送系の関与が指摘されている。核酸輸送に関しては、ヌクレオシ ド 輸 送 の 研 究 が 先 行 し て お り 、 輸 送 系 の 分 子 実 体 と し て 、equilibrative nucleoside transporters(ENTs)2,3) やconcentrative nucleoside transporters(CNTs)4,5) などが既に同定されている。また、近年、ヌクレオシド輸送に関する研究が先行したのは、核酸塩基よりもヌクレ オシドの利用が優位であるとされているためであるが、古くから赤血球などにおいて核酸塩基の特 異的輸送の報告が多数あり 6-12)、核酸塩基利用の必要性も無視できないとみられる。しかし、核酸 塩基輸送系の分子実体に関しては、一部のENT の弱い核酸塩基輸送能が知られている13,14) の みで、実質的には不明のままであった。 このような状況のもと、最近になって、哺乳類における核酸塩基特異的なトランスポータ ーとしてrat sodium-dependent nucleobase transporter 1(rSNBT1)が当研究室におい て同定された15)。しかしながら、rSNBT1 のオーソログが欠損しているヒトにおいては、
核酸塩基トランスポーターは未だ不明のままであった。
本研究では、ヒトにおける核酸塩基の動態を明らかにするため、核酸塩基輸送系の分子 実体の同定に取り組み、アミノ酸トランスポーター群であるsolute carrier(SLC)43 family に属する機能未知のトランスポーター様タンパク質として知られていたSLC43A3 16)を、核
酸塩基輸送能を有するトランスポーターとして同定することに成功した。さらに、その輸 送様式が促進拡散型であることが見出されたことを踏まえ、このトランスポーターを equilibrative nucleobase transporter 1(ENBT1)と命名することとした(Table 1)。この ENBT1の同定及び輸送機能解析と合わせて、核酸塩基利用における細胞内核酸塩基代謝酵 素との協働的機能の解析にも取り組んだ。そして、その応用利用の試みとして、ヒト単純 ヘルペスウイルス由来チミジンキナーゼ(HSV-TK)による代謝活性化によって抗がん剤と して働くプリン核酸塩基類似薬物である ganciclovir(GCV)の ENBT1による輸送及び HSV-TK による代謝との機能的協働についての解析に取り組んだ。なお、この GCV と HSV-TK を組み合わせて用いる抗がん療法は、HSV-TK/GCV 自殺遺伝子治療として提唱さ れ、開発が進められている新しい治療法である。最後に、各種の臓器由来細胞やモデル細 胞 で の 細 胞 膜 輸 送 に お け る ENBT1の寄与の評価に利用できる特異的阻害薬として decynium-22を見出し、その阻害特性の解析等に取り組んだ。
3
本論
第1章 ENBT1の核酸塩基トランスポーターとしての同定と核酸塩基代謝酵素との機 能的協働 1 ENBT1の核酸塩基トランスポーターとしての同定 核酸塩基特異的トランスポーターの探索に当たって、まず、担体介在性の核酸塩基輸送 機構が認められている赤血球に高発現している機能未知のトランスポーター様遺伝子を 中心として、染色体データベースなどから得た情報をもとに、候補遺伝子(cDNA)の検 索を行った。次に、候補cDNAを単離し、哺乳類発現ベクターであるpCI-neo vectorに組 み込んでHEK293細胞に一過性に発現させ、[3H]adenineの細胞内取り込み輸送活性を評 価した。その結果、92個の遺伝子の内の一つ(ENBT1/SLC43A3)のみにおいてadenin e取り込み活性の顕著な増大が認められ、機能未知のENBT1が核酸塩基のトランスポータ ーであることが示唆された。 ENBT1は、アミノ酸トランスポーター群に属するSLC43 familyに分類されているが、 SLC43A1(LAT3)及びSLC43A2(LAT4))アミノ酸を輸送する17,18)のに対し、アミノ 酸に対する輸送活性は認められず、機能未知のトランスポーター様タンパク質として知ら れていた。一方で、ENBT1は胚発生時の腎形成において誘導されるembryonic epitheli a gene 1(EEG1)遺伝子としても報告されている他19)、血管新生時の血管内皮細胞や甲 状腺がん組織においてENBT1の発現上昇が認められている20,21)。さらに、 lipopolysaccharide(LPS)を投与した健常者の末梢血由来の単球においてENBT1の発現 が有意に上昇することが報告されている22)。従って、ENBT1はがんなどの細胞増殖の盛 んな組織や炎症反応に関与する可能性が考えられる。そこで、ENBT1の核酸塩基トラン スポーターとしての機能解析及びその生理的役割の解明を行うこととした。 Fig. 1に示したように、ヒトENBT1を一過性に発現させたHEK293細胞におけるaden ine取り込みは、コントロール(mock細胞での取り込み)の7倍程度に達し、高い輸送活 性が示唆された。また、hypoxanthine取り込みについても、同様に取り込み増大がみら れ、高い輸送活性が示唆された。4
Adenine
Hypoxanthine
Upt
ak
e rat
e
(p
mo
l/mi
n/
mg
p
ro
te
in
)
0.0
0.1
0.2
0.3
0.4
*
*
Fig. 1. ENBT1 mediates the uptakes of adenine and hypoxanthine
The uptakes of [3H]adenine and [3H]hypoxanthine (5 nM for each) were evaluated at 37°C and pH
7.4 for 1 min in HEK293 cells transiently expressing ENBT1 (open bars) and mock cells (filled bars) for control. Data are presented as means ± SE (n = 4). * p < 0.05 compared with each control by Sudent’s t-test.
5 2 ENBT1の核酸塩基輸送機能の解析
ENBT1を安定発現させたイヌ腎臓由来細胞である MDCKII 細胞を作製し、ENBT1によ る核酸塩基輸送の詳細な解析を行った。まず、ENBT1の各種核酸関連物質に対する輸送活 性の比較検討を行った(Fig. 2A)。その結果、プリン塩基である adenine、guanine、 hypoxanthine に対する輸送活性は、コントロールである mock 細胞の30倍程度を示した。 一方で、プリン塩基の一種であるxanthine 及びピリミジン塩基である uracil に対する輸送 活性は極めて小さいことが示された。また、ヌクレオシドである adenosine、thymidine、 uridine に対しても輸送活性を示さなかった。なお、哺乳類における ascorbate のトランス ポーターがバクテリアの核酸塩基トランスポーターのオーソログであることから23)、 ascorbate が ENBT1の基質となる可能性を考えたが、輸送活性は認められなかった。 以後の実験では、高い輸送活性の認められたプリン塩基類の一つとして主にadenine を 取り上げ、ENBT1の輸送機能評価を行った。また、ENBT1導入細胞での取り込みから mock 細胞での取り込みを差し引くことにより、ENBT1による特異的な取り込みを評価した。 ENBT1による adenine 輸送の駆動力を調べるため、細胞外のイオン及び pH が adenine の 取り込みに与える影響について検討したが、いずれの影響も認められなかった(Fig. 2B and C)。従って、ENBT1は、生体エネルギーの供給により形成されるこれらのイオン類の濃度 勾配を必要としない、促進拡散型の輸送様式で機能することが示唆された。
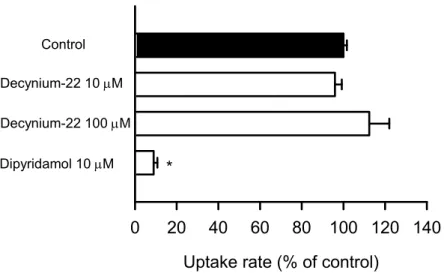
ENBT1によるプリン塩基輸送 ENBT1の基質認識特性をさらに把握するために、ENBT1 による adenine 輸送に対する各種化合物が及ぼす影響について検討を行った(Fig. 2D)。 ENBT1による輸送活性の認められた adenine、guanine、hypoxanthine は ENBT1による [3H]adenine 取り込みに対して顕著な阻害効果を示した。また、purine 及びプリン塩基の 誘導体である6-thioguanine、mercaptopurine が阻害効果を示した。その一方で、ピリミ ジン塩基及びヌクレオシドでは、ENBT1の adenine 取り込みに対する影響は認められなか った。ENT1の特異的阻害剤である NBMPR (IC50 = 0.4 - 1.4 nM)24,25)のENBT1に対す る阻害活性は、ENT1と比較して極めて弱く、ENT1及び ENT2の特異的阻害剤である dipyridamole 24,25)は阻害効果を示さなかったことから、ENBT1によるプリン塩基輸送は ENT 類とは異なる特性を有することが示唆された。その他の薬物では、核酸塩基類似医薬 品であり抗ウイルス薬として用いられているacyclovir が阻害効果を示し、核酸塩基輸送を 阻害することが報告されているpapaverine 及び decynium-22 26)も有意な阻害効果を示し た。
7 3 ENBT1による核酸塩基輸送の速度論的解析
