循環系の基礎と臨床
(5)リンパ管発生
東京女子医科大学医学部解剖学・発生生物学講座 エ ザ キ タ イ チ江
太一
(受理 平成 29 年 7 月 26 日)Circulatory System: Basic and Clinical Research (5) Development of Lymphatic Vessels
Taichi EZAKI
Department of Anatomy and Developmental Biology, School of Medicine, Tokyo Women s Medical University
Research on the lymphatic vascular system has made relatively slow progress compared to that of the blood vascular system. The major reasons for this include a lack of reliable techniques for the identification of lym-phatic vessels in tissue, and that blood circulation is considered to be directly connected with life, and therefore requires urgent investigation of associated pathogenesis and treatments. However, in recent decades, research on the lymphatic system has made remarkably rapid progress because of the discovery of specific markers for lymphatic endothelial cells, various factors and genes relating to their proliferation and regeneration, and also sig-nificant advancements in lymphatic vessel imaging techniques. We have entered a new era of lymphatic system studies in both basic and clinical research fields. The importance of lymphatic vessels in local microcirculation is being realized through improved understanding of the functional significance of the lymphatic system in the nor-mal body fluid circulation, and its contribution to defense against various infections and cancers. The establish-ment of treatestablish-ments for lymphatic diseases relating to edema (lymphedema), immune disorders including organ transplantation, and lymphogenic metastasis of malignant tumors is a matter of great urgency. In the present view, I present the phylogeny, ontogeny, and mechanisms for lymphangiogenesis/lymphvasculogenesis, and re-port some imre-portant disorders relating to lymphangiogenesis, and discuss promising recently published strate-gies for their treatment.
Key Words: microcirculation, lymphatic endothelial cell, lymphangiogenesis, lymphvasculogenesis, lymphogenic metastasis
はじめに
日本におけるリンパ管研究は杉田玄白の『解体新
書』
(1774 年)に始まるが,その後約 100 年の空白時
代を経て,その本格的な進歩は我が国の脈管学の祖
である足立文太郎とその門下である木原卓三郎
1),さ
らにその弟子らによって,一大発展を遂げた
2).はじ
め,血液は組織に開放されていた(開放性血管)が,
動物の構造が複雑になるにつれて,組織液を回収し
て心臓に返すための管(静脈)ができ,血管系が閉
鎖する(閉鎖性血管).さらにからだが複雑化・大型
:江 太一 〒162―8666 東京都新宿区河田町 8―1 東京女子医科大学医学部解剖学・発生生物学講座 Email: [email protected] doi: 10.24488/jtwmu.87.5_1352
Fig. 1 Generalized schematic of body fluid circulation
Long solid arrows show blood flow in the blood vascular system. Dotted arrows show lymph flow in the lymphatic system. Smaller arrows show bidirectional exchanges of body fluid via the extravascular fluid pathway in the interstitial spaces.
化すると組織液の回収が静脈だけでは不十分とな
り,組織液の回収を目的とした新たな管(吸収管),
つまりリンパ管が分化する.したがって,リンパ管
系は血管系を補うために発生するのであり,系統発
生学的にも個体発生学的にも,血管系より遅れて分
化する(Fig. 1).われわれ哺乳類においては,リンパ
管は血管と同様に循環系を構成する重要な脈管系の
1 つであるが,臨床的にも心・血管系に比べると
“命”に直結する緊急性が低いことから,昔からより
軽視される傾向にあった.つまり,リンパ管研究の
歴史は血管生物学のめざましい発展に比べて,つい
15 年ほど前までは大きく遅れをとっていた.ところ
が,近年リンパ管の内皮の特異的なマーカーやリン
パ管内皮の増殖を促進する種々の因子の発見によっ
て,基礎医学・臨床医学の領域を問わず,今まさに
新たな時代を迎えている
3)∼5).
本稿では,これまでの長いリンパ管研究の歴史を
垣間見ながら,リンパ管の系統発生,ならびに個体
発生,さらにはリンパ管新生・形成機構とリンパ管
新生と関連する種々の病態とその対策などを最近の
トピックスを交えて考察してみたい.
リンパ管の発生
1.リンパ管の系統発生
そもそも,循環系は植物性器官
6)のうち,すべての
生物の営む生命現象の根幹である“食と性”の観点
から吸収系(すなわち消化管系)と排出系(すなわ
ち泌尿生殖器系)の間に介在し,生体にとって必須
の栄養と生殖の両機能を果たすために両者をつなぐ
体液の流路として出現する.生命の誕生から多細胞
生物としての発生過程の中で,太古の海水の形象が
体液,つまり「血液」となって,次第にそれを包む
管としての「血管」の中に閉じ込められる過程が起
こる(Fig. 2A).特にわれわれ脊椎動物では,それま
で“開放循環”であったものから体液をより効率的
に循環させるために体液からの「血液の分離」が系
統発生の途上で,つまり古生代末の上陸とともに,
急速に進行したと考えられる.脊椎動物の血液と血
管が誕生する最初の姿としては,卵黄囊の壁の周囲
の間葉細胞の細胞集団(血島 blood island)の中から,
hemoangioblast という血液細胞と血管内皮の両者
の幹となる細胞から分化するといわれる.
