乳がん細胞株
MCF-7 および MDA-MB-231における転写因子
E2F5 の機能解析日本大学大学院医学研究科博士課程 内科系内分泌代謝内科学専攻
稲垣 喜則
修了年
2018年
指導教員 相馬 正義
【目次】
概要 ・・・・・・・・・・・・・・・・・・・
1緒言 ・・・・・・・・・・・・・・・・・・・
6対象と方法 ・・・・・・・・・・・・・・・・
10結果 ・・・・・・・・・・・・・・・・・・・
15考察 ・・・・・・・・・・・・・・・・・・・
23まとめ ・・・・・・・・・・・・・・・・・・
27謝辞 ・・・・・・・・・・・・・・・・・・・
28表 ・・・・・・・・・・・・・・・・・・・・
29図 ・・・・・・・・・・・・・・・・・・・・
31図説 ・・・・・・・・・・・・・・・・・・・
48引用文献 ・・・・・・・・・・・・・・・・・
53研究業績 ・・・・・・・・・・・・・・・・・
581
【概要】
はじめに
本邦における乳がん患者の粗死亡率, 年齢調整死亡率はともに, ここ数十年増 加傾向が続いている
. 2013年の女性の乳がん患者の死亡者数は
13,000人程度 であり, 年齢調整死亡率は大腸がんに続いて高く, 第 2 位である. 食事やライ フスタイルの欧米化が進む発展途上国でも今後罹患者数の増大が見込まれ
,全 世界的に乳がんの有効な予防, 診断, 治療法の開発が喫緊の課題である.
乳がんの罹患のリスクのうち生活習慣要因としては
,妊娠・出産経験の有無
,初経年齢, アルコール, 肥満などが知られている. また遺伝的要因としてはがん 抑制遺伝子の変異があり
,中でも代表的な
BRCA1や
BRCA2の変異は家族性 の乳がん患者の 2 割程度で認められる. しかしながら, これらの因子では説明 ができない症例も多々あり
,早期の診断
,新規の治療法を開発する上で
,引き続 き新たな乳がん関連因子の探索を行っていくことが必要である.
乳がん にお いて
estrogen receptor (ER), progesterone receptor (PR), epidermal growth factor receptor 2 (HER2)は, 増殖や浸潤において重要な役割を持ち, 治療 標的ともなっている
.そのため
,これらの発現のパターンを基に乳がんのタイ プを分類し (Liminal A タイプ, Luminal B タイプ, HRR2 陽性タイプ
, TripleNegative (TN)
タイプ
,その他
),治療方針が決定されている
.しかしながらこれら
の受容体をいずれも発現していない TN 乳がんでは, 薬剤が作用せず予後も悪 いため新規治療薬の開発が待たれている
.近年
TN乳がんにおいて転写因子
E2F5
が過剰発現し, E2F5 が高発現であるほど生命予後が悪いことが報告され
た
.また食道がん
,肝細胞がん
,卵巣がん
,前立腺がん
,大腸がんなど各種がん
2
組織においても
E2F5が高発現しているという報告が続いている
.このため
E2F5
はがん遺伝子として機能している可能性があると考えられた. そこで本研
究では
,異なる
2株の乳がん細胞株を用いることで乳がん細胞株における転写 因子 E2F5 の機能解析を検討することを目的として以下の研究を行った.
目的
乳がん細胞の細胞機能における
E2F5の役割を細胞生物学的に検討し
,更に
E2F5
がそれらの機能を発揮する上で重要なシグナル伝達経路について検討を
行う
.対象と方法
当研究グループで保管している 7 株のヒト乳がん細胞株について, Real-time
PCRに よ り
E2F5の 発 現 量 を 確 認 し
,一 定 の 発 現 量 を 示 す
p53野 生 型
, Luminal Aタイプの MCF-7 と p53 変異型, TN タイプの MDA-MB-231 を使 用した
.細胞内における
E2F5の局在は蛍光免疫染色により行った
. E2F5の発 現抑制は lipofection 法による
siRNA導入により行い
, Negative controlには
Control siRNAを用いた
.細胞増殖能の検討は
WST8アッセイにより
,コロニ ー形成能はコロニー形成試験にて行った. 細胞周期の解析および死細胞の割合 は
, Propidium Iodidese染色した細胞を
FACSにて解析し
,核内
DNA量の分布 を調べることにより解析した. E2F5, p53 およびその下流遺伝子の mRNA の発 現は
Real-time PCRにより
,蛋白質の発現解析は
Western blottingにより行った
.E2F5
と p53 の蛋白レベルでの結合状態は免疫沈降試験にて解析し, p21
WAF1プ
ロモーターへの
E2F5および
p53の結合はクロマチン免疫沈降試験
(ChIPア
ッセイ) により行った. 統計学的解析は, Student’s t 検定によっておこなった.
