日本ならびにその近海で記録されたクドア属粘液胞子虫
(1930 〜 2016)
佐 藤 宏、笠 井 亨 浩
山口大学 共同獣医学部 寄生虫学教室1.はじめに
クドア属 (Kudoa Meglitsch, 1947;以下クドア) は刺 胞動物門ミクソゾア亜門粘液胞子虫綱多殻目(Cnidaria: Myxozoa: Myxosporea: Multivalvulida)に分類され[27, 29, 56, 71]、2016 年末現在、Table 1 に示す 108 種が記 録されている[25, 67, 72]。日本人にとっては親しみの もてる属名「Kudoa」は、東京大学で博士号を取得後に 渡米し、粘液胞子虫や微胞子虫研究に多大な功績を残し たイリノイ大学動物学教授 Richard Rokusaburo Kudo (1886−1967)に由来する[52, 53, 61]。 クドアは魚類の諸臓器(脳、筋肉、心嚢、消化管壁、 卵巣、鰓など)に寄生し、母細胞内に多数の娘細胞を 内包したプラズモディウム plasmodium の中で、娘細胞要 約
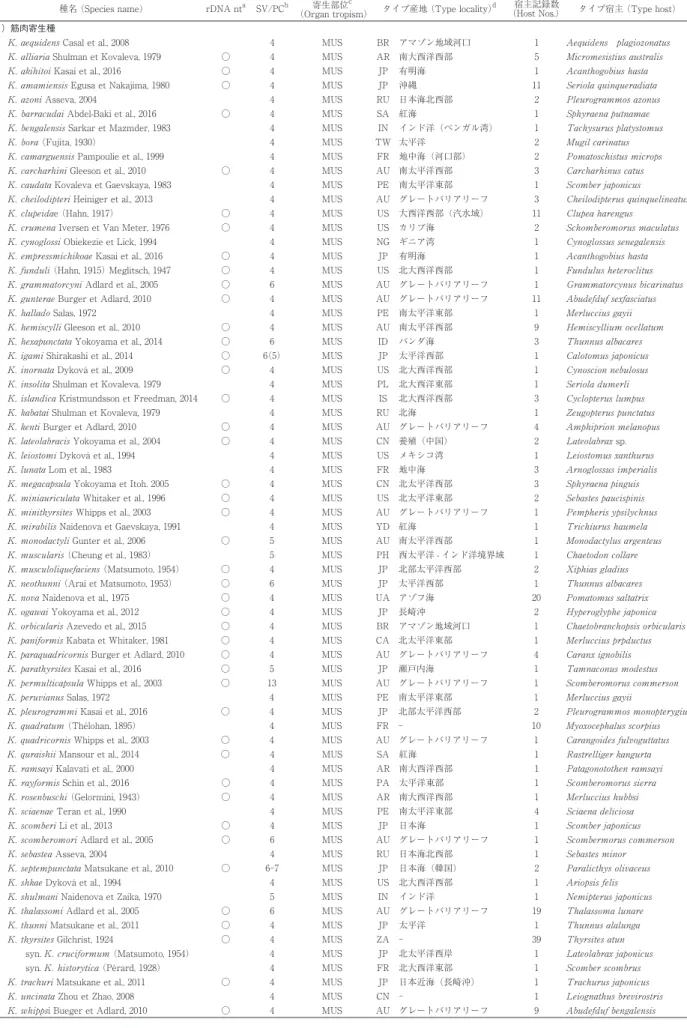
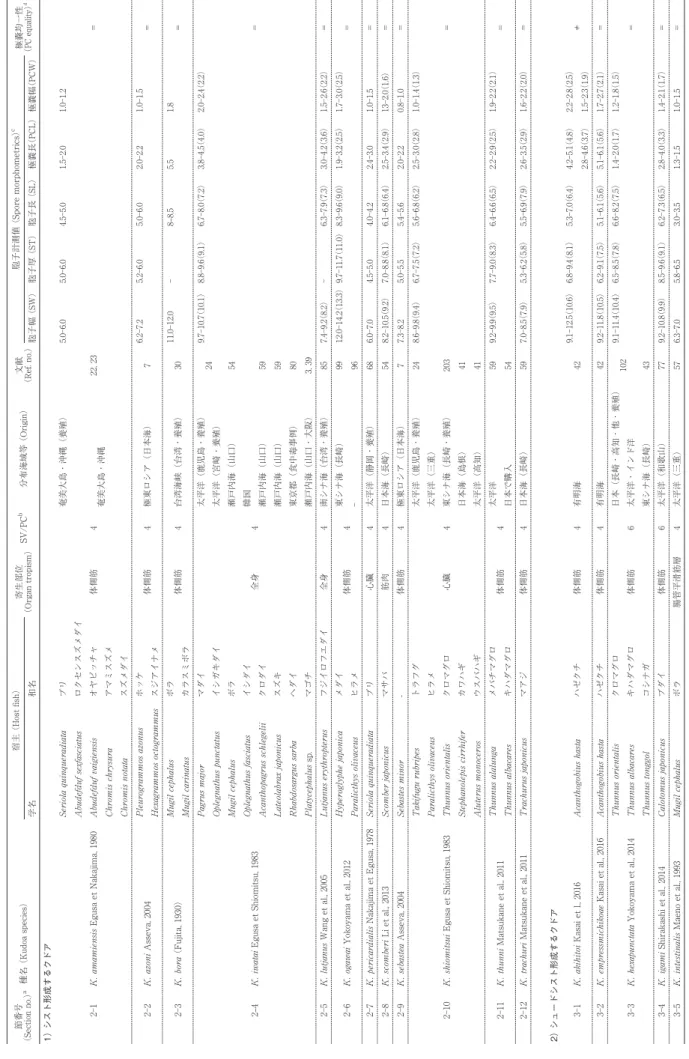
クドア属(Kudoa Meglitsch, 1947;以下、クドア)は刺胞動物門ミクソゾア亜門粘液胞子虫綱多殻目に分類され、 現在までに 108 種が記録されている。脳や筋肉、心嚢、消化管壁などに寄生し、異常遊泳や死後筋肉融解あるいは白 点多発による生産性低下や市販価値喪失などを引き起こす重要な魚類病原体として知られている。加えて 2011 年 6 月以降、新たな寄生虫性食中毒であるクドア食中毒の原因として公衆衛生上の関心も高まっている。日本近海を含め た極東地域からは 33 種が報告され、そのうち日本国内で水揚げあるいは市販されている海産魚からは 25 種のクドア が確認されている。ここでは、極東海域もしくは日本近海(日本海、東シナ海、西太平洋)で確認されている 33 種 の確認状況や胞子形態の特徴などの概要をまとめた。クドアはシスト形成型とシュードシスト形成型に大別され、前 者は肉眼的に切り身などに白点としてみられ、後者は筋細胞内などに被膜なしで局在し切り身での肉眼的な確認は難 しい。シスト形成型あるいはシュードシスト形成型のいずれであっても、その中にプラズモディウムと呼ぶ母細胞が あって、その細胞内に 5 〜 15μm 程度の胞子が多数形成される。胞子の外壁を構成する殻片と交互宿主接着のため の極糸を容れる極嚢、交互宿主へ侵入する胞子細胞体が主たる構成細胞である胞子は、小さく単純な構造であり形態 学的な特徴づけが難しい。クドアは 4 つ以上の殻片 / 極嚢をもつ胞子であるが、この明確な形態指標であっても変異 がしばしば見られる。リボソーム RNA 遺伝子を指標とした分子系統学的解析で種同定が行える基盤情報が充実して きたが、日本国内でも 2010 年に以降に 11 新種が記録されているように、種多様性理解は現在もまだ不十分である。 これまでに日本を含めた極東海域で確認されたクドアの概要と今後の課題を記し、社会的関心も増してきたクドア感 染の水産現場や公衆衛生現場での参考となること、更に、今後のクドア粘液胞子虫に関する基盤研究の進展に寄与で きることを願っている。 Key words:クドア、粘液胞子虫、種鑑別、胞子形態、極東、日本 である胞子形成細胞 sporogonic cell と周細胞 pericyte がペアとなって粘液胞子 myxospore を形成するための 細胞分裂・分化が進行する特異な寄生体である[56]。 すなわち、周細胞に包まれた中で、胞子形成細胞は殻 片形成細胞 valvogenic cell、極嚢形成細胞 capsulogenic cell、胞子原形質体形成細胞 sporoplasmogenic cell へと 細胞分裂・分化して、胞子として完成することになる。 胞子は殻片 shell valve で囲まれ、殻片の接合部を縫合 線 sutural line と呼び、その内部には極糸 polar filament を容れる極嚢 polar capsule と 2 細胞性の胞子原形質体 sporoplasm が配置されている。極嚢数は殻片数と一致 し、クドアでは 4 以上となる[89]。なお、殻片 / 極嚢 数が 3 つ以上の粘液胞子虫を多殻目として、2 つのもの を双殻目としている[56]。多殻目のうち、殻片 / 極嚢数が 3 つのものとしては Trilospora Noble, 1939 あるい は Unicapsula Davis, 1924 が知られ、今世紀初頭までの 形態分類学では、殻片 / 極嚢数が 4、5、6、7 の多殻目 粘液胞子虫については Kudoa、Pentacapsula Naidenoa et Zaika, 1970、Hexacapsula Arai et Matsumoto, 1953、 および Septemcapsula Hsieh et Chen, 1984 と区別され た[6, 34, 38, 56]。2003 年、Whipps et al. [87]によっ てオーストラリア産ヨコシマサワラ Scomberomorus commerson から 13 の殻片 / 極嚢数をもつ粘液胞子虫 が見出され、その 18S リボソーム RNA 遺伝子(18S rDNA) 塩基配列が 4 つの殻片 / 極嚢をもつクドアと分 子系統樹において同じクレードに入ることが報告され、 更に Pentacapsula 属や Hexacapsula 属についても同様 であったことから[89]、Kudoa 属の定義を修正して、 4 つ以上の殻片 / 極嚢数の粘液胞子虫をすべて「Kudoa 属」として再分類することになった。後述するように、 生鮮ヒラメ喫食に伴う食中毒の原因とされる Kudoa septempunctata では 6 〜 7 の殻片 / 極嚢数が確認されて おり[41, 58]、また Kudoa yasunagai においても殻片 / 極嚢数の変異についての報告がある[11, 20, 21]。外に も、K. chaetodoni Burger et al., 2007 や K. thalassomi Adlard et al., 2005 で同様の現象が報告されている[12, 13, 73]。このことは、粘液胞子虫の種分類において殻片 / 極嚢数は重要な指標となる一方で[56]、Kudoa 属にお いては種内変異が起こり不安定な指標でもあることを考 慮するならば、4 以上の殻片 / 極嚢数をもつ粘液胞子虫 をクドアと一括する分類体系は妥当性がある。過去の形 態分類体系にあっては実際に、江草[21]はスズキの異 常遊泳の原因となる脳寄生クドア(当時の Septemcapsula yasunagai Hsieh et Chen, 1984、現在の K. yasunagai) の種同定にあたり、7 殻片 / 極嚢あるいは 6 殻片 / 極嚢 の 2 形態型の胞子を得て、個体により両者の比率は一定 しないことから、その分類学的位置づけに当惑している ことを書き残している。クドアの種鑑別において、胞子 の形態を特徴づけるために胞子構造の計測値が用いられ るが、過去には研究者によりその計測基準が様々であり 混乱もみられた。今日、Lom & Arthur [55]が提唱し た計測部位と呼称が標準的に用いられている(Fig. 1)。 日本での特筆されるクドア感染の社会的認知は、1975 年に開催された沖縄国際海洋博覧会(以下、沖縄海洋博、 会期:1975 年 7 月 20 日−1976 年 1 月 18 日)で試みら れた海洋牧場(ブリ Seriola quinqueradiata 養殖)の失 敗に遡ることができる。養殖ブリの体側筋に白点を多発 させた原因は東京大学農学部水産学科魚病学研究室(現 東京大学大学院農学生命科学研究科水圏生物科学専攻魚 病学研究室)を中心として Kudoa amamiensis Egusa et Nakajima, 1980 と特定され[22, 23]、その後に続発する 養殖魚のクドア感染症においても世界的にも高く評価さ れる研究が重ねられている。それ以前にも、水産業が主 要産業の1つである日本の研究者たちは、粘液胞子虫に 関わる研究を病態の原因解明とこれに付随する新種記載 として行ってきていたことは後述するとおりである。 さて、魚病として市場価値の維持や養殖生産性の観点 から問題とされてきたクドアであるが、平成 23 年 6 月 17 日付食安 0617 第 3 号「生食用生鮮食品による病因物 質不明有症事例への対応について」において、生食用の ヒラメの K. septempunctata ならびに馬刺しの Sarcocystis fayeri が原因として考えられる有症事例を食中毒として 扱うよう通知が出された[50, 78]。これまで水産上での みその病害性が知られていたクドアが、ヒトへの病害性 を示し得ることが行政的に認知されたことになる。これ に伴い、クドアは公衆衛生領域において重要な病原体と して位置づけられ、その診断や防遏に関する多角的な研 究の対象ともなってきた[78]。また、K. septemunctata 等の粘液胞子虫寄生魚の喫食に起因する食中毒を「クド ア食中毒」と呼称することとなった[78]。 クドア寄生、すなわちプラズモディウムの局在には 2 型が区別される。シスト(cyst) 形成型とシュード Fig. 1. クドア胞子の計測部位と呼称 . (A) 対称性のあ る胞子;(B) 非対称性胞子 . L, 胞子長;PCL, 極嚢長;PCW, 極嚢幅;ST, 縫合線部胞子厚; T, 胞子厚;W, 胞子幅 . (山口獣医誌より転載・許諾済み [72])
Table 1. 2016 年末までに報告のあるクドア種 (A list of nominal Kudoa spp. until the end of 2016)
種名 (Species name) rDNA nta SV/PCb 寄生部位c
(Organ tropism) タイプ産地 (Type locality)d (Host Nos.)宿主記録数 タイプ宿主 (Type host) 1)筋肉寄生種