その際,循環系には個体の栄養機能と並行して個
体の防禦機能が同時に備わってくる.つまり,個体
の中では生命維持のために一種の“交易”と“警備”
のために,入出物の授受や侵入物の処理は,当初は
もっぱら流動系の「血液」によって果たされる.そ
の中心となる場が,
“栄養の吸収”と“生体防御”の
要である原始腸管とその隣接器官である(Fig. 2B).
やがて,その細胞は古い骨髄系と新しいリンパ系の
2 種類に分かれ,それぞれ独自の場で生産が営まれ
るが,その造血組織の中で,後者のリンパ組織の分
化と並行して組織間の余剰の体液を導く,新たな還
Fig. 2 Origin of vascular systems and relationship between the myelopoietic and
lympho-poietic systems in association with the primitive gut
A: The vascular system is formed as a network in the inter-dermal areas (*), where body fluid can flow freely. Blood vessels form vascular networks in these areas. B: Dis-tribution of myelopoietic system (dotted) and lymphopoietic system (slashed) against for-eign objects ingested into the primitive gut. The former consists of primary myelopoietic organs (e.g. blood island, liver) on the ventral side of the primitive gut and secondary myelopoietic organs (e.g. spleen) on the dorsal side. The latter consists of primary lym-phopoietic organs (primitive thymus at the entrance of the primitive gut, bursa of Fabri-cius in the cloaca at the end) and secondary lymphopoietic organs (e.g. spleen, Peyer s patches, lymph nodes) along the intestine.
All illustrations are reproduced and modified with permission from the original figures by Miki (1989)6).
流路すなわち「リンパ管系」が静脈路とは独立して
組織間隙に出現する.
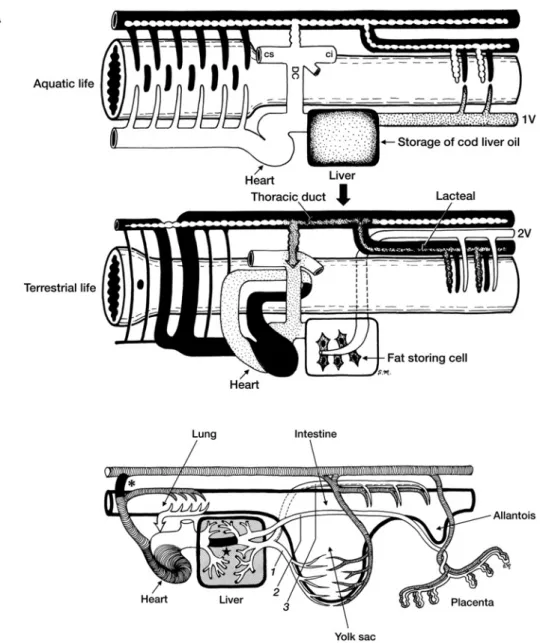
さらに,この両者の違いとして栄養の面から「脂
質」の吸収とその貯蔵方法の違いを招いた.そこで,
リンパ系と脂肪代謝の観点から,肝臓という臓器の
意義を考えてみる(Fig. 3).われわれ哺乳類では,三
大栄養素のうち,脂肪(厳密にいうと炭素数が C12
以上の長鎖脂肪)の吸収は,腸の毛細血管からは吸
収できず,毛細リンパ管(中心乳び管 lacteal)から
のみ吸収されて,門脈系―肝臓を介さずに,別のルー
トで腸管リンパ系―胸管を介して直接静脈角に合流
する.三木
6)によれば,もともと肝臓は消化管上皮か
ら発生して,消化機能と吸収機能の両面を持ってい
て,消化管腔内で「管腔内消化」を果たしていたと
いう.ところが,次第に消化管を離れ,門脈という
特殊な専属血管系を伴って消化管で吸収された栄養
物を一挙に「管腔外消化」するとともに,それらの
栄養源を貯蔵するようになった.特に無脊椎動物ま
では,肝臓では糖質・タンパク質・脂質の三大栄養
素の代謝と貯蔵を行っている.ところが,この中で
脂肪については,脊椎動物が水棲から陸棲に移行す
るにつれて,次第に肝臓から遠のいていく.つまり,
水棲時代の肝臓は現在も魚類の“肝油”として知ら
れるように,腸管より吸収された脂肪流のすべてを
門脈経由で受取り,これを重要な栄養源として自ら
の中に貯蔵する.ところが,陸上生活と共に陸棲動
物の脂肪流は,その経路を門脈からリンパ管に大き
く変更して,次第に胸管に合流して直接血液内に合
4
Fig. 3 Changes in fat flow across liver phylogeny and sources of the hepatic portal vein
during ontogenesis
A-top: In aquatic organisms, fat (dotted) as well carbohydrates and proteins are trans-ported via the portal vein (1V) to the liver. The fat is stored in the liver as “cod liver oil”.A-bottom: In terrestrial organisms, fat (dotted) is only absorbed by the lacteals and transported via the thoracic duct to the general blood circulation without first accumu-lating in the liver. The portal vein (2V) only transports carbohydrates and proteins to the liver. In the human liver, fat storing cells known as Ito s fat storing cells are only located in the space of Disse. B: During ontogenesis, the mammalian hepatic portal vein chrono-logically originates from the yolk sac (1), placenta (2), and, after birth, the digestive tract (3). The ductus venosus ( ★ ) and ductus arteriosus (*) are represented.
All illustrations are reproduced and modified with permission from the original figures by Miki (1989)6).
