河川基金助成事業
「溶菌性ウイルスに着目したメタボロミクス解析
による琵琶湖難分解性有機物の起源解明」
助成番号:
28 – 5211 – 041
京都大学大学院工学研究科附属
流域圏総合環境質研究センター
教授 清水芳久
平成
28 年度
様式6・2 1.調査・研究 [概要版報告書] 助成番号 助成事業名 所属・助成事業者氏名 28 – 5211 – 041 溶菌性ウイルスに着目した メタボロミクス解析による 琵琶湖難分解性有機物の起源解明 京都大学大学院工学研究科附属 流圏総合環境質研究センター 清水 芳久 助 成 事 業 の 要 旨 〔目 的〕 本研究では、琵琶湖物質循環においてウイルス分流を経由して放出される DOM(Dissolved Organic Matter:溶存有機物)量を推定することを目的として、具体的には以下の課題を追求した。 (1) 琵琶湖北湖と南湖での観測からウイルスおよび細菌の存在量を季節や水深で比較し、その 変動要因を検討する。 (2) 琵琶湖北湖沖帯(水深約 90 m)におけるウイルス感染率についての季節変化を把握する。 〔内 容〕 本研究では、ウイルスとそれに続く溶菌プロセスの琵琶湖物質循環における寄与を推定するた め、毎月の現場観測(7~12 月)から細菌・ウイルス数やウイルスによる細菌の感染率を調査し た。琵琶湖北湖今津沖中央においては、細菌数は表水層で 0.8~3.2 × 106 cells mL-1、深水層で 0.4 ~0.8 × 106 cells mL-1であった。また、ウイルス数は表水層で 2.6~9.8 × 106 mL-1、深水層で 1.3~ 3.7 × 107 mL-1であった。表水層での細菌数とウイルス数は 7~12 月にかけて深水層での値に近づ いた。また、感染率は表水層で 0.9~2.7%、深水層で 1.4~3.0 であった。推定された感染成功率 は深水層(4.8~20.4%)の方が表水層(1.7~11.8%)に比べて高く、細菌数の少ない深水層にお いて感染成功率を上げる生存戦略をウイルスは取っていることが示唆された。また、年間のウイ ルス感染により放出される DOC(Dissolved Organic Carbon)は約 6,900 tonC であり、細菌炭素生 産量の約 20%、一次生産の約 6%であることが分かった。深水層に限れば、ウイルス感染により 放出される DOC 量は、深水層での細菌炭素生産量の約 36%を占めた。以上のことから、ウイル スが琵琶湖物質循環において重要な役割を果たしていることが示された。 〔結 果〕 本研究で推定したウイルス感染により放出される年間 DOC 量は、表水層で 3,770 tonC、深水層 で 3,150 tocC であった。これらは表水層の細菌生産量の 15.8%、深水層の 36.1%である。琵琶湖 全体で考えると、内部生産 107,337 tocC/y のうち 30.4%(32,600 tocC/y)が細菌生産に消費さ れており、細菌生産量うちの 21.2%(6,920 tonC/y)がウイルス感染により DOC に回帰している。 この値は海洋で報告されている値よりも低いが、この理由としては、海洋(主に外洋)に比べて 一次生産が活発である点が挙げられる。また、琵琶湖では内部生産の約 20%が沈降フラックス (5,699 tonC/y)として深水層へ移動し、深水層での重要な有機栄養源となっている。しかし, 本研究で推定されたウイルス感染により放出される DOC 量は深水層で 3,150 tonC/y であり、沈降 フラックスに匹敵する重要な有機栄養源であることが分かる。DON(Dissolved organic nitrogen) も同様にウイルス感染によって放出される。放出される DON は細胞内でほとんどがアミノ酸や DNA など易分解性の有機物に含まれていることから、放出後は細菌によりすぐに利用されると考えら れる。さらに利用された易分解性の細胞由来の有機物は無機化されて窒素やリンなどの栄養塩と して環境水中に放出される。このようにウイルスとその後に続く細胞破壊プロセスは、細菌や藻 類への重要な利用性の高い栄養源として琵琶湖水中で重要な役割を果たしている。 調査対象水系・河川 データベースに登 録するキーワード 部門 大分類 中分類 小分類 調査部門 環境 水環境 その他 ※データベースに登録するキーワードは、実施の手引 P.47 の表から 代表的なものを一つ記入 して下さ
様式6・3 1.調査・研究 [自己評価シート] 助成番号 助成事業名 所属・助成事業者氏名 28 – 5211 – 041 溶菌性ウイルスに着目した メタボロミクス解析による 琵琶湖難分解性有機物の起源解明 京都大学大学院工学研究科附属 流圏総合環境質研究センター 清水 芳久 助 成 事 業 実 施 成 果 の 自 己 評 価 〔計画の妥当性〕 琵琶湖では、1980 年代中頃より CODMnとBOD との乖離が問題となっており、難分解性有 機物が増加・蓄積していると指摘されている。なぜ難分解性有機物が増加し始めたのか、その 解明はされておらず、現在でも、その起源について多くの研究がなされている。藻類や細菌の 種組成や活性の変化による「内部生産由来説」、流入河川中の難分解性有機物の増加による「陸 域由来説」、そして気候変動等によって引き起こされた水・物質循環の変化による「汚濁機構 変化説」の3 つに大別しており、生分解試験およびモデルによる推計から、「内部生産由来説」 を支持している。しかし、日本各地の湖沼および琵琶湖の流入河川でも同様の乖離問題が同時 期に起きていると報告されている。これは、地球温暖化のような外的環境要因の変化が湖沼の 環境に作用しており、日本各地の湖沼が応答を示したことによると考えられる。そのため、物 質循環の変化による汚濁機構変化説を容易に否定するのは危険であり、藻類や細菌の内部生産 も含めた物質循環全体を解明する必要があると考えた。 〔当初目標の達成度〕 物質循環において、ウイルス分流という仮説がある。ウイルスの感染によって宿主細菌の細 胞が破壊・溶菌され、その成分が溶存態有機物(DOM:Dissolved Organic Matter)および粒 子態有機物(POM:Particulate Organic Matter)として水中に放出される。溶菌に伴い放出 された DOM は比較的すぐに細菌に利用され、細菌→DOM→細菌という循環的な流れが形成 され、またCO2として水中から除去される。