K. aequidens Casal et al., 2008 4 MUS BR アマゾン地域河口 1 Aequidens plagiozonatus K. alliaria Shulman et Kovaleva, 1979 ○ 4 MUS AR 南大西洋西部 5 Micromesistius australis K. akihitoi Kasai et al., 2016 ○ 4 MUS JP 有明海 1 Acanthogobius hasta K. amamiensis Egusa et Nakajima, 1980 ○ 4 MUS JP 沖縄 11 Seriola quinqueradiata K. azoni Asseva, 2004 4 MUS RU 日本海北西部 2 Pleurogrammos azonus K. barracudai Abdel-Baki et al., 2016 ○ 4 MUS SA 紅海 1 Sphyraena putnamae K. bengalensis Sarkar et Mazmder, 1983 4 MUS IN インド洋(ベンガル湾) 1 Tachysurus platystomus K. bora (Fujita, 1930) 4 MUS TW 太平洋 2 Mugil carinatus K. camarguensis Pampoulie et al., 1999 4 MUS FR 地中海(河口部) 2 Pomatoschistus microps K. carcharhini Gleeson et al., 2010 ○ 4 MUS AU 南太平洋西部 3 Carcharhinus catus K. caudata Kovaleva et Gaevskaya, 1983 4 MUS PE 南太平洋東部 1 Scomber japonicus K. cheilodipteri Heiniger et al., 2013 4 MUS AU グレートバリアリーフ 3 Cheilodipterus quinquelineatus K. clupeidae (Hahn, 1917) ○ 4 MUS US 大西洋西部(汽水域) 11 Clupea harengus
K. crumena Iversen et Van Meter, 1976 ○ 4 MUS US カリブ海 2 Schomberomorus maculatus K. cynoglossi Obiekezie et Lick, 1994 4 MUS NG ギニア湾 1 Cynoglossus senegalensis K. empressmichikoae Kasai et al., 2016 ○ 4 MUS JP 有明海 1 Acanthogobius hasta K. funduli (Hahn, 1915) Meglitsch, 1947 ○ 4 MUS US 北大西洋西部 1 Fundulus heteroclitus K. grammatorcyni Adlard et al., 2005 ○ 6 MUS AU グレートバリアリーフ 1 Grammatorcynus bicarinatus K. gunterae Burger et Adlard, 2010 ○ 4 MUS AU グレートバリアリーフ 11 Abudefduf sexfasciatus K. hallado Salas, 1972 4 MUS PE 南太平洋東部 1 Merluccius gayii K. hemiscylli Gleeson et al., 2010 ○ 4 MUS AU 南太平洋西部 9 Hemiscyllium ocellatum K. hexapunctata Yokoyama et al., 2014 ○ 6 MUS ID バンダ海 3 Thunnus albacares K. igami Shirakashi et al., 2014 ○ 6(5) MUS JP 太平洋西部 1 Calotomus japonicus K. inornata Dyková et al., 2009 ○ 4 MUS US 北大西洋西部 1 Cynoscion nebulosus K. insolita Shulman et Kovaleva, 1979 4 MUS PL 北大西洋東部 1 Seriola dumerli K. islandica Kristmundsson et Freedman, 2014 ○ 4 MUS IS 北大西洋西部 3 Cyclopterus lumpus K. kabatai Shulman et Kovaleva, 1979 4 MUS RU 北海 1 Zeugopterus punctatus K. kenti Burger et Adlard, 2010 ○ 4 MUS AU グレートバリアリーフ 4 Amphiprion melanopus K. lateolabracis Yokoyama et al., 2004 ○ 4 MUS CN 養殖(中国) 2 Lateolabrax sp. K. leiostomi Dyková et al., 1994 4 MUS US メキシコ湾 1 Leiostomus xanthurus K. lunata Lom et al., 1983 4 MUS FR 地中海 3 Arnoglossus imperialis K. megacapsula Yokoyama et Itoh. 2005 ○ 4 MUS CN 北太平洋西部 3 Sphyraena pinguis K. miniauriculata Whitaker et al., 1996 ○ 4 MUS US 北太平洋東部 2 Sebastes paucispinis K. minithyrsites Whipps et al., 2003 ○ 4 MUS AU グレートバリアリーフ 1 Pempheris ypsilychnus K. mirabilis Naidenova et Gaevskaya, 1991 4 MUS YD 紅海 1 Trichiurus haumela K. monodactyli Gunter et al., 2006 ○ 5 MUS AU 南太平洋西部 1 Monodactylus argenteus K. muscularis (Cheung et al., 1983) 5 MUS PH 西太平洋 - インド洋境界域 1 Chaetodon collare K. musculoliquefaciens (Matsumoto, 1954) ○ 4 MUS JP 北部太平洋西部 2 Xiphias gladius K. neothunni (Arai et Matsumoto, 1953) ○ 6 MUS JP 太平洋西部 1 Thunnus albacares K. nova Naidenova et al., 1975 ○ 4 MUS UA アゾフ海 20 Pomatomus saltatrix K. ogawai Yokoyama et al., 2012 ○ 4 MUS JP 長崎沖 2 Hyperoglyphe japonica K. orbicularis Azevedo et al., 2015 ○ 4 MUS BR アマゾン地域河口 1 Chaetobranchopsis orbicularis K. paniformis Kabata et Whitaker, 1981 ○ 4 MUS CA 北太平洋東部 1 Merluccius prpductus K. paraquadricornis Burger et Adlard, 2010 ○ 4 MUS AU グレートバリアリーフ 4 Caranx ignobilis K. parathyrsites Kasai et al., 2016 ○ 5 MUS JP 瀬戸内海 1 Tamnaconus modestus K. permulticapsula Whipps et al., 2003 ○ 13 MUS AU グレートバリアリーフ 1 Scomberomorus commerson K. peruvianus Salas, 1972 4 MUS PE 南太平洋東部 1 Merluccius gayii
K. pleurogrammi Kasai et al., 2016 ○ 4 MUS JP 北部太平洋西部 2 Pleurogrammos monopterygius K. quadratum (Thélohan, 1895) 4 MUS FR - 10 Myoxocephalus scorpius K. quadricornis Whipps et al., 2003 ○ 4 MUS AU グレートバリアリーフ 1 Carangoides fulvoguttatus K. quraishii Mansour et al., 2014 ○ 4 MUS SA 紅海 1 Rastrelliger kangurta K. ramsayi Kalavati et al., 2000 4 MUS AR 南大西洋西部 1 Patagonotothen ramsayi K. rayformis Schin et al., 2016 ○ 4 MUS PA 太平洋東部 1 Scomberomorus sierra K. rosenbuschi (Gelormini, 1943) ○ 4 MUS AR 南大西洋西部 1 Merluccius hubbsi K. sciaenae Teran et al., 1990 4 MUS PE 南太平洋東部 4 Sciaena deliciosa K. scomberi Li et al., 2013 ○ 4 MUS JP 日本海 1 Scomber japonicus K. scomberomori Adlard et al., 2005 ○ 6 MUS AU グレートバリアリーフ 1 Scombermorus commerson K. sebastea Asseva, 2004 4 MUS RU 日本海北西部 1 Sebastes minor K. septempunctata Matsukane et al., 2010 ○ 6-7 MUS JP 日本海(韓国) 2 Paralicthys olivaceus K. shkae Dyková et al., 1994 4 MUS US 北大西洋西部 1 Ariopsis felis K. shulmani Naidenova et Zaika, 1970 5 MUS IN インド洋 1 Nemipterus japonicus K. thalassomi Adlard et al., 2005 ○ 6 MUS AU グレートバリアリーフ 19 Thalassoma lunare K. thunni Matsukane et al., 2011 ○ 4 MUS JP 太平洋 1 Thunnus alalunga K. thyrsites Gilchrist, 1924 ○ 4 MUS ZA - 39 Thyrsites atun syn. K. cruciformum (Matsumoto, 1954) 4 MUS JP 北太平洋西岸 1 Lateolabrax japonicus syn. K. historytica (Pérard, 1928) 4 MUS FR 北大西洋東部 1 Scomber scombrus K. trachuri Matsukane et al., 2011 ○ 4 MUS JP 日本近海(長崎沖) 1 Trachurus japonicus K. uncinata Zhou et Zhao, 2008 4 MUS CN - 1 Leiognathus brevirostris K. whippsi Bueger et Adlard, 2010 ○ 4 MUS AU グレートバリアリーフ 9 Abudefduf bengalensis