3
結果
はじめに MCF-7 について解析を行った. E2F5 は核内だけでなく細胞質にも 存在していることを蛍光免疫染色により確認した
. E2F5の発現を抑制した細胞 では, WST8 アッセイにより有意な細胞生存率の低下を認め (p<0.01), コロニー 形成能も低下していた
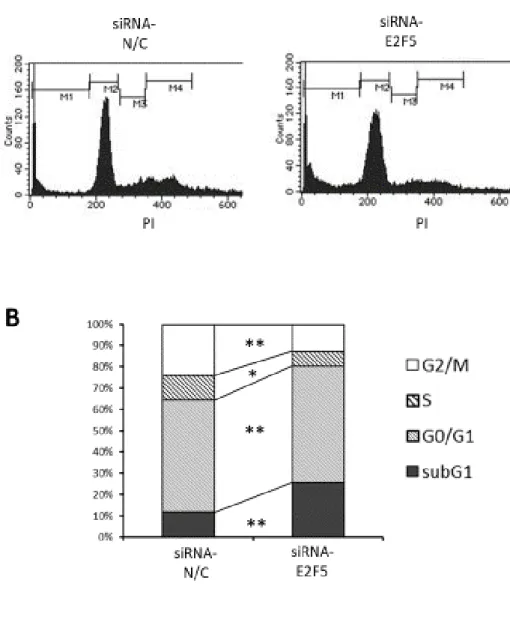
. FACS解析では
, E2F5 siRNA投与群で死細胞
(sub-G1期) の割合 (p<0.01) および G0/G1 期の細胞の割合 (P<0.05) が有意に上昇し ていることを確認した
.そこで, E2F5 発現抑制下での p53 およびその下流遺伝子 mRNA の発現レ ベ ル を
Real-time PCRに よ り 解 析 し た と こ ろ
,細 胞 周 期 調 節 因 子 で あ る
p21WAF1 (p<0.01)およびアポトーシス関連遺伝子 である
NOXA (p<0.01)と
PUMA (p<0.01)
において有意な発現上昇を認めた
. p53とそのファミリーメン
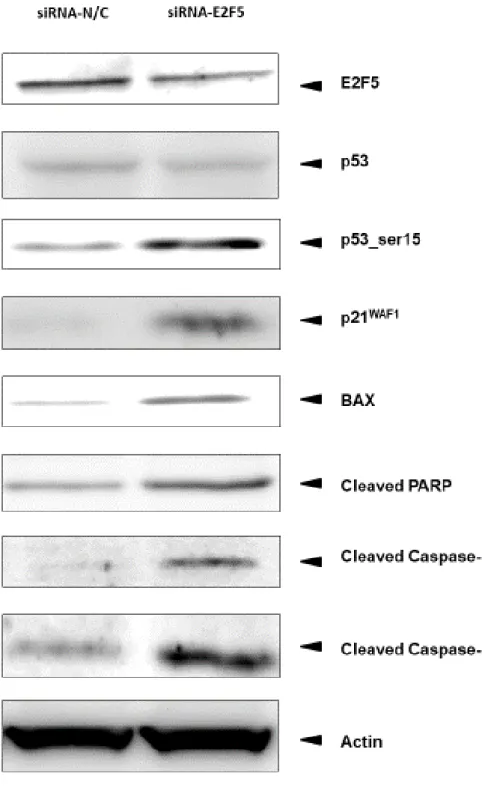
バーである p73 の発現には有意な差は認めなかった. また Western blotting を 用いた蛋白発現解析では
, p21WAF1およびアポトーシス関連蛋白である
BAX, Cleaved PARP, Cleaved Caspase-3, Cleaved Capase-9では明確な発現の上昇を認め
た
.また
, p53それ自体の発現の亢進は認めなかったが
, p53のリン酸化の亢進
を確認した.
以上の結果は
, E2F5の発現抑制により
p53シグナルが活性化され
,増殖停止 や細胞死の誘導死をもたらしたと考えられた. このため E2F5 の作用点を解明 するために
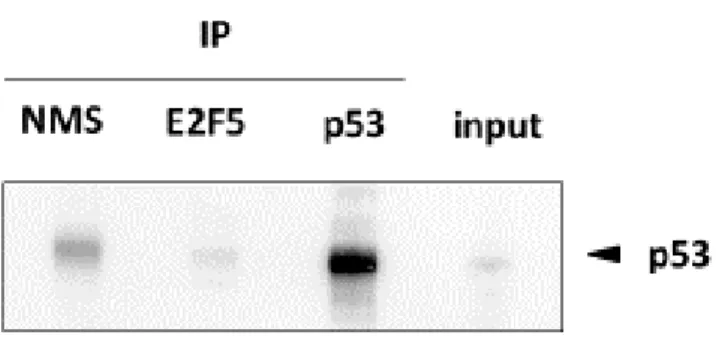
, E2F5と
p53の相互作用について免疫沈降試験で解析を行ったが
,E2F5
と p53 が共沈しているという結果は認めなかった. さらに, E2F5 が直接
p53
の標的遺伝子の発現を抑制している可能性を調べるために
, ChIPアッセイ を行ったが, E2F5 の p21
WAF1プロモーター領域への結合は確認できなかった.
また
, p21WAF1プロモーターへの
p53蛋白の結合が
, E2F5の発現抑制により亢
進するか解析をしたが, 予想とは逆に E2F5 の発現抑制により, p21
WAF1プロモ
4
ーターへの
p53の結合は抑制されていた
.次に MDA-MB231 について解析を行った. E2F5 の発現抑制により細胞の生 存率が有意に低下し
(p<0.01),コロニー形成能も抑制されることを確認した
. FACS解析では, E2F5 siRNA 投与群で死細胞 (sub-G1 期) および G0/G1 期の 細胞の割合は上昇していなかった
.一方で
, G2/M期の細胞の割合が有意に上昇 していた (p<0.05).
また
, E2F5発現抑制下での
Real-time PCRによる解析では
, p53およびその下 流遺伝子 mRNA の発現レベルは, p73 (p<0.05), p21
WAF1 (p<0.05)および NOXA
(p<0.05)と
PUMA (p<0.05) , BAX (p<0.05)において有意な発現上昇を認めたが
,MCF-7
と比較して顕著ではなかった. また p53 およびその下流遺伝子の蛋白
レベルでの発現解析では
E2F5抑制群とコントロールの間で明確な差は認めな かった. また, p53 それ自体の発現量やリン酸化レベルにも変化は認めなかった.
考察
本研究では
, 2種類の異なった乳がん細胞株である
, Luminal Aタイプのヒト乳 が ん 細 胞 株 で あ る
MCF-7と
TNタ イ プ で あ る ヒ ト 乳 が ん 細 胞 株
MDA-MB231
を用いて
, E2F5の機能解析を行った
. 2種類のタイプでともに
, E2F5の
発現抑制により, 細胞生存率, コロニー形成能が低下したが, その機序は異なっ ていると考えられた
. MCF-7では
, E2F5の発現抑制により
, G0/G1アレストと 細胞死の誘導が生じていたが, MDA-MB231 ではそのどちらも起こっておらず,
G2/Mアレストのみを認めた
.この結果は
, E2F5が
p53野生型と
p53変異型で は異なる経路で細胞増殖・細胞死誘導を制御する可能性を示唆していると考え られた
.p53
野生型の MCF-7 では, E2F5 の発現抑制により, p53 がリン酸化され, そ
5
の下流である遺伝子が発現することに伴うメカニズムにより
,細胞周期の停止 ならびにアポトーシスが亢進すると考えられた. その機序の解明を試みたが,
E2F5
と
p53の蛋白レベルでの相互作用は確認できなかった
.また直接
p21遺
伝子プロモーター領域に結合している証左も得られなかったことから, E2F5 は
p53のさらなる上流で
p53の活性化を制御する分子の機能に影響を与えてい る可能性が高いと考えられた.
