報告
オウギガニの胃咀嚼器断面観察による年齢査定の試み
A preliminary investigation of the direct age determination using band counts in the gastric mill
of the xanthid crab Leptodius exaratus (H. Milne Edwards, 1834)
森川英祐
1・渡邊隆司
1*・浜崎活幸
1Eisuke Morikawa, Ryuji Watanabe and Katsuyuki Hamasaki
はじめに 年齢査定は水産資源の適切な利用を進めるうえで 基礎的な調査項目であるが,最近まで甲殻類は年齢 形質として有効な硬組織(鱗,耳石,脊椎骨,貝殻 など)を持たないと考えられてきた.このため,甲 殻類の年齢は体サイズの頻度分布におけるモード解 析,飼育による脱皮・成長量の把握,蛍光性色素リ ポフスチンの頻度分布解析などにより推定されてき た(渡邊ら,2012).特に蛍光性色素リポフスチン は,加齢とともに一定の速度で蓄積していくため, 近年ではリポフスチン解析による年齢推定が広く利 用されている.しかし,リポフスチン解析による年 齢推定は1個体の年齢を直接査定できるものではな く,また対象種は加入時期が1年の中で明確に分け
られる種に限定される(Bluhm & Brey, 2001).さら に,水温をはじめとする環境要因によってリポフス チンの蓄積量は変化することが知られている (Wahle et al., 1996).このため,甲殻類の新たな年齢形質が 長年にわたって探索されてきた. このような中で,最近,胃小骨の一部である胃咀 嚼器(gastric mill)あるいは眼柄(eyestalk)が甲殻 類の年齢形質として報告された(Leland et al., 2011; Kilada et al., 2012, 2016; Kilada & Ibrahim, 2016).こ れは胃咀嚼器あるいは眼柄の切断面を観察した際, 内クチクラ層に年齢を示すバンドを確認できるとい うものである.なお,甲殻類各種の胃咀嚼器の形態 の比較および機能の詳細については田崎(1995)に 詳しい.この手法を用いた年齢査定が,ズワイガニ Chionoecetes opilio,ホンホッコクアカエビPandalus borealis (ともにKilada et al., 2012),タイワンガザミ Portunus pelagicus (Kilada & Ibrahim, 2016),ナンキョ クオキアミEuphausia superba(Kraft et al., 2016)な どで行われている.しかし,これまで広く用いられ てきたリポフスチンによる年齢推定結果との比較を 行なった事例は報告されていない.
オ ウ ギ ガ ニLeptodius exaratus (H. Milne Edwards, 1834) は岩礁やサンゴ礁に生息する十脚目,短尾下 目,オウギガニ科に属するカニ類であり,わが国にお いては房総半島から八重山列島に,さらにはインド・ 西太平洋にかけて広く分布している (三宅,1983). 本種は東京海洋大学水圏科学フィールド教育研究セ ンター館山ステーション(千葉県館山市)地先の潮 間帯にも多く生息しており,繁殖期などの生活史特 性および成長に関する研究事例がいくつか報告され ている.特に渡邊ら(2012)は館山湾で得られた本 種を用いてリポフスチン解析による年齢推定を行 い,その結果本種は雌が4年級群,雄が5年級群に 分けられることを報告した.そこで本研究では,館 山湾にて採集したオウギガニを用いて胃咀嚼器断面 1 東京海洋大学海洋生物資源学部門 〒108–8477 東京都港区港南4–5–7
Department of Marine Biosciences, Tokyo University of Marine Science and Technology, 4–5–7 Konan, Minato, Tokyo 108–8477, Japan