したがって、ウイルス感染は一次生産の速度にも 深く関わっている。また、易分解性有機物は細菌によって利用され難分解性有機物に変換され、 水中に残存することから、ウイルス感染によって放出されたDOM の一部も難分解性有機物に 変換されていると考えられる。このように、水中バイオマスの90%以上を占める細菌の一部が ウイルス感染によりDOM に変換されることから、ウイルス分流の強さが水中の難分解性有機 物の増減を支配しているのではないかと予想できる。この予想の妥当性を検証するためには、 まず琵琶湖におけるウイルス分流の寄与や、細菌との相互作用を明確にする必要があった。 本研究では、当初、まずメタゲノム解析を用いて細菌と溶菌性ウイルスの存在実態を時空間 的に把握し、次に全有機物を対象に網羅的解析手法を用いて難分解性有機物の起源とその生成 過程を解明することを目指した。しかしながら、予備実験の結果、前者のウイルスの時空間的 把握に多くの時間が必要であったことから、主としてこれに重点を置いて研究を実施した。こ のために、当初購入を予定していたDNA 抽出キットについては実施金額が少なくなっている。 また、安定同位体試薬と次世代シーケンサー分析試薬は、過年度より使用してきたものを使用 することが可能であったため、そのまま利用した。さらに、本研究の内容は研究室に所属する 大学院生の修士論文研究の一部としたため、研究協力者謝金使用は行なわなかった。これらを 消耗品等の購入資金に充てたことにより、より効果的に研究を進めることができたと考える。 〔事業の効果〕 ウイルス分流はウイルスが細菌に感染することで始まる。しかし、感染率、つまり衝突頻度 と感染成功率がどのような因子に支配されているかはよく分かっていない。その要因として、 宿主の多様性やウイルスとの組み合わせ、あるいは細胞への付着に関与する物質や水温などの 物理因子などが考えられるが、解明には至っていない。今後は、感染率の制御因子の探索が必 要になると考えられる。また、実際に琵琶湖水中に存在しているウイルスのうちシアノファー ジとバクテリオファージがどの程度の割合を占めているか、あるいは、それらの不活化率を把 握することは、より正確なウイルスの寄与を評価する上で重要な知見となり得る。 〔河川管理者等との連携状況〕 本研究は、滋賀県環境科学琵琶湖研究センターの協力の下に実ンプリングにおいては、同セ ンターの「はっけん号」に同乗させてもらっての実施であった。
1. はじめに
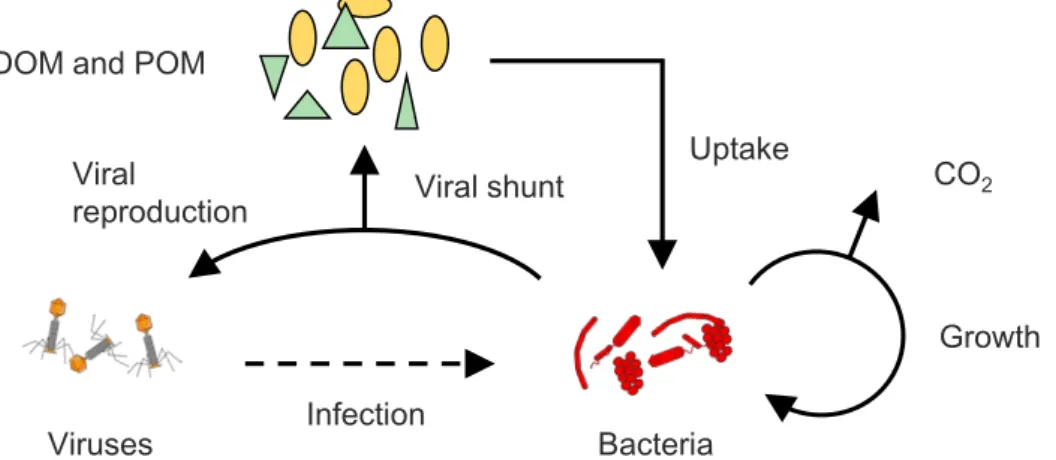
1.1 研究の背景 琵琶湖では,1980 年代中頃より CODMn(化学的酸素要求量)とBOD(生物化学的酸素 要求量)との乖離が問題となっており,生物の分解を受けにくい難分解性有機物が増加・ 蓄積していると指摘されている(岡本ら,2011 および 2012;今井ら,1998).近年では, この乖離問題も落ち着きを見せはじめているものの,なぜ難分解性有機物が増加し始めた のか,依然としてその解明はされていない.問題が起こり始めて 30 年が経過した現在で も,その起源について数多くの研究がなされている.佐藤ら(2016)は,難分解性有機物 の起源について,藻類や細菌の種組成や活性の変化による「内部生産由来説」,流入河川中 の難分解性有機物の増加による「陸域由来説」,そして気候変動等によって引き起こされた 水・物質循環の変化による「汚濁機構変化説」の 3 つに大別しており,生分解試験および ボックスモデルによる推計から,「内部生産由来説」を支持している.しかし,日本各地の 湖沼および琵琶湖の流入河川でも同様の乖離問題が同時期に起きていると報告されている (滋賀県,2014).これは,地球温暖化のような外的環境要因の変化が湖沼の環境に作用し ており,日本各地の湖沼が応答を示したことによると考えられる.そのため,物質循環の 変化による汚濁機構変化説を容易に否定するのは危険であり,藻類や細菌の内部生産も含 めた物質循環全体を解明する必要がある. 物質循環において,ウイルス分流(Viral Shunt)という仮説がある(Figure 1.1).ウイ ルスの感染によって宿主細菌の細胞が破壊・溶菌され,その成分が溶存態有機物(DOM,Dissolved Organic Matter)および粒子態有機物(POM, Particulate Organic Matter)と して水中に放出される.溶菌に伴い放出された DOM は比較的すぐに細菌に利用され,細 菌→DOM→細菌という循環的な流れが形成され,また CO2として水中から除去される. この際,制限元素であるリンや鉄も同時に放出されることから,ウイルス感染は一次生産 の速度にも深く関わっている.また,易分解性有機物は細菌によって利用され難分解性有 機物に変換され,水中に長期間残存することから(Ogawa et al., 2001),ウイルス感染に よって放出された DOM の一部も難分解性有機物に変換されていると考えられる.このよ うに,水中バイオマスの 90%以上を占める細菌(Suttle, 2007)の一部がウイルス感染に Viral shunt Infection Bacteria Viruses Viral reproduction DOM and POM
Uptake
Growth CO2
より DOM に変換されることから,ウイルス分流の強さが水中の難分解性有機物の増減を 支 配 し て い る の で は な い か と 予 想 で き る . 実 際 に , 海 洋 で は , 溶 菌 に よ っ て 放 出 さ れる DOM(Virally mediated DOM)は年間 3-20 Gt C と報告されており(Wilhelm and Suttle, 1999),これは海洋での 1 年間の一次生産量である 50 Gt C に対して大きな割合を占めて いる.この予想の妥当性を検証するためには,まず琵琶湖におけるウイルス分流の寄与や, ウイルスと細菌の相互作用が果たす役割を明確に記述する必要がある. 1.2 研究の目的 本研究では,琵琶湖物質循環においてウイルス分流を経由して放出される DOM 量を推 定することを目的とした.具体的には以下の課題を追求した. (1) 琵琶湖北湖と南湖での観測からウイルスおよび細菌の存在量を季節や水深で比較 し,その変動要因について検討する. (2) 琵琶湖北湖沖帯(全水深約 90 m)におけるウイルス感染率について表層と深層で の季節変化を把握する.