一方で
, p53変異型の
MDA-MB231では
, E2F5の発現抑制による
p53下流遺 伝子の活性化や細胞死誘導, 蛋白発現の上昇は観察されなかった. また, G2/M 期における細胞周期の停止が観察されたことからは
, E2F5が複数のターゲット を持ち, p53 経路が十分に働かない細胞では, p53 非依存的な経路での細胞増殖 能の制御を行っていると考えられた
.今回の結果より
,ヒト乳がん細胞株にお ける E2F5 の機能解析には, p53 野生型と p53 変異型を区別して検討する必要 があると考えられた
.結語
今回の実験結果からは, ヒト乳がん細胞株における転写因子 E2F5 による腫瘍
制御機構は
p53野生型と
p53変異型で大きく異なることが明らかになった
.このため今後は, がん遺伝子の候補の一つであると考えられる E2F5 の解析を
おこなう上で
p53野生型と
p53変異型を分けて機能解析する必要があると考
えられた. また今回の結果からは, E2F5 はがん遺伝子の一つであることを示唆
し
,さらに他の乳がん細胞株や各種がん組織の解析を進めることで
, E2F5の詳
細な機能解析につながっていくと考える.
6
【緒言】
乳がん
乳房は乳腺と,それらを支持する脂肪組織などから構成されている. 乳腺は乳 管と多数の小葉からなり
,小葉で作られた乳汁は乳管を通過して
,乳管洞に貯 められている. 乳がんの多くは乳管から発生し, 乳管がんと呼ばれ, 小葉から発 生するがんは小葉がんと呼ばれている
.このほかのタイプも存在するが
,ほと んどの乳がんはこの 2 種類のタイプに分類される.
本邦における乳がん患者の粗死亡率
,年齢調整死亡率ともに
, 1960年代以降一 貫して増加傾向にある. 2013 年の女性の乳がん死亡者数は, 13,148 人であり, 粗 死亡率は
,大腸
,肺
,胃
,膵臓に次いで高く
,人口
10万人対
20.4人である
(2013
年). 年齢調整死亡率は, 大腸がんに次いで高く, 次に胃, 肺が続き, 人口
10
万人対
12.0人である
(乳がん
,大腸がん
,肺がんはほぼ同率
).年齢別死亡率 は 50 歳代まで直線的に増加し, その後は 70 歳代までほぼ一定である (1). 欧 米諸国の乳がん調整死亡率は
,本邦と比較してかなり高い
.しかしながら
,欧米 諸国では, 1990 年あたりをピークに減少傾向に転じており, 上昇している本邦 との差は縮まる傾向にある
(2, 3).このため本邦だけではなく
,食事やライフス タイルの欧米化が進む発展途上国なども含めて, 今後はこれまで以上に乳がん 患者は世界全体では増加すると考えられ
,新規診断方法
,新規治療薬などの開 発が望まれる (4).
乳がんの罹患のリスク要因として最も良く知られているものとして
,妊娠・出
産経験の有無, 初経年齢, 閉経年齢がある. これは, 女性ホルモンへの暴露期間
の長さが乳がん発生率に影響するためであり
,暴露期間が長いほど罹患リスク
は高くなる (5). 他にも飲酒, 喫煙などが知られているが, 遺伝的要因としては
7
いくつかのがん抑制遺伝子の変異が乳がん発症のリスクを上昇させることが判 っており, 代表的なところでは BRCA1 や BRCA2 の変異が家族性乳がん患者 の
20~25%で観察される
(6).また
p53の変異を持つ
Li-Fraumendi症候群に おいては閉経前乳がんの発症頻度が高く, 平均発症年齢が 33 歳との報告があ
る
(7).これらの遺伝子には
,生殖細胞系列変異だけでなく
,乳がん組織におけ
る体細胞変異も観察されることから, リスク予測マーカーとしても, 治療標的 としても非常に重要な変異であると言える
.しかしながらこれらの遺伝子に変 異が見られないケースも少なくなく, 未だに発見されていない乳がん関連遺伝 子があると考えられ
,解析が続けられている
.乳がんの病期 (ステージ) は腫瘍の乳房内での広がり, リンパ節転移の有無,
がん細胞の遠隔転移によって規定される
. TNM分類を用い区分を規定し
,推奨
される外科手術, 根治的放射線療法, 化学療法 (内分泌薬・抗がん剤・分子標的
薬
)を含めた治療法の中から推奨される治療法が選択される
.さらに
,乳がんで
は化学療法施行時において, 生物学的要因である臨床病理学的特徴が非常に重
要である
.代表的なものとして
, estrogen receptor (ER)や
progesterone receptor (PR)などのホルモン受容体及び epidermal growth factor receptor 2 (HER2) があ
る
.これらは乳がん細胞の増殖や浸潤において非常に重要な役割を持つことか
ら, これらの受容体などを基にして乳がんのタイプを分類し, 治療方針を検討
することが行われている
. ER(+)/HER2(-)/PR(+/-)で
Ki67(-)の
Luminal Aタイプ
, ER(+)/HER2(-)/PR(+/-)で Ki67(+) の Luminal B タイプ, ER(-)/HER2(+)/PR(-) の
HER2タイプ
,そして
ER(-)/HER2(-)/PR(-)の
Triple Negative (TN)タイプ
,そ
の他のタイプである (8) (表 1). 一般的に乳がん治療においては, これらの受容
体を標的とした薬剤やホルモン療法が用いられているが
, ER, PRそして
HER2をいずれも発現していない TN 乳がんにはこれらの薬剤が作用せず, 新規の治
8
療薬の開発が切望されている
.近年, 細胞周期関連転写因子である E2F5 が食道がん (9), 肝細胞がん (10),
卵巣がん
(11),前立腺がん
(12),大腸がん
(13)などの各種がん組織において
発現上昇を示すことが明らかになっている. TN 乳がんにおいても E2F5 が過剰 発現していること
,リンパ節転移の無いケースでは
, E2F5が高発現であるほど 無病生存期間が短いことが報告された (14).