次に、ENBT1による核酸塩基取り込みの速度論的特性を解析した。ENBT1の輸送機能解 析を進めるに当たり、adenine (5 nM) 取り込みの時間推移を解析した(Fig. 3A)。ENBT1 を導入した細胞でのadenine 取り込みは、mock 細胞での取り込みよりも著しく大きく、1 分程度までほぼ時間に比例して増大していた。これに基づき、ENBT1の取り込み初速度の 評価に適する1分以内の取り込み時間として40秒を選び、以後の実験を行うことにした。 ENBT1安定発現 MDCKII 細胞における核酸塩基(adenine, guanine, hypoxanthine)の取り 込み速度は、濃度の上昇に伴って増大したが、担体輸送に特徴的な飽和性が見られた(Fig. 3B)。この濃度依存的な取り込み速度の変化を Michaelis-Menten 型の担体輸送を仮定した モデルで解析した結果、ENBT1の adenine、guanine、hypoxanthine に対する Michaelis 定数(Km)は、それぞれ0.940 M、1.70 M、1.32 M と算出された。 一方で、adenine 輸送に対する guanine、hypoxanthine の阻害効果の濃度依存性の解析 から、IC50はそれぞれ70, 350 M と算出された(Fig. 3C)。また、hypoxanthine 輸送に対す るadenine の阻害効果の濃度依存性の解析から、IC50は13 M と算出され、これら3種の核 酸塩基においてKmとIC50が大きく異なった。一般に、トランスポーターの輸送基質は競合 阻害物質となり、KmとIC50は理論的には一致する。ここで見られた両者の乖離の問題につ
いては、adenine の例では、取り込みのKmが、salvage 経路における adenine の代謝酵素
であるadenine phosphoribosyltransferase(APRT)による adenine 代謝のKmに近いこ
と(0.6-0.9 M)と、本研究では3H 標識体の adenine を用いていることから、APRT によ
る代謝過程(APRT への親和性)を主に反映した見かけの Km が取り込み(3H 標識体の
adenine 及び代謝物の蓄積)過程の解析から評価されている可能性が考えられる。ENBT1 を介して細胞内に取り込まれたadenine が APRT によって代謝されて細胞内に蓄積する一 方で、細胞内 adenine 濃度は低く維持され、細胞外との濃度勾配に依存した促進拡散型の ENBT1介在 adenine 輸送が効率的に進行するものと考えられ、ENBT1と APRT が機能的 協 働 関 係 に あ る と み る こ と が で き る 。 一 方 で 、adenine は guanine 代 謝 に 関わ る hypoxanthine phosphoribosyltransferase 1(HPRT1)に親和性を持たないため、IC50は
ENBT1への作用(ENBT1への親和性)を反映しているものと考えられる。guanine 及び hypoxanthine に関しても、adenine の場合と同様に、ENBT1と HPRT1との機能的協働が 考えられる。
9
4 核酸塩基の細胞内取り込みにおける ENBT1と核酸塩基代謝酵素との機能的協働
ENBT1 と 核 酸 塩 基 代 謝 酵 素 と の 機 能 的 協 働 に つ い て 、 さ ら に 検 証 す る た め 、 APRT/HPRT1欠損の A9細胞(マウス繊維芽細胞由来)を用いて検討を行った(Fig. 4)。ヒ トENBT1のみを一過性に導入した場合には、adenine 取り込みの変化はみられなかったが、 ENBT1をヒト APRT と共に導入した場合には、著しい adenine 取り込みの上昇が認められ た。この結果は、APRT による adenine 代謝に伴い、adenine の濃度勾配が維持されるた め、ENBT1による adenine 輸送が促進され、かつ、放射性同位体標識された adenine の代 謝物が細胞内に蓄積することによると考えられる。また、APRT のみを導入した場合にお いても、adenine 取り込みの増大がみられたが、A9細胞において内因性に発現する核酸塩 基トランスポーターが働いているためであると考えられる。一方で、APRT に代えて guanine の代謝酵素であるヒト HPRT1を導入した場合では、ENBT1の有無に関わらず adenine 取り込みに変化はみられず、ENBT1と協働した APRT の adenine に対する特異的 な作用が確認された。また、guanine に関しても、同様に、ヒト HPRT1との機能的協働に よるとみられるENBT1介在性取り込みの増大が確認された。すなわち、guanine 取り込み は HPRT1の導入により増大し、ENBT1を共導入することによってさらに増大した。一方 で、APRT の導入や ENBT1単独の導入による guanine 取り込みへの影響はなかった。こ れらの結果より、adenine 及び guanine のそれぞれに特異的な代謝酵素の存在下において、 代謝処理に連動して ENBT1による各核酸塩基の輸送が効率的に進行することが示唆され た。
さらに、内因性のadenine 取り込みが認められる HeLa 細胞において、ENBT1及び APRT の特異的siRNA による遺伝子発現抑制が adenine 取り込みに与える影響についても検討し たところ、ENBT1の発現抑制により adenine 取り込みの著しい低下がみられた(70%程度の 低下)一方で、APRT の発現抑制による adenine 取り込みの低下(60%程度の低下)も ENBT1 の発現抑制による効果と同等であった(Fig. 5A)。また、HeLa 細胞における ENBT1及び APRT のタンパク質量をウェスタンブロットにより測定をした結果、ENBT1及び APRT の siRNA の導入により、ENBT1及び APRT の特異的な発現抑制が認められた(Fig. 5B)。こ れらの結果は、HeLa 細胞におけるプリン塩基取り込みの主要な経路として ENBT1が機能 していること、また、salvage 経路における ENBT1を介するプリン塩基の供給(取り込み輸 送)に対する核酸塩基代謝酵素との機能的協働を強く示唆するものである。尚、得られたバ ンドのシグナル強度を定量化した際に、ENBT1の発現抑制によるタンパク質量の低下率 (55%程度の低下)と adenine 取り込みの低下率(70%程度の低下)が乖離する結果とな り、両者の相関を見出すには至らなかった。
10 B A D C Mock ( control ) ENBT1 APRT ENBT1 +APRT [ 3 H] Ad enin e upt ak e (p mol /mg protei n) 0.0 0.1 0.2 0.3 0.4 0.5 Mock ( control ) ENBT1 HPRT1 ENBT1 +HPRT1 [ 3 H] A deni ne up ta ke (pmol/mg pr o tein) 0.0 0.1 0.2 0.3 0.4 0.5 * Mock ( control ) ENBT1 HPRT1 ENBT1 +HPRT 1 [ 3 H] G uan ine u pt ake (p mol/ mg protei n) 0.0 0.1 0.2 0.3 0.4 0.5 * Mock ( control ) ENBT1 APRT ENBT1 +APRT [ 3 H] Gua nine upt ak e (pmol /mg protei n) 0.0 0.1 0.2 0.3 0.4 0.5 † *, *,†
Fig. 4. Cooperative operation of ENBT1 with salvage enzymes for purine nucleobases.
The uptake of [3H]adenine (5 nM) was evaluated at 37°C and pH 7.4 for 1 min in APRT/HPRT1-deficient A9
cells transfected with a plasmid for ENBT1 and one for APRT (A) or HPRT1 (B) with 1 : 1 ratio (1 μg of total plasmid). Each plasmid was replaced with empty pCI-neo vector when it was not used. The uptake of [3H]guanine (5 nM) was evaluated similarly in A9 cells transfected with a plasmid for ENBT1 and one for APRT (C) or HPRT1 (D). *Significantly different from control at p < 0.05. †Significantly different from APRT or HPRT1 alone at p < 0.05. All the data are presented as the means ± SE (n = 3 or 4).
11 Controlsi-ENBT1 si-APRT
Upt ak e (% of c ont rol) 0 20 40 60 80 100 120
*
*
A BControl si-ENBT1 si-APRT
ENBT1 -Actin APRT -Actin 1.00 1.45 0.463 1.34 0.447 1.00
Fig. 5. Effect of silencing of ENBT1 and APRT on [3H]adenine uptake in HeLa cells.