* 現所属 (Present address):(株)サンケイ分析センター 〒170–0013 東京都豊島区東池袋2–23–2
Sankei Analysis Center, 2–23–2 Higashi-Ikebukuro, Toshima, Tokyo 170–0013, Japan
観察による年齢査定を行ない,先行研究であるリポ フスチン解析による年齢推定結果との比較・考察を 試みた. 材料と方法 採 集 2016年7月20日~22日にかけて,東京海洋大学水 圏科学フィールド教育研究センター館山ステーショ ン地先の潮間帯にて,オウギガニの雌19個体と雄 29個体を採集した. 採集個体は研究室に持ち帰り, 腹節幅から雌雄を判別した後,ノギスを用いて甲幅 を0.05 mmの精度で測定した.測定後の個体は冷凍 保存した. 年齢査定 本研究で用いた年齢査定方法は以下のとおりであ る.なお,基本的な操作は増田ら(2003)による魚 類の耳石処理の手法を参考にした. 解剖 オウギガニを解凍したのち,腹節との接合 部の両端から眼窩にかけて背甲を切断・除去し,胃 を取り出した.取り出した胃は蒸留水中で解剖して 胃壁,胃内容物,筋肉組織などの不要な部分を取り 除き,胃咀嚼器(胃小骨のうち尾噴門骨 (urocardiac), 中央噴門骨 (mesocardiac), 翼噴門骨 (pterocardiac), 対 噴門骨 (zygocardiac) を含む部分)をエタノール・グ リセリン液 (70%エタノール,26%蒸留水,4%グリ セリン;以下,保存液と記載する)に液浸した.得 られた胃咀嚼器は顕微鏡 (SMZ800, Nikon社製) で観 察し,その観察像をデジタルカメラ (DS-Fi1, Nikon社 製) で撮影,保存し,画像解析ソフト (NIS-Elements, Nikon社製)を用いて翼噴門骨長および尾/中央噴門 骨長を測定した(図1). 樹脂包埋 バンドの観察には胃咀嚼器の対噴門骨 を用いた.なお主に左側の対噴門骨を使用し,左側 が欠損等していた場合は右側を使用した.保存液か ら取り出した対噴門骨は室温で約1時間乾燥させ, エポキシ樹脂に包埋した.包埋にはエポキシ樹脂お よび硬化剤 (Epofix, いずれもStruers社製) を重量比 17 : 2で混合したものを使用した.混合した樹脂は シリコーン離型剤(KF412SP, 信越化学工業)を吹 き付けたテフロン製の樹脂包埋用モールド(Teflon
mounting moulds ϕ20 mm, PRESI社製)に流し込み, 乾燥させた試料を,ピンセットを用いて樹脂中に包 埋した.エポキシ樹脂に包埋した試料は直ちに恒温 器に入れ,30℃で12時間静置して樹脂を硬化させ, 樹脂包埋用モールドから取り出した. 樹脂研磨および観察 包埋した樹脂は電動の刃物 研磨機(モデル9820,マキタ製作所製)を用いて研 磨し,対噴門骨の横断面を露出させた.なお,荒削 りには耐水ペーパー(粒度320番)および粒度60番 の砥石を,仕上げには粒度1000番の砥石を用いた. 研磨を終えた試料は,樹脂表面の凹凸による乱反射 を抑えるため保存液中に液浸し,そのまま検鏡に供 した.観察にはNikon社製SMZ800実体顕微鏡を使 用し,内クチクラ層のバンドを計数した.また,観 察像はNikon社製DS-Fi1デジタルカメラによって撮 影,記録した. 相対成長解析 甲幅と翼噴門骨長および尾/中央噴門骨長の関係 を相対成長式(1)によって解析した.相対成長式の aは初期成長定数,bは相対成長係数とよばれ,MS-Excelの回帰分析で推定したbが1より有意に大きい 場合は優成長,小さい場合は劣成長,有意差がない 場合は等成長とした.また,相対成長式の雌雄差を, 統計解析ソフトRのlm関数を用いた共分散分析に よって検定した. y=axb (1) 図1. 腹側から観察したオウギガニの胃咀嚼器と 各部名称および測定部位(横両矢印が翼噴 門骨長,縦の両矢印が尾/中央噴門骨長).
