チトクロム酸化酵素の配位子結合構造に基づく反応機構
兵庫県立大学大学院生命理学研究科 村本和優 チトクロム酸化酵素(CcO)は原核生物から真核生物まで普遍的に保持され、呼吸鎖電子 伝達系の末端酵素(複合体IV)として細胞におけるエネルギー生産機能を担っている。呼 吸鎖電子伝達系は真核生物ではミトコンドリア内膜中に存在し、複合体I, II, III, IV と呼ば れる4 つの膜タンパク質超分子複合体によって構成される(図 1)。電子はミトコンドリア マトリックスのクエン酸回路で生成されたNADH またはコハク酸の酸化によって供与され、 電子伝達系の酸化還元中心の還元電位の勾配に従ってNADH→複合体 I→III→IV→O2、また はコハク酸→複合体 II→III→IV→O2→の順に最終の電子受容体である酸素(O2)まで伝達 される。電子伝達に伴い放出されるエネルギーは膜を介したプロトンの電気化学ポテンシ ャルエネルギーに変換され、このエネルギーは複合体V による ATP 合成、膜タンパク質に よる物質輸送、細胞運動など様々な生命活動に利用される。CcO は O2の還元反応を触媒す るとともに、それと共役したプロトン能動輸送を行うことによってプロトンの電気化学ポ テンシャル差を形成している。CcO における O2還元反応とプロトン電気化学ポテンシャル 差形成のエネルギー変換効率は70%に達すると推定され、高効率なエネルギー変換機構の 物理化学的解明は長年にわたって生体エネルギー学分野における中心的な研究課題である。 (4)H+ (2)H+ NADH NAD+ + H+ チトクロム c H2O 2e -2e -2e- 2e -2H+ 2H+ UQ UQH2 ADP + Pi ATP (3)H+ 複合体 I 複合体III チトクロム bc1 複合体IV チトクロム酸化酵素 複合体V ATP 合成酵素 ミトコンドリア 2H+ ½O2 膜間腔 マトリックス 4H+ 図1 ミトコンドリア呼吸鎖の模式図。電子伝達経路は青、プロトン移動方向は赤で表し ている。複合体I と III 間の電子伝達は膜内在性の低分子化合物ユビキノン(UQ)が、複合 体III と IV 間の電子伝達は膜表在性のタンパク質チトクロム c が担っている。我々の研究グループでは、ウシ心筋ミトコンドリアのCcO を研究対象に用いて、X 線結 晶構造解析を主な方法として反応機構の解明を目指している。図2A は X 線構造解析によっ て決定されたウシCcO のペプチド α 炭素骨格構造である(1, 2)。ウシ CcO は 13 種類の異 なるサブユニット(ポリペプチド)で構成される。それらが集合して機能単位である分子 量約210 kDa、28 本の膜貫通 α ヘリックスを持つ超分子複合体を形成し、それがさらに 2 量体を形成している。サブユニットI, II, III は機能上必須の膜貫通ドメインで、構造的な保
存性も高い。サブユニットI は補因子としてヘム分子(heme a と heme a3)と銅原子(CuB)
を結合し(図2B)、サブユニット II は銅原子(CuA)を結合している。Heme a の鉄原子(Fea)
は軸配位子としてHis61 と His378 が結合した 6 配位構造をとっている。Heme a3の鉄原子
(Fea3)にはHis376 が配位し、Fea3から5 Å 程度離れたところにある CuBにはHis90, His91,
His240 が配位している。Heme a3の第6 配位座と CuBの第4 配位座は外部配位子を結合する
性質を持ち、O2の還元反応の触媒部位として機能する。O2を還元するための電子は還元型
チトクロム c から CuA、heme a, heme a3, CuBの順に伝達される。Heme a3-CuBサイトが還元
されるとO2の結合が可能になり、heme a3-CuBサイトからO2へ電子が移動することにより O-O 結合の解裂が起こる。生成した酸素種はプロトンと反応することにより最終的に 2 分 子の水(H2O)となる。O2をH2O に還元する反応には 4 電子と 4 プロトンが必要である。 膜間腔 マトリックス ミトコンド リア内膜 Tyr244 CuB heme a heme a3 His240 His290 His291 His376 His61 His378 O2+ 4e–+ 4H+ 2H 2O A B 図2 (A)ウシ心筋チトクロム酸化酵素 2 量体の全体構造。サブユニット I は黄色、サ ブユニットII は水色で表している。サブユニット I 内部にあるヘム分子は赤で表している。