(A) The uptake of [3H]adenine (5 nM) was evaluated at 37°C and pH 7.4 for 1 min in HeLa cells transfected with siRNA for ENBT1 or APRT. *Significantly different from control at p < 0.05. Data are presented as the means ± SE (n = 4). (B) The endogenous protein levels for ENBT1, APRT and β-actin in those cells were analyzed by western blotting. The bands of ENBT1, APRT and β-actin were detected at 54 kDa, 30 kDa and 42 kDa, respectively. The ratio of protein level of ENBT1/β-actin, as determined by NIH-Image software, is shown under each lane. Western blots shows the representative results in at least two independent experiments.
14 6 まとめ 機能未知のトランスポーター様タンパク質として報告されていた ENBT1を核酸塩基ト ランスポーターとして見出した。ENBT1は adenine、guanine、hypoxanthine 等のプリン 塩基に特異的に高い輸送活性を示し、その輸送様式は促進拡散型であることが示唆された。 また、ENBT1による adenine 輸送に対する阻害実験から、プリン塩基及びその誘導体によ る顕著な阻害効果が認められたほか、decynium-22が ENBT1に極めて高い阻害活性を示す ことが明らかとなった。さらに、ENBT1に対する adenine、guanine、hypoxanthine の Kmは、それぞれ0.94 μM、1.70 μM、1.32 μM と算出された。その一方で IC50は、それぞ れ13 μM、70 μM、350 μM と算出され、Kmと大きく異なる結果となった。このKm とIC50 の乖離は、細胞内に存在する核酸塩基酵素によるプリン塩基の代謝過程がENBT1によるプ リン塩基輸送に寄与する可能性を示唆するものであり、今回得られたKmは核酸塩基代謝酵 素に対する親和性を、IC50はENBT1に対する親和性を反映しているものと考えられる。核 酸塩基代謝酵素であるAPRT 及び HPRT を欠損する A9細胞において、ENBT1によるプリ ン塩基取り込みはAPRT 及び HPRT1との共発現により顕著に増大し、また、内因性の核酸 塩基輸送の認められるHeLa 細胞においても、APRT の発現抑制による adenine 取り込み の低下は、ENBT1の発現抑制による adenine 取り込みの低下に匹敵したことから、ENBT1 と核酸塩基代謝酵素との機能的協働がさらに強く示唆された。 ヒトにおけるENBT1遺伝子の発現は様々な臓器において認められ、特に肝臓及び肺にお いて高いことが明らかとなった。肝臓におけるENBT1の細胞内局在は、肝実質細胞の血管 側膜である類洞膜に局在することが明らかとなった。 肝臓が核酸(ヌクレオチドや核酸塩基)の de novo 合成を盛んに行っている臓器である ことを考えると、肝臓でde novo 合成により生成された核酸塩基の一部(過剰分)は ENBT1 を介して血液へと排出されると考えられる。血液中に供給された核酸塩基が ENBT1と salvage 酵素の協働的作用によって効率的に肺をはじめとする細胞損傷の修復の盛んな各 種臓器の細胞内に取り込まれることで、salvage 合成に必要な核酸塩基の需要を満たしてい ることが推察される。
15 第2章 HSV-TK/GCV自殺遺伝子治療におけるENBT1の役割:ENBT1と代謝酵素との 機能的協働の応用利用 1 ENBT1によるGCV輸送の解析 ウイルス由来のリン酸化酵素による代謝に始まる活性化過程を経て作用する抗ウイル ス薬として知られるGCVは、宿主細胞に対する殺細胞効果も示す31)。一方で、抗ウイル ス治療では副作用となるこの宿主細胞への作用をがん治療に応用する試みとして、がん組 織の細胞特異的にHSV-TKの遺伝子を導入し、発現させた上で、GCVを投与して殺細胞 作用を惹起させる治療法(HSV-TK/GCV自殺遺伝子治療法)が提唱されており32-34)、非 がん組織への副作用を回避できる治療法として期待されているが、その有効性は十分には 検証されていない35-39)。そこで、この治療法の前提となるGCVの細胞内取り込みのメカ ニズムが未解明であること40-43)及び、プリン核酸塩基類似の化学構造を持つGCVの取り 込みにENBT1が関与する可能性があることに着目し、ENBT1と代謝酵素の機能的協働の 応用利用の観点から、ENBT1によるGCV輸送及びHSV-TK/GCV自殺遺伝子治療におけ るENBT1の役割を探ることにした。 まず、ENBT1のGCV輸送能を探るため、ENBT1を一過性発現させたHEK293細胞に おいてGCV取り込みの解析を行った。その結果、ENBT1の導入によりGCVの取り込みは、 mock細胞での取り込みの2.5倍程度に増大したことから、ENBT1のGCVに対する輸送能 が見出された。比較のため、一部の核酸塩基類に対する輸送能が知られているENT1及び ENT2に関しても同様の解析を行ったが、これらはGCVに対する輸送活性を示さなかった (Fig. 8)。
Moc
k (co
ntro
l)
ENB
T1 ENT1 ENT2
U
ptake (pmo
l/mg protein
)
0.0
0.1
0.2
0.3
0.4
0.5
0.6
*
Fig. 8. GCV uptake in HEK293 cells transiently expressing ENBT1, ENT1 or ENT2
The uptake of [3H]ganciclovir (60 nM) was evaluated for 5 min at 37°C and pH 7.4. Data are presented as means ± SE (n = 4). *, p < 0.05 compared with the values at control.
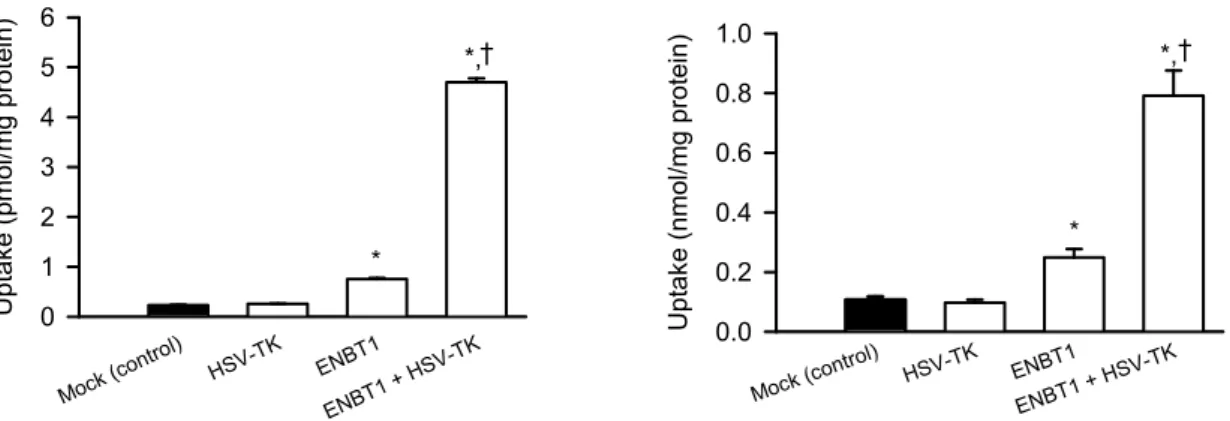
18 2 GCV の細胞内取り込みにおける ENBT1と HSV-TK との機能的協働 ENBT1の GCV 輸送能が見出されたことより、ENBT1と HSV-TK との機能的協働の解 析を行った(Fig. 11)。HEK293細胞において、HSV-TK のみを一過性に導入した場合には、 GCV 取り込みは変化しなかったが、ENBT1を HSV-TK と共に導入した場合には、ENBT1 のみを導入した場合を上回る GCV 取り込みが認められ、HSV-TK との機能的協働による ENBT1介在性 GCV 取り込みの増大が示唆された。この結果は、HSV-TK による GCV の 代謝により、細胞内GCV 濃度が低く維持され、ENBT1を介する促進拡散型の GCV 輸送が 効率的に進行すると共に、GCV 代謝物の細胞内蓄積が進んだことによるものと考えられる。 さらに、ENBT1及び HSV-TK を共に安定発現させた MDCKII 細胞(ENBT1/HSV-TK 細 胞)において、臨床で想定される水準のGCV の血中濃度(30 M)と同濃度で、同様に両 者の機能的協働によるGCV 取り込みの増大が確認された(Fig. 12)。また、ENBT1/HSV-TK 細胞においてのみ、GCV 存在下での72 h の培養後の細胞生存率の低下(殺細胞効果)が認 められた(Fig. 13)。この殺細胞効果は、GCV 濃度依存的に強くなり、30 M で最大となっ た(70%程度の生存率低下)。一方で、HSV-TK または ENBT1のみを導入した細胞では、 mock 細胞の場合と同様に、試験濃度の全範囲(100 M まで)において、殺細胞効果はみ られなかった。 Mock ( control ) HSV-T K ENBT1 ENBT1 + HSV -TK Upt a ke ( pm ol /m g pr ot ei n) 0 1 2 3 4 5 6 * *,† Mock (co ntrol) HSV-TK ENBT1 ENBT1 + HSV-TK Uptak e (nmol /mg protein ) 0.0 0.2 0.4 0.6 0.8 1.0 * *,†
Fig. 11. Effect of transient expression of ENBT1 and HSV-TK on GCV uptake in HEK293 cells
The uptake of [3H]ganciclovir (60 nM) was evaluated for 5 min at 37C and pH 7.4. HEK293 cells were transiently transfected with a plasmid for ENBT1 and one for HSV-TK with 1:1 ratio (1 μg of total plasmid). Each plasmid was replaced with empty pCI-neo vector when it was not used. Data are presented as means ± SE (n = 4). *, p < 0.05 compared with mock; †, p < 0.05 compared with ENBT1.