転写因子 E2F5
E2F5
は
E2Fファミリーに属する転写因子であり
,ヒトでは
8番染色体の
q21
に存在する
E2F5遺伝子にコードされている. E2F 転写因子ファミリーは
E2F1~
8のメンバーで構成されるが
,いずれも増殖因子シグナルカスケードの 下流に位置する転写因子であり, 主に細胞周期やアポトーシスに関連した細胞 機能の制御に関わっている
.ファミリーメンバーのうち
E2F1~
5は
Rbファミ リー蛋白と複合体を形成し, 細胞周期を制御する. E2F1~3 は
Rb蛋白と, E2F4 は
Rbもしくは関連の
pocket protein p107や
p130と結合し
, E2F5は主に
p130
のみと結合することが判っている. E2F1 ~3 は転写活性化因子, 一方で
E2F4
~
8が転写抑制因子に分類されている(
15)
.前者の
E2F1~
3は休止期に ある細胞を S 期に移行させる作用を持つ (16, 17, 18). また細胞周期の進行や
複製
, DNA合成に関与する遺伝子の転写を活性化する機能を持つ (
19, 20)
.一
方で E2F-4 および 5 は主に細胞の G0/G1 の標的遺伝子プロモーターに結合
しているが
,その標的については不明な点が多い
. E2F5のノックアウトマウス
は脳脊髄液の過剰合成による水頭症を発症するとの報告があるが, 発がん感受
性への影響については特に知られていない
.一方
,通常
p16を過剰発現させる
と細胞周期の停止が生じるが, E2F4 と E2F5 を両方ノックアウトしたマウス胎
9
児由来の線維芽細胞に
p16を過剰発現させても
G1アレストが生じないこと がわかった. またこのマウスは胎生致死であった (21)
.これらの報告より, 当 初は
E2F1~
3はがん遺伝子として
, E2F4~
8はがん抑制遺伝子として機能する ことが予測されていたが, ヒト腫瘍組織の解析からは, 必ずしもそれを支持す る結果は得られていない
.先にも記した通り
, E2F5は
TN乳がんやその他のが ん腫においてはむしろ高発現していることから, がん遺伝子としての機能を持 つ可能性が十分あると考えられた
.しかしながら
,その作用機序についてはほ とんど報告がなく現時点では不明な点が多い.
そこで
,本研究では
,異なる
2種類の乳がん細胞の細胞株を用いて
E2F5の
機能解析を行い, 乳がん細胞における E2F5 の役割の解明を試みた.
10
【対象と方法】
1.
ヒト乳がん細胞株
ヒト乳がん細胞株
MCF-7, MDA-MB-231, Hs578T, BT474, SK-BR-3, T47D, MDA-MB-175Ⅶ
は日本大学医学部・増田しのぶ教授より譲渡を受けた
. MCF-7は
10%
ウシ胎児血清
(FBS) (Nichirei Bioscience, Tokyo, Japan), 100IU/ml Penicilin- Streptomycin-Glutamine (PSG) (Thermo Fisher Scientific, Walthan, MA, USA)を含有 する
RPMI培地
(Nacalai Tesque, Kyoto, Japan)を使用し
, MDA-MB-231および
MDA-MB-175Ⅶは 10% FBS および 100IU/ml PSG を含有する Leivovitz L-15 培地
(Thermo Fisher Scientific)を使用して培養した
. Hs578Tの培養には
10%FBS
と 100IU/ml PSG, 0.1U/ml bovine insulin (SIGMA, St. Louis, MO, USA) を含 有する
DMEM培地
, BT474は
10% FBSと
100IU/ml PSGを含有する
Hybri- care medium (ATCC46-X) (ATCC, Manassas, VA, USA), SK-BR-3は 10% FBS と
100IU/ml PSGを含有する
McCoy5A培地
(Thermo Fisher Scientific), T47Dは
10% FBSと 100IU/ml PSG, 0.2U/ml bovine insulin を含有する RPMI 1640 培地 を用いた
.培養は全て
37℃
, 5% CO2の条件下で行った
.2. siRNA
の 導入
E2F5
の発現抑制は
, siRNAを用いて行った
.各 siRNA は
Ambion (Thermo Fisher Scientific)より購入した
. E2F5 siRNAを最終濃度各
30nMとなるように
調整し, lipofection 法で導入した. 導入 は
Lipofectamine 3000 (Thermo Fisher Scientific)を用いて推奨プロトコールに従い
, reverse transfection法で行った
.陰
性コントロールとして, Ambion Control siRNA (Thermo Fisher Scientific) を同様に
導入して作成した
.11 3.
増殖能の検討
: WST-8アッセイ
細胞を 10,000 cells/well の密度で 96well plate に播種し, E2F5 siRNA および
Control siRNAを
transfectした
.その後培養を行い
,一定時間後に
Cell Count Reagent SF (Nacalai tesque)を含む培地に置換した
. 1時間後の呈色反応後,
Wallac 1420 ARVO MX (Perkin Elmer, Waltham, MA, USA)を用いて吸光度を測定 した. 各条件につき 4well 以上測定を行った.
4.
コロニー形成能の検討
細胞を
5,000 cells/wellの密度で
6well plateに播種し
, E2F5 siRNAおよび
Control siRNAを transfect した. その後培養を行い, 10 日後に培地を除去して
PBSで細胞表面を洗い
10%ホルマリンで固定した
.その後
, Diff Quick固定液 にて染色しコロニーを観察した.
5. RNA
の抽出と cDNA の合成
細胞からの
RNA抽出は
, RNeasy Mini kit (Qiagen, Hilden, Germany)を用いて
,推奨プロトコールに従い行った. 1 反応あたり
500ngのトータル RNA より,
Prime Script RT Master Mix (Takara Bio Inc, Shigma, Japan)を用いて逆転写反応を 行い, complementary DNA (cDNA) を作成した.
6. mRNA
レベルの遺伝子発現解析
合成した
cDNAを用いて
Real-time polymerase chain reaction (以下
, Real-time PCR)を行い, 各転写産物の発現量を解析した. E2F5 および
Actinは Taqman
pobe (Thermo Fisher Scientific)を用い
, Premix Ex Taq (Takara Bio Inc, Kyoto, Japan)にて標準プロトコールに従い行った. E2F5,
Actin以外については
STBER12
Premix Ex Taq
Ⅱ
(Takara Bio Inc)を用いて行った
.使用した各プライマーの詳 細を表 1 に示す.
Actinもしくは GAPDH を内因性コントロールとして発現 レベルの補正を行った
.各条件につき
3回測定を行い
,統計解析を施行した
.7.