成長様式 得られた年齢情報から雌雄別に3つの成長式(von Bertalanffy式,Gompertz式,Logistic式)を求めた. 成長式のパラメータ推定は統計解析ソフトRのnls 関数を用いて行なった.各式は赤嶺 (2016)に従い, 以下の (2)~(4)式に示すとおりである.推定するパ ラメータの持つ意味は各式によって異なるが,tと l∞は共通して年齢および極限体長を表す.またvon Bertalanffy式におけるcは,理論上の体長が0 mmの ときの年齢を表している.なお採集した個体の年齢 は,オウギガニの抱卵個体が6月から確認されるこ
と (Than Than Lwin, 2008)から,観察されたバンド 数に0.2を足したものと仮定して計算した. von Bertalanffy式 l(t)=l∞*(1-exp(-k(t-c))) (2) Gompertz式 l(t)=l∞*exp(-e-k(t-c)) (3) Logistic式 l(t)=l∞/(1+exp(-k(t-c))) (4) 結 果 相対成長解析 採集したオウギガニの甲幅の頻度分布を図2に示 す.甲幅の平均値±標準偏差は,雌で17.6±3.4 mm, 雄で20.5±7.3 mmであった.翼噴門骨長と尾/中央 噴門骨長は甲幅にともない大型化し (図3),それぞ れの甲幅に対する相対成長は,雌では等成長(それ ぞれp=0.09, p=0.8),雄では劣成長(それぞれp= 0.002, p=0.01) を示した.次に,雌雄で比較すると, 甲幅と翼噴門骨長の相対成長式では,切片,傾きと もに雌雄差がみられ(それぞれp=0.02, p<0.001), 雌の切片が小さく傾きが大きかった.甲幅と尾/中 央噴門骨長の相対成長式では,雌の切片は雄より大 図2. 採集したオウギガニの体長組成ヒストグラム. 図3. オウギガニの甲幅と翼噴門骨長および尾/中央噴門骨長の関係.
きく有意差がみられたが (p<0.001),傾きでは有意 差はみられなかった(p=0.1). 年齢査定 内クチクラ層のバンドは雄1個体を除く47個体 (雌19個体,雄28個体)で観察された(図4).バン ド数は,雌では0~3本,雄では0~5本であり,甲 幅はバンド数にともない大きくなる傾向がみられた (表1).なお,対噴門骨断面におけるバンドの観察 部位は個体によって異なるため,バンド間の長さを 測定することはできなかった.また,バンドが確認 できなかった1個体は,採集時に外骨格が柔らか く,脱皮直後の個体であった. 成長様式 成長式(2)~(4)のパラメータ推定結果を表2に 示す.雄では3つのパラメータを推定できたが,雌 では値が収束せず,パラメータを推定することがで きなかった.このため,本研究ではcに雄の値を代 入することによりl∞およびkの値を推定し,参考値 とした.得られた結果を成長式に代入し,各成長曲 線を図5に示す.実測した年級群の範囲でみると, 雌雄とも3つの式で同様の成長曲線を示し,2年級 群以降は雄が雌の成長を上回っていた. 考 察 胃咀嚼器の長さである翼噴門骨長と尾/中央噴門 骨長は雌雄とも成長にともない大型化したが,雌で は両サイズともに等成長を示したのに対し,雄では どちらも劣成長であった.したがって,雌では体の 成長と胃咀嚼器の成長率が同じであるが,雄では 図4. オウギガニの対噴門骨断面観察像(雄個体, 甲幅26.5 mm).黒矢印は外–内クチクラ層 境界,白矢印は内クチクラ層に形成された バンドを示し,バンドは0から計数した. 表2. オウギガニの雌雄別に推定した各成長式のパラメータ. 成長式 l∞ k c 雌 von Bertalanffy 19.7999 0.7887 -0.8872 Gompertz 39.0140 0.1613 0.9225 Logistic 31.2754 0.2954 1.4833 雄 von Bertalanffy 43.8652 0.1897 -0.8872 Gompertz 33.8519 0.4629 0.9225 Logistic 30.8592 0.7393 1.4833 雌はすべてのパラメータを推定できなかったため,cに雄で推定した値を代入してその他の値を推定した. 