(B)O2還元部位付近の構造。Heme a と heme a3 は赤、CuBは水色で表している。アミノ
CcO における O2の4 電子還元反応では、中間生成物としてスーパーオキシドやペルオキ
シドといった反応性が高く細胞に危害を与える酸素種を遊離させることなく、反応を進行
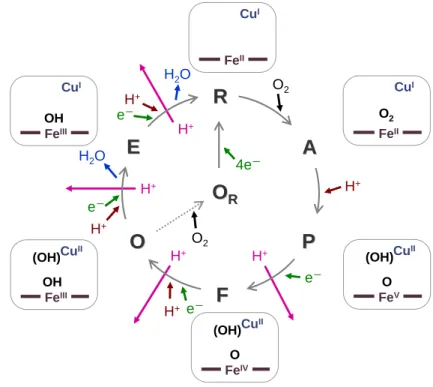
させなければならない。これまでの分光学的研究から、CcO の O2還元反応サイクルにおけ
る中間体が同定されている(図3)。還元型(R)の heme a3-CuBサイトは外部配位子を結合
しておらず、Fea3は2 価 5 配位、CuBは1 価 3 配位の状態にある。O2がFea32+に結合すると
酸素化型( A )が生成する。酸素化型の次に観測される中間体は P 型と呼ばれ、P 型では O2 のO-O 結合は既に切れた状態にあることが時間分解共鳴ラマン分光法により示されている (3)。つまり、O2が結合した後、O2の4 電子還元反応は一気に進行する。4 つの電子のうち 3 つは Fea3とCuBから供与され、4 つ目の電子は heme a3またはHis240 側鎖と共有結合した Tyr244 側鎖から供与されると考えられる。P 型が 1 電子還元されると F 型、更に 1 電子還元 されると酸化型(O)になる。酸化型が 1 電子還元されると E 型、更に 1 電子還元されると 還元型に戻る。2 分子の H2O を生成するために必要な 4 つのプロトンは反応過程の各ステ ップにおいてheme a3-CuBサイトに取り込まれると考えられる。また、生成したH2O は酸化 型から還元型への還元過程でCcO から放出されると考えられる。H2O の生成とは別に、能 動輸送されるプロトンはP 型から還元型への反応過程の各ステップにおいて、電子移動と 共役してCcO から放出されると考えられる。これらの反応中間体以外に、単離精製された 後の酵素は休止酸化型(OR)と呼ばれる状態で存在することが知られている。 H+ H+ H+ H+ H2O e- H+
F
F
e- FeIV O CuII (OH)A
A
O2 FeII O2 CuIP
P
H+ FeV O CuII (OH)E
E
H2O e- H+ FeIII OH CuIO
O
H+ e- FeIII OH CuII (OH)R
R
O
O
RR 4e- FeII CuI O2図3 CcO の反応サイクルモデル。Heme a3-CuBサイトへの電子の移動を緑色、プロトンの
移動を茶色で表している。プロトン能動輸送(プロトンポンプ)を赤で表している。各反
応中間体において存在が推測されるCuB配位子の酸素種を括弧で表している。休止酸化型
CcO の反応機構を解明するためには、各反応中間体構造や反応過程における構造変化の 詳細な情報が必要である。我々はこれまでの研究において、CcO 結晶の反応中間体を作製 し、加えてheme a3-CuBサイトに結合する呼吸阻害剤をプローブとして用い、X 線構造解析 によりO2還元反応とプロトンポンプ機構に関与する構造情報を得てきた(4, 5)。以下に heme a3-CuBサイトの配位子結合構造に基づいて得られたO2還元反応機構に関する知見を紹介す る。
休止酸化型構造(図4A) CcO を単離、精製し X 線構造解析を行った結果、Fea33+とCuB2+
の間を架橋するperoxide の電子密度が観測された。酸化型の 1 電子還元によってプロトン
ポンプが誘起されるのに対し、休止酸化型では1 電子還元と共役したプロトンポンプ機能
が認められない。よって、heme a3-CuBサイトへのperoxide の結合はプロトンポンプ機能を
阻害することが示唆される(6, 7)。
還元型構造(図4B) 休止酸化型 CcO 結晶に還元剤を作用させると Fea32+, CuB1+の還元
型が生成する。Fea3は5 配位構造で、ポルフィリン面から 0.3 Å 程度 His376 側へ出た位置
にある。CuBは3 配位構造で、配位子である His 側鎖の 3 つの窒素が作る平面内に位置して
いる(6, 8)。
CuB1+–CO 結合構造(図 4C) 還元型 CcO と一酸化炭素(CO)を反応させると CO は Fea32+
に結合する。Fea32+–CO 結合型 CcO に低温状態で光を照射すると、CO が Fea3から解離して