蛋白の抽出と濃縮
細胞を
PBSにて洗浄後
, protease inhibitor (Roche Applied Science, Penzburg, Germany)を添加した
Pierce RIPA buffer (Thermo Fisher Scientific)を用いて溶解 した. その後 BioLupter (Cosmo Bio, Tokyo, Japan) にて超音波処理を行い, 4℃,
12,000rpmで
10分間遠心して, その上清を回収した
.8. Western blotting
精製した蛋白抽出液の蛋白濃度は BCA Protein Assay kit (Thermo Fisher
Scientific)
を用いて定量した
.蛋白を
30μ
g/laneの濃度で
NuPAGE 4-12% Bis- tris protein gel (Thermo Fisher Scientific)にアプライし, NuPAGE MOPS-SDS
(Thermo Fisher scientific)を
Running bufferとして
30mA定電流で電気泳動し た. ゲル内の蛋白質を Semi-dry 法もしくは Wet 法にて
Polyvinylidenedifluoride (PVDF) membrane
へ転写し
, Blocking One (Nacalai tesque)を用いてブロッキングした. 各一次抗体を Can get Signal Immunoreaction
enhancer Solition 1 (Toyobo, OSKA, Japan)
で
1:500に希釈し
, 4℃ で一晩反応さ せた. 用いた抗体は以下の通りである. マウス抗
E2F5抗体 (Abcam,
Cambridge, UK),
マウス抗
p53抗体
DO-1 (Sigma),ウサギ抗
p53-ser15 (Cell Signaling Technology, Beverly, MA, USA),ウサギ抗
p21WAF1抗体 (Santa Cruz,
Biotechnology, Santa Cruz, CA, USA),ウサギ抗
BAX抗体
(Cell SignalingTechnology),
ウサギ抗
PARP抗体 (Cell Signaling Technology), ウサギ抗
Caspase-13
3
抗体
(Cell Signaling Technology),ウサギ抗
Caspase-9抗体
(Cell Signaling Technology),マウス抗Actin 抗体 (Sigma). 一次抗体処理の後 PVDF
membraneを
0.1% Tween含有
Phosphate Buffered Saline(
PBS-T) を用いて 洗浄し, 二次抗体を反応させた. 二次抗体は, horseradish peroxidase (HRP) 標識 抗マウス
IgG抗体(
GE Healthcare Life Science, Buckinghamshire, UK) もしくは
HRP標識抗ラビット
IgG抗体を PBS-T で 1:2000 希釈し, 室温で 1 時間反応 させ
, PBS-Tを用いて洗浄した
. Chemi-Lumi One Super(
Nacalai Tesque) で化 学発光させたのち Bioimage Analyser LAS-4000 (Fujifilm, Osaka, Japan) を用 いて観察・撮影した
.9.
免疫沈降試験
細胞を φ6 cm dish に播種し, 80% コンフルエントになった時点で, PBS にて 洗浄し
, protease inhibitor含有
Pierce RIPAを
500μ
l/dish加え溶解した
.溶解液 を 500μl/tube ずつ 1.5ml エッペンドルチューブに移し, 各チューブに抗
E2F5抗体
,抗
p53抗体
,マウス血清をそれぞれ
2ug/tube添加し
, 4℃ で
12時間でゆ っくり振とうした. その後, Protein G Sepharose 4 Fast Flow (GE Healthcare) を添 加し
,更に
2時間
4℃ でゆっくり振とうした
.その後遠心により
Sepharoseビーズを回収し, PBST による洗浄後, 2xSDS sample buffer を添加し, 95℃ 3 分 間の熱処理を行った
.その後遠心を行い
,上清
20μ
lを
NuPAGE 4-12% Bis-tris protein gel各レーンにアプライし, 電気泳動を行った. その後 Western blotting
を行い
, PVDF膜を抗
p53抗体と反応させ
, p53蛋白の沈降レベルを解析した
.泳動条件や Western blotting の条件等は上記 7. と同様である. Input としては,
細胞溶解液を
2xSDS sample bufferと
1:1で混合したものを用いた
.14 10.
クロマチン免疫沈降試験
(ChIPアッセイ
)細胞を φ10 cm dish に播種し, 80% コンフルエントになった時点で終濃度 1%
のホルムアルデヒドを添加し
,クロスリンク反応を行った
.その後
EpiScope ChIPKit Anti-mouse IgG (Takara)を用い, 推奨プロトコールに従いクロマチン免 疫沈降物の抽出を行った
.抗体は抗
E2F5抗体
,抗
p53抗体
,マウス血清をそれ ぞれ用いた. 沈降物の中に含まれる p21
WAF1プロモーター領域の量を, Real-time
PCRにより定量した
.用いたプライマーを表
1に示す
.またこの際
, ChIPアッ セイを行う前のクロマチン断片含有溶液についても同様に Real-time PCR を行
い
,これを
inputとした
.各サンプルについて得られた値を
inputの値で割っ
たもの (%input) を算出し, これを各サンプル中の p21
WAF1プロモーター領域 含有用とした
. PCR反応は各条件につき
4回ずつ行い
,統計的な有意差の検定 を行った.
11.
統計処理
実験群間の統計的有意性は
, Student’s t検定で解析し
,値は平均値±標準偏差で
表記し, p<0.05 以下を統計学的有意差ありとした.
15
【結果】
1.
各種乳がん細胞株における転写因子 E2F5 と p53 蛋白量の解析
1-1.
各種乳がん細胞株における転写因子
E2F5の発現量
Triple Negative
タイプである Hs578T と MDA-MB-231, HER2 陽性タイプで ある
BT474と
SK-BR-3, Luminal Aタイプである
T47D, MCF-7と
MDA-MB-175Ⅶ
の計 7 種類の乳がん細胞株における E2F5 の発現量を Real-time PCR
により解析した
.解析の結果
,乳がんのタイプにおける
E2F5の発現パターン には明確な傾向はみられなかった (図 1-A).
1-2.
各種乳がん細胞株における p53 蛋白量の発現量
Triple Negative
タイプである
Hs578Tと
MDA-MB-231, HER2陽性タイプで ある BT474 と SK-BR-3, Luminal A タイプである T47D, MCF-7 と MDA-MB-
175Ⅶ の計
7種類の乳がん細胞株における
p53蛋白量の発現量を
Western blottingにより定量した
. p53野生型である, MCF-7 と MDA-MB-175Ⅶ では
p53蛋白量の発現は
, p53変異型である
, Hs578Tと
MDA-MB-231, BT474, SK- BR-3と T47D と比較して低かった (図 1-B).
1-3.
各種乳がん細胞株における転写因子
E2F5と p53 蛋白量の発現量の関係
Triple Negative
タイプである
Hs578Tと
MDA-MB-231, HER2陽性タイプで
ある BT474 と SK-BR-3, Luminal A タイプである T47D, MCF-7 と MDA-MB-
175Ⅶ の計
7種類の乳がん細胞株における
E2F5と
p53蛋白量との関係を解
析した. p53 野生型 (MCF-7, MDA-MB-175Ⅶ) では, E2F5 の発現量は比較的一
定であったが
, p53変異型
(Hs578T, MDA-MB-231, BT474, SK-BR-3, T47D)では
,16
E2F5
の発現量と
p53蛋白量の発現量の間に相関が見られ
, p53の発現量が高
い細胞株では E2F5 の発現量も高かった (図 1-C).
2.Luminal A
タイプ乳がん由来 MCF-7 細胞株を用いた解析
2-1.