流する.以来,脂肪は動物性器官のうち,体壁系(皮
膚・骨格筋)の特に皮下組織から筋組織の内部に貯
蔵され,ついには血管壁にまで及ぶことに成り兼ね
なくなった.こうして,陸棲動物の肝臓は,水棲時
代から持ち続けていた脂肪の貯蔵能の大半を失い,
辛うじてその名残として,肝 Disse 腔内にいる脂肪
摂取細胞(伊東細胞)の姿を留めるに過ぎない.
小谷
7)によれば,独立したリンパ管系が初めて確認
できるのが閉鎖血管系が確立した硬骨魚類からだと
いう.無脊椎動物・甲殻類では,開放血管系を持つ
ためリンパ管系は存在しない.また,ヤツメウナギ
に代表される円孔類では,腸管からの栄養を吸収し,
Fig. 4 Phylogeny of the lymphatic system and lymph hearts in various species
A: The thoracic duct (T) enveloping the aorta (Ao) of a snake. A valve equipped with smooth muscle fibers is indicated by an arrow. B: The lymph heart of a lizard. A valve with smooth muscle fibers (arrow) can be seen at the entrance of the lymph heart filled with Berline blue. C: The lymph hearts of salamanders ( ① ), frogs ( ② ), snakes ( ③ ), and fowls ( ④ ). Reproduced with permission from Kotani (1990)10).
腸粘膜ヒダの中の(哺乳類の中心リンパ管にあたる)
脈管中を流れるので,血乳び管ともいうべきもので,
独立したリンパ管とはいえない
8).サメなど一部の軟
骨魚類には,血液を含まない乳び管を持つものがい
るともいわれる
9)が,それは例外的らしい.
リンパ管の形状も動物によって著しい差がある.
イモリやサンショウウオなど有尾両生類のリンパ管
はよく発達しているが,硬骨魚類と同様に細い.一
方,カエルなど無尾両生類やカメ・ヘビなどの爬虫
類のリンパ管は非常によく発達し,太いリンパ洞あ
るいは広いリンパ囊を形成する.しかも,その太い
リンパ洞の中を動静脈が走っている.ヘビでは甲状
腺も胸腺も,脂肪体,さらに肝臓すら広いリンパ腔
に包まれているという
10)(Fig. 4A).このような広い
リンパ洞やリンパ囊を持つ謎を解く鍵として,オー
ストラリアの砂漠に住むカエルが夏の暑さを避けて
夏眠する際に皮下のリンパ囊が水分の補給と断熱の
効果があるといわれている.さらには,ガラパゴス
島のイグアナやナイル川のワニは変温動物(冷血動
物)であるがゆえに,自分で熱産生ができないため
に太陽からの熱を熱伝導の低いリンパ液を満たした
リンパ洞で溜め込むことでその中を通過する血管内
の血液を保温し体温維持をしているといわれる.つ
まり,体温をより安定に保つための断熱材や保温材
の役割を果たすのではないだろうか.鳥類になると,
リンパ管は急に細くなり,主な血管系は独立して走
るが,その全周または一部をリンパ管に包まれた血
管も多い.哺乳動物のリンパ管も細く,血管系とリ
ンパ管系は完全に独立している.血管系は末梢部と
中枢部では太さに大きな差があるが,リンパ管は極
端な差がない.したがって,毛細リンパ管といって
も毛細血管の数倍∼それ以上の内径を持つことがあ
6
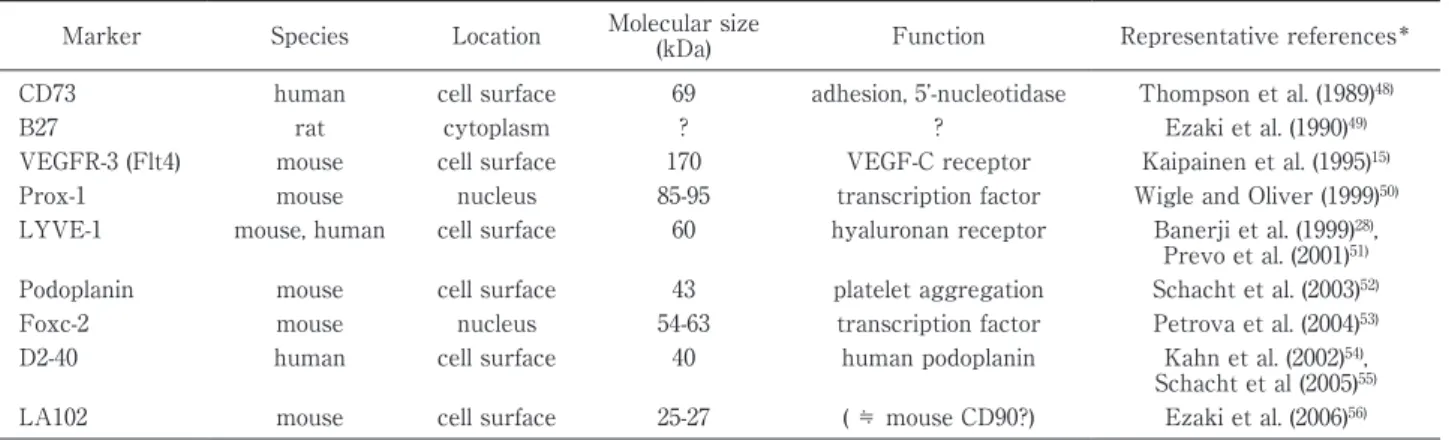
Table 1 Specific markers for lymphatic endothelial cells
Marker Species Location Molecular size
(kDa) Function Representative references* CD73 human cell surface 69 adhesion, 5 -nucleotidase Thompson et al. (1989)48)
B27 rat cytoplasm ? ? Ezaki et al. (1990)49)
VEGFR-3 (Flt4) mouse cell surface 170 VEGF-C receptor Kaipainen et al. (1995)15) Prox-1 mouse nucleus 85-95 transcription factor Wigle and Oliver (1999)50) LYVE-1 mouse, human cell surface 60 hyaluronan receptor Banerji et al. (1999)28),
Prevo et al. (2001)51) Podoplanin mouse cell surface 43 platelet aggregation Schacht et al. (2003)52) Foxc-2 mouse nucleus 54-63 transcription factor Petrova et al. (2004)53) D2-40 human cell surface 40 human podoplanin Kahn et al. (2002)54),
Schacht et al (2005)55) LA102 mouse cell surface 25-27 ( ≒ mouse CD90?) Ezaki et al. (2006)56) *Only original or representative papers are listed.