Fig. 12. Effect of stable expression of ENBT1 and HSV-TK on GCV uptake in MDCKII cells
The uptake of [3H]ganciclovir (30 μM) was evaluated for 60 min at 37°C and pH 7.4 in MDCKII cells stably expressing HSV-TK, ENBT1 or ENBT1/HSV-TK. Data are presented as means ± SE (n = 4). *, p < 0.05 compared with mock; †, p < 0.05 compared with ENBT1.
22 3 各種がん由来細胞株における HSV-TK 一過性導入が GCV 取り込み及び殺細胞効果に及 ぼす影響 ここまでの結果から、ENBT1は GCV の細胞内取り込み経路として、その殺細胞効果の 惹起に大きく関与するものと考えられる。ENBT1以外の GCV トランスポーターが関与す る場合がある可能性を否定できないが、少なくとも、がん細胞におけるENBT1の発現の有 無(程度)は、GCV の殺細胞効果に影響し、HSV-TK/GCV 自殺遺伝子治療の成否に関わ る要因となり得ると考えられる。そこで、この仮説の検証のため、種々のがん由来細胞株 を用い、HSV-TK の一過性導入の効果を比較検討することにした。
まず、FLAG 発現 vector に組み込んだ HSV-TK(HSV-TK/FLAG)を一過性発現させた 各種がん由来細胞株におけるENBT1及び HSV-TK/FLAG の発現を、ウェスタンブロット により解析した(Fig. 17)。その結果、ENBT1の発現は、肺がん由来 A549細胞、結腸がん由 来HCT-15細胞、乳がん由来 MCF-7細胞では低い一方で、腎がん由来 Caki-1細胞、子宮頸 がん由来HeLa 細胞、肝がん由来 HepG2細胞においては ENBT1の発現が高いことが明ら かとなった。以後、これらの細胞株を、ENBT1高発現細胞(Caki-1、HeLa、HepG2)及 びENBT1低発現細胞(A549、HCT-15、MCF-7)と称することにした。尚、ENBT1高発 現細胞においてENBT1の発現を示すバンドが2つ認められたが、ENBT1には long form(60 kDa, NM_001278206.1)と short form(55 kDa, NM_001278201.1)の2つの splicing variant が存在するためだと考えられる。また、HSV-TK/FLAG の発現は、A549、HCT-15、 MCF-7、HeLa 細胞では同程度であったが、これらと比較し、Caki-1細胞でやや低く、HepG2 細胞ではかなり低かった。
A549 HCT-15 MCF-7 Caki-1 HeLa HepG2
ENBT1
HSV-TK
-actin
ENBT1 ENBT1
Fig. 17. Western blot analysis of endogenous ENBT1 and FLAG-tagged HSV-TK transiently expressed in various cancer cell lines
The protein levels of endogenous ENBT1 and β-actin, and exogenous HSV-TK/FLAG in various cancer cell lines were analyzed by Western blotting. Western blots shows the representative results in at least two independent experiments.
23
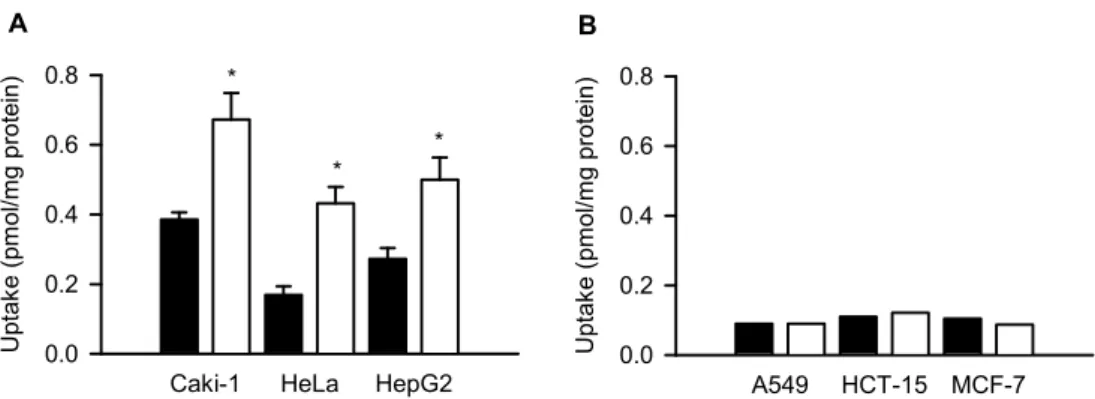
続いて、これらのがん由来細胞株におけるHSV-TK 一過性導入が GCV 取り込み及び GCV 処理による殺細胞効果に与える影響について解析した(Figs. 18 and 19)。その結果、ENBT1 高発現細胞では、GCV 取り込みの増大と細胞生存率の低下がみられた一方で、ENBT1低発 現型細胞では、いずれの変化もほとんどみられず、ENBT1の有無(発現レベル)が GCV の効果に関わるという仮説の妥当性が示唆された。尚、HSV-TK の発現効率の低い Caki-1 及びHepG2の両細胞株において、HSV-TK の一過性導入による GCV 取り込み及び殺細胞 効果の上昇が認められた点からも、HSV-TK の発現寮と比較して、ENBT1の発現量が HSV-TK/GCV 自殺遺伝子治療の効果に対してより強く影響する可能性が考えられる。
Caki-1 HeLa HepG2
Up ta ke (p m ol /m g prot ei n) 0.0 0.2 0.4 0.6 0.8 * * * A A549 HCT-15 MCF-7 Up ta ke ( pm ol /m g prot ei n) 0.0 0.2 0.4 0.6 0.8 B
Fig. 18. Effect of transient introduction of HSV-TK on GCV uptake in various cancer cell lines
The uptake of [3H]GCV (30 μM) was evaluated at 37°C and pH 7.4 for 5 min in various cancer cells
transiently expressing HSV-TK (open bars) and mock cells (filled bars) for control. Data are presented as means ± SE (n = 4). *, p < 0.05 compared with control by Student’s t-test.
Caki-1 HeLa HepG2
Cell vi ab ility ( % o f c on tr ol) 0 20 40 60 80 100 * * * A A549 HCT-15 MCF-7 Cell v ia bility ( % o f c on tr ol) 0 20 40 60 80 100 120 * B
Fig. 19. Effect of transient introduction of HSV-TK on GCV-induced cytotoxicity in various cancer cell lines
Cells were cultured for 72 h in the presence or absence (control, filled bars) of ganciclovir (30 μM, open bars) and the numbers of cells were determined by MTT assay. Data are presented as means ± SE (n = 4). *, p < 0.05 compared with the value for control by Student’s t-test.
24
さらに、HSV-TK 導入の効果が最も顕著であった HeLa 細胞を用いた試験では、HSV-TK の存在下において(一過性導入)、ENBT1特異的な siRNA を用いて ENBT1遺伝子発現抑 制を行うことにより、GCV の取り込み及び殺細胞効果が低下することも確認できた(Fig. 20)。これにより、HeLa 細胞における HSV-TK 導入による GCV の効果の惹起において、 ENBT1が寄与することが明らかとなった。以上より、がん細胞における ENBT1の発現が HSV-TK/GCV 自殺遺伝子治療の効果に大きく関与する可能性が示唆された。 Uptak e (nmol/mg pro tein) 0.00 0.05 0.10 0.15 0.20 0.25 * * HSVTK + si-ENBT 1 HSVTK + si-contr ol pCI-neo + si-con trol pCI-neo + si-EN BT1 A C el l v ia bi lit y (% o f c on tr ol) 0 20 40 60 80 100 120 140 * * HSVTK + si-ENBT 1 HSVTK + si-contr ol pCI-neo + si-con trol pCI-neo + si-EN BT1 B
sicontrol siENBT1sicontrolsiENBT1
ENBT1
HSV-TK
-actin
C
Fig. 20. Effect of silencing of ENBT1 on GCV uptake and GCV-induced cytotoxicity in HeLa cells transiently expressing HSV-TK(A) Mean ± SE (n = 4); uptake of [3H]ganciclovir (30 μM) for 5 min at 37°C and pH 7.4; *, p < 0.05. (B) Mean ± SE (n = 4); cells were cultured for 72 h in the presence or absence (control) of ganciclovir (30 μM) and the numbers of cells were determined by MTT assay; *, p < 0.05.