乳がん細胞株
MCF-7における転写因子
E2F5の細胞内局在の結果
本研究では, 最初に一定の E2F5 の発現量を示す Luminal A タイプ細胞株
MCF-7
を用いて
E2F5の機能解析を行った
.蛍光免疫染色により
,細胞内にお
ける E2F5 の局在を調べた. 核の位置は DAPI 染色により確認した. 観察結果
から
, E2F5は核内だけでなく
,細胞質にも存在していることが確認された
(図
2).
2-2.乳がん細胞株 MCF-7
における
E2F5 siRNA導入効果の検討
乳がん細胞の細胞機能に対する
E2F5の役割を検討するため
, E2F5 siRNAを 用 い て E2F5 の発現 抑制を行った
. MCF-7に
E2F5 siRNAおよび Control
siRNAを投与し
, 48時間後における
E2F5 mRNAの発現量を
Real-time PCRにより定量した結果, E2F5 siRNA 投与群で有意な発現低下を認めた (p<0.01)
(図
3 A).また
,蛋白レベルにおける
E2F5の発現も
E2F5 siRNA投与群で抑制 されていることを Western blotting で確認した (図 3B).
2-3.E2F5
の発現を抑制した MCF-7 細胞の細胞増殖能の検討 (WST8 アッセイ
およびコロニー形成試験
)E2F5
の発現抑制が細胞増殖に及ぼす役割を検討するために, MCF-7 に E2F5
siRNA
および
Control siRNAを導入し
, 4日後に生存細胞率を
WST8アッセイ
にて解析した (図 4A). Control 群と比較して, E2F5 siRNA 投与群では有意な細
17
胞生存率の低下を認めた
.また低密度で播種した MCF-7 に E2F5 siRNA および Control siRNA を導入
し
, 10日後に染色してコロニー形成状態を観察した
. Control群と比較して
,E2F5 siRNA
投与群では明確なコロニー形成数の低下を認めた (図 4B).
2-4. E2F5
の発現を抑制した MCF-7 細胞の細胞周期・死細胞の割合の解析
E2F5
の発現抑制による細胞生存率の低下が
,細胞死の誘導によるものか
,増
殖抑制効果によるものかを確認するために, FACS 解析を用いた核内 DNA 量 の定量により
,細胞周期および死細胞の割合を調べた
. MCF-7に
E2F5 siRNAもしくは Control siRNA を導入し, 4 日後に細胞を回収した. Propidium Iodide を 用いた核内
DNAの染色後
, FACS解析を行った
(図
5A).その結果
, Control群 と比較して, E2F5 siRNA 投与群では死細胞 (sub-G1 期) の割合が有意な上昇を 示していた
(p<0.01).また
G0/G1期の細胞の割合も増加していた
(p<0.05) (図
5B).2-5. E2F5
の発現を抑制した MCF-7 細胞における p53 およびその下流遺伝
子
mRNAの発現解析
上記の解析の結果, E2F5 の発現抑制により細胞死の誘導と G0/G1 期における 細胞周期の停止が観察されたことから
,細胞増殖抑制や細胞死の誘導において 中心的な役割を果たす p53 およびその下流遺伝子の発現レベルについて検討 を行った
. MCF-7に
E2F5 siRNAまたは
Control siRNAを導入し
, 4日後に
Real-time PCR
を行い, p53 およびその制御下にある遺伝子の mRNA の発現に
ついて解析した
(図
6).その結果
,細胞周期調節因子である
p21 (p<0.01)およ
びアポトーシス関連遺伝子である NOXA (p<0.01) と PUMA (p<0.01) が有意な
18
発現の上昇を認めた
. p53自体と
,そのファミリーメンバーである
p73の発現 レベルには, E2F5 の発現抑制細胞とコントロール細胞の間で有意な差は認めら れなかった
.2-6. E2F5
の発現を抑制した
MCF-7細胞における
p53およびその下流遺伝
子の蛋白発現解析
これらの遺伝子について蛋白レベルでの発現変化についても検討を行った
. MCF-7に E2F5 siRNA および Control siRNA を導入し, 48 時間後に各蛋白の 発現量を
Western blottingにより解析をした
(図
7).細胞周期調節因子である
p21お よ び ア ポ ト ー シ ス 関 連 遺 伝 子 で あ る
BAX, Cleaved PARP, Cleaved Caspase-3と
Cleaved Caspase-9の有意な発現の上昇を
Western blottingで確認 した. また p53 の発現はほとんど変化を認めなかったが, p53 のリン酸化の亢 進を確認した
.2-7.
転写因子
E2F5と
p53蛋白質の結合状態の解析
(免疫沈降試験
)以上の結果は, E2F5 の発現抑制により p53 シグナルが活性化され, 増殖停止 や細胞死の誘導が生じている可能性を示唆した
.そこで
, E2F5の作用点を解明 するために, 最初に E2F5 と p53 の相互作用について免疫沈降実験により解析
を行った
. MCF-7細胞より抽出した蛋白質に抗
E2F5抗体を加えて沈降させ
,沈
降物を電気泳動し, Western blotting 後に抗
p53抗体の反応を行った. その結果,
抗
E2F5沈降物はコントロール血清による沈降物と比べても
p53の含有量が少
なく, E2F5 が p53 と共沈しているという結果は得られなかった (図 8).
19
2-8. p21WAF1
遺伝子プロモーターにおける転写因子
E2F5の結合状態の解析
(クロマチン免疫沈降試験)
次に
, E2F5が直接
p53の標的遺伝子の発現を抑制している可能性について
検討を行った。非小細胞肺がん細胞株において, E2F5 が p21
WAF1プロモーター
に結合し
, p21 WAF1の発現を抑えているとの報告があるため
(22),同様のこと
が MCF-7 細胞でも観察されるかどうか, ChIP アッセイで確認した. クロマチ ン断片に抗
E2F5抗体もしくはコントロール血清を投与し
,沈降物に含まれる
p21 WAF1
プロモーター領域の
DNA断片の量を Real-time PCR にて検討した.
p21 WAF1
遺伝子のプロモーター
2か所に対するプライマーを用いて解析を行
ったが, 抗
E2F5抗体による沈降物中の p21
WAF1のプロモーター DNA 量は、
コントロール血清と比べて増えてはいなかった
(図
9).2-9. p21 WAF1
遺伝子プロモーターにおける転写因子
E2F5の結合状態の解析
(クロマチン免疫沈降試験)
次に
, p21 WAF1プロモーターへの
p53蛋白の結合が
, E2F5の発現抑制により
亢進するかどうかを検討するために, MCF-7 に E2F5 siRNA および Control
siRNA
を導入し
, 4日後に各細胞よりクロマチン断片を回収し
,抗
p53抗体を用
いた免疫沈降を行った. Control siRNA を感染させた群では, 抗
p53抗体により
p21 WAF1
プロモーター領域の
DNA断片量がコントロール血清と比べて有意に
増加していたが, E2F5 siRNA を感染させた群では, 予想とは逆に, 抗
p53抗体
による
p21 WAF1プロモーターの沈降量がコントロール血清と同様のレベルま
で下がっていた (図 10). この結果は 2 種類のプライマー両方において確認さ
れた
.20
3
.