る.ちなみに,毛細リンパ管とは,
「明瞭な基底膜や
弁構造を欠き単層!平な内皮細胞のみで構成される
最末端側のリンパ管」であり,その起始部は盲端に
始まる.血管と同様に基底膜や弁構造,さらに筋層
を有するのは次の集合リンパ管のレベルからである
が,これらのリンパ管が互いに合流して中枢側に近
付くとしても,内径の大きさは極端には差がない.
リンパ管と血管の交通やリンパ流の動きについて
も動物によって著しい差がある
10).両生類や爬虫類
にはリンパ心臓といわれるポンプ機能を持つ装置が
存在する(Fig. 4B,C).サンショウウオなどの有尾
両生類には各体節ごとに 1 対,カエルなどの無尾両
生類では前後各 1 対のリンパ心臓に減少し,ヘビや
カメなどの爬虫類ではさらに減少して後リンパ心臓
1 対のみとなる.鳥類では胚時期に後リンパ心臓が
一時的に出現するものの,孵化後は見られないとい
う
11).ヒトを含めた哺乳類ではリンパ心臓はなく,弁
を持つ集合リンパ管の走行において,リンパ輸送の
機能単位と考えられているリンパ管分節が,リンパ
心臓に相当するものと考えられている.ちなみに,
両生類や爬虫類では広いリンパ洞のように拡張した
リンパ管系を有するために,リンパの流れは特定方
向ではなく,多方向に向かうため貯留することがな
く,浮腫は起こらない.このようなリンパ管の系統
発生の過程で,リンパ心臓が減少することにより,
ヒトにおいては逆にリンパ浮腫が発症しやすくなる
傾向があるというのは極めて興味深い
12).
2.リンパ管の個体発生
リンパ管の個体発生については古来から論争が尽
きない.リンパ管が静脈内皮の発芽によって末梢側
に向かって伸張していくとする「遠心説 centrifugal
theory」
13),静脈周囲の間葉性組織間隙の細胞が!平
化して小腔を作り,形成された腔と腔が互いに連結
して原始的なリンパ囊となり,さらに繋がってリン
パ管となり中枢側に向かうとする「求心説
centripe-tal theory」
14)である.その際のリンパ管と静脈との
連結は二次的に生じるとされた.その他,両者の「折
中説」として,胸管は胎生静脈から出芽し,他は組
織間隙から発生して中枢側に向かうとするもの,あ
るいは頸リンパ囊は静脈性で,他は組織間隙からと
するものなどがある.
リンパ管発生機序について 100 年以上も論争が続
いてきた根本的な理由は,血管とリンパ管を厳密に
同定する方法がなかったことである.血管内皮の特
異的マーカーがいち早く発見されて以来,血管生物
学は飛躍的に進歩したのに比べ,リンパ管内皮の特
異的マーカーの発見は 20 年近くも後になったこと
から,リンパ管研究は大きく遅れをとってしまった.
つまり,1995 年に血管内皮増殖因子受容体
3(vascu-lar endothelial growth factor receptor-3 : VEGFR
3)がリンパ管内皮に特異的に発現することが発表
15)されて以来,ようやく本格的に種々の分子マーカー
が報告されてきた(Table 1).このようなリンパ管特
異的なマーカーによってリンパ管を厳格に同定する
ことによって初めて,リンパ管と血管との相関性が
証明できるようになった.したがって,近年では分
子生物学的手法を用いて遠心説を支持する報告
16)が
多くなっている.しかしその一方,ゼブラフィッシュ
を用いたライブイメージングによる解析から,求心
説を支持する所見や,リンパ管内皮の由来とその分
化を制御するメカニズムが頭頸部と体幹部では異な
るとする説も報告
17)され,新たな波紋を呼んでいる.