2. 実験材料と方法
2.1 採水地点 採水は滋賀県琵琶湖環境科学研究センター(以下,琵環研セ)が毎月実施する水深別調 査に同乗して実施した.観測期間は 2016 年 7 月~12 月の毎月で,採水地点は琵琶湖北湖 の 4 地点(17A,17B,17C’,12B)と南湖の 1 地点(6B)で実施した(Table 2.1).地点 17B および 12B では水深別に採水を実施した. 2.2 採水方法および前処理Station Location Sampling depth (m) Northern lake Biwa
17A 35° 23’ 58”N, 136° 02’ 30”E 0.5
17B 35° 23’ 41”N, 136° 07’ 57”E 0.5, 5, 10, 15, 20, 30, 40, 60, 80 17C’ 35° 22’ 17”N, 136° 15’ 22”E 0.5
12B 35° 11’ 39”N, 135° 59’ 39”E 0.5, 40 Southern lake Biwa
6B 35° 02’ 40”N, 135° 53’ 36”E 0.5
Table 2.1 Description of stations sampled in the Northern and Southern Lake Biwa from July through December in 2016.
各採水地点の表水層試料はステンレスバケツで採水した.また,17B および 12B の表水 層 以 外 の 水 深 別 試 料 は バ ン ド ー ン 採 水 器 ま た は ペ リ コ ン ポ ン プ ( XX80EL000,Merk-Millipore)を用いて採水した.バンドーン採水器は琵環研セが地点 17B において水深別の 試料を採水する際に使用する採水容器である.化学分析用試料の採水には硼珪酸ガラス製 のねじ口瓶を,生物分析用には滅菌済みボトル(ポリスチレン,CORNING)を用いた. 全ての採水容器は試料で共洗いを 1 度行った.また生物分析用試料は採水後にすぐに船上 で 50%グルタルアルデヒド(最終濃度 1vol%)による化学固定を行い,冷蔵保存(4 °C) により実験室に持ち帰った.その後,分析まで冷凍保存(-30 °C)した. 2.3 現場の観測データおよび水質項目 水温データについては,平成 28 年度公共用水域・地下水水質測定計画(滋賀県)に基づ き,滋賀県琵琶湖環境科学研究センターにより測定されたものである.これらは,本論文 執筆時点では速報値である. 2.4 全菌数,ウイルス数および VBR
全菌数およびウイルス数の計数は Patel et al. (2007)や Noble and Fuhrman (1998)によ る方法をもとに琵琶湖水試料に対して条件を最適化して実施した.以下にこの方法を簡単 に記す.グルタルアルデヒドにより化学固定(最終濃度 1vol%)した試料(0.5~2 mL)を, 孔径 0.8 μm のフィルター(セルロース混合エステル,Merck Millipore) をバッキングフィ ルターとして用いて,Anodisc filter(孔径 0.02 μm,Whatman)上にろ過した.次に自然乾 燥させた Anodisc filter を 400 倍希釈した SYBR Gold で 18 分間染色し,再び自然乾燥させ た.この Anodisc filter を退色防止試薬とともにスライドガラスとカバーガラスの間にマウ ントした.作成したスライガラスは観察日まで-30 °C で冷凍保存した.観察には蛍光顕微 鏡(BZ-9000,KEYENCE)を用いた.また,超純水によるブランクのスライドガラスを作 成し,観察に使用した試薬や実験器具などによるコンタミがないことを毎回確認した.ま た,VBR(Viruses to bacteria ratio)はウイルス数を細菌数で除して求めた.
2.5 透過型電子顕微鏡(TEM)を使用した感染率(FVIC)の評価
感染率およびバーストサイズの評価には,地点 17B の水深 0.5 m と 60 m のサンプルを 使用した.サンプル中の細菌は超遠心(Himac CS 100GXII, Hitachi; S52ST スイングロータ, 70,000 × g,20 分,4 °C,Pradeep Ram et al,. 2010)によって TEM グリッド(400-mesh, carbon-coated Formvar film)に捕集した.1 サンプルに対して TEM グリッドを 3 枚用意し(N = 3),それぞれのグリッドを EM ステイナー(希釈なし,日新 EM)で 30 分間,室温で染色 した.その後,超純水で 5 回洗浄し,自然乾燥させた.このグリッドは透過型電子顕微鏡 H-7650(80 kV,Hitachi)により観察(3,000~50,000 倍)した. 本研究では,1 細胞に 3 つ以上のウイルス粒子が明瞭に確認できるものを VIC(Visibly Infected Cell)とした.化学固定の過程でメソソームと呼ばれる人工産物が細胞内に見られ ることがあり,また細胞分裂中にはそれが 2 つ確認できる.メソソームは形状と色がウイ ルス粒子に似ており,これらが存在する細胞を誤って VIC と判定するのを避けるために, 感染の判定基準を 3 つ以上とした.
グリッド 1 枚につき 500 細胞以上を観察し,FVIC(Frequency of Visibly Infected Cell)を 計算した.TEM でウイルス粒子を宿主細胞の中に確認できる期間は,ウイルスの生活史の 10-20%程度であり,感染した直後や暗黒期にあるウイルスは観察できない.よって FVIC を式(2.1)に従い,FIC(Frequency of Infected Cell)に変換する必要がある(Weinbauer et al., 2002).
256
.
3
524
.
9
−
=
FVIC
FIC
(2.1) また,ウ イル スによ る 感染が細 菌の 死亡率 に 対して占 める 割合( Frequency of bacterial Mortality due to Viral Lysis,FMVL)を式(2.2)に従い,計算することが出来る(Binder, 1999). またバーストサイズは宿主細胞中のウイルス粒子を計数して求めた.FIC
FIC
FIC
FMVL
2
.
1
1
6
.