表1. オウギガニの胃咀嚼器の対噴門骨断面に観察されたバンド数と体サイズの関係. バンド数 雌(n=19) 雄(n=28) 個体数 平均甲幅 (mm) 標準偏差 個体数 平均甲幅 (mm) 標準偏差 0 2 12.08 2.79 4 8.21 1.29 1 2 17.75 2.12 5 14.45 3.43 2 6 16.01 2.05 5 18.78 1.97 3 9 19.89 2.37 4 24.88 3.06 4 0 ― ― 9 26.86 2.63 5 0 ― ― 1 30.45 ―
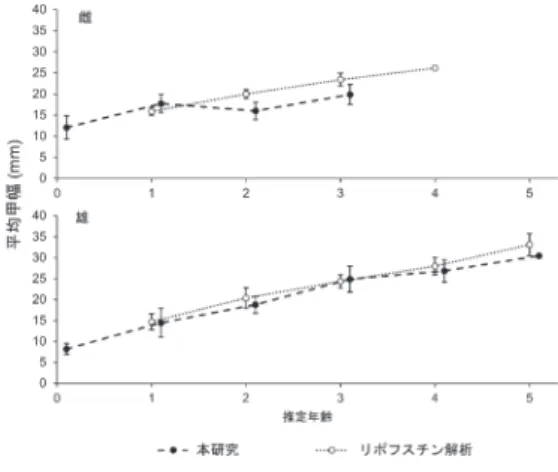
徐々に胃咀嚼器の成長率が鈍くなっていくといえ る.このことは,雌は雄に比べて生殖腺の発達に大 きなエネルギーを投資するため,雄よりも多くの餌 を摂取する必要があるのかもしれない. 本研究では,胃咀嚼器断面観察による年齢査定法 によって,オウギガニの雌では最大3本,雄では最大 5本のバンドが確認された.また,脱皮直後の個体 では胃咀嚼器にバンドが確認されなかった.Vatcher et al. (2015) は胃咀嚼器が外骨格と同様に脱皮するこ とを明確な写真を用いて示したうえで,胃咀嚼器は 年齢形質として不適切であると結論づけている.一 方でLeland et al. (2015) は,カルセイン染色を施した 個体を長期間飼育した結果,胃咀嚼器の染色された 面の外側に新たな層が形成されることを示し,胃咀 嚼器は脱皮によって失われないと報告していること から,脱皮後の再石灰化の過程でバンドが確認でき るようになることも考えられる.胃咀嚼器における バンドの形成メカニズムについては全く解明されて いないことから,今後の検討が望まれる. 雌雄それぞれの成長式にvon Bertalanffy式,Gompertz 式,Logistic式を適用したところ,いずれの成長式 においても2歳以降では雄が雌の成長を上回り,大 型化することが示された.このことは,Than Than Lwin(2008),渡邊ら(2012)で報告された体長組 成の頻度分布解析によるオウギガニの成長様式と一 致していた. ここで,渡邊ら (2012) によるオウギガニのリポフ スチン解析結果(雌99個体,甲幅12.4~28.3 mm; 雄104個体,甲幅11.8~36.7 mm) から,各年級群の 平均甲幅(標準偏差)を算出し,本研究の結果と併 せて図6に示す.なお,オウギガニは生まれた年の 12月には甲幅が約12 mmの大きさまで成長すること が報告されているため(Than Than Lwin, 2008),リ ポフスチン解析による年齢推定結果の最小群は本研 究における1歳群に相当するものとした.この結果 から,雌では数値に多少の乖離がみられるものの, リポフスチン解析法と胃咀嚼器断面観察法とによる 年齢推定・査定結果はほとんど同じ傾向を示すこと が明らかになった.このことは,胃咀嚼器に見られ るバンドが年齢を表していることを支持するもので ある.また,この2手法による年齢情報の比較から, オウギガニの雄は0~5歳の6年級に,雌は0~4歳 の5年級群に分けられる.渡邊ら(2012)は寿命に 性差が生じることについて,繁殖行動にともなうエ ネルギーの消費によって雌が短命となると考察して おり,本研究で得られた結果は前述の甲幅と胃咀嚼 器長の相対成長における性差と合わせて,この説を 支持するものと考えられる. 以上,本研究によって,胃咀嚼器断面観察による オウギガニの年齢査定の可能性が示唆された.今後 は個体数を増やして本研究の結果を補強するととも に,加齢にともなうバンド数の増加について飼育実 験等によって確認する必要がある. 図5. 推 定 し たvon Bertalanffy式,Gompertz式, Logistic式によるオウギガニの成長曲線. 図6. オウギガニの胃咀嚼器による年齢査定法とリポフスチン解析による年齢推定法におけ る各年級群の平均甲幅の比較.エラーバー は標準偏差を示す.