(Table 1, continued)
種名 (Species name) rDNA nta SV/PCb 寄生部位c
(Organ tropism) タイプ産地 (Type locality)d (Host Nos.)宿主記録数 タイプ宿主 (Type host) 2)全身臓器寄生種
K. iwatai Egusa et Shiomitsu, 1983 ○ 4 SYS JP 日本近海 22 Chrysophtys major K. lutjanus Wang et al., 2005 ○ 4 SYS TW 台湾南部(養殖) 1 Lutjanus erythropterus
3)脳寄生種
K. cerebralis Paperna et Zwerner, 1974 4 CNS US 北大西洋西部 1 Morone saxatilis K. chaetodoni Burger et al., 2007 ○ 8(7-9) CNS AU グレートバリアリーフ 1 Chaetodon unimaculatus K. lemniscati Miller et Adlard, 2012 ○ 7-8 CNS AU グレートバリアリーフ 1 Lutjanus lemniscatus K. lethrini Burger et al., 2007 ○ 7 (6) CNS AU グレートバリアリーフ 2 Gymnocranius audleyi K. neurophila (Grossel et al., 2003) ○ 5 CNS AU 南太平洋西部 2 Latris lineata K. paralichthys Cho et Kim, 2003 4 CNS KR 韓国沿岸(養殖) 1 Paralicthys olivaceus K. prunusi Meng et al., 2011 ○ 5 CNS JP 太平洋西部 1 Thunnus orientalis K. tetraspora Narasimhamurti et Kalavati, 1979 4 CNS IN インド沿岸 1 Mugil cephalus K. yasunagai (Hsieh et Chen, 1984) ○ 7(5-8) CNS JP 日本近海 7 Lateolabrax japonicus
4)心嚢腔寄生種
K. aegyptica Koura, 2000 4 PEC EG - 1 Rhabdosargus haffara K. hypoepicardialis Blaylock et al., 2004 ○ 4 PEC US メキシコ湾 7 Nomeus gronovii K. leptacanthae Heiniger et Adlard, 2012 ○ 4 PEC AU グレートバリアリーフ 1 Zoramia leptacantha K. pagrusi Al Quraishy et al., 2008 ○ 4 PEC EG 紅海 1 Pagrus pagrus K. pericardialis Nakajima et Egusa, 1978 4 PEC JP 太平洋西部 1 Seriola quinqueradiata K. shiomitsui Egusa et Shiomitsu, 1983 ○ 4 PEC JP 太平洋(養殖) 5 Takifugu rubripes
5)消化管(平滑筋・結合組織等)寄生種
K. chilkaensis Tripathi, 1951 4 ESO IN チルカ湖(汽水湖) 1 Strongylura strongylura K. ciliatae Lom et al., 1992 ○ 4 GUT AU ニューサウスウェルズ沿岸 3 Sillago cliata K. cookii Heiniger et al., 2013 ○ 4 GUT AU グレートバリアリーフ 1 Ostorhinchus cookii K. dianae Dyková et al., 2002 ○ 4 ESO MX 北太平洋西部 1 Sphoeroides annulatus K. intestinalis Maeno et al., 1993 4 GUT JP 太平洋西部 1 Mugil cephalus K. sphyraeni Narasimhamurti et Kalavati, 1979 4 GUT IN インド洋(ベンガル湾) 1 Sphyraena jello K. unicapsula Yurakhno et al., 2007 ○ 4 GUT ES 地中海 2 Liza ramada K. valamugili Kalavati et Anuradha, 1993 4 GUT IN インド洋(ベンガル湾) 1 Moolgarda cunnesius 6)鰓寄生種
K. atropi Sandeep et al., 1986 4(5) GIL IN インド洋(ベンガル湾) 1 Atropus atropus K. boopsi Kpatacha et al., 1999 4 GIL SN 大西洋 1 Boops boops K. branchiata Joy, 1972 4 GIL US 大西洋 1 Leiostomus xanthurus K. eleotrisi Siau, 1971 4 GIL BJ ギニア湾 1 Eleotris kribensis
7)卵細胞 / 卵巣寄生種
K. azevedoi Mansour et al., 2013 ○ 4 OVA TN 地中海 1 Trachurus trachrus K. ovivora Swearer et Robertson, 1999 ○ 4 OVA PA カリブ海 7 Thalassoma bifasciatum K. saudiensis Mansour et al., 2015 ○ 4 OVA SA 紅海 1 Rastrelliger kangurta
8)腸管膜 / 腹膜寄生種
K. cascasia Sarkar etChaudhury, 1996 4 MES IN インド洋(ベンガル湾) 1 Sicamugil cascasia K. trifolia Holzer et al., 2006 ○ 4 PER ES 地中海 2 Liza aurata
9)腎臓・膀胱寄生種
K. stellula Yurakhno, 1991 4 KID UA 黒海 1 Atherina hepsetus K. vesica Kovaleva et Gaevskava, 1984 4 UBL AQ 南極海 1 Pseudoicichthys australis
10)皮下組織寄生種
K. cutanea Kovaleva et Gaevskava, 1984 5 SUB AQ 南極海 1 Echiodon sp.
11)胆嚢組織寄生種
K. haridasae Sarkar et Ghosh, 1991 4 GBL IN インド洋(ベンガル湾) 1 Mugil persina K. petala Zhou et Zhao, 2008 4 GBL CN - 1 Sillago shihama K. pyramidalis Zhou et Zhao, 2008 4 GBL CN - 1 Caranx djeddaba K. tachysurae Sarkar et Mazumder, 1983 4 GBL IN インド洋(ベンガル湾) 1 Plicohollis tenuispinis
a rDNA 塩基配列登録 (Deposition of rDNA nucleotide sequences at DDBJ/EMBL/GenBank). b 殻片 / 極嚢数 (No. of SV/PC). c (CNS) 脳;(ESO) 食道(平滑筋もしくは結合組織);(GBL) 胆嚢;(GIL) 鰓;(GUT) 腸管壁(平滑筋もしくは結合組織);(KID) 腎臓;(MES) 腸間膜;(MUS) 体側筋; (OVA) 卵細胞 / 卵巣;(PEC) 囲心腔;(PER) 腹膜;(SUB) 皮下組織;(SYS) 全身臓器;(UBL) 膀胱 . d (AQ) 南極;(AR) アルゼンチン;(AU) オーストラリア;(BJ) ベナン;(BR) ブラジル;(CA) カナダ;(CN) 中国;(ES) スペイン;(EG) エジプト;(FR) フランス;(ID) インド;(IN) インド;(IS) アイスランド;(JP) 日本;(KR) 韓国;(MX) メキシコ;(NG) ナイジェリア;(PA) パナマ;(PE) ペルー;(PH) フィリピン;(PL) ポーラ ンド;(RU) ロシア;(SA) サウジアラビア;(SN) セネガル;(TN) チュニジア;(TW) 台湾;(UA) ウクライナ;(US) 米国;(YD) イエメン;(ZA) 南アフリカ .