CuB1+–CO 結合構造をとることが分かった。CO は CuBに対してside-on 型の結合をしており、
この構造はO2が還元型Fea3に結合する過程で一時的にCuBに結合する状態に対応すると考 えられる。CuBは還元型の構造と比べてCO 寄りに位置する。 Fea32+ C- H2O N Tyr N Fea33+ N O --O Fea32+ CuB1+ N His His His Tyr O C Fea32+ CuB1+ O N His His His Tyr N Fea32+ CuB1+ His His His Tyr N CuB2+ His His His Tyr CuB1+ His His His A B C D E
図4 Heme a3-CuBサイトの配位子結合構造。(A)休止酸化型 (B)還元型 (C)CO
結合還元型(光解離後) (D)NO 結合還元型 (E)CN–結合還元型 いずれの構造も温
Fea32+–NO 結合構造(図 4D) 還元型 CcO と亜硝酸イオン(NO2–)を反応させるとFea32+
にNO が結合する。解析の結果、NO は Fea3に対しbent end-on 型の結合をしており、N-O
軸はCuBの第4 配位座の方向を向いていることが分かった。この構造は Fea32+–O2のモデル と考えられる。NO が結合した Fea3はポルフィリン面内に位置する。CuBは配位子が作る面 近くに位置することから、NO と CuBとの相互作用は弱いと考えられる。Fea32+–O2中間体の Fe–O2伸縮振動の共鳴ラマンスペクトルがヘモグロビンやミオグロビンのそれらと酷似し ていることから、O2とCuBとの相互作用は弱いと推定されていた(3)。この X 線構造解析結 果は、共鳴ラマン分光解析結果を導く構造要因を示すものと考えられる。 Fea32+ CN– CuB1+結合構造(図4E) 陰イオンである CN–は一般に酸化型のheme(Fe3+)
に強く結合するが、CcO においては CN–は還元型のheme a3-CuBサイトにも結合する。解析
の結果、CN–がFea32+に配位するとheme a3のポルフィリン面が僅かにスライドし、Tyr244
とCN–を水素結合で架橋する位置に水分子が取り込まれることが分かった。この結果は、
Fea32+に結合したO2(Fe2+–O2 ↔ Fe3+–O2–)も同様に水分子を取り込むことを示唆している。
この水分子が配置されることはFea3に結合しているO2への4 電子還元の引き金になると考
えられる。
以上の知見をもとに、我々は次のO2還元機構を提唱している(図5)。O2は還元型heme
a3-CuBサイトのCuB1+に一時的に結合した後、Fea3にbent end-on 型の配位をする。CuBとの
相互作用は弱く、(Fea32+–O2 ↔ Fea33+–O2–)構造は安定化される。O2とTyr244 の間に H2O
が取り込まれて水素結合が形成されると、それが引き金となってO2の4 電子還元が起きる。
Tyr244 は K-pathway と呼ばれるプロトン輸送経路の末端に位置しており、O2の還元に伴い
プロトンがK-pathway から heme a3-CuBサイトに取り込まれてP 型中間体が形成される。
H+
Transition state
Fea32+ CuB 1+ O His His N His Tyr HO H O O H2O K -pathw ay Fea3 2+ CuB1+ O O His His N His Tyr HO H OA
A state
K-p a th w a y Fea3 2+ CuB 1+ O O N His His His Tyr HO H OR
R state
+
O
2 K-p a th w a y Fea35+ CuB 2+ His His N His Tyr HO H O OH -OP
P state
H2O K -pathw ay H2O 図5 O2還元反応過程(還元型からP 型中間体まで)のスキーム。謝辞 本研究は兵庫県立大学大学院生命理学研究科の吉川信也教授の研究室において、伊藤・ 新澤恭子助教、前田友子研究員、小倉尚志教授(兵庫県立大学)、月原冨武教授(兵庫県立 大学)、山下栄樹助教(大阪大学)、青山浩准教授(大阪大学)らとの共同研究で行われた ものです。この場をお借りして深く感謝いたします。 文献