0
2−
+
=
(2.2) 2.6 細菌の細胞体積 細胞体積の計算には 50%グルタルアルデヒド(最終濃度 1vol%)により固定した細菌を 利用し,その形状は両端が半球の円柱型であると仮定した.細菌は FVIC の評価のために 得た TEM 画像から無作為に 100 細胞を抽出し,長径 L と短径 W から以下の式(2.3)に従 い,体積 V を算出した.この式は桿菌と球菌(L=W)の両方に適応可能である.また,長 径(L)と短径(W)の計測にはオープンソースであるソフトウェア,ImageJ を用いた.
−
=
3
4
2W
L
W
V
π
(2.3) 2.7 ウイルスと細菌の衝突頻度 ウイルスは水中をブラウン運動する溶質であると解釈すると,ウイルスと細菌が衝突す るという現象は,ウイルスの粒子(宿主菌および非宿主菌,あるいは生物粒子や非生物粒 子)への移動であると捉えることが出来る.よってウイルスと細菌の衝突頻度 R(contacts mL-1 d-1)は,以下の式(2.4)により求めることが出来る(Murray and Jackson, 1992).BA
VA
D
S
R
=
(
2
πϖ
V)
⋅
(2.4) 式(2.4)で,S はシャーウッド数(無次元),ω は細菌の長径(cm),DVはウイルスの拡散係数(4.32×10-3 cm2 d-1, Dubin et al., 1970),VA はウイルス数(mL-1),BA は細菌数(cells
mL-1)である.
海洋でのデータではあるが,細菌群集のうち運動性を有するのは 10%未満であることが 報告されている(Mitchell et al., 1995).したがって本研究では運動性を有する細菌を 10%, 非運動性の細菌を 90%と設定した.運動性と非運動性の細菌のシャーウッド数はそれぞれ S = 1.53,1.01 であるので(Murray and Jackson, 1992),それぞれの存在量を考慮したシャ ーウッド数 1.06 を本研究で使用した.また衝突頻度 R を細菌数で除したものを,1 細菌・
1 日あたりの衝突頻度 RBacteria(contacts cell-1 d-1)として計算した.さらに溶菌により死亡 した細菌数を,細菌 1 個あたりの衝突頻度 RBacteriaで除して 100 倍したものを感染の成功 率とした.
3. 結果
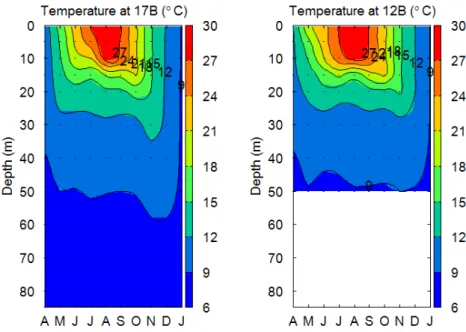
3.1 水温 地点 17B の表水層(0.5 m)では観測期間(2016 年 7 月~12 月)を通して,11.6~28.9 °C の間を推移した(Figure 3.1).また深水層(60 m)では,8.5~8.9 °C と安定していた.地 点 12B の表水層(0.5 m)においては 12.0~29.8 °C,深水層(40 m)では 9.3~10.2 °C の間 を推移していた.また地点 12B,17B ともに 4 月には弱いながらも 40 m 付近に躍層が形成 されており,8 月には 10~15 m の水深で最も強い躍層が形成された.そして 8 月から 12 月にかけて表水層の水温が下がると,躍層は弱まりながら,その深度も深くなっている. 12 月での水温躍層は地点 12B で 30~40 m,地点 17B で 40~60 m の水深で形成されてい た.2017 年 1 月では,地点 12B の全水深で水温が 8.8 °C,17B が 8.6~8.7 °C となってお り,全循環が起きたと考えられた.したがって本章で得られた地点 17B における 7~12 月 のデータ(水深 0.5 m および 60 m)は成層期でのデータであると言える.Figure 3.1 Seasonal variation of temperature in water column at 17B and 12B from April in 2016 through January in 2017.
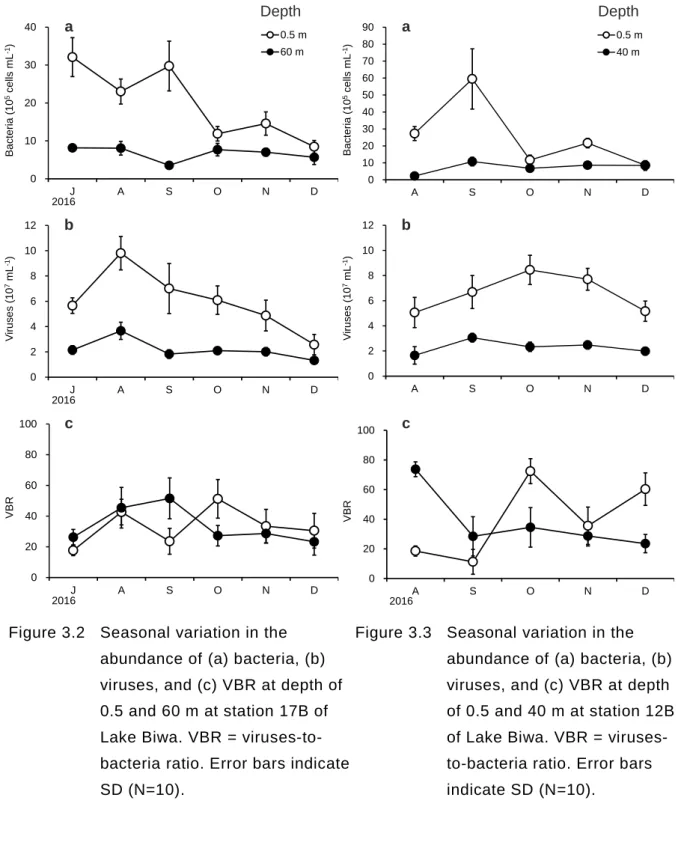
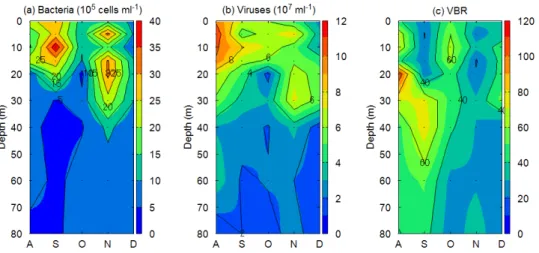
3.2 全菌数,ウイルス数および VNR (1)地点 17B 表水層(0.5m)における細菌数とウイルス数は,成層期が深水層(60 m)に比べて高い 結果となった(Figure 3.2a, b).観測期間(7~12 月)を通して,表水層での細菌は 2.0 ± 1.0 × 106 cells mL-1,ウイルスは 6.0 ± 2.4 × 107 mL-1となっているのに対し,深水層では細 菌は 0.7 ± 0.2 × 106 cells mL-1,ウイルスは 2.2 ± 0.8 × 107 mL-1となっており,表水層と深 水層で有意な差を示した(p < 0.05).また 7 月から 12 月にかけ水温躍層が弱くなるにつ れ,表水層と深水層における細菌とウイルス数の差は小さくなった.しかし,細菌数に対 するウイルス数の比は成層期においても表水層(33.2 ± 12.3)と深水層(33.7 ± 11.7)で有 意な差はなかった(p > 0.05).また,季節変動も有意に差があるとは言えず,表水層,深 水層ともに概ね一定の値をとっていた(Figure 3.2c). 地点 17B での観測期間における各水深の細菌数およびウイルス数を Figure 3.4 に示す.細 菌数,ウイルス数ともに極大値は表水層(0.5 m)ではなく,水深 5-10 m に存在した.また 11 月では水深 5 m と 20 m 付近が細菌,ウイルスのもっとも多い水深となった. (2)地点 12B 表水層の細菌数(8.6~59.5 × 105 cells mL-1)は,地点 17B と同様に夏から 12 月にかけ
て深水層(2.2~10.8 × 105 cells mL-1)での値に近づいていた(Figure 3.3a).しかし,ウイ
ルス数にその傾向はみられず,観測期間を通して,表水層(6.6 ±1.5 × 107 mL-1)が深水層 (2.3 ± 0.5 × 107 mL-1)に比べて常に高い値となった(p < 0.05).また VBR は,観測期間を 通して,表水層では増加傾向にあり,深水層では減少傾向にあった. (3)表水層の細菌数およびウイルス数 細菌数は 8 月から 12 月にかけてどの地点においても減少傾向にあった(Figure 3.5a). ウイルス数においても減少傾向が観られたが,地点 12B と 6B では減少傾向が見られなか った.