Triple Negativeタイプ乳がん由来
MDA-MB-231細胞株を用いた解析
3-1.乳がん細胞株 MDA-MB-231
における E2F5 siRNA 導入効果の検討
MDA-MB-231
細胞における
E2F5の発現抑制を行うために
, E2F5 siRNAおよ び Control siRNA を導入した. 培養 48 時間後における E2F5 mRNA の発現量
を
Real-time PCRにより定量した結果
, E2F5 siRNA投与群で有意な発現低下を
認めた (p<0.01) (図
11 A).また, 蛋白レベルにおける E2F5 の発現も E2F5
siRNA投与群で十分に抑制されていることを
Western blottingで確認した
(図
11 B).3-2.E2F5
の発現を抑制した MDA-MB-231 細胞の細胞増殖能の検討 (WST-8
アッセイおよびコロニー形成試験
)E2F5
の発現抑制が細胞増殖に及ぼす役割を検討するために, MDA-MB-231 に
E2F5 siRNA
および
Control siRNAを導入し
, 4日後に生存細胞率を
WST-8ア ッセイにて経時的に解析した (図 12 A). この細胞においても, E2F5 siRNA 投与 群では
Control群と比較して有意な細胞生存率の低下を認めた
(p<0.01).ま た 低 密 度 で 播 種 し た
MDA-MB-231に
E2F5 siRNAも し く は
ControlsiRNA
を導入し
, 10日後に染色してコロニー形成状態を観察した実験において
も, E2F5 siRNA 投与群は Control 群と比較してコロニー形成数の明らかな低下
を示した
(図
12 B).21
3-3. E2F5
の発現を抑制した
MDA-MB-231細胞の細胞周期・死細胞の割合の
解析
E2F5
の発現抑制により
,細胞死の誘導と
,細胞増殖抑制のどちらが生じてい
たのかを確認するために, FACS 解析による核内 DNA 量の定量を行い, 細胞周 期 お よ び 死 細 胞 の 割 合 を 調 べ た
. MDA-MB-231に
E2F5 siRNAも し く は
Control siRNAを導入し, 4 日後に細胞を回収した. Propidium Iodide を用いて核 内
DNAを染色し
, FACS解析を行ったところ
, Control群と比較して
, E2F5siRNA
投与群では死細胞 (sub-G1 期) の割合はほとんど増加せず, G0/G1 期の
細胞増加も確認できなかった
.しかしその一方で
, G2/M期の細胞の割合が有意 に増加していることが確認できた (P<0.05) (図 13 B).
3-4. E2F5
の発現を抑制した MDA-MB-231 細胞における p53 およびその下
流遺伝子
mRNAの発現解析
MDA-MB-231
細胞における E2F5 の発現を抑制した際の, p53 およびその下
流遺伝子の発現レベルについて検討を行った
. MDA-MB-231に
E2F5 siRNAま
たは
Control siRNAを導入し, 4 日後に real-time PCR を行い, p53 およびその制
御下にある遺伝子の
mRNAの発現について解析した
(図
14).その結果
,細胞
周期調節因子である p21 (p<0.05) およびアポトーシス関連遺伝子である BAX
(p<0.05), NOXA (p<0.05)と
PUMA (p<0.05),がん抑制遺伝子である
p53のファ
ミリーメンバーである p73 (p<0.05) において有意な発現の上昇を認めた. しか
しこれらの発現の上昇は
, MCF-7と比較するとそれほど顕著ではなかった
.22
3-5. E2F5
の発現を抑制した
MDA-MB-231細胞における
p53およびその下 流遺伝子の蛋白発現解析
これらの遺伝子について蛋白レベルでの発現変化についても検討を行った
, MDA-MB-231に E2F5 siRNA もしくは Control siRNA を導入し, 4 日後に各蛋
白の発現量を
Western blottingにより解析をした
(図
15).細胞周期調節因子で
ある p21 およびアポトーシス関連遺伝子である BAX, Cleaved PARP, Cleaved
Caspase-3と
Cleaved Caspase-9の発現レベルには
E2F5抑制による変化がみら
れなかった. また p53 の発現やリン酸化レベルにも
Control群との違いが見ら
れなかった
.23
【考察】
本研究では, Luminal A タイプのヒト乳がん細胞株 MCF-7 と Triple negative タイプのヒト乳がん細胞株
MDA-MB231を用い
, E2F5の機能解析を行った
.いずれの細胞においても, E2F5 の発現抑制により細胞生存率, コロニー形成能 が低下したが
,その機序は異なっていた
. MCF-7細胞では
E2F5の発現抑制に より G0/G1 アレストと細胞死の誘導が生じていたが, MDA-MB231 ではそのど ちらも起こっておらず
, G2/Mアレストのみを認めた
.この結果は
, E2F5が
p53野生型と変異型とで異なる経路で細胞増殖・細胞死誘導を制御している可能性 を示唆する
.p53
がん抑制遺伝子は, 393 アミノ酸から構成される核内たんぱく質 p53 を コードする
. p53は主に転写因子としての機能を持ち
,細胞周期の停止やアポト ーシス誘導, DNA 修復促進や血管新生抑制などの重要な細胞機能に関連した数 多くの遺伝子をその下流に持つ
(23, 24).通常状態ではユビキチン・プロテオソ ーム系による分解を受け, 低い発現レベルに抑えられているが, DNA 損傷を引 き起こす各種細胞ストレスなどによりリン酸化
,アセチル化などの翻訳語修飾 を受け, 活性化される. その結果 p53 は核内に蓄積し, 複数の下流遺伝子プロ モーター上の特異的な塩基に結合し
,転写を活性化する
.これまでに多くのヒ トのがんにおいて p53 変異が報告され, 腫瘍の種類にもよるが, ヒトがんの全 体の約半分に
p53変異があるとされる
(25, 26).乳がんにおいては約
20%の頻 度で p53 の体細胞変異が見られるとの報告や (27), p53 の生殖細胞系列変異を
持つ
Li-Fraumendi症候群においては若年性の乳がんの発症率が高いとの報告
がある (7).