シスト (pseudocyst) 形成型である。前述のブリの K. amamiensis 寄生は前者、ヒラメの K. septempunctata 寄生は後者である。シスト形成型では、組織内に局在す るプラズモディウムは宿主由来の結合組織性被膜に包ま れており、肉眼的に白点として確認されることがしばし ばである。シュードシスト形成型では、筋細胞など大型 の細胞の細胞質内や脳脊髄液にプラズモディウムが被膜 なしで局在、もしくは浮遊し、その寄生を肉眼的に確認 することは難しいのが一般的である。 2016 年 12 月現在、クドアとして種名が確定している ものは全世界で 108 種であることは前述した (Table 1)。 そのうち、極東地域(日本近海、中国、台湾、韓国及び ロシア極東部)からは 33 種が報告されているが、その うち日本国内で水揚げあるいは市販されている海産魚か らは 25 種のクドアの寄生が報告されている(Table 2)。 ここでは、極東海域もしくは日本近海(日本海、東シナ 海、北西太平洋)で確認されている 33 種の概要につい てまとめ、社会的関心も増してきたクドア寄生の水産現 場や公衆衛生現場での参照となること、あるいは、今後 のクドア粘液胞子虫に関する基盤研究の進展に寄与でき ることを願っている。なお、顕微鏡検査の際にクドア胞 子の形態がイメージできるように、それぞれの種記載報 告を参照して描画を独自に用意したが(Fig. 2)、種の確 定作業の中では原記録をご参照いただきたい。また、以 下のクドア胞子の計測値は代表的な値を報告から選び、 「最小値−最大値(平均値)」で示すようにしたが、原報 告によっては範囲あるいは平均値の一方のみを示す場合 もある。
2.シスト形成するクドア
2-1.Kudoa amamiensis Egusa et Nakajima, 1980 奄美・沖縄海域に導入飼養されたブリの体側筋に夥し い数の白色で類円形のシストを形成したことから 1970 年代に注目され、1980 年に新種として記載された[22, 23]。和名は「アマミクドア」。本種のシストは、ブリ の横紋筋 (体側筋、頭部諸筋、鰭筋など)、心筋、鰭 膜内に発生し、芥子粒大〜米粒大 (3.2-4.6mm × 2.2-2.6mm)、最大で 5mm に達することもある[22]。本症 の国内発生はブリの非分布海域である奄美・沖縄海域に 限られ、1970 年代初頭の奄美大島養鰤場での数万尾単 位の寄生事例や 1975 年の沖縄海洋博の開催に際しての 海洋牧場での寄生事例 (導入飼養した 2.5 万尾すべてが 高度に感染) が有名で[22]、その後、奄美・沖縄海域 では高い感染感受性魚であるブリの養殖は行われてい ない。奄美大島養鰤場での発生においては、海に設置 した生け簀の位置により罹患率が異なったことが報告 されている。すなわち、海岸に近いものではほぼ 100% の感染率であったが、沖合に設置した生け簀ではほと んど感染が見られなかった。この事実は、感染源とな る放線胞子あるいは交互宿主 alternate host となる環形 動物(おそらく多毛類)への暴露機会と関連づけて考 えると興味深い。ブリでのアマミクドア症発生海域に 生息する在来宿主探索が奄美大島ならびに沖縄で行わ れ(少なくとも 17 科 28 属 45 種 1,400 尾余)、スズメダ イ類が自然宿主となっている可能性が示唆されたこと から、更にスズメダイ類(スズキ目スズメダイ亜目ス ズメダイ科)7 属 20 種 2,251 尾を肉眼で精査して、スズ メダイ属 (スズメダイ Chromis notata 115 尾、アマミ スズミ Chromis chrysura 2 尾)、オヤビッチャ属 (オヤ ビッチャ Abudefduf vagiensis 2 尾、ロクセンスズメダ イ Abudefduf sexfasciatus 4 尾)、ルリスズメダイ属 (ル リスズメ Chrysiptera cyanea 4 尾)、計 3 属 5 種 127 尾 に感染が確認された[22]。これら在来魚での寄生部位 は脊椎骨の神経棘間あるいは血管棘間、椎体周辺、背鰭・ 臀鰭・尾鰭の鰭骨間の骨格筋での寄生であり、宿主1個 体当たりのシスト数は 1 〜数個、大きさも 2mm を超す ものはなかった[22]。 1990 年に、沖縄海洋博の開催地であった沖縄本島本 部町でのカンパチ Seriola dumerili 飼育試験事業で改め て発生したことを受けて、沖縄各地の海域でのブリおよ びカンパチの飼養実験が実施された[79]。本種の汚染 海域は、ほぼ本部町沖合に限定されることが明らかにな り、交互宿主と推定される多毛類が生息できる海域環境 の重要性が指摘されているが[79]、未だ交互宿主の特 定ならびに放線胞子虫の発見には成功していない。汚染 海域に浮遊する放線胞子虫は砂濾過(径 10-30μm 以上 の粒子を除去)を通過し、短波長紫外線照射(253.7nm UV; 216 mJ/cm2)で感染性を失うことが確認されてい る[76]。 本種は奄美・沖縄の環礁に生息するスズメダイ類が自 然宿主となっていることから、豪州の大堡礁、すなわち グレート・バリア・リーフ (Lizard 島および Heron 島 周辺)に生息する類縁魚種 31 種 210 尾を対象として調 査が行われ、2 種 9 尾 (Abudefduf bengalensis (6/12) および Abudefduf whitleyi (3/22)) に寄生が確認され るとともに、別に入手した同海域のアジ科の Caranx sexfasciatus 1 尾にも同様の寄生が確認された[14]。
シストは 3-5mm で、スズメダイ科では1尾あたり 2-4 個、アジ科では多数のシストが検出された。外に、 Whipps et al. [88]は、グレート・バリア・リーフで 得たハタンポ類 (スズキ目スズキ亜目ハタンポ科)の Pempheris ypsilychnus での寄生を記録している。更に 遡り、1970 年代に Heron 島で養殖が試みられたヒラマ サ Seriola lalandi で発生したクドア高度感染の原因が K. amamiensis ではないかと推測されている[14]。 感染魚のシストに見出される胞子は 4 殻片 / 極嚢をも つ亜四角形の頂上像をもち、全体としては正位置の富有 柿のイメージである。それぞれの殻片には小さな頂上突 起と側方上部の外面に複数個の疣状突起が観察される。 側面像は類円形である。Burger et al. [14]による計測 では、胞子幅 6.8-8.4 (7.8)μm、胞子厚 5.8-7.3 (6.7)μm、 胞子長 4.6-6.0 (5.3)μm、極嚢長 1.8-2.7 (2.0)μm、極嚢 幅 1.1-1.8 (1.4)μm で小さめの胞子であるが、Egusa & Nakajima [23]の原記載では更に小型の胞子として特 徴づけられている。日本および豪州の各種魚類宿主 12 種から得た K. amamiensis の分子遺伝学的な解析では、 18S rDNA 1518 塩基長で最大7塩基の種内変異 (塩基 配列同一性は ≥99.5%) が確認されている[14]。 2-2.Kudoa azoni Asseva, 2004
北海道の対岸に位置する沿海地方ウスリー湾の浅海 (日本海)で漁獲されたホッケ Pleurogrammus azonus (74 尾中 3 尾)とスジアイナメHexagrammos octogrammus (6 尾中 2 尾)の体側筋に紡錘形のシスト(2.5-5.0mm × 0.5-1.2mm)を形成する4殻片 / 極嚢のクドアで、頂 上像は亜四角形、側面像では頂上の丸いピラミッド形で あり、殻片遠位端が尖る胞子形態を取る[7]。北海道沖 合の日本海で漁獲されたホッケの体側筋からは後述す る Kudoa pleurogrammi の寄生が知られるが、当該種 はシュードシストを形成する4殻片 / 極嚢のクドアであ り、明確に区別される。胞子幅 6.2-7.2μm、胞子厚 5.2-6.0μm、胞子長 5.0-6.0μm、極嚢長 2.0-2.2μm、極嚢幅 1.0-1.5μm で小さめの胞子である[7]。
2-3.Kudoa bora Fujita, 1930
台湾南部の水産試験場で飼養されていたボラ Mugil cephalus の体側筋に、約 2mm 径のシストを形成する4 殻片 / 極嚢のクドアとして報告された[30]。胞子は頂 上像で円形、胞子幅ならびに胞子厚は 11-12μm、胞子 長 8.0-8.5μm であり、殻片間の縫合線は明瞭、指状の 頂上突起をもつ。極嚢は長洋梨状で、極嚢長 5.5μm、 極嚢幅 1.8μm で前端近くが屈曲する。藤田[30]以降、 本種の報告はなく、日本沿岸のボラでは未確認である [21]。胞子の計測値は微妙に異なるが、下述のように、 日本近海や地中海の種々のスズキ目魚類に寄生が知られ る K. iwatai あるいは台湾南部の養殖場のフエダイの全 身組織に感染し種記載された K. lutjanus とよく類似し た胞子形態である。実際、Diamante et al. [19]は地中 海産ボラの筋肉に寄生するクドアを K. iwatai として報 告している。また、Narasimhamurti & Kalavati [69] がインド産ボラの視床葉にシスト形成するクドア(胞子 は亜四角形、9.0 × 9.0μm)として Kudoa tetraspora を 報告しているが、本種も同様に K. iwatai あるいはその 類縁クドアとして、今後の再確認が必要である。 2-4.Kudoa iwatai Egusa et Shiomitsu, 1983
宮崎県産養殖イシガキダイ Oplegnathus punctatus お よび鹿児島県産養殖マダイ Pagrus major の体側筋に 1mm 程度のシストを形成する4殻片 / 極嚢のクドアと して報告された[24]。その後、国内からは、クロダイ Acanthopagrus schlegelii、ブリ、サワラ Scomberomorus niphonius、 ス ズ キ Lateolabrax japonicus、 ヘ ダ イ Rhabdosargus sarba、ボラ、マゴチ Platycephalus sp. な どでの寄生が記録されている[3, 21, 39, 59, 80]。シス トの大きさは 0.5-1.5mm の球形もしくは卵円形[24]、 あるいは 0.5-2.8mm × 0.3-1.6mm の卵円形である[59]。 Diamant et al. [19]は、地中海で養殖されるヨーロッパ ヘダイ Sparus aurata、ヨーロッパスズキ Dicentrarchus labrax、ボラや同海域の 10 種以上の天然海産魚におけ る K. iwatai の寄生を報告し、また 18S rDNA 塩基配列 を日本産マダイ由来株、地中海産ヨーロッパヘダイ由来 株、および Siganus rivulatus 由来株で比較し、1560 塩 基長で3つの塩基置換を確認している。国内で分離され る K. iwatai の塩基配列はすべて前述の日本産マダイ由 来株と完全に一致している[39, 59]。地中海産各種魚 類での K. iwatai の寄生部位は、体側筋、頭蓋腔内の脂 肪組織、神経束、口唇やその周囲、眼窩組織、腸管膜お よび体腔、鰾、腸管筋層、心臓や心嚢、腎臓、卵巣など 全身臓器・組織に及ぶ[19]。国内産魚からは主として 体側筋からの確認であるが、眼窩組織などでも確認はさ れている[59]。東京都の食中毒事例では、生鮮ヘダイ Rhabdosargus sarba の喫食が原因と疑われる事例もあ り[80]、また、桃山・天社[65]は周防灘で漁獲され たコイチ Nibea albiflora の体側筋に形成された白色シス トから 4 殻片 / 極嚢の Kudoa sp. を記録するとともに、