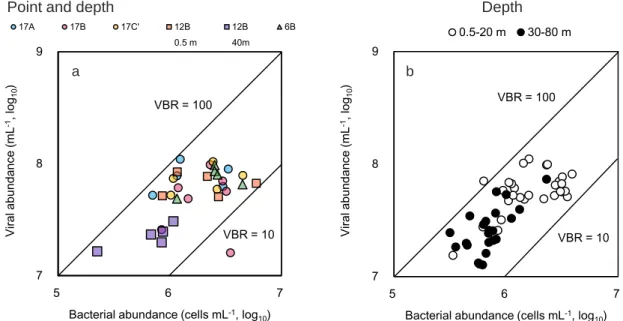
(4)VBR(viruses to bacteria ratio)
琵琶湖の 17A,17B,17C’,12B,6B の表水層および 12B の水深 40 m において,VBR は 10~100 の間にあった(Figure 3.6a).また地点 17B の全水深においても VBR は 10~100 の間を推移した(Figure 3.6b).このことから,琵琶湖において水深や水域によらず VBR は 10~100 の間を推移することが示唆された.ただし,琵琶湖の表水層(17A,17B,17C’, 12B および 6B)では細菌数とウイルス数の間の相関は弱く,また 17B において水深が深く なるにつれウイルスと細菌に強い正の相関が観られた.同様に 12B の水深 40 m において 強い正の相関が観られた.このことから,表水層でのウイルスの一部にはシアノファージ が含まれており,深水層ではほぼバクテリオファージであることが示唆された.
Figure 3.2 Seasonal variation in the abundance of (a) bacteria, (b) viruses, and (c) VBR at depth of 0.5 and 60 m at station 17B of Lake Biwa. VBR = viruses-to-bacteria ratio. Error bars indicate SD (N=10).
Figure 3.3 Seasonal variation in the abundance of (a) bacteria, (b) viruses, and (c) VBR at depth of 0.5 and 40 m at station 12B of Lake Biwa. VBR = viruses-to-bacteria ratio. Error bars indicate SD (N=10). 0 10 20 30 40 J A S O N D B ac ter ia ( 10 5c e lls m L -1) 2016 0.5 m 60 m 0 2 4 6 8 10 12 J A S O N D V ir us es ( 10 7mL -1) 2016 0 20 40 60 80 100 J A S O N D VBR 2016 a b c 0 10 20 30 40 50 60 70 80 90 A S O N D B ac ter ia ( 10 5c e lls m L -1) 0.5 m 40 m 0 2 4 6 8 10 12 A S O N D V ir us es ( 10 7mL -1) 0 20 40 60 80 100 A S O N D VBR 2016 a b c Depth Depth
Figure 3.4 Seasonal variation of (a) bacterial abundance, (b) viral abundance, and (c) VBR in water column at 17B from August through December in 2016.
Figure 3.5 Seasonal variation of (a) bacterial abundance and (b) viral abundance at depth of 0.5 m at 17A, 17B, 17C’, 12B and 6B from August through December in 2016. Error bars indicate SD (N=10).