リンパ管の個体発生を考える上で,胎生期のリン
パ管形成について遠心説の立場から考えると,静脈
Adhesion molecules Integrin α5 Integrin β5 ICAM-1, ICAM-2 N-cadherin Selectin P, Selectin E CD44 Integrin α9 Integrin α1
Macrophage mannose receptor I
Cytoskeletal proteins Vinculin
Claudin 7 Actin α2 Profilin 2 Desmoplakin I and II Adducin γ Plakoglobin
α-actinin-2 associated LIM protein Extracellular matrix proteins Collagens 8A1, 6A1, 1A2
Lamininγ2, α5 Versican Proteoglycan1
Matrix Gla protein
Extracellular matrix modulation MMP-1, MMP-14 uPA
Plasminogen activator inhibitor I Cathepsin C
TIMP-3
Cytokines, chemokines & receptors IL-8, IL-6
Monocyte chemoattractant protein 1 UFO/axl CXCR4 CCRL2/CKRX IL-4 receptor IL-7 SDF-1b
Growth factors VEGF-C
Tie2 Angiopoietin 2VEGFR-3
# The data listed in this table are reproduced and modified from the original reports by Petrova et al (2002)26) & Kobo (2004)44).
から発芽する「リンパ管発生(lymphvasculogene-sis)」と,既存のリンパ管から新たにリンパ管が伸張
する「リンパ管新生(lymphangiogenesis)」を区別し
て考えることができる.Buttler ら
18)は胎生期は静脈
から発芽するリンパ管発生と同時に末梢の間葉由来
のマクロファージ様細胞が取り込まれながらリンパ
管を形成することを報告している.また,Kato とそ
の同僚ら
19)は酵素組織化学的手法によって,種々の
臓器の間質組織に,しばしば原始的なリンパ囊様の
構造(リンパ島 lymphatic island)が出現し,さらに
そこから小突起が伸びて互いに融合・連結して次第
に有弁性のリンパ管網が形成される一連の過程を報
告
20)している.
いずれにしても,成体におけるリンパ管形成は,
既存のリンパ管から新たにリンパ管が伸張するリン
パ管新生が主体となると考えられ,例えば血管に認
められる脈管形成 vasculogenesis に相当するよう
な,末梢血中でのリンパ管内皮前駆細胞の存在など
については今のところよく解っていない.
3.リンパ管新生・形成の分子機構
まず,血管の発生と新生の起こるメカニズムにつ
いて,分子レベルで解明が進み始めたのは,血管内
皮 増 殖 因 子(vascular endothelial growth factor:
VEGF)とその受容体群が発見されてからである.リ
ンパ管新生研究の幕開けとなったのも,VEGF 受容
体ファミリーの 1 つである VEGFR3 の発見
21)と,こ
れがリンパ管内皮細胞に特異的に発現すること
15),
ならびに VEGFR3 のリガンドとしての VEGF-C が
リンパ管新生因子であること
22)が報告されたことで
あった.さらに,血管内皮細胞との関係で対比され
る分子として,neuropilin-2 の KO マウスで皮下にリ
ン パ 管 が 欠 損 し て 浮 腫 を 起 こ す こ と
23),ま た
angiopoietin-2 の KO マウスでは腸管リンパ管の形
成不全と壁細胞欠損で生直後に乳び腹水で致死とな
ることが報告
24)されている.このように,リンパ管の
発生や新生の分子機構にも血管との相同性,関連性
の深さが常に付きまとい,2 つの脈管系が互いに孤
立無援ではあり得ないことを示している
25).また一
方では,両者の内皮細胞株同士の遺伝子解析から,
それらの遺伝子プロフィールには多くの相違点も明
らかになった
26)(Table 2).
さらに,遺伝子レベル,あるいは種々の転写因子
レベルでリンパ管の初期発生やリンパ管新生に関与
するものが存在することが,これまで数多く報告さ
れている(Fig. 5).例えば,遠心説の立場からでは,
リンパ管の初期発生は,まず静脈内皮細胞における
8
Fig. 5 The process of lymphatic endothelial cell differentiation from venous endothelial
cells, and related target genes expressed at each step in mice
At the embryonic stage of 8.5 days (E8.5), venous differentiation occurs with coinciding high expression of VEGFR2. At E9.5 ∼ E12.5 some venous endothelial cells become posi-tive for LYVE-1 and Sox18, and acquire the ability to differentiate into lymphatic vessels. The expression of Sox18 induces the expression of Prox1, a master control gene in the process of lymphatic endothelial cell differentiation. The expression of Prox1 induces its target genes, such as VEGFR3 (a receptor for VEGF-C), and podoplanin, which increases cell mobility. Prox1 also temporally suppresses VE-cadherin, enabling free migration, so that the cells can detach from each other and move towards VEGF-C to form a stable lymphatic vessel.
核 内 受 容 体 で も あ る chicken ovalbumin upstream
promoter transcription factor II(COUP-TF II)とい
う転写因子が VEGF の下流で Notch シグナルを抑
制することで動脈分化から静脈の分化に転じるとさ
れている
27).リンパ管発生において最初の段階に観
察されるのは,静脈内皮細胞においてヒアルロン酸
受容体(lymphatic vessel endothelial hyaluronan
re-ceptor 1:LYVE-1)を発現すること
28)であり,これに
よってリンパ管内皮細胞に分化する能力を獲得す
る,つまりリンパ管内皮前駆細胞が出現すると考え
られている.ここで,Sry-related HMG box family
転写因子である Sox18 が発現するようになると,既
に静脈内皮で発現していた COUP-TF II と共役し
て prospero related homeobox 1(Prox-1)遺伝子発現
を誘導する
29).この Prox-1 遺伝子はショウジョウバ
エのホメオボックス遺伝子のオーソログであり,レ
ンズ,網膜,膵臓,肝臓などにも発現するが,脈管
ではリンパ管のみに発現しており,リンパ管分化の
マスター転写因子である
30)31).つまり,この転写因子
が 発 現 す る
と,VEGFR3,FGFR-3,LYVE-1,podo-planin をはじめとする種々のリンパ管内皮特異的な
マーカーの発現が上昇する
32).Prox1 陽性細胞は静
脈から出芽して周囲の VEGF-C 発現領域に遊走し
て初期のリンパ囊が形成されるが,さらに,リンパ
管 内 皮 細 胞 が 出 芽 に よ っ て 伸 張 す る た め に は
VEGF-C が必須である
33).