1. Tsukihara T, Aoyama H, Yamashita E, Tomizaki T, Yamaguchi H, Shinzawa-Itoh K, Nakashima R, Yaono R, Yoshikawa S. (1995) Structures of metal sites of oxidized bovine heart cytochrome c oxidase at 2.8 A. Science 269, 1069-74.
2. Tsukihara, T., Aoyama, H., Yamashita, E., Tomizaki, T., Yamaguchi, H., Shinzawa-Itoh, K., Nakashima, R., Yaono, R. and Yoshikawa, S. (1996) The whole structure of the 13-subunit oxidized cytochrome c oxidase at 2.8 A. Science 272, 1136-1144.
3. Kitagawa T., Ogura T. (1997) Oxygen activation mechanism at the binuclear site of heme-copper oxidase superfamily as revealed by time-resolved resonance Raman spectroscopy. Prog. Inorg. Chem. 45, 431-479.
4. Yoshikawa S, Muramoto K, Shinzawa-Itoh K, Aoyama H, Tsukihara T, Ogura T, Shimokata K, Katayama Y, Shimada H. (2006) Reaction mechanism of bovine heart cytochrome c oxidase. Biochim Biophys Acta. 1757, 395-400.
5. Yoshikawa S, Muramoto K, Shinzawa-Itoh K, Aoyama H, Tsukihara T, Shimokata K, Katayama Y, Shimada H. (2006) Proton pumping mechanism of bovine heart cytochrome c oxidase. Biochim Biophys Acta. 1757, 1110-1116.
6. Yoshikawa S, Shinzawa-Itoh K, Nakashima R, Yaono R, Yamashita E, Inoue N, Yao M, Fei MJ, Libeu CP, Mizushima T, Yamaguchi H, Tomizaki T, Tsukihara T. (1998) Redox-coupled crystal structural changes in bovine heart cytochrome c oxidase. Science 280, 1723-1729.
7. Aoyama H, Muramoto K, Shinzawa-Itoh K, Hirata K, Yamashita E, Tsukihara T, Ogura T, Yoshikawa S. (2009) A peroxide bridge between Fe and Cu ions in the O2 reduction site of fully
oxidized cytochrome c oxidase could suppress the proton pump. Proc. Natl. Acad. Sci. USA (in press)
8. Tsukihara T, Shimokata K, Katayama Y, Shimada H, Muramoto K, Aoyama H, Mochizuki M, Shinzawa-Itoh K, Yamashita E, Yao M, Ishimura Y, Yoshikawa S. (2003) The low-spin heme of cytochrome c oxidase as the driving element of the proton-pumping process. Proc. Natl. Acad. Sci. USA 100, 15304-15309.