0 10 20 30 40 50 60 70 A S O N D B ac ter ia ( 10 5c e lls m L -1) 17A 17B 17C' 0 10 20 30 40 50 60 70 A S O N D B ac ter ia ( 10 5c e lls m L -1) 2016 12B 6B 0 3 6 9 12 15 A S O N D V ir us es ( 10 7mL -1) 17A 17B 17C' 0 3 6 9 12 15 A S O N D V ir us es ( 10 7mL -1) 12B 6B a b d b Point Point Point Point
Figure 3.6 VBR (viruses to bacteria ratio) at (a) surface water in 17A, B, C’, 12B, 6B and 40 m in 12B and (b) water column in 17B, through July to December in 2016. 7 8 9 5 6 7 Vi ral abundanc e ( m L -1, l og10 )
Bacterial abundance (cells mL-1, log10)
17A 17B 17C' 12B 0.5 m 12B 40m 6B VBR = 10 VBR = 100 7 8 9 5 6 7 Vi ral abundanc e ( m L -1, l og10 )
Bacterial abundance (cells mL-1, log10)
0.5-20 m 30-80 m
VBR = 100
VBR = 10
a b
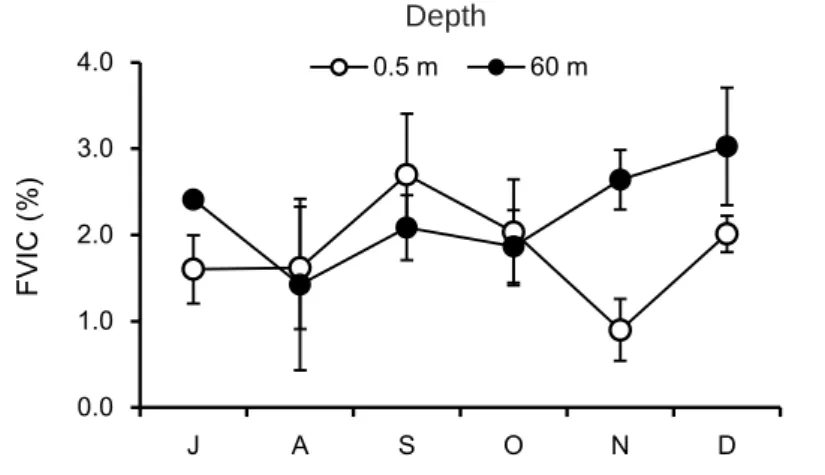
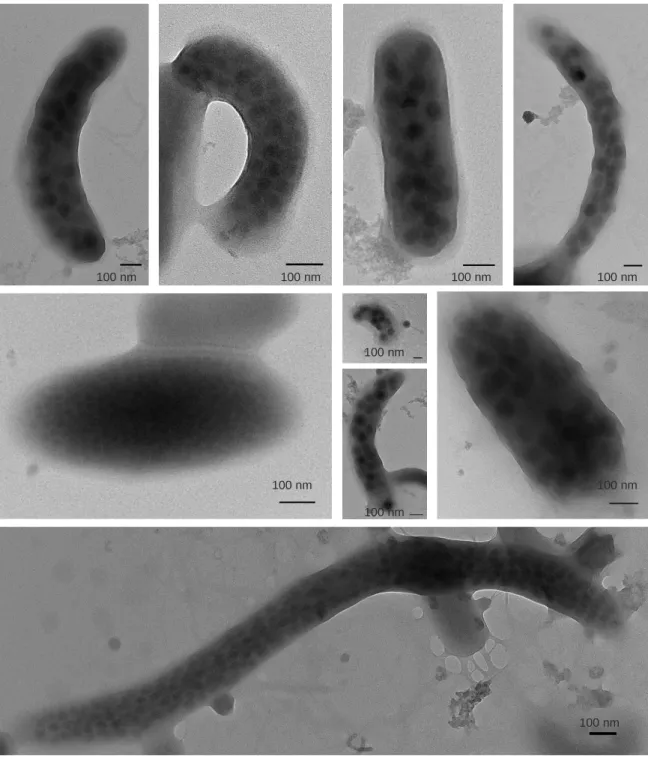
3.3 感染率(FVIC)およびバーストサイズ 観測期間を通して,地点 17B 表水層(0.5 m)での感染率は 0.9~2.7%を推移し,平均は 1.8 ± 0.6%であったのに対し,深水層(60 m)では 1.4~3.0%を推移し,平均は 2.2 ± 0.6% であった(Figure 3.7).成層期において,深水層が表水層に比べ平均値は高いものの,有 意な差はなかった(p > 0.05).FMVL は表水層で 5.9~35.9%であったのに対して深水層で は 19.3~43.0%であった.また得られた TEM 画像から計数したバーストサイズは 5~150 以上の範囲内であり,表水層では 13.4 ± 6.3,深水層では 15.1 ± 10.6 となり,2 水深間ある いは月別での明確な違いはなかった. Figure 3.8 に感染した細菌の TEM 写真を示す.細菌の細胞内に見られる黒い粒子がウイ ルス粒子である.観察された細胞内のウイルス粒子は大きさが約 30~120 nm の範囲で存 在した.
Figure 3.7 Seasonal variation of FVIC (Frequency of visibly infected cells) at 17B in Lake Biwa, Japan from July through December in 2016.
0.0 1.0 2.0 3.0 4.0 J A S O N D FV IC (% ) 0.5 m 60 m Depth
Figure 3.8 Transmission electron micrographs (TEM) of bacterial cells infected by viruses. 100 nm 100 nm 100 nm 100 nm 100 nm 100 nm 100 nm 100 nm 100 nm
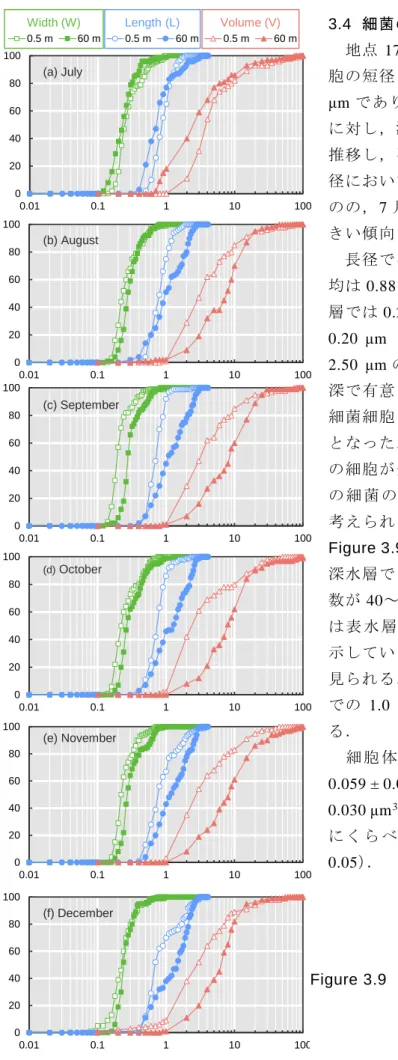
3.4 細菌の細胞体積 地点 17B 表水層(0.5 m)における細菌細 胞の短径(W)は観測期間を通して,0.08-1.10 μm であり平均は 0.27 ± 0.03 μm であったの に対し,深水層(60 m)では 0.12-1.29 μm を 推移し,平均は 0.31 ± 0.04 μm となった.短 径においては 2 水深で明確な差はなかったも のの,7 月を除いて深水層の細菌のほうが大 きい傾向にあった. 長径では,表水層が 0.18-3.71 μm であり平 均は 0.88 ± 0.10 μm であったのに対し,深水 層では 0.25-3.09 μm を推移し,平均は 1.27 ± 0.20 μm となった.ほとんどの細菌は 0.50-2.50 μm の範囲にあった.またこれらの 2 水 深で有意な差があり(p < 0.01),深水層での 細菌細胞は表水層に比べ 1.5 倍程度長い結果 となった.この長径の差は,深水層での細菌 の細胞が全体的に大きいわけではなく,一部 の 細 菌 の 長 径 が 長 い た め に 生 じ た も の だ と 考えられる(Figure 3.9).その根拠として, Figure 3.9 の長径において,累積度数曲線は 深水層で 2 種類の上昇を示している.累積度 数が 40~60%付近(7 月のみ 90%付近)まで は 表 水 層 の 累 積 度 数 曲 線 と 類 似 し た 上 昇 を 示しているが,40~60%付近から違う上昇が 見られる.これは,二段階目の上昇が深水層 での 1.0 μm 以上の細菌数の多さを表してい る. 細 胞 体 積 は 観 測 期 間 を 通 し て , 表 水 層 で 0.059 ± 0.015 μm3であり,深水層では 0.097 ± 0.030 μm3であった.深水層での細菌は表水層 に く ら べ 2 倍 弱 大 き い 結 果 と な っ た ( p < 0.05). 0 20 40 60 80 100 0.01 0.1 1 10 100 C um ul at iv e V al ue ( % ) 0.5 m 60 m 0.5 m 60 m 0.5 m 60 m 0 20 40 60 80 100 0.01 0.1 1 10 100 C um ul at iv e V al ue ( % ) 0 20 40 60 80 100 0.01 0.1 1 10 100 C um ul at iv e V al ue ( % ) 0 20 40 60 80 100 0.01 0.1 1 10 100 C um ul at iv e V al ue ( % ) 0 20 40 60 80 100 0.01 0.1 1 10 100 C um ul at iv e V al ue ( % ) 0 20 40 60 80 100 0.01 0.1 1 10 100 C um ul at iv e V al ue ( % )
Width (W) Length (L) Volume (V)
(a) July (b) August (c) September (d) October (e) November (f) December
Figure 3.9 Cumulative curve of width, length and volume of bacterial cells at the depth of 0.5 m and 60 m at 17B.