その生鮮魚肉を喫食した 3 名が食中毒を発症し、一方、 加熱した魚肉を喫食した 1 名は無事であったことを記録 している。この寄生種も K. iwatai と考えられる[39]。 胞子は頭頂像で亜四角形、縫合線は明瞭、側面像は巾 着形で、胞子幅 8.8-11.6 (10.1)μm、胞子厚 7.9-10.4 (9.7) μm、胞子長 7.2-9.4 (8.4)μm、極嚢は長洋梨状で極嚢 長 3.7-5.4 (4.5)μm、極嚢幅 1.5-2.7 (2.2)μm、頂上突起 は指状で明確に観察される[24, 59]。
2-5.Kudoa lutjanus Wang et al., 2005
台湾南部で養殖されていたフジイロフエダイ Lutjanus erythropterus の全身組織 (脳、眼組織、鰾、体側筋、各 種内臓およびその漿膜)に最大 6.5mm の円形〜楕円の 乳白色のシストを形成する4殻片 / 極嚢のクドアとして 報告された[85]。この養殖場での集団感染では、無作 為に選んだ感染魚 10 尾において、脳に見つかったシス ト数が平均 180 (最大 839) に対して、体側筋および内 臓では平均 30-40 (最大 159) であったと記録されてい る。胞子は頂上像で亜四角形、縫合線は明瞭、側面像は 巾着形で、胞子幅 7.4-9.2 (8.2)μm、胞子長 6.3-7.9 (7.3) μm、極嚢長 3.0-4.2 (3.6)μm、極嚢幅 1.5-2.6 (2.2)μm で極嚢は長洋梨状、明確な指状の頂上突起をもつ点は K. iwatai に酷似する[85]。胞子の形態の類似性に加え、 18S rDNA 塩基配列においても、829 塩基長と部分的な 塩基配列しか利用できないが、日本産 K. iwatai とは 3 塩基置換、地中海産 K. iwatai とも 3 塩基置換、そして 日本産および地中海産 K. iwatai 株間には同一塩基長で 2 塩基置換があり、K. iwatai と同一種の可能性を考え る研究者もいる[59]。前述の台湾南部養殖ボラ寄生の K. bora との異同も含め、今後の精査が必要である。 2-6.Kudoa ogawai Yokoyama et al., 2012
2008 年 12 月 に 長 崎 県 で 水 揚 げ さ れ た メ ダ イ Hyperoglyphe japonica の体側筋に白色〜クリーム色の 楕円形のシストをつくり、そのサイズは 1-2mm であっ た。その冷凍切り身を神奈川県水産試験場から提供され た Yokoyama et al. [99]が精査し新種とした。胞子は 大きさの等しい 4 つの殻片 / 極嚢をもち、頂上像は各殻 片の遠位端が縫合部に対して花弁状に大きく丸く張り出 す特徴的な形態を示す。側面像はニンニク玉状で、各殻 片の遠位端が頂上像と同様大きく丸く張り出す。極嚢は 胞子サイズに比して小さめで、短洋梨状である。胞子の 計測値は、胞子幅 12.0-14.2 (13.3)μm、胞子厚 9.7-11.7 (11.0)μm、胞子長 8.3-9.6 (9.0)μm、極嚢長 1.9-3.2 (2.5) μm、極嚢幅 1.7-3.0 (2.5)μm である[99]。また、横山・ 長澤[96]はその著作「養殖魚介類の寄生虫の標準和名 目録」において、本種の宿主としてヒラメを追加している。 2-7.Kudoa pericardialis Nakajima et Egusa, 1978 1966 年 9 月に愛媛県の養殖ブリの心膜腔にクドアに よる白色シストが見いだされたのを皮切りに、鹿児島、 宮崎、高知、徳島、三重、そして静岡と西日本を中心と した複数の県の養魚場においてブリの心膜腔寄生クドア が確認された[68]。そこで Nakajima & Egusa [68] は 1976 年 9 月、静岡県網代湾の養殖場で 30 尾のブリを 対象に心膜腔のクドア感染について調査し、14 尾に感 染を検出した。白色ないしは黄色味を帯びたシストは 楕円形でその大きさは変化に富み(0.03-2.7mm × 0.02-1.2mm)、胞子は縫合線の明瞭な円形で、4 つの細長い 洋梨状の極嚢を有し、後述する同じく心膜腔寄生の K. shiomitsui と類似した胞子形態である。胞子の計測値 は、胞子幅 6.0-7.0μm、胞子厚 4.5-5.0μm、胞子長 4.0-4.2 μm、極嚢長 2.4-3.0μm、極嚢幅 1.0-1.5μm である[68]。 江草・塩満[24]は、1982 年に鹿児島県の養殖トラフ グ Takifugu rubripes か ら 心 膜 腔 寄 生 の K. shiomitsui を見出しているが、寄生部位や胞子の形態が類似する本 種とは、胞子サイズの違いや胞子頂上部の構造の違いで 別種とした。種記載以降に本種の記録はなく、一方の K. shiomitsui は各種魚類から報告されている。本種の胞子 頂上部の特徴とされた構造は明らかに走査電子顕微鏡標 本作製に伴うアーティファクトであり、両種の違いは胞 子計測値のみであるが、こちらも固定状態の差を反映し ている可能性がある[41]。両種の異同については今後 の大きな課題の1つである。
2-8.Kudoa scomberi Li et al., 2013
山口市内で市販されていた長崎県産のマサバ Scomber japonicus から検出され報告された[54]。本種のシス トは微小ながら肉眼で確認可能な 0.30-0.75mm × 0.20-0.40mm の白点として見られた。胞子の頂上像は亜四 角形、側面像は楕円形であり、洋梨形の大きさの等し い 4 つの極嚢を持つ。殻片表面は平滑である。ペルー 沖の南東太平洋のマサバから Kudoa caudata Kovaljova et Gajevskaja, 1983 が報告されているが、本種と違い、 殻片の側方にそれぞれ1本の線状構造が突出している [51]。胞子の計測値は、胞子幅 8.2-10.5 (9.2)μm、胞 子厚 7.0-8.8 (8.1)μm、胞子長 6.1-6.8 (6.4)μm、極嚢長 2.5-3.4 (2.9)μm、極嚢幅 1.3-2.0 (1.6)μm である[54]。
2-9.Kudoa sebastea Asseva, 2004 北海道の対岸に位置する沿海地方の浅い湾海(日本 海)で漁獲されたメバル属魚類の 1 種 Sebastes minor の体側筋(6 尾中 2 尾)の体側筋にシスト(2.0-5.0mm × 1.0-2.5mm)を形成する 4 殻片 / 極嚢の種で、頂上像 は亜四角形、側面像では栗形であり、前端が尖り底面が 丸い[7]。胞子幅 7.3-8.2μm、胞子厚 5.0-5.5μm、胞子 長 5.4-5.6μm、極嚢長 2.0-2.2μm、極嚢幅 0.8-1.0μm で 小さめの胞子である[7]。
2-10.Kudoa shiomitsui Egusa et Shiomitsu, 1983 江草・塩満[24]は、1982 年に鹿児島県内の複数の 養魚場から任意に採集したトラフグ Takifugu rubripes 計 30 尾を検査し、そのうちの 16 尾の心膜腔にクドアの シストを見出して報告した。心膜腔内のシストは白色な いしクリーム色のやや扁平な楕円体で、シストの数は数 個から約 200 個と様々で、その数が多いほどシストの平 均的な大きさは小さい傾向にあった。胞子の頂上像は縫 合線の明瞭な円形で、側面像は下部が丸く膨らみ、上面 が平らな柿の実様であった。極嚢は 4 つでやや曲がった バナナ状で長軸をほぼ水平にして四方に放射するように 位置していた。米国メキシコ湾に導入された養殖トラ フグでも検出されている[9]。また、Kasai et al. [41] は益田港水揚げの天然カワハギ Stephanolepis cirrhifer と高知県水揚げの天然ウスバハギ Aluterus monoceros の心膜腔での寄生を報告している。外に、Zhang et al. [103]は 2007 年から 2008 年にかけて長崎県での養殖ク ロマグロのクドア感染についての調査し、感染率には 季節性があるものの、高い感染率(最大 100%)で本種 の寄生が確認されることを報告した。また Tsutsumi et al. (未発表)は三重で水揚げされたヒラメから得られた 本種の 18S rDNA 塩基配列を GenBank に登録している (DDBJ/EMBL/GenBank accession no. AB183718)。 こ の塩基配列は、上記メキシコ湾産トラフグ由来株、なら びに日本海産カワハギ由来株と 1,700 塩基長で完全に一 致するが、長崎県産クロマグロとは 99.7% (1,724/1,730) の一致にとどまる[41]。K. shiomitsui の 28S rDNA 塩 基配列は最近まで得られておらず[41]、解像度の高い 28S rDNA 塩基配列の利用を通して分離株間の関係を更 に解析していくことも有意義と考えられる。豪州グレー ト・バリア・リーフ (Lizard 島周辺) のイトヒキテンジ クダイ Zoramia leptacantha の心膜腔にシスト形成する クドアとして Kudoa leptacanthae Heiniger et Adlard, 2012 が報告されているが、K. shiomitsui カワハギ由来株 との 18S rDNA 塩基配列での同一性は 99.6%(1649/1656) と高い[41]。洋梨状とバナナ状という極嚢の形状に違 いがあるとされるが[36]、計測値には大きな違いが見 出せない。わが国では本種の外に心膜腔寄生のクドアと して K. pericardialis が Nakajima & Egusa [68]によっ て記載されているが、これら 2 種の異同については今後 検討する余地があることは前述したとおりである。また、 紅海産ヨーロッパマダイ Pagrus pagrus の心膜腔にシス ト形成する Kudoa pagrusi や米国メキシコ湾のエボシダ イ Nomeus gronovii の心膜腔にシスト形成する Kudoa hypoepicardialis など[1, 9]、海産魚の心膜腔にシスト 形成するクドア種は 18S rDNA 分子系統樹上に1つの クレードをつくり類縁性が高いが、これらがどのように 種分化し地理上の分散を遂げたのか、今後の研究に期待 したい。K. shiomitsui 胞子の計測値は、胞子幅 8.6-9.8 (9.4)μm、胞子厚 6.7-7.5 (7.2)μm、胞子長 5.6-6.8 (6.2) μm、極嚢長 2.5-3.0 (2.8)μm、極嚢幅 1.0-1.4 (1.3)μm である[24]。