まず
, p53野生型
MCF-7に関する検討結果について考察する
. MCF-7は
p5324
野生型である
. MCF-7においては
, E2F5の発現抑制により
p53のリン酸化が 亢進し, それに伴い p53 の下流遺伝子である p21 やアポトーシス関連遺伝子 の発現が上昇していた
(図
6, 7).一方
, p53変異型の
MDA-MB-231においては
, p53のリン酸化および, 下流遺伝子の発現亢進は確認できなかった (図 15). 以 上の結果からは
, E2F5が野生型
p53の活性を抑制することにより細胞増殖の 亢進, 細胞死の阻害を行う機能を持つ事を示唆する. より詳細なメカニズムを 探るために、免疫沈降実験により
E2F5と
p53の結合状態について検討したが
,この二つの蛋白の結合は証明できなかった (図 8). また, p53 が p21 のプロモ ーターに結合して
p21の転写を活性化する際に
, E2F5を抑制するとより活性 が上昇するか ChIP アッセイにより検討したが, 予想とは逆に E2F5 の発現抑
制により
, p21プロモーター領域への
p53蛋白の結合は抑制されていた
(図
10).
また, 非小細胞肺がん細胞を用いた解析では, E2F5 そのものが p21 プロ
モーターに結合し
, p21の発現を抑制している事が報告されているが
(22),MCF-7
を用いた今回の実験では p21 プロモーターへの E2F5 の結合は確認で
きなかった
(図
9).以上のように
, E2F5の発現抑制により
p53のリン酸化が 亢進するものの, E2F5 が直接
p53, p21遺伝子プロモーター領域に結合している 証左は得られなかったことから
, p53のさらに上流で
p53の活性化に関与する 何らかの分子の機能に影響を与えている可能性が高い (図 16).
次に
, p53変異型である
MDA-MB231に関する検討結果について考察する
.MDA-MB231
は p53 の codon 280 のアミノ酸がアルギニンからリジンに代わ
るミスセンス変異を持つ
.この変異は他のほとんどの腫瘍で観察される変異と
同様, p53 の DNA ドメインに位置しており, これにより p53 の立体構造が変
化し
,下流遺伝子の転写を誘導する機能が阻害される
(28).今回得られた結果
においても, E2F5 の発現抑制によって, 野生型 p53 を持つ MCF-7 に見られた
25
ような
, p53下流遺伝子の活性化や細胞死誘導は観察されなかったことは
,この
p53
の変異のためであると考えられる. 一方, MDA-MB-231 においては G2/M 期において細胞周期が停止している細胞の割合が増加していた
.このことは
E2F5
が複数の標的分子を持ち, p53 経路が不全の細胞においても, p53 非依存
的な経路での細胞増殖能の制御を行っていることを意味する
.さらに近年, p53 により E2F5 が発現制御を受けることが報告されている.
非小細胞肺がん細胞を用いた解析では
, E2F5の発現を抑制する
microRNA128-2が, 変異型 p53 により発現誘導を受けることが示された (22). また, 乳がん細 胞や膵がん細胞を用いた解析から
, E2F5の発現を抑制する
microRNA let7iが
,変異型 p53 により発現抑制を受けることが示された (29). 前者は変異型 p53 が
microRNAを介して
E2F5の発現を抑制し
,後者では
E2F5の発現を抑制 する microRNA の転写抑制を行うため, 変異型 p53 は E2F5 の発現を促進し ているという結果になった
.今回
,我々の行った解析においても変異型
p53を 持つ細胞株では p53 の発現レベルと E2F5 の発現レベルが相関しており (図
1)
,乳がん細胞では
Subramanianらの観察結果
(29)と同様であった
. E2F5と
p53の関係について理解するにはさらにデータの蓄積が必要であるが, 今回得 られた結果とこれらの報告を考え合わせると
,この
2種類の遺伝子は相互に制 御し合う密接な関係がある可能性が高い(図
16).乳がんでは
,悪性度が高い
TN乳がんで
E2F5の高発現が報告された
(14).また, 乳がんのみならず, 食道がん (9) においても悪性度の高さと
E2F5の発 現レベルに相関のあること
,肝細胞がん
(10)や卵巣がん
(11)においても正常 組織と比べてがん組織では, E2F5 の発現が亢進していることが報告され, 前立 腺がんにおいては
, E2F5の位置する染色体領域
8q21-24が増幅し
,それに伴い
E2F5
の発現が亢進していることが報告されている (12). 今回の検討結果から,
26
E2F5
の発現量は
, TN乳がん培養細胞株で特異的に高いという結果は得られな
かったが, p53 変異型の細胞株においては E2F5 の発現量と p53 蛋白量の発現 量に相関があることを示す結果を得た
. p53変異型細胞株においては
, E2F5発 現量に関する何らかの調節が関与している可能性がある. TN 乳がんには p53 変異が高頻度に指摘されていることから
, E2F5の高発現が
TN乳がんという よりは, むしろ p53 変異の有無とより関係している可能性も示唆された. ヒト 乳がんの発生・進展における
E2F5の機能解析には
, p53野生型と
p53変異型 を区別して検討する必要が明らかとなった.
ヒト乳がん組織に関する解析では
,細胞質における
E2F5の発現亢進が観察さ れた (14). 培養細胞を用いた我々の解析においても, E2F5 は核と細胞質の両方 に存在することを確認した
.E2F1~6
は転写因子として機能するために Dimerization Protein (DP) と結合し て複合体を作る必要がある
(30, 31).特に
, E2F4と
E2F5はその配列中に核外 搬出シグナルを持ち, DP 蛋白が結合していない状態では核内に移行すること ができない
(18, 32).転写因子である
E2F5の発現が細胞質内で亢進すること が, 乳がんの悪性化にどのように関係するのか, 現時点では全く不明である.
E2F5
が転写因子として以外の機能をもつ可能性について検討することも今後
の検討課題と言える.
27
【まとめ】
ヒト乳がん細胞株である, MCF-7 と MDA-MB-231 において転写因子 E2F5 の発現抑制は細胞の生存率を低下させることが確認できた
.野生型
p53を持つ
MCF-7
細胞では E2F5 の発現抑制により G0/G1 アレストと細胞死の誘導が生
じ
,変異型
p53を持つ
MDA-MB-231では
G2/Mアレストのみを認めたこと
から, E2F5 は p53 シグナル経路および p53 非依存の経路の両方を通じて細胞
増殖
,生存を制御している可能性が示された
.今回の結果は
E2F5ががん遺伝
子の一つであることを示唆し, さらに他の乳がん細胞株やがん組織の解析を進
めることで
, E2F5の詳細な機能解析につながっていくと考える
.28