4. 考察
前述の結果から,本章ではいくつかの結論を得た.細菌数が少ない環境において感染成 功率はウイルスの生存を決定する要因の一つになっている.また琵琶湖の物質循環におい てウイルス感染に伴い放出される DOM は有機物あるいは栄養塩を再利用する重要な供給 源として駆動している.とくに深水層では,細菌がウイルス感染によって死亡する割合は 高く,他の細菌の重要な栄養源になっている.ここでは得られた結果を更に解析し,考察 を加える. 4.1 ウイルスの回転速度および感染成功率ウイルスの生産速度 VP (Viral production,mL-1 d-1)は,細菌の生産速度 BP(Bacterial
production,cells mL-1 d-1),バーストサイズ S (Burst size,viruses cell-1),および細菌の生
産速度に占めるウイルス感染による死亡の割合 F(FMVL; Frequency of bacterial mortality due to viral lysis,%)を用いて,式(4.1)のように表すことが出来る.この式(4.1)は, 単位時間あたりに生産された細菌数のうち,ある一定の割合の細菌数がウイルス感染によ って死亡し,その際に各死亡細胞から S だけのウイルスが放出されることを意味している.
F
S
BP
VP
=
⋅
⋅
(4.1) また,細菌の生産速度 BP は細菌増殖速度 μ と細菌数 BA (cells mL-1),を用いると,以 下の関係式が成り立つ.BA
BP
=
µ
⋅
(4.2) さらに,ウイルスの回転速度(TV [d])と細菌の回転速度(TB [d],Turnover time)は以 下の式に従い求めることが出来る.ただし,VA(Viral abundance)はウイルス数(mL-1)を 表す.VP
VA
T
V=
(4.3)BP
BA
T
B=
(4.4) 式(4.3)と式(4.4)を式(4.1)に代入すると,式(4.5)が得られる. B VT
BA
VA
F
S
T
⋅
⋅
⋅
=
1
(4.5) 式(4.5)は,ウイルスの回転時間 TVは細菌の回転時間 TBとともに変化することを示し ている.そして,バーストサイズ S の逆数,ウイルス感染による死亡が細菌生産速度に占 める割合 F の逆数,さらに VBR(VA/BA)の関数である.また,衝突頻度 R(Contact rate [contacts mL-1 d-1])を細菌数 BA で除することで,1 細胞当たりのウイルスとの衝突頻度Rbacteria(contacts cell-1 d-1)が得られる.
BA
R
R
bacteria=
(4.6) また,ウイルスの宿主への感染成功率 IS(Infection success,%(cell/contancts))は以下の 式(4.7)により求めることができる.100
⋅
⋅
=
bacteriaR
F
IS
µ
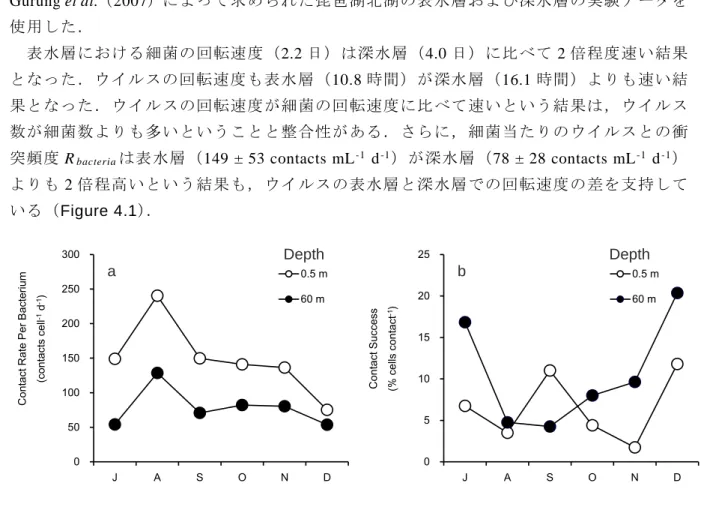
(4.7) 式(4.7)において,分子は増殖した細菌のうちウイルス感染によって死亡する細菌の割合 を表している.したがって感染成功率とは,一匹の細菌がウイルスと衝突した回数のうち, 感染が起こり死亡する割合を示している. これらの式を用いて,本章で得られたデータから,実際に琵琶湖でウイルスが回転する 速度や,細菌の回転速度に対する比を求めた(Table 4.1).ただし,細菌の増殖速度 μ は Gurung et al.(2007)によって求められた琵琶湖北湖の表水層および深水層の実験データを 使用した. 表水層における細菌の回転速度(2.2 日)は深水層(4.0 日)に比べて 2 倍程度速い結果 となった.ウイルスの回転速度も表水層(10.8 時間)が深水層(16.1 時間)よりも速い結 果となった.ウイルスの回転速度が細菌の回転速度に比べて速いという結果は,ウイルス 数が細菌数よりも多いということと整合性がある.さらに,細菌当たりのウイルスとの衝 突頻度 Rbacteriaは表水層(149 ± 53 contacts mL-1 d-1)が深水層(78 ± 28 contacts mL-1 d-1)よりも 2 倍程高いという結果も,ウイルスの表水層と深水層での回転速度の差を支持して いる(Figure 4.1). 0 50 100 150 200 250 300 J A S O N D C ont ac t R at e Per B ac ter ium (co nt act s ce ll -1d -1) 0.5 m 60 m 0 5 10 15 20 25 J A S O N D C ont ac t S uc ces s (% ce lls co nt act -1) 0.5 m 60 m a b Depth Depth
Figure 4.1 Seasonal variation of (a) contact rate per bacterium and (b) contact success at 2 layers (0.5 and 60 m) in 17B from July through December in 2016.