(C) The protein levels of endogenous ENBT1 and β-actin, and exogenous HSV-TK/FLAG in HeLa cells were analyzed by western blotting.
HSV-TK/FLAG was used in this set of experiments to assess its expression by probing for FLAG in Western blot analysis. Western blots shows the representative results in at least two independent experiments.
26 第3章 ENBT1特異的阻害薬としてのdecynium-22の特性 1 ENBT1によるadenine輸送に対するdecynium-22の阻害特性:ENT1及びENT2によ るadenine輸送との比較 各種の組織由来細胞等での核酸塩基関連医薬品等の取り込みにおける ENBT1の寄与に 関する情報は、その種の医薬品等の体内動態特性を的確に把握し、開発及び使用を効率的 ないし効果的に行う上で有用である。また、一般に、薬物等の細胞膜輸送における各種ト ランスポーターの寄与を探る目的で、特異的阻害剤による阻害率を指標とする手法が汎用 されている。そこで、ENBT1に関して、特異的阻害剤の検索及びその阻害特性の把握する ことにした。 核酸塩基関連医薬品等の細胞膜輸送におけるENBT1の寄与の評価に際しては、一部の核 酸塩基等に対する輸送活性が知られている ENT1及び ENT2の寄与との識別が特に重要と なると考えられる。第一章では、各種薬物等を用いた一連の ENBT1阻害試験に行い、 ENT1/2の特異的阻害剤として知られる dipyridamol(200 M)24,25)が阻害活性を示さず、 またENT1の特異的阻害剤として知られる NBMPR(200 M)24,25)の阻害活性もかなり弱 い一方で、核酸塩基輸送の阻害剤として知られる decynium-22(10 M)が強い阻害活性 を示すことが見出された。そこで、decynium-22の ENBT1特異的阻害薬として有用である かを解析した。 decynium-22の各種核酸トランスポーターに対する阻害効果の解析に先立ち、adenine を モデル核酸塩基基質として、ENBT1及び ENT1、ENT2一過性発現 HEK293細胞における 輸送解析を行った(Fig. 22)。ヒト ENBT1(hENBT1)発現細胞での adenine 取り込みは、 1分まで取り込み時間にほぼ比例して増大した。そこで、hENBT1による取り込み初速度の 評価のための取り込み時間を1分と設定し、以後の実験を行った。ヒト ENT2(hENT2)に ついても、同様に、取り込み時間を1分と設定した。また、マウス ENBT1(mENBT1)及 びマウス ENT2(mENT2)に関しても、同様に取り込み時間を1分と設定した。ヒト及び マウス ENT1については、HEK293細胞での一過性発現における取り込み時間1分での adenine 取り込みに関して有意な増大が認められず、adenine 輸送能を持たないことが示唆 された。
続いて、ヒト及びマウスのENBT1及び ENT2による adenine 輸送の濃度依存性に関する 検討を行った(Table 2)。いずれのトランスポーターに関しても5 nM から100 nM の範囲で 取り込み速度/濃度比は一定となり、Km よりも低濃度で、取り込み速度が濃度に比例する
速度論的線形領域にあることが確認された。これに基づき、5 nM の adenine 濃度設定で、 以後の阻害試験を行うことにした。
29
Uptake rate (% of control)
0
20
40
60
80 100 120 140
Control Decynium-22 10 M Decynium-22 100 M Dipyridamol 10 M
*
Fig. 24. Effects of decynium-22 and dipyridamole on uridine uptake by hENT1 transiently expressed in HEK293 cells transiently expressing hENT1
The specific uptake rate of [3H]uridine (5 nM) was evaluated for 1 min at 37°C and pH 7.4 in the presence of decynium-22 (10 µM or 100 µM), dipyridamole (10 µM), or in their absence. The control value was 0.194 pmol/min/mg protein. Data are presented as means ± SE (n = 4). *, p < 0.05 compared with control.
31
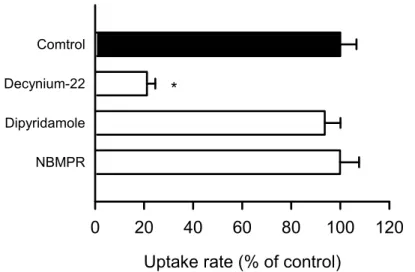
Uptake rate (% of control)
0
20
40
60
80
100
120
NBMPR Dipyridamole Decynium-22 Comtrol*
Fig. 26. Effect of decynium-22, dipyridamol and NBMPR on the carrier-mediated component of adenine uptake in HepG2 cells
The uptake rate of [3H]adenine (5 nM) was evaluated for 1 min at 37°C and pH 7.4 in the presence of decynium-22 (10 µM), dipyridamol (10 µM) or NBMPR (100 nM), or in their absence and the nonsaturable component of the uptake rate was subtracted from that. The control value was 0.177 pmol/min/mg protein. Data are presented as means ± SE (n = 4). *, p < 0.05 compared with control.
32 3 まとめ
hENBT1及び hENT2による adenine 輸送に対する decynium-22のIC50は、それぞれ2.90
M 及び111 M と算出され、大きく異なった。さらに、mENBT1及び mENT2に対する decynium-22のIC50は、それぞれ2.30 M 及び134 M と得られ、ヒトの場合と同程度の阻 害活性及び差異を示した。一方で、adenine に対する輸送活性の認められなかった hENT1 においては、そのuridine 輸送に対する decynium-22のIC50は100 M を大きく上回ること が明らかになった。これより、核酸塩基輸送において、10 M の decynium-22は ENBT1 の選択的かつ強力な阻害薬として働き、ENBT1の寄与の把握に役立つものと考えられる。 また、ENBT1の発現が認められる HepG2細胞における adenine 輸送では、担体輸送の関 与が大きく、10 M の decynium-22によりその担体輸送はほぼ完全に阻害された一方で、 ENT1/2を完全に阻害する10 M の dipyridamole は阻害効果を示さなかったことから、そ の担体輸送のほとんどがhENBT1の寄与によるものであることが示唆された。 decynium-22の薬理作用としては、dopamine や serotonin などのモノアミンの再取り込 みに対する阻害作用及び抗うつ様作用を示すことが報告されている46,47)。現在抗うつ薬とし て用いられている三環系抗うつ薬、四環系抗うつ薬及び選択的セロトニン再取り込み阻害 薬は、モノアミンの再取り込みを阻害することで抗うつ作用を示す。decynium-22とこれら の抗うつ薬との間には複数の環構造からなる化学構造を有するという共通点が存在するこ とからも、decynium-22は抗うつ作用を持つ可能性は高い。さらに、三環系抗うつ薬である amitriptyline や四環系抗うつ薬である mianserin、選択的セロトニン再取り込み阻害薬で あるparoxetine 等は、鎮痛作用を有するため、慢性疼痛に対して適応されている。近年、 paroxetine は、慢性疼痛の一種である神経障害性疼痛抑制作用を有することが報告されて おり、paroxetine による神経障害性疼痛抑制作用はセロトニン神経非存在下においても認 められることから、モノアミンの再取り込みの阻害とは異なる作用機序で生じている可能 性が高い48)。従って、ENBT1を標的とした阻害薬は、うつ病及び疼痛をはじめとする炎症 性疾患に対する新たな治療薬となる可能性が考えられる。
33
結論