2-11.Kudoa trachuri Matsukane et al., 2011
2008 年 9 月と 12 月に長崎県で水揚げされた日本海産 マアジ Trachurus japonicus の体側筋から見出され報告 された[59]。本種は体側筋に楕円形の白色シストを形 成し、胞子は 4 殻片 / 極嚢で亜四角形の頂上像と楕円形 の側面像をもち、極嚢は大きさの等しい洋梨状であった。 胞子の形態は K. thunni と類似しているが、胞子計測値 は、胞子幅 7.0-8.5 (7.9)μm、胞子厚 5.3-6.2 (5.8)μm、 胞子長 5.5-6.9 (6.1)μm、極嚢長 2.6-3.5 (2.9)μm、極嚢 幅 1.6-2.2 (2.0)μm で、本種の方が胞子幅と胞子厚が小 さく、また、両種間には 18S rDNA および 28S rDNA の塩基配列に有意な差があることで区別できる[59]。 Kasai et al. [39]は 2014 年 6 月に山口県萩沖の日本海 で水揚げされたマアジから得た本種について internal transcribed spacer (ITS) 領域を含めた 1984 塩基長の 塩基配列を報告し、Matsukane et al. [59]が報告した 塩基配列を再確認している。
2-12.Kudoa thunni Matsukane et al., 2011
2008 年 10 月に山口市内で市販されていた太平洋産 メバチマグロ Thunnus alalunga の刺身に白色球形のシ ストが確認されたことで報告された[59]。胞子は 4 殻 片 / 極嚢で亜四角形の頂上像と楕円形の側面像をもち、 極嚢は大きさの等しい洋梨状であった。胞子の計測値 は、胞子幅 9.2-9.9 (9.5)μm、胞子厚 7.7-9.0 (8.3)μm、
胞子長 6.4-6.6 (6.5)μm、極嚢長 2.2-2.9 (2.5)μm、極嚢 幅 1.9-2.2 (2.1)μm であった[59]。DNA データベース (DDBJ/EMBL/GenBank)に登録される『K. crumena』 との異同が問題となるが、米国のノースキャロライナ沖 大西洋産キハダマグロ Thunnus albacares (18S rDNA− AF378347)と豪州のタスマニア島沖の太平洋産ミナミマ グロ Thunnus maccoyii (28S rDNA−FJ417057)か ら の クドアには形態記録がないままに K. crumena Iversen et van Meter, 1967 として塩基配列が登録されている。 当該種は大西洋フロリダ沖のスパニッシュ・マカレル Scomberomorus maculatus (サバ科、サワラの類) の体 側筋に見出された種で、その胞子の側面像は巾着状で あり、上記の K. thunni 胞子が側面像では楕円である点 で大きく異なっている。最近、Griffin et al. [33]はカ リブ海と北大西洋との境界に位置するセントキッツ島 沖で漁獲されたタイセイヨウマグロ Thunnus atlanticus から本種を見出し報告した。また、Kasai et al. [43] は、東南アジア沖の太平洋で漁獲されたキハダマグロ Thunnus albacares から本種 4 分離株を得て 18S rDNA および 28S rDNA 塩基配列についてこれまでの報告を 整理し、現在のところ K. thunni は一定範囲の種内変異 が両 rDNA 領域で見られること、複合種である可能性 も含めて更なる材料収集と塩基配列の収集が必要である としている。なお、18S rDNA および 28S rDNA に基 づく分子系統樹では、マアジからの K. trachuri、マサ バからの K. scomberi、豪州グレート・バリア・リーフ のスズメダイ科からの Kudoa kenti Burger et Adlard, 2009 といった体側筋にシストを形成する種との近縁性 が明らかであるが、現在までのところ、マグロ寄生 K. thunni 分離株との分子系統樹上で混在はなく、すべて が独立種であると考えられる[43]。
3.シュードシスト形成するクドア
3-1.Kudoa akihitoi Kasai et al., 20162016 年 2 月に有明海に棲息するハゼクチ Acanthogobius hasta 6 尾を検査した Kasai et al. [42]は、そのうちの 2 尾の体側筋に本種のシュードシストを検出した。下述 の K. empressmichikoae との混合感染であり、1 尾当た り 74 個もしくは 106 個の総検出プラズモディウムのう ちのどれだけが本種に因るものかは不明である。プラズ モディウムのサイズは 0.79-5.40 (1.98) mm × 0.12-0.44 (0.26) mm で、こちらについても 2 種間の違いは混合 感染のため不明である。4 殻片 / 極嚢で頭頂像は菱形の クドアで、4 つの極嚢のうちの 1 つが特に大きい胞子の 形状は K. thyrsites や K. whippsi 胞子に類似するが、 その大きさは小さめである[42]。側面像は対象性に欠 けたピラミッド形で殻片遠位部が尖り、胞子幅 9.1-12.5 (10.6)μm、胞子厚 6.8-9.4 (8.1)μm、縫合部胞子厚 5.3-6.7 (6.1)μm、胞子長 5.3-7.0 (6.4)μm、極嚢長(大) 4.2-5.1 (4.8)μm、極嚢幅(大) 2.2-2.8 (2.5)μm、極嚢長(小) 2.8-4.6 (3.7)μm、極嚢幅(小) 1.5-2.3 (1.9)μm である [42]。18S および 28S rDMA 塩基配列に基づく分子系 統樹では、胞子形態が酷似する前述の K. thyrsites や K. whippsi など一群のクドアと同一のクレードに入る が、他種とは明確に区別される。すなわち、18S rDNA での塩基配列の同一性は 96.6-98.2%、28S rDNA での 塩基配列の同一性は 91.4-95.6% であり[42]、塩基配列 の同一性に基づく種鑑別は容易である。なお、胞子形 態が菱形で 4 極嚢のうちの 1 つが特に大きい胞子をも つクドアは、本種を含めて日本近海では 6 種が報告さ れている。すなわち、K. thyrsites、K. lateolabracis、 K. megacapsula、K. parathyrsites、K. whippsi、K. akihitoi である[41, 42, 94, 97]。
3-2.Kudoa empressmichikoae Kasai et al., 2016 前述の K. akihitoi と共に、2016 年 2 月に入手した有 明海に棲息するハゼクチ 6 尾すべての体側筋に本種の シュードシストが検出されている[42]。上述のとおり、 K. akihitoi とプラズモディウムのサイズ差は明らかで ない。混合感染のあった 2 尾を除いた単独感染 4 尾での プラズモディウム検出数は 1 尾当たり 51-70 (57) 個で あった。4 殻片 / 極嚢で頂上像は縫合線の明瞭な球形の 胞子体が、小三角形に突出する殻片下部に載っている かのような特異な形状をもつクドアで、4 つの極嚢は短 洋梨形で小さめである。側面像は、丸い胞子体をつく る殻片がその下部で突出するかたちをとり、殻片毎に 1 つの指状に伸びる明瞭な頂上突起が確認できる[42]。 胞子計測値は、胞子幅 9.2-11.8 (10.5)μm、胞子厚 6.2-9.1 (7.5)μm、縫合部胞子厚 4.8-7.0 (5.7)μm、胞子長 5.1-6.1 (5.6)μm、極嚢長 1.7-2.7 (2.1)μm、極嚢幅 1.2-1.8 (1.5)μm である[42]。殻片下部が突出して本種に 似るクドアとしては、豪州グレート・バリア・リーフ に棲息するホシカイワリ Carangoides fulvoguttatus、 インドカイワリ Carangoides plagiotaenia、ロウニンア ジ Carax ignobilis などのアジ科魚類から記録された K. quadricornis Whipps et al. 2003 および K. paraquadricornis Burger et Adlard, 2010 などが知られるが[10, 87]、後
2種の胞子体部は卵円形で、極嚢も長く伸びて棍棒状に 近い。また、18S および 28S rDNA 塩基配列に基づく分 子系統樹では、豪州で記録された2種(K. quadricornis と K. paraquadricornis)は1つのクレードに入るが、K. empressmichikoae は瀬戸内海産のウマヅラハギの体側 筋から見いだされた K. parathyrsites とクレードをつく り、胞子形態が菱形で4極嚢のうちの1つが特に大きい 胞子をもつ一群のクドアと類縁性が高いことが示されて いる[42]。なお、K. parathyrsites との 18S rDNA での 塩基配列の同一性は 99.4%(1,560 塩基長)、28S rDNA での塩基配列の同一性は 95.7%(553 塩基長)である。 3-3.Kudoa hexapunctata Yokoyama et al., 2014 Li et al. [54]ならびに Abe & Maehara [2]によっ て、キハダマグロ Thunnus albacares およびクロマグロ Thunnus orientalis の体側筋にシュードシストを形成し て寄生する 6 殻片 / 極嚢のクドアの分子遺伝学的特徴づ けが報告された。Arai & Matsumoto [6]がバンダ海域 で漁獲され築地市場に搬入されたキハダマグロで確認し た死後筋肉融解現象の原因 Kudoa neothunni と類似す る形態から、異なるマグロ種に寄生する K. neothunni の 2 遺伝子型としての報告であった。Yokoyama et al. [102]は、キハダマグロに専ら寄生する種としての K. neothunni と、クロマグロならびにキハダマグロに寄生 する種として K. hexapunctata を形態学的に区別すると ともに(頂上像での殻片遠端部の鋭度が主たる鑑別点 となる)、これまでに報告されていた 18S rDNA (両種 間の塩基置換は 1,741 塩基長で 2 塩基部位)および 28S rDNA (両種間の塩基置換は 2,876 塩基長で 23 塩基部 位)塩基配列をそれぞれの種の鑑別点とした。実際、異 なる研究グループが異なるソースから収集した両種の 18S rDNA ならびに 28S rDNA 塩基配列の種内変異は ほぼ皆無である[2, 43, 54, 102]。生鮮ヒラメ喫食によ る食中毒とともに、クロマグロ若魚 (ヨコワ) 喫食によ る食中毒集団発生も疑われる事例も頻発していることか ら[80]、ヒラメ寄生の食中毒原因種 K. septempunctata と同様の食中毒病原性の in vitro 解析が実施され、その 病原性は確認されたが、前述種ほどには強くなかったと された[81]。最近になり、長崎県水揚げの東シナ海産 コシナガ Thunnus tonggol での軽度の寄生も確認され ている[43]。胞子幅 8.0-11.8 (10.4)μm、胞子厚 7.0-10.2 (9.1)μm、胞子長 5.0-8.4 (7.5)μm、極嚢長 2.2-4.0 (2.5) μm、極嚢幅 1.3-2.2 (1.7)μm である[102]。