感染成功率 IS は表水層(6.5 ± 4.1%)と深水層(10.6 ± 6.6%)で逆転している(Figure 4.1b).表水層のウイルス数が深水層に比べて多いこと(Figure 3.2)は,細菌あたりのウ イルスとの衝突頻度が,表水層が深水層よりも高いことに繋がる.しかし,感染の成功率 が逆転していることから,表水層では,太陽光によって不活化しているウイルスが多い可 能性を示している.また,深水層での感染成功率の高さは,深水層でのウイルスが太陽光 によってほとんど不活化していないと考えられることから説明できる. さらに,海洋のデータではあるが,ウイルスが 1 種類の細菌にのみ感染力を示すとすれ ば(Børsheim, 1993),深水層での感染成功率の高さはその水域の細菌の多様性が低いこと を意味している.ただし,近年になり数種類(近縁種)の細菌に感染するウイルスの存在 も報告されており(Børsheim, 1993),必ずしも多様性とは結び付けられるわけではない. しかし,琵琶湖における深水層での生物多様性の低さの根拠として,CL500-11 系統の細菌 が琵琶湖の深水層で 11 月から 12 月に優先することが挙げられる(Okazaki et al., 2013). Okazaki et al.(2013)は 2 年間にわたる琵琶湖北湖沖帯での調査により,成層の形成が始 まると深水層で CL500-11 の現存量が増加していき,11~12 月の成層期末期で現存量は最
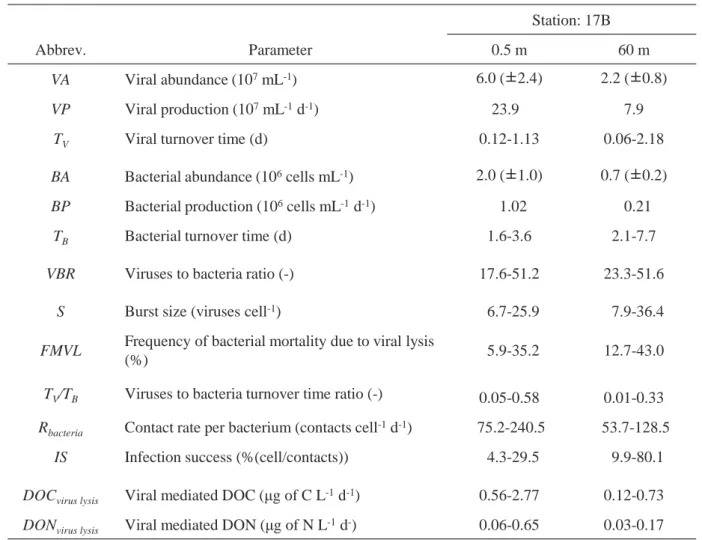
Table 4.1 Impact of viruses on bacteria at the surface layer and deep layer in Lake Biwa from July to December in 2016. Viral abundance and bacterial abundance are mean of 6 months (±SD). Ranges for each parameter show minimum and maximum during sampling period.
Station: 17B
Abbrev. Parameter 0.5 m 60 m
VA Viral abundance (107mL-1) 6.0 (±2.4) 2.2 (±0.8)
VP Viral production (107mL-1d-1) 23.90 7.9
TV Viral turnover time (d) 0.12-1.13 0.06-2.18
BA Bacterial abundance (106cells mL-1) 2.0 (±1.0) 0.7 (±0.2)
BP Bacterial production (106cells mL-1d-1) 01.02 00.21
TB Bacterial turnover time (d) 1.6-3.6 2.1-7.7
VBR Viruses to bacteria ratio (-) 17.6-51.2 23.3-51.6
S Burst size (viruses cell-1) 06.7-25.9 07.9-36.4
FMVL Frequency of bacterial mortality due to viral lysis
(%) 05.9-35.2 12.7-43.0
TV/TB Viruses to bacteria turnover time ratio (-) 0.05-0.58 0.01-0.33
Rbacteria Contact rate per bacterium (contacts cell-1d-1) 075.2-240.5. 053.7-128.5.
IS Infection success (%(cell/contacts)) 04.3-29.5 09.9-80.1
DOCvirus lysis Viral mediated DOC (μg of C L-1d-1) 0.56-2.77 0.12-0.73
大値を取ること,それが全細菌の 10.0~16.5%を占めること,そして循環期には検出限界 以下に減少することを報告している.これは Figure 4.1b での深水層の感染成功率が 8~12 月にかけて上昇していることと一致している.つまり,観測期間において深水層のウイル ス数はほぼ一定であるが(Figure 3.2),多様性が低下することで,それに伴うウイルスの 多様性も低下する.よって,ウイルスが感染可能な細菌が増加し,結果として感染の成功 率が増加する.また,CL500-11 は細胞サイズが 1~2 μm 程度であることも,成層期末期に な る に 従 い 長 径 が 長 い 細 菌 が 深 水 層 で 増 加 し て い る TEM の観察結 果とも一致している (Figure 3.9).これらのことから,細菌数の低い深水層において感染成功率はウイルスの生 存にかかわる一つの要因であることが示唆された. 4.2 ウイルスの琵琶湖物質循環への寄与
細菌の単位時間・単位体積あたりの炭素生産速度 BCP(Bacterial carbon production, μg of C mL-1 d-1)は,一細胞当たりの体積 v(μm3 cell-1),細胞体積あたりの炭素含有量 c(μg of C μm-3),および細菌生産速度 BP を用いて,式(4.8)で表すことが出来る.
BP
v
c
BCP
=
⋅
⋅
(4.8) また,湖水中における細菌数が定常状態にあると仮定すると,細菌の生産速度と死亡速 度は見掛け上釣り合っているので,ウイルス感染に伴い放出される DOC(DOCviral lysis; Viralmediated dissolved organic carbon)は,FMVL(F)を用いて求められる.
BCP
F
DOC
virallysis=
⋅
(4.9)