本研究では、ENBT1を核酸塩基トランスポーターとして同定し、その輸送機能解析と合 わせて、核酸塩基利用における細胞内核酸塩基代謝酵素との協働的機能の解析、協働的機 能の医療面での応用利用等に取り組み、以下の成果を挙げることができた。 1. ENBT1を新たに核酸塩基トランスポーターとして同定し、プリン核酸塩基を特異的に 認識し、促進拡散型の輸送様式で機能することを明らかにした。さらに、細胞内の核酸 塩基代謝酵素と機能的協働関係にあり、代謝産物の生成・蓄積と連動して、細胞内核酸 塩基濃度が低く維持されることで、細胞外との濃度勾配に依存した促進拡散型の ENBT1介在核酸塩基輸送が効率的に進行することが示唆された。また、ENBT1の生理 的な役割としては、de novo経路が活発に働く肝臓(実質細胞)から血中への核酸塩基 の排出輸送経路(供給経路)としての可能性が注目される。一方で、核酸塩基の供給を 受ける側の諸臓器では、ENBT1が核酸塩基の取り込み輸送経路として働き、細胞内の 核酸代謝酵素と連携して構成されるsalvage経路の最初の段階の役割を担っているもの と考えられる。 2. プリン核酸塩基類似の化学構造を持つ GCV の取り込みに ENBT1が関与する可能性があ ることに着目し、ENBT1と代謝酵素の機能的協働の応用利用の観点から、ENBT1による GCV 輸送及び HSV-TK/GCV 自殺遺伝子治療における ENBT1の役割を探ることにも取 り組んだ。これにより、ENBT1が高い GCV 輸送能を持つことが明らかとなり、GCV の 代謝活性化及び殺細胞効果の惹起につながるGCV の供給経路としての ENBT1の重要性 が示唆された。従って、ENBT1を高発現するがん種・細胞に HSV-TK/GCV 自殺遺伝子 治療を適応することで、よりがん選択的な細胞傷害性を有し、副作用の小さいがん治療 法となることが期待される。 3. 核 酸 塩 基 関 連 医 薬 品 等 の 細 胞 膜 輸 送 に お け る ENBT1 の 特 異 的 阻 害 薬 と し て 、 decynium-22を発見した。この知見は、核酸塩基関連医薬品等の細胞内取り込みにおけ るENBT1の寄与を評価するために有用であると考えられる。 本研究の成果は、先天性の HPRT1の先天的欠損による遺伝子疾患であるレッシュ・ナイハン 症候群等の核酸代謝の異常を端とするような病態の解析や、ウイルス感染症やがん・免疫系 疾患の治療の用いられる核酸塩基類似医薬品等の開発及び使用の最適化のための基礎情報と して有用と考えられる。34
実験の部
1 実験材料
1 試薬
[3H]adenine(25.0 Ci/mmol)、[3H]hypoxanthine (27.0 Ci/mmol) 、[3H]uracil(42.8
Ci/mmol)、 [3H]uridine(30 Ci/mmol)、[3H]xanthine(12.8 Ci/mmol)、[3H]adenosine (39.2
Ci/mmol)、[3H]acyclovir (15.3 Ci/mmol)、[3H]ganciclovir (6.9 Ci/mmol)は、Moravek
Biochemicals(Brea, CA, USA)製のものを購入して用いた。[14C]guanine hydrochloride
(55 mCi/mmol) は American Radiolabeled Chemicals (St. Louis, MO, USA) 製のものを 購入して用いた。[14C]ascorbate (8.5 mCi/mmol) は PerkinElmer Life Sciences (Boston,
MA, USA) 製のものを購入して用いた。シンチレーションカクテルは、Nacalai Tesque (Kyoto, Japan) 製の Clear-sol I を用いた。また、遺伝子操作用キットとして、TRI REAGENT® BD (Sigma-Aldrich, St. Louis, MO, USA)、QIAEX® Ⅱ Gel Extraction Kit
(QIAGEN, Valensia, CA, USA)、PureLink™ HiPure Plasmid Midiprep Kit (Invitrogen,
Carsbad, CA, USA)を使用した。DNA polymerase としては、Toyobo (Osaka, Japan)製の KOD-FX もしくは KOD-Plus Neo を、ベクターは Promega (San Luis Obispo, CA, USA) 製のpCI-neo、pCI-neo-N-FLAG、Clontech (Palo Alto, CA, USA)製の pEGFP-C1、Takara Bio(Ohtsu, Japan)製の pIRESpuro3 vector を用いた。なお、pCI-neo-N-FLAG vector に 関しては、pCI-neo vector を NheI と XhoI の制限酵素で処理し、DYKDDDDK (5’- GACTACAAGGACGACGATGACAAG -3’)の配列を組み込むことにより作製した。制限酵 素はすべて Toyobo 製のものを用い、配列の組み込みには T4 DNA Ligase (Fermentas, Ontario, Canada)及び Ligation-Convenience Kit (Nippon gene, Tokyo, Japan)を用いた。 また、ヒトのcDNA ライブラリを作製するにあたり、ヒト組織の total RNA は Clontech から購入し、逆転写酵素はReverTra Ace (Toyobo)を用いた。その他の試薬は全て特級品を 使用した。免疫染色には、Human Normal Adult Liver Frozen Tissue Section (BioChain Institute, Newark, CA, USA)を購入して用いた。その他の試薬は、全て特級品を使用した。
2 培養細胞
HEK293、MDCKII、A9、HeLa、HepG2、Caki-1、A549、HCT-15、MCF-7の各細胞 は、すべて東北大学加齢医学研究所医用細胞資源センター(Sendai, Japan)より提供を受け た。
35 2 実験方法
1 各種遺伝子のクローニング
1-1 human equilibrative nucleobase transporter 1 (hENBT1)
hENBT1 (GenBank accession no, NM_017611.2)のクローニングには、human lung total RNA を用いた。ReverTra Ace を用いて total RNA から逆転写を行うことにより cDNA を得た。この cDNA を鋳型とし、hENBT1の cDNA を得るために PCR (LittleGene TC-25/H, BioFlux)を行った。DNA polymerase としては KOD-Plus DNA polymerase を用 い、プライマーとしては以下のものを使用した。
forward primer: 5’-ATTTTCCAAGTGCTCAAACGC-3’ reverse primer: 5’-CTGCCAAGGCTAAGTGCAAGG-3’
これによってできたPCR 産物を鋳型として、制限酵素サイトをデザインしたプライマー を用い、再びPCR を行った。プライマーとしては以下のものを使用した。
forward primer, +EcoRI site (下線) : 5’-AGGAATTCTGCTCATGGCGGGCCA-3’ reverse primer, +XbaI site (下線): 5’-GCTCTAGAACTATGCAATTGCAGA-3’
得られたPCR 産物を、制限酵素 EcoRI と XbaI で処理し、その後、電気泳動し QIAEX®II Gel Extraction Kit を用いて精製、抽出した。哺乳類発現ベクターである pCI-neo vector も同様の 制限酵素処理を行った後、精製した。これらをT4 DNA Ligase を用いてライゲーションし、 ヒ ー ト シ ョ ッ ク (42C、1分) によって大腸菌 (DH5) に導入した。その大腸菌を ampicillin 添加した LB プレートに播き、37C で一晩 (約12時間) 培養した後、プレート 上のコロニーを採取した。そこからプラスミドDNA を抽出し、制限酵素処理後、電気泳動 を行うことによりインサート部分を確認した。予想された長さのインサート部分が確認さ れたコロニーから得られたプラスミドは、シークエンスを確認し、インサート部分の塩基 配列に変異がないことを確認した (ABI PRISM® 3100-Avant Genetic Analyzer)。なお、大
腸菌 (DH5) のコンピテントセルは、Inoue らの方法49)に準じて作製したものを用いた。
また、Green fluorescent protein (GFP) 遺伝子を組み込んだ pEGFP-C1 vector も同様 の制限酵素処理を行い、hENBT1の cDNA を導入し、GFP-hENBT1プラスミドを作製した。
1-2 human adenine phosphoribosyltransferase (hAPRT)
hAPRT の遺伝子導入用プラスミドの作製にあたっては、human liver total RNA から ReverTra Ace を用いて逆転写することによって cDNA を得た。この cDNA を鋳型とし、 hAPRT (GenBank accession no.,NM_000485.2)の cDNA を得るために PCR を行った。
36
polymerase は KOD-plus-neo DNA polymerase を用い、プライマーは以下のものを使用し た。
forward primer: 5’-CTGCCGCTGGCTCTTCGCACG-3’ reverse primer: 5’-GCAGCCGGTGCCCCTGGTCACT-3’
これによってできたPCR 産物を鋳型として、制限酵素サイトをデザインしたプライマー を用い、再びPCR を行った。プライマーとしては以下のものを使用した。
forward primer, +EcoRI site (下線) :5’-ATGAATTCAGCCATGGCCGACTCC-3’ reverse primer, +XbaI site (下線): 5’-GACTCTAGAGAGGCCCTGTGGTCA-5’
以後、hENBT1の場合と同様の手順で操作を行い、pCI-neo vector に組み込んだプラス ミドを得た。
1-3 human hypoxanthine-guanine phosphoribosyltransferase 1 (hHPRT1)
hHPRT1の遺伝子導入用プラスミドの作製にあたっては、human liver total RNA から ReverTra Ace を用いて逆転写することによって cDNA を得た。この cDNA を鋳型とし、 hHPRT1 (GenBank accession no.,NM_000194.2)の cDNA を得るために PCR を行った。 polymerase は KOD-plus-neo DNA polymerase を用い、プライマーは以下のものを使用し た。