VEGF-C で活性化された幼若なリンパ管内皮は,
さらに種々の分子シグナルによってリンパ管特有の
走行あるいは弁形成や平滑筋による被覆を伴う成熟
の過程を取り始める
34).まず,リンパ管形成の成熟期
には forkhead box protein C2(Foxc2),nuclear
fac-tor of activated T-cells cytoplasmic 1(NFATc1),
GATA binding protein 2(GATA-2)などの転写因子
が関与する.Lymphedema distichiasis(LD)症候群
いること
35),GATA-2 KO マウスにおいても前 2 者と
同様にリンパ管の弁形成不全を来すことが報告
36)さ
れている.さらに最近では,その他にも多種多様な
分 子,例 え ば connexin,reelin,elastin microfibril
interface 1(EMILIN-1),semaphorin 3A-NRP1 など
がリンパ管の成熟過程に関与することが報告され,
これらの分子機能異常によりリンパ浮腫の原因とな
ることも分かっている
5).
4.病態時のリンパ管新生とその制御の試み
創傷治癒はもとより,身体内の様々な病的炎症の
場において,血管新生と共にリンパ管新生が観察さ
れる.このような局所の微小環境において,NFκB
によるプロモーターの活性化を介して炎症細胞から
VEGF-C の産生が起こること,Prox1 と協調してリ
ンパ管内皮細胞の VEGFR3 発現を高めることなど
が報告されている.また,脂質メディエーターが炎
症巣や腫瘍性癌病巣の間質細胞,マクロファージ,
顆粒球などに作用して VEGF-C/D を誘導してリン
パ管の新生増強作用を発揮していることが分かって
いる
37).さらに,腎臓や角膜などの移植部位において
も,リンパ管内皮細胞由来のケモカイン CCL21 が移
植片由来の CCR7 陽性樹状細胞を呼び寄せ,移植片
の拒絶反応の引き金を引くといわれているが,この
ような場では,リンパ管新生を抑制することの有効
性が種々の移植実験で試行されている
38).
また,臨床的にも特に問題になるのが,腫瘍の増
殖に伴うリンパ管新生とリンパ節転移の問題であ
る.悪性腫瘍の遠隔転移には血行性とリンパ行性が
あり,癌による死亡の決定的要因の 1 つである.従
来から癌腫はリンパ行性に,肉腫は血行性に転移し
やすいと言われてきたが,その理由は十分に証明さ
れていない
39).腫瘍のリンパ節転移については,最近
まで腫瘍が増大する過程で既存のリンパ管に偶発的
に侵入し,リンパ節へ到達するという考えが主流で
あった.また,腫瘍組織内にはリンパ管は存在しな
いと考えられていた
40).ところが,腫瘍が VEGF-C
41)や VEGF-D
42)を発現・分泌することでリンパ管新生
を誘導し,リンパ行性に転移するケースが報告され
ている.さらに,VEGF-C の発現とリンパ節転移と
の相関が胃,大腸,肺,前立腺,甲状腺,メラノー
マなどの腫瘍例でも報告されており,腫瘍内および
腫瘍周囲に新生するリンパ管は「腫瘍リンパ管」と
おわりに
リンパ管研究は今や,基礎医学と臨床医学の両面
から,正に新しい時代を迎えている
4).それを確実に
しているのが,リンパ管のイメージング技術の飛躍
的な進歩である
45)46)といっても過言ではない.そこに
は,依 然 と し て 未 解 決 の 謎 や 問 題 点 も 多 い も の
の
39)47),リンパ管の発生過程の足取りとそのメカニズ
ムを理解することによって,今後ともリンパ管の機
能とその生理学的意義,免疫学的意義,腫瘍学的意
義が少しずつでも自ずと理解されよう.そして,我々
の体内において,血管系を上水道に例えるならば,
リンパ管系はまさに下水道に当たる.忘れてならな
いのは,ライフラインとして命と直結している血管
系に対して,リンパ管系は緊急性はないものの,こ
れら下水道が詰まって上手く流れなくなると,とた
んに生命活動が成り立たなくなってしまうことであ
る.正に,
「病気が発症するのも,治癒するのも,す
べてリンパ管系を含む局所の微小循環系から始ま
る」といっても過言ではない.この言葉とともに,
本稿が今後の基礎医学と臨床医学の発展に少しでも
役に立てば幸いである.