引用文献
赤嶺達郎,2016. 水産総合研究センター叢書 水産資
源解析のエッセンス.第4章水産資源解析のエッ
センス.恒星社厚生閣,東京,pp. 46–59.
Bluhm, B. A., & Brey, T., 2001. Age determination in the Antarctic shrimp Notocrangon antarcticus (Pfeffer, 1887) (Crustacea: Decapoda) using the autofluorescent pigment lipofuscin. Marine Biology, 138: 247–257. Kilada R., Sainte-Marie, B., Rochette, R., Davis, N., Vanier,
C., & Campana, S., 2012. Direct determination of age in shrimps, crabs, and lobsters. Canadian Journal of Fisheries and Aquatic Sciences, 69: 1728–1733. Kilada, R., & Ibrahim, N. K., 2016. Preliminary
investiga-tion of direct age determinainvestiga-tion using band counts in the gastric mill of the blue swimmer crab (Portunus pe-lagicus Linnaeus, 1758) in two salt-water lakes in the eastern Mediterranean. Journal of Crustacean Biology, 36(2): 119–128.
Kraft, B. A., Kvalsund, M., Søvik, G., Farestveit, E., & Ag-nalt, Ann-L., 2016. Detection of growth zones in the eyestalk of the Antarctic krill Euphausia superba (Dana, 1852) (Euphausiacea). Journal of Crustacean Biology, 36(3): 267–273.
Leland, J. C., Coughran, J., & Bucher, D. J., 2011. A prelimi-nary investigation into the potential value of gastric mills for ageing crustaceans. In: Akira Asakura (ed.), New Frontiers in Crustacean Biology: Proceeding of the TCS Summer Meeting, Tokyo, 20–24 September
2009. BRILL, Leiden, pp. 57–68.
Leland, J. C., Bucher, D. J., & Coughran, J., 2015. Direct age determination of a subtropical freshwater crayfish (redclaw, Cherax quadricarinatus) using ossicular growth marks. PLoS ONE, 10(8): e0134966. Doi: 10.1371/journal.pone.0134966. 増田育司・野呂忠秀,2003. 耳石横断薄層切片を用い た魚類の年齢査定の薦め.鹿児島大学水産学部紀 要,52: 51–56. 三宅貞祥.1983. 原色日本大型甲殻類図鑑(II).保育 社,大阪,277 pp. 田崎健郎.1995. 甲殻類の口胃神経系III その比較生 理学.比較生理生化学,12(4): 398–415.
Than Than Lwin, 2008. Population biology of the xanthid crab Leptodius exaratus (H. Milne-Edwards, 1834) in Tateyama Bay, Japan. PhD Thesis, Graduate school of Marine science and Technology, Tokyo university of Marine science and Technology.
Vatcher, H. E., Roer, R. D., & Dilaman, R. M., 2015. Struc-ture, molting, and mineralization of the dorsal ossicle complex in the gastric mill of the blue crab, Callinectes sapidus. Journal of Morphology, 276: 1358–1367. Wahle R. A., Tully, O., & O’Donovan, V., 1996. Lipofuscin
as an indicator of age in crustaceans: analysis of the pigment in the American lobster Homarus americanus. Marine Ecology Progress Series, 138: 117–123. 渡邊隆司・浜崎活幸・横田賢史・Strüssmann C. A.・渡
邊精一,2012. Lipofuscinを年齢形質としたオウギ ガニの年級群推定.Cancer, 21: 7–12.