3-4.Kudoa igami Shirakashi et al., 2014
和歌山県沖の北西太平洋で漁獲されたブダイCalotomus japonicus 41 尾のうち 12 尾 (29.3%) の体側筋にシュー ドシストを形成する 6 殻片 / 極嚢の胞子をもつ種で、胞 子の頂上像は K. hexapunctata に類似しているが、殻片 の遠端部はより丸味をもつ[77]。側面像は蜜柑のよう な類円形である。6 殻片 / 極嚢の胞子が優勢であるが、 5 殻片 / 極嚢の胞子も混在し、両者の比率は 81:19 であっ た[77]。本種が検出されたブダイからは、体側筋に 同じくシュードシスト形成する K. lateolabrasis (17/41 [41.5%])と K. thalassomi (1/41 [2.4%])の寄生、脳 に寄生する K. yasunagai (16/17 [94.1%]) の寄生も確 認された。胞子幅 9.2-10.8 (9.9)μm、縫合部胞子幅 7.0-9.1 (7.9)μm、胞子厚 8.5-9.6 (9.1)μm、胞子長 6.2-7.3 (6.5) μm、極嚢長 2.8-4.0 (3.3)μm、極嚢幅 1.4-2.1 (1.7)μm である[77]。
3-5.Kudoa intestinalis Maeno et al., 1993
三重県志摩半島の五ヶ所湾水揚げのボラMugil cephalus の腸管平滑筋にシュードシスト形成する4殻片 / 極嚢の クドアとして報告された[57]。胞子は頂上像で亜四角形、 縫合線は明瞭、側面像は巾着形で底面は凹み、胞子幅 6.3-7.0 (6.5)μm、胞子厚 5.8-6.5 (6.1)μm、胞子長 3.0-3.5 (3.4)μm、極嚢長 1.3-1.5 (1.5)μm、極嚢幅 1.0-1.5 (1.2) μm で極嚢は小さな類円形、極糸は 2.5-3.5 (2.7)μm と 短く、また、小さな頂上突起をもつ[57]。
3-6. Kudoa lateolabracis Yokoyama, Whipps et Kent, 2004 中国から稚魚を輸入し愛媛県沖の瀬戸内海で養殖され ていたタイリクスズキ Lateolabrax sp. の体側筋に死後 筋肉融解現象を引き起こしたクドアとして種記載され た。4 殻片 / 極嚢で頂上像は菱形のクドアで、4 つの極 嚢のうちの 1 つが特に大きい胞子の形状は K. thyrsites 胞子に酷似するが、その大きさはやや小さめである[97]。 タイリクスズキは、日本近海産のスズキ Laterolabrax japonicus に比べ、淡水あるいは汽水への順応性があり、 成長が早いことから養殖魚としての大きな可能性を秘め ていると目論まれたが、輸入する種苗によっては出荷 段階において本種による市場価値喪失が起こり得る。 最近、西日本での養殖ヒラメ Paralichthys olivaceus や 和歌山県沖の北西太平洋産ブダイでの寄生報告もある [32, 77]。胞子の側面像は対象性に欠けたピラミッド形 で、胞子幅 9.9-12.9 (11.5)μm、胞子厚 8.4-9.9 (9.3)μm、
胞子長 5.4-6.9 (6.4)μm、極嚢は洋梨状で、極嚢長(大) 4.0-5.9 (5.2)μm、極嚢幅(大) 2.5-3.5 (2.8)μm、極嚢 長(小) 3.0-4.0 (3.6)μm、極嚢幅(小) 1.5-2.0 (1.9)μm である[97]。
3-7.Kudoa megacapsula Yokoyama et Itoh, 2005 中国産アカカマスを輸入して干物に加工する過程で筋 肉が融解した原因として報告された。体側筋にシュード シストを形成するクドアで、4殻片 / 極嚢の胞子をもつ が、1つの極嚢のみが特別に大きく、残りの3つの極嚢 は痕跡的といえるほどに小さい[94]。胞子は4つの殻 片がヒトデの腕のように伸び、殻片の先から先まで最大 で 30.3-44.7 (40.0)μm、最小で 25.6-34.9 (30.9)μm、縫 合部胞子厚 11.0-12.9 (12.0)μm、胞子長 9.3-15.4 (13.4) μm、特大の極嚢は極嚢長 11.7-14.2 (12.7)μm、極嚢幅 4.9-6.8 (5.8)μm、小さい極嚢は極嚢長 1.5-4.4 (2.8)μm、 極嚢幅 1.0-1.5 (1.2)μm である[94]。ブリでは、韓国 産の種苗を用いて養殖した 1 歳魚に見られ報告された [98]。この寄生魚では、死後筋肉融解現象は見られず、 代わりに、黒色変性したプラズモディウムを囲む肉芽腫 があたかもシストのように体側筋に散在していた。国内 の養殖魚では見られないことから、大陸側の日本海ある いは東シナ海に分布するクドアであろうと推測されてい る[98]。
3-8. Kudoa musculoliquefaciens (Matsumoto, 1954) Egusa, 1986 1953 年 7 月、宮城県気仙沼水揚げの死後筋肉融解現 象を呈した三陸沖産メカジキ Xiphias gladius の体側筋 を検査した Matsumoto [60]が、シュードシスト形成 する粘液胞子虫 Chloromyxum musculoliquefaciens とし て当初報告した。本種は体側筋の筋線維内にシュードシ ストを形成し、4 つの大きさの等しい殻片 / 極嚢の胞子 を持つ。胞子の頂上像は縫合線の明瞭な亜四角形で円形 に近く、側面像は楕円形である。胞子の計測値は、胞 子幅 7.4-9.9 (8.3)μm、胞子厚 7.0-8.9 (7.8)μm、胞子長 5.3-7.3 (6.2)μm、極嚢は類円形で極嚢長 1.7-2.8 (2.1) μm、極嚢幅 1.7-2.5 (2.0)μm である[60]。Kasai et al. [40]は、これまで形態学的な記述にとどまり分子系統 学的な位置づけがなされていなかった本種について解析 を目論み、日本国内で購入した三陸沖産および北インド 洋産メカジキと高知県水揚げのインドー太平洋産バショ ウカジキ Istiophorus platypterus における本種の感染を 確認するとともに、分子系統樹解析の結果、本種は豪州 の軟骨魚類から得られているクドア 2 種 (K. hemiscylli と K. carcharhini) に最も近縁であることを示した。な お、これら 2 種の胞子形態も K. musculoliquefaciens に 酷似しているが[31]、実際の 18S および 28S rDNA 塩 基配列の同一性はそれぞれ 95.24%-96.08% (1,575 ある いは 1,580 塩基長)、71.82%-73.76% (785 あるいは 763 塩基長)と必ずしも高くはない[40]。小長谷[47]は 本種による死後筋肉融解現象の組織学的観察を報告して いる。
3-9. Kudoa neothunni (Arai et Matsumoto, 1953) Whipps et al., 2004 1952 年 8 月、バンダ海域で漁獲され築地市場に搬入 されたキハダマグロにジェリーミートが多発し、その原 因究明にあたった Arai & Matsumoto [6]により 6 つ の殻片 / 極嚢を持つ星形に殻片遠位端が突出する新属新 種の粘液胞子虫 Hexapunctata neothunni として報告さ れた。その後、小長谷 [45, 46, 48]によって、1969 年 同海域で漁獲されたキハダマグロ、1989 年台湾沖で漁 獲されたメバチマグロ Thunnus obesus、1980 年三陸沖 で漁獲されたクロマグロの各ジェリーミートからも検 出されている。前述したように、Li et al. [54]ならび に Abe & Maehara [2]はキハダマグロおよびクロマ グロ若魚(メジマグロ)の体側筋にシュードシストを 形成する 6 殻片 / 極嚢のクドアを Kudoa neothunni の 2 遺伝子型としたが、後者クロマグロ寄生種については Yokoyama et al. [102]が K. hexapunctata として区別 するとともに、キハダマグロでの寄生も見られることを 示した。Kasai et al. [43]は、長崎県水揚げの東シナ海 産コシナガマグロ Thunnus tonggol から得られた 7 つ の殻片 / 極嚢のクドアについて、18S および 28S rDNA 塩基配列のほぼ完璧な一致から本種として報告してい る。すなわち、キハダマグロ寄生では 6 殻片 / 極嚢の胞 子が確認され、コシナガマグロでは 7 殻片 / 極嚢の胞子 が見られることになるが、異なる形態型として固定され たポピュレーションであるかどうかは、今後の検討課 題である。キハダマグロ寄生での胞子計測値は、胞子 幅 9.1-13.0 (11.0)μm、胞子厚 5.9-8.7 (7.1)μm、胞子長 5.3-7.3 (6.2)μm、極嚢長 2.0-3.1 (2.5)μm、極嚢幅 1.3-2.1 (1.6)μm である[6]。
3-10.Kudoa paralichthys Cho et Kim, 2003
Cho & Kim [6]が韓国の養殖ヒラメの脳に寄生する 4 殻片 / 極嚢のクドアとして報告した。大きさの等しい
4 殻片 / 極嚢を持つ胞子亜正四角形の頂上像をもち、楕 円形の側面像を持つ、極嚢は短洋梨状で胞子の頂上部に 構造物は見られなかった。胞子サイズは、胞子幅 7.4-8.9 (8.2)μm、胞子厚 6.3-7.7 (6.9)μm、胞子長 4.5-6.0 (5.2) μm、極嚢長 1.9-2.7 (2.2)μm、極嚢幅 1.0-1.4 (1.2) μm である[16]。
3-11.Kudoa parathyrsites Kasai et al., 2016