謝 辞 本稿をまとめるに当たって,長年の間たくさんの方々 のご協力・ご支援を受けた.特に,恩師の小谷正彦熊本 大学名誉教授, 出崎順三博士(元愛媛大学医学部講師), ならびに当教室の森川俊一講師,清水一彦助教,故山崎 康子氏,中田和子氏,佐川弘美氏,元丸佳恵氏に心より 感謝いたします. 開示すべき利益相反はない. 文 献1)Kihara T: Anatomie der Japaner 3; Lfg. 2. Das tiefe Lymphgefäßsystem der Japaner. Deutsch-Japanisches Kulturinstitut, Kyoto (1953)
2)小谷正彦:付録リンパ管研究の歴史.「リンパ管 形態・機能・発生」(大谷 修,加藤征治,内野滋雄 編),pp311―320,西村書店,新潟(1997) 3)江 太一:リンパ管をいかにして把握するか? リ ンパ学 35(2):105―107,2012 4)大橋俊夫,河合佳子:循環生理学・免疫学・腫瘍学 と連携した新しいリンパ学の提唱.リンパ学 38 (1):15―18,2015
5)Aspelund A, Robciuc MR, Karaman S et al: Lym-phatic system in cardiovascular medicine. Circ Res
10 118: 515―530, 2016 6)解剖学論集.「生命形態の自然誌 第 1 巻」(三木成 夫編),うぶすな書院,東京(1989) 7)小谷正彦:6 リンパ管の分化と発生.「リンパ管 形態・機能・発生」(大谷 修,加藤征治,内野滋雄 編),pp255―259,西村書店,新潟(1997) 8)木原卓三郎:琳巴管系ノ分化ニ就イテ.解剖誌 14:9―15,1939
9)Kampmeier OF: Evolution and Comparative Mor-phology of the Lymphatic System, Thomas, Spring-field, Ill (1969)
10)Kotani M: The lymphatics and lymphoreticular tis-sues in relation to the action of sex hormones. Arch Histol Cytol 53 (Suppl): 1―76, 1990
11)Kutsuna M: Beitrage zur Kenntinis der Entwick-lung des Lymphagefassystems der Vorgel. Acta Scholae Med Univ Kyoto 16: 6―12, 1933
12)「新しいリンパ学 微小循環・免疫・腫瘍とリンパ
系」(加藤征治,須網博夫編),金芳堂,京都(2015)
13)Sabin FR: On the origin of the lymphatic system from the veins and the development of the lymph hearts and thoracic duct in the pig. Am J Anat 1: 367―389, 1902
14)Huntington GC, McClure CFW: The anatomy and development of the jugular lymph sacs in the do-mestic cat(Felis dodo-mestica). Am J Anat 10: 177―311, 1910
15)Kaipainen A, Korhonen J, Mustonen T et al: Ex-pression of the fms-like tyrosine kinase 4 gene be-comes restricted to lymphatic endothelium during development. Proc Natl Acad Sci USA 92 (6): 3566― 3570, 1995
16)Oliver G: Lymphatic vasculature development. Nat Rev Immunol 4: 35―45, 2004
17)磯貝純夫:リンパ管をめぐる新知見 リンパ管内 皮への分化制御.炎症と免疫 24(5):367―371, 2016
18)Buttler K, Ezaki T, Wilting J: Proliferating meso-dermal cells in murine embryos exhibiting macro-phage and lymendothelial characteristics. BMC Dev Biol 22 (8): 43―55, 2008
19)Kato S, Miura M, Miyauchi R: Structural organi-zation of the initial lymphatics in the monkey mes-entery and intestinal wall as revealed by an enzyme-histochemical method. Arch Histol Cytol
56: 149―160, 1993
20)三浦真弘,米村 豊:リンパ管形成とリンパの流れ リンパ管新生と生後成長様式の形態学的特徴.リン パ学 37:10―15,2014
21)Aprelikova O, Pajusola K, Partanen J et al: FLT 4, a novel class III receptor tyrosine kinase in chro-mosome 5q33-qter. Cancer Res 52 (3): 746―748, 1992 22)Jeltsch M, Kaipainen A, Joukov V et al:
Hyperpla-sia of lymphatic vessels in VEGF-C transgenic mice. Science 276 (5317): 1423―1425, 1997
23)Yuan L, Moyon D, Pardanaud L et al: Abnormal lymphatic vessel development in neuropillin 2 mu-tant mice. Development 129 (20): 4797―4806, 2002 24)Gale NW, Thurston G, Hackett SF et al:
Angiopoietin-2 is required for postnatal angiogene-sis and lymphatic patterning, and only the latter
role is rescued by Angiopoietin-1. Dev Cell 3: 411― 423, 2002
25)Adams RH, Alitalo K: Molecular regulation of an-giogenesis and lymphanan-giogenesis. Nat Rev Mol Cell Biol 8: 464―478, 2007
26)Petrova TV, Mäkinen T, Mäkelä TP et al: Lym-phatic endothelial reprogramming of vascular en-dothelial cells by the Prox-1 homeobox transcrip-tion factor. EMBO J 21 (17): 4593―4599, 2002 27)You LR, Lin FJ, Lee CT et al: Suppression of
Notch signalling by the COUP-TFII transcription factor regulates vein identity. Nature 435 ( 7038 ) : 98―104, 2005
28)Banerji S, Ni J, Wang SX et al: LYVE-1, a new homologue of the CD44 glycoprotein, is a lymph-specific receptor for hyaluronan. J Cell Biol 144 (4): 789―801, 1999
29)吉松康裕:リンパ管をめぐる新知見 リンパ管形 成における転写因子の役割.炎症と免疫 24(5): 372―378,2016