forward primer: 5’-CCTCCTCCTGAGCAGTCAGC-3’ reverse primer: 5’-TTTAGGAATGCAGCAACTGACA-3’
これによってできたPCR 産物を鋳型として、制限酵素サイトをデザインしたプライマー を用い、再びPCR を行った。プライマーとしては以下のものを使用した。
forward primer, +EcoRI site (下線) :5’-AGTGAATTCCGTTATGGCGACCCGCA-3’ reverse primer, +XbaI site (下線): 5’-GCCTCTAGAACATTGATAATTTTAC-3’
以後、hENBT1の場合と同様の手順で操作を行い、pCI-neo vector に組み込んだプラス ミドを得た。
1-4 human equilibrative nucleoside transporter 1 (hENT1)
pcDNA3.1 vector (Invirogen) に 組 み 込 ま れ た hENT1 (GenBank accession no, NM_001078175.1)のプラスミドは、北海道大学大学院薬学研究科 薬物動態解析学研究室 菅原 満 教授より提供されたものを用いた50)。このプラスミドを制限酵素EcoRI と XbaI
で処理し、その後、電気泳動しQIAEX®Ⅱ Gel Extraction Kit を用いて精製、抽出した。
哺乳類発現ベクターであるpCI-neo vector も同様の制限酵素処理を行った後、精製した。 以後、hENBT1の場合と同様の処理により、pCI-neo vector に組み込んだプラスミドを得
37 た。
1-5 human equilibrative nucleoside transporter 2 (hENT2)
pcDNA3.1 vector に組み込まれた hENT2 (GenBank accession no, NM_001532.2)のプ ラスミドは、北海道大学大学院薬学研究科 薬物動態解析学研究室 菅原 満 教授より 提供されたものを用いた。このプラスミドを鋳型として、制限酵素サイトをデザインした プライマーを用い、PCR を行った。プライマーとしては以下のものを使用した。
forward primer, +EcoRI site (下線) : 5’-GACGAATTCGGCCATGGCGCGAGGAGA-3’ reverse primer, +XbaI site (下線): 5’-GGCATCTAGATCAGAGCAGCGCCTTGAA-3’ 制限酵素EcoRI と XbaI で処理し、その後、電気泳動し QIAEX®Ⅱ Gel Extraction Kit
を用いて精製、抽出した。哺乳類発現ベクターであるpCI-neo vector も同様の制限酵素処 理を行った後、精製した。
以後、hENBT1の場合と同様の処理により、pCI-neo vector に組み込んだプラスミドを 得た。
1-6 mouse equilibrative nucleobase transporter 1 (mENBT1)
mENBT1の遺伝子導入用プラスミドの作製にあたっては、A9細胞(マウス線維芽細胞由 来)からグアニジン-イソチオシアネート法にて抽出したtotal RNA から、ReverTra Ace を用いて逆転写することによって cDNA を得た。この cDNA を鋳型とし、mENBT1 (GenBank accession no.,NM_000194.2)の cDNA を得るために PCR を行った。polymerase はKOD-plus-neo DNA polymerase を用い、プライマーは以下のものを使用した。
forward primer: 5’-CAGTGGTCCTCTCTGAAGCTC-3’ reverse primer: 5’-GAAAAGGCATCATCGAGGCTA-3’
これによってできたPCR 産物を鋳型として、制限酵素サイトをデザインしたプライマー を用い、再びPCR を行った。プライマーとしては以下のものを使用した。
forward primer, +EcoRI site (下線) : 5’-GCAGAATTCGACCATGGCAAGCAAG-3’ reverse primer, +XbaI site (下線): 5’-CTGTCTAGACTAGAGAGTGGACTA-3’
以後、hENBT1の場合と同様の手順で操作を行い、pCI-neo vector に組み込んだプラス ミドを得た。
1-7 mouse equilibrative nucleoside transporter 1 (mENT1)
mENT1の遺伝子導入用プラスミドの作製にあたっては、A9細胞(マウス線維芽細胞由来) からグアニジン-イソチオシアネート法にて抽出したtotal RNA から、ReverTra Ace を用
38
いて逆転写することによってcDNA を得た。この cDNA を鋳型とし、mENT1 (GenBank accession no., NM_001199113.1)の cDNA を得るために PCR を行った。polymerase は KOD-plus-neo DNA polymerase を用い、プライマーは以下のものを使用した。
forward primer: 5’-TCACCGTCCTGCCAACATCG-3’ reverse primer: 5’-ATTGAGCCCTTGCCATTCTGAC-3’
これによってできたPCR 産物を鋳型として、制限酵素サイトをデザインしたプライマー を用い、再びPCR を行った。プライマーとしては以下のものを使用した。
forward primer, +SalI site (下線) : 5’-GGAGTCGACAGCCATGACAACCAGT-3’ reverse primer, +NotI site (下線): 5’CTTGCGGCCGCGTCCCCACAGGGTCA-3’ 以後、hENBT1の場合と同様の手順で操作を行い、pCI-neo vector に組み込んだプラス ミドを得た。
1-8 mouse equilibrative nucleoside transporter 2 (mENT2)
mENT2の遺伝子導入用プラスミドの作製にあたっては、A9細胞(マウス線維芽細胞由来) からグアニジン-イソチオシアネート法にて抽出したtotal RNA から、ReverTra Ace を用 いて逆転写することによってcDNA を得た。この cDNA を鋳型とし、mENT2 (GenBank accession no., NM_000194.2)の cDNA を得るために PCR を行った。polymerase は KOD-plus-neo DNA polymerase を用い、プライマーは以下のものを使用した。
forward primer: 5’-GCAATAGGACTGCGGCAACCA-3’ reverse primer: 5’-TCCTTCCCTGTGAACGTCCCA-3’
これによってできたPCR 産物を鋳型として、制限酵素サイトをデザインしたプライマー を用い、再びPCR を行った。プライマーとしては以下のものを使用した。
forward primer, +EcoRI site (下線) : 5’-CAAGAATTCTACCATGGCGCGGGGA-3’ reverse primer, +XbaI site (下線): 5’-CAGTCTAGACACTGGTGGTCCTCA-3’
以後、hENBT1の場合と同様の手順で操作を行い、pCI-neo vector に組み込んだプラス ミドを得た。
1-9 human herpesvirus 1 thymidine kinase (HSV-TK)
HSV-TK (GenBank accession no, AF057310.1) の 遺 伝 子 に つ い て は 、 Operon Biotechnology (Tokyo, Japan)において合成されたものを、pUC57 vector に組み込まれた プラスミドとして入手した。
このプラスミドを制限酵素EcoRI と XbaI で処理し、その後、電気泳動し QIAEX®Ⅱ Gel
Extraction Kit を用いて精製、抽出した。哺乳類発現ベクターである pCI-neo vector も同 様の制限酵素処理を行った後、精製した。
39
以後、hENBT1の場合と同様の処理により、pCI-neo vector 及び pCI-neo-N-FLAG vector に組み込んだプラスミドを得た。
また、HSV-TK の cDNA を pIRESpuro3 vector に組み込んだプラスミドを作製した。そ の作製に当たっては、HSV-TK の cDNA を pCI-neo vector に組み込んだプラスミドを NheI とNotI で処理して HSV-TK の cDNA を得て、同様の制限酵素処理を行った pIRESpuro3 vector に組み込んだ。
2 細胞の培養方法
HEK293、A9、HeLa、HepG2、Caki-1、A549、HCT-15、MCF-7の各細胞の培養には、 TPP (Trasadingen, Switzerland) 製の組織培養用フラスコおよび24 well プレートを用い た。MDCKII 細胞の培養には TPP 製の組織培養用フラスコおよびポリスチレン製の蛍光測 定用96 well プレートを用いた。組織培養メディウムとして10% FBS (BioSolutions, Coquilam, BC, Canada)、1% penicillin/streptomycin (Sigma-Aldrich)を含む DMEM 及び RPMI1640を使用し、37ºC、95% air-5% CO2インキュベーター内で培養した。
3 トランスフェクション
3-1 一過性発現系細胞の作製
HEK293細胞は、継代に際し、ビュルケルチュルク盤を用いて細胞数を計測した後、2.0 105 cells/ml となるように調製し、poly-L-lysine (Sigma-Aldrich)コートした24 well プレー
ト (底面積1.91 cm2/well)に1 ml/well ずつ播種した。その後、約12時間培養し、細胞の
confluency が90%程度であることを確認したうえで、抗生物質の入っていない400 l の培 地に交換した。その後、Lipofectamine 2000 (Invitrogen)を用いてリポフェクション法によ り目的のプラスミドDNA(1 g/well)を導入した。4時間後、抗生物質を含む培地に交換 し、さらにその44時間後(トランスフェクション後、計48時間後)に取り込み実験に用いた。 また、遺伝子を導入しないmock 細胞として、pCI-neo vector のみを導入し、上記の手順で 作製したものを用いた。
A9細胞は、継代に際し、ビュルケルチュルク盤を用いて細胞数を計測した後、2.0 105
cells/ml となるように調製し、24 well プレート (底面積1.91 cm2/well) に1 ml/well ずつ播
種した。その直後に Lipofectamine LTX(Invitrogen)を用いて各プラスミドを導入し、 48時間の培養によって、一過性にそれぞれの遺伝子を発現させた。遺伝子(プラスミド) の導入量は1 g/well とした。2種のプラスミドを共導入する実験では、両者を1:1の比(合 計1 g)で導入した。比較のために単一のプラスミドを導入する場合には、導入しないプラ スミドをpCI-neo vector で置き換えた。
40
HeLa、HepG2、Caki-1、A549、MCF-7細胞は、継代に際し、ビュルケルチュルク盤を 用いて細胞数を計測した後、1.5 105 cells/ml となるように調製し、24 well プレート (底
面積1.91 cm2/well) に1 ml/well ずつ播種した。その後、約12時間培養し、細胞の confluency
が90%程度であることを確認したうえで、Lipofectamine 3000(Invitrogen)を用いて各プ ラスミドを導入し、48時間の培養によって、一過性にそれぞれの遺伝子を発現させた。
HCT-15細胞は、継代に際し、ビュルケルチュルク盤を用いて細胞数を計測した後、1.5 105 cells/ml となるように調製し、24 well プレート (底面積1.91 cm2/well) に1 ml/well ず
つ播種した。その後、約36時間培養し、細胞の confluency が90%程度であることを確認し たうえで、Lipofectamine 3000を用いて各プラスミドを導入し、48時間の培養によって、 一過性にそれぞれの遺伝子を発現させた。
3-2 MDCKII 安定発現系細胞の作製
MDCKII 細胞は継代に際し、ビュルケルチュルク盤を用いて細胞数を計測した後、4.0 105 cells/ml となるように調製し、24 well プレートに500 l/well ずつ播種した。その直後
に、Lipofectamine 2000を用いてリポフェクション法により目的のプラスミド DNA を導入 した。6時間後、抗生物質を含む培地に交換し、さらにその24時間後1 well 中の全細胞を剥 がし、300倍に希釈、96 well プレート (底面積0.31 cm2/well)に100 l/well ずつ細胞を播い
た。pCI-neo 及び pEGFP-C1 vector に組み込まれたプラスミドを導入した細胞では、その 24時間後、0.8 mg/ml の G418 (Sigma-Aldrich) を含んだ培地に交換し、pIRESpuro3 vector に組み込まれたプラスミドを導入した細胞では、puromycin(Nakarai, Kyoto, Japan)を 含んだ培地に交換することで、遺伝子導入細胞のセレクションを10日間ほど行った。G418 及びpuromycin を含んだ培地は2、3日おきに交換した。セレクションを行った後は、G418 及びpuromycin を含まない培地で培養し、それを安定発現株として用いた。また、遺伝子 を導入しないmock 細胞として、pCI-neo vector、pEGFP-C1 vector、pIRESpuro3 vector のみを導入し、上記の手順で作製したものを用いた。
3-3 siRNA による遺伝子発現抑制実験
HeLa 細胞は、継代に際し、2.5 104 cells/ml となるように調製し、24 well プレートに
1 ml/well ずつ播種した。その後、約6時間培養し、細胞の confluency が30%程度であるこ とを確認したうえで、hENBT1、hAPRT に特異的な Stealth TM Select RNAi (Invitrogen) 、
4 pmol/well を Lipofectamine RNAiMAX (Invitrogen)を用いて細胞に導入した。用いた siRNA の配列は、以下のとおりである。
41
hENBT1 antisense: 5’-UCAUGCUUGCCAUUCUUCUGACAUU-3’
hAPRT sense: 5’-AGAUGUCCCUGAAUACCACGCCUGG-3’ hAPRT antisense: 5’-CCAGGCGUGGUAUUCAGGGACAUCU-3’
導入を行う際の無血清培地としてはOpti-MEM I (Invitrogen)を用い、対照群(control)と して hENBT1、hAPRT とは無関係な siRNA である Stealth TM RNAi NEGATIVE
CONTROL Low GC Duplex (Invitrogen)を同様に細胞に導入した (4 pmol/well)。72時間 の培養を行い、遺伝子発現抑制系の対照群(control)とした。24 well プレートは、あらか じめpoly-L-lysine によりコーティングしたものを用いた。
また、HSV-TK/FLAG と hENBT1の siRNA の共導入に関しては、HeLa 細胞は、継代に 際し、1.5 105 cells/ml となるように調製し、24 well プレートに1 ml/well ずつ播種した。
その後、約12時間培養し、細胞の confluency が90%程度であることを確認したうえで、 HSV-TK/pCI-neo-N-FLAG (1 μg/well)及び hENBT1に特異的な Stealth TM Select RNAi (4
pmol/well)を Lipofectamine 3000を用いて細胞に導入した。
導入を行う際の無血清培地としては Opti-MEM I を用い、対照群(control)として、 pCI-neo-N-FLAG 及び hENBT1とは無関係な siRNA である Stealth TM RNAi NEGATIVE
CONTROL Low GC Duplex を同様に細胞に導入した。48時間の培養を行い、共導入系と した。
4 取り込み実験
今回の実験で用いたbuffer の組成は以下のとおりである。
緩衝剤は、HEPES を用いた。pH 5.0 – 6.0の範囲では、HEPES と等 mol の MES を用 いた。
・Hanks’ Na+ buffer: 136.7 mM NaCl, 5.36 mM KCl, 0.812 mM MgSO4, 0.411 mM KH2PO4,
0.385 mM Na2HPO4, 0.952 mM CaCl2, 25 mM D-glucose, 10 mM HEPES
・Hanks’ K+ buffer: 142.06 mM KCl, 0.812 mM MgSO4, 0.441mM KH2PO4, 0.379 mM K2HPO4,
0.952 mM CaCl2, 25 mM D-glucose, 10 mM HEPES, pH 7.4
・Hanks’ Na-gluconate buffer: 136.7 mM Na-gluconate, 0.812 mM MgSO4, 0.441 mM KH2PO4,
0.385 mM Na2HPO4, 0.952 mM Ca-gluconate, 25 mM D-glucose, 10 mM HEPES, pH 7.4
・Hanks’ K-gluconate buffer: 142.06 mM K-gluconate, 0.812 mM MgSO4, 0.379 mM K2HPO4,
0.379 mM K2HPO4, 0.952 mM Ca-gluconate, 25 mM D-glucose, 10 mM HEPES, pH 7.4
・Hanks’ Mannitol buffer: 273 mM mannitol, 0.2 mM KCl, 1.2 mM MgSO4, 0.441 mM KH2PO4,


![Fig. 5. Effect of silencing of ENBT1 and APRT on [ 3 H]adenine uptake in HeLa cells.](https://thumb-ap.123doks.com/thumbv2/123deta/9926276.1385724/18.892.133.738.167.440/effect-silencing-enbt-aprt-adenine-uptake-hela-cells.webp)
![Fig. 8. GCV uptake in HEK293 cells transiently expressing ENBT1, ENT1 or ENT2 The uptake of [ 3 H]ganciclovir (60 nM) was evaluated for 5 min at 37°C and pH 7.4](https://thumb-ap.123doks.com/thumbv2/123deta/9926276.1385724/22.892.262.629.795.1067/uptake-cells-transiently-expressing-enbt-uptake-ganciclovir-evaluated.webp)