瀬戸内海産のウマヅラハギ Tamnaconus modestus の 体側筋から見いだされた 4 殻片 / 極嚢のクドアである が、4つのうちの1つの極嚢が特に大きい[41]。2013 年 12 月から 2015 年 7 月の期間に、西日本各地で水揚げ されたカワハギ科魚種 4 種 75 尾を対象にクドア感染に ついて検査し、ウマヅラハギ 51 尾の内 1 尾の体側筋か ら K. septempunctata との混合感染として 2 個のシュー ドシストを得た。胞子は K. thyrsites に類似しており、 不等辺の星形で大きさが不揃いな 4 つの殻片と極嚢を 持っていたが、殻片遠位端の角が丸みを帯びているこ とが K. thyrsites と異なる点であった。18S および 28S rDNA 塩基配列に基づいた分子系統解析の結果、本種 は K. thyrsites など不等辺の星形で大きさが不揃いな 4 つの殻片 / 極嚢をもつクドアと同一のクレードに属した が、既知の K. thyrsites や類縁種とは有意な相違が見ら れた。前述したように、胞子形態が大きく異なるハゼク チ寄生の K. empressmichikoae と 18S および 28S rDNA 塩基配列が最も近い[42]。胞子の計測サイズは、胞子 幅 14.0-16.8 (15.2)μm、 胞 子 厚 9.7-13.3 (11.5)μm、 胞 子縫合部厚 7.7-10.5 (9.1)μm、胞子長 8.1-10.9 (9.5)μm、 極嚢長(大) 4.0-6.7 (5.1)μm、極嚢幅(大) 1.8-2.9 (2.5) μm、極嚢長(小) 2.5-4.6 (3.5)μm、極嚢幅(小) 1.3-2.6 (1.8)μm である[41]。現在までのところ、極めて限ら れた寄生記録であり、今後、更に自然界での寄生状況を 確認する必要がある。
3-12.Kudoa pleurogrammi Kasai et al., 2016
2014 年 7 月、山口市内で販売されていたキタノホッケ (いわゆるシマホッケ)Pleurogrammos monopterygius の開き干しの肉質が異常に柔らかいとして山口大学獣医 寄生虫学教室に持ち込まれ、筋肉にクドアのシュードシ ストが多数確認された[40]。その後、北海道沿岸で漁 獲されたホッケ Pleurogrammus azonus を 2015 年 9 月 に太平洋産(青森)と日本海産(北海道)6 尾ずつ計 12 尾を調べた Kasai et al. [40]は、そのうちの 7 尾にキ タノホッケ寄生種と同一のクドアを確認して、ホッケお よびキタノホッケ寄生種として報告した。本種は体側筋 に細長いシュードシスト(1.2-3.9mm × 0.11-0.26mm) を形成する 4 殻片 / 極嚢の種で、頂上像は亜四角形、側 面像では二枚貝状であり、頂部と底部が丸く膨らみ、殻 片中央部が両側に尖る。小さな頂上突起をもつ。胞子 幅 8.2-9.1 (8.6)μm、胞子厚 7.1-8.2 (7.4)μm、胞子縫合 部厚 5.4-7.7 (6.6)μm、胞子長 5.6-6.8 (6.3)μm、極嚢長 2.7-2.9 (2.8)μm、極嚢幅 1.4-2.0 (1.6)μm である[40]。 3-13.Kudoa prunusi Meng et al., 2011
和歌山県の養殖クロマグロの脳から見いだされて報告 された[62]。すなわち、2009 年 7 月から 8 月にかけて 和歌山県の養殖クロマグロ幼魚(平均体重 156.6g、平均 体長 19.3cm)を対象にクドア感染について検査したと ころ、9 尾すべての脳にクドアの感染が確認された。脳 の表面に多数のプラズモディウムがみられ、胞子は殻片 / 極嚢数が 5 ないし 6 で、その割合は 80:20 であった。 胞子の頂上像は角の丸い 5 放射状で桜の花の様な形状を 呈し、側面像は丸味のある巾着型、極嚢は洋梨状である。 胞子の計測値は、胞子幅 8.5-10.3 (9.6)μm、胞子長 6.7-8.6 (7.5)μm、 極 嚢 長 2.8-4.1(3.7)μm、 極 嚢 幅 1.7-2.2 (2.0)μm である[62]。クロマグロの脳寄生クドアとし ては、わが国では本種の外に K. yasunagai が知られて いる[103]。
3-14.Kudoa septempunctata Matsukane et al., 2010 2009 年 2 月から 3 月にかけて 60 尾の養殖ヒラメ(30 尾は日本産、30 尾は韓国産)を対象にクドア感染の有 無を検査した Matsukane et al. [58]は、韓国産養殖ヒ ラメ 1 尾の体側筋から大きさが不揃いな 7 つの殻片 / 極 嚢を持つクドアを得て報告した。和名は「ナナホシクド ア」。本種は体側筋にシュードシストを形成し、胞子は いびつな放射状の頂上像で、殻片 / 極嚢数は 7 つのもの が主で 6 のものも見られた(両者の比は 74:26)。その 後、他の感染魚についても同様に 7 殻片 / 極嚢と 6 殻 片 / 極嚢の胞子比をみると、67:33、40:60,34:66 と個体もしくはプラズモディウムによる変異が大きい ことが報告されている[54]。平成 23 年 6 月 17 日付食 安 0617 第 3 号「生食用生鮮食品による病因物質不明有 症事例への対応について」において、生食用のヒラメの K. septempunctata 並びに馬刺しの Sarcocystis fayeri が 原因として考えられる有症事例を今後食中毒事例として 扱うよう通知が出され、本種のヒトへの病害性が行政的 に認知され、公衆衛生領域における重要な病原体として
位置づけられた[44, 50, 78]。国内でのクドア食中毒の 発生は、2011 年が 34 件(発症者 473 名)、2012 年が 41 件(発症者 418 名)、2013 年が 20 件(発症者 288 人) であった[78]。当初は専ら養殖ヒラメを原因食とする 集団発症事例が多かったが、最近では天然ヒラメを原因 食とする集団発症事例も増えてきた[83]。Kasai et al. [41]は西日本各地水揚げのカワハギ科魚種 4 種 75 尾を 対象にクドア感染について検査し、ウマヅラハギ 51 尾 のうち日本海産(鳥取県水揚げ)1 尾と瀬戸内海産(山 口県水揚げ)計 2 尾の体側筋に本種の寄生を確認し、ヒ ラメ以外の魚種からの初記録を報告している。当該研究 で得られた本種の胞子は、殻片 / 極嚢の数が瀬戸内海産 で 5 から 8 個(7 が優勢)、日本海産で 5-6 個(6 が優 勢)と変異が見られたが、18S および 28S rDNA 塩基配 列は Matsukane et al. [58]で得られた韓国産ヒラメ寄 生の塩基配列と完全に一致している。胞子の計測値は、 胞子幅 11.1-13.1 (11.8)μm、胞子厚 8.9-10.0 (9.4)μm、 胞子長 7.9-8.9 (8.5)μm、極嚢長 3.7-5.3 (4.6)μm、極嚢 幅 2.2-2.8 (2.4)μm である[58]。 Takeuchi et al. [83]は、本種のミトコンドリア DNA、 す な わ ち cytochrome c oxidase subunit I (cox-1) と large-subunit rRNA (rnl)に注目し、区別された 3 つの サブポピューレーションと分布海域(日本と韓国)との 関連性を探り、特定系統が一方に頻発する傾向があった ことを報告した。
3-15.Kudoa thalassomi Adlard et al., 2005
2000 年から 2001 年にかけて豪州グレート・バリア・ リーフで 69 種類の珊瑚礁に棲息する天然魚を対象にク ドア感染について検査した Adlard et al. [4]は、ベラ 科魚類のムーンラス Thalassoma lunare の 3 尾中 1 尾の 体側筋にシュードシストを形成するクドアを報告した。 胞子は 6 殻片 / 極嚢であった。また、Burger & Adlard [12]はグレート・バリア・リーフの 2 環礁(Helon 島 周辺と Lizard 島周辺)の 41 科 214 魚種を対象にクドア 感染について検査し、18 魚種で本種の感染を記録して いる。わが国では Shirakashi et al. [77]によって和歌 山県産のブダイの体側筋から見いだされた。同研究に よると 2012 年から 2013 年にかけて和歌山県で採取さ れたブダイ 41 尾を対象にクドア感染について検査し、 1 尾に 6-7 殻片 / 極嚢の本種が確認されている。なお、 同研究では 41 尾中 17 尾から K. lateolabracis、12 尾か ら新種 K. igami、脳を検査した 17 尾中 16 尾から K. yasunagai の寄生を確認している。胞子サイズの計測値 は胞子幅 12.1-15.0 (13.7)μm、胞子厚 8.5-9.6 (9.1)μm、 胞子長 6.6-8.9 (7.7)μm、極嚢長 2.8-4.0 (3.3)μm、極嚢 幅 1.4-2.1 (1.7)μm である[77]。
3-16. Kudoa thyrsites (Gilchrist, 1924) Meglitsch, 1947 本種は広い宿主域と分布域をもち、死後筋肉融解現象 を引き起こす種として古くから知られる[66, 86]。日 本国内では Yokoyama et al. [97]が愛媛県の水産加工 場において筋肉の軟化を呈したヒラメの体側筋から見 いだしている。宿主のヒラメは韓国の種苗を愛媛県で 約1年間にわたり陸上養殖したもので、養殖場におい て肉質の変化が見られたヒラメは全体の 1% 未満であっ たとされている。また、Kasai et al. [41]はウマヅラハ ギの体側筋に寄生を確認した。本種は体側筋にシュード シストを形成し、菱形の頂上像と大小不同の 4 つの殻片 / 極嚢をもち、1つの極嚢が特に大きいことが特徴とな る。Yokoyama et al. [97]はタイリクスズキ寄生の K. lateolabrasis の新種記載に際し、Matsumoto [60]がス ズキの体側筋から見いだした K. cruciformum は本種の シノニムである可能性が高いと指摘している。また、同 様に、北大西洋ビスケー湾の Scomber scombrus (サバ科) の 体 側 筋 に 見 出 さ れ た K. histolytica (Peraad, 1928) Meglitsch, 1947 も本種のシノニムと考えられている [86]。胞子サイズは、最大胞子幅 7.9-11.9 (10.0)μm、最 小胞子幅 6.9-7.9 (7.6)μm、胞子厚 12.9-17.8 (14.7)μm、 胞子長 6.9-8.9 (7.9)μm、極嚢長(大) 4.0-5.9 (5.0) μm、 極嚢幅(大) 1.5-3.0 (2.5)μm、極嚢長(小) 2.0-4.0 (-2.8) μm、極嚢幅(小) 1.5-2.5 (1.6)μm である[97]。 世界各地の様々な魚種から報告のある K. thyrsites は 18S および 28S rDNA 塩基配列にもかなりの種内変異 がみられることから複合種と考える研究者もいるが、 Whipps & Kent [86]は5つの地理的系統(英国、南ア フリカ、オーストリア、カナダ、日本)にまとめられ る可能性を示した。日本産 K. thyrsites は、他の地理的 系統との間での 18S rDNA 塩基配列での同一性は 98.2% (1,692/1,723)-99.5% (1,558/1,566)、28S rDNA 塩基配列 での同一性は 91.7% (650/709)-94.1% (753/800)である。 ITS-1 領域ではより顕著な地理的系統間の塩基配列の違 いがみられるが[86]、2 もしくは数塩基を単位とした 繰り返しが多発することから、解析は必ずしも容易では ない[41]。 なお、胞子形態が菱形で4極嚢のうちの1つが特に 大きい胞子をもつクドアは、本種を含めて世界的に