30)Oliver G, Sosa-Pineda B, Geisendorf S et al: Prox 1, a prospero-related homeobox gene expressed during mouse development. Mech Dev 44 (1): 3―16, 1993
31)Hong YK, Harvey N, Noh YH et al: Prox 1 is a master control gene in the program specifying lym-phatic endothelial cell fate. Dev Dun 225: 351―357, 2002
32)Wigle JT, Harvey N, Detmar M et al: An essential role for Prox1 in the induction of the lymphatic en-dothelial cell phenotype. EMBO J 21 : 1505 ― 1513, 2002
33)Karkkainen MJ, Haiko P, Sainio K et al: Vascular endothelial growth factor C is required for sprout-ing of the first lymphatic vessels from embryonic veins. Nat Immunol 5: 74―80, 2004
34)平島正則:リンパ管をめぐる新知見 リンパ管パ ターニングにかかわる分子シグナル.炎症と免疫
24(5):379―384,2016
35)Norrmén C, Ivanov KI, Cheng J et al: FOXC2 con-trols formation and maturation of lymphatic collect-ing vessels through cooperation with NFATc 1. J Cell Biol 185 (3): 439―457, 2009
36)Kazenwadel J, Betterman KL, Chong CE et al: GATA2 is required for lymphatic vessel valve de-velopment and maintenance. J Clin Invest 125 (8): 2979―2994, 2015
37)馬嶋正隆,天野英樹,細野加奈子:リンパ管をめぐ る新知見 プロスタノイドによるリンパ管新生と がんリンパ節転移の制御.炎症と免疫 24(5): 391―400,2016
38)Alitalo K : The lymphatic vasculature in disease. Nat Med 17 (11): 1371―1380, 2011
39)小 谷 正 彦:リ ン パ 管 の 7 不 思 議.リ ン パ 学 35 (2):66―80,2012
40)Padera TP, Kadambi A, di Tomaso E et al: Lym-phatic metastasis in the absence of functional intra-tumor lymphatics. Science 296 (5574 ): 1883 ― 1886, 2002
41)Skobe M, Hawighorst T, Jackson DG et al: Induc-tion of tumor lymphangiogenesis by VEGF-C
pro-循環系の基礎と臨床―掲載予定― 執筆者 所属 テーマ 掲載号 澤田達男 病理学(第一) 1.脳の微小循環 87(1・2) 槍澤大樹 輸血・細胞プロセシング科 2.血管新生 87(1・2) 森本 聡 高血圧・内分泌内科 3.高血圧 87(3) 瀧田守親 薬理学 4.転移 87(4) 江 太一 解剖学・発生生物学 5.リンパ管発生 87(5) 小川哲也 腎臓内科 6.動脈硬化(透析も含めた腎血管) 87(6)
promotes the metastatic spread of tumor cells via the lymphatics. Nat Med 7 (2): 186―191, 2001 43)Stacker SA, Achen MG, Jussila L et al:
Lymphan-giogenesis and cancer metastasis. Nature Rev 2 : 573―583, 2002
44)久保 肇:リンパ管の形成機構.生化学 76(9): 1210―1216,2004
45)Munn LL, Padera TP: Imaging the lymphatic sys-tem. Microvasc Res 96: 55―63, 2014
46)江 太一:特集 リンパ管イメージングの最前線 総論 リンパ管可視化のあゆみと展望.細胞 47 (13):622―624,2015 47)江 太一:リンパ管形成とリンパの流れ リンパ 学に残された謎をめぐって―リンパと脂肪との関 わり―.リンパ学 37(1):4―9,2014
48)Thompson LF, Ruedi JM, Glass A et al: Antibod-ies to 5 -nucleotidase ( CD 73 ) , a glycosyl-phosphatidylinositol-anchored protein, cause hu-man peripheral blood T cells to proliferate. J Immu-nol 143: 1815―1821, 1989
49)Ezaki T, Matsuno K, Fujii H et al: A new ap-proach for identification of rat lymphatic capillaries using a monoclonal antibody. Arch Histol Cytol 53 (Suppl): 77―86, 1990
50)Wigle JT, Oliver G: Proxl function is required for
LYVE-1 is an endocytic receptor for hyaluronan in lymphatic endothelium. J Bio Chem 276 : 19420 ― 19430, 2001
52)Schacht V, Ramirez MI, Hong YK et al: T1alpha/ podoplanin deficiency disrupts normal lymphatic vasculature formation and causes lymphedema. EMBO J 22: 3546―3556, 2003
53)Petrova TV, Karpanen T, Norrmén C et al: De-fetive valves and abnormal mural cell recruitment underlie lymphatic vascular failure in lymphedema distichiasis. Nat Med 10: 974―981, 2004
54)Kahn HJ, Bailey D, Mark A: Monoclonal antibody D2-40, a new marker of lymphatic endothelium, re-acts with Kaposi s sarcoma and a subset of angio-sarcomas. Mod Pathol 15: 434―440, 2002
55)Schacht V, Dadras SS, Johnson LA et al: Up-regulation of the lymphatic marker podoplanin , a mucin-type transmembrane glycoprotein, in human squamous cell carcinomas and germ cell tumors. Am J Pathol 166: 913―921, 2005
56)Ezaki T, Kuwahara K, Morikawa S et al: Produc-tion of two novel monoclonal antibodies that distin-guish mouse lymphatic and blood vascular endothe-lial cells. Anat Embryol 211: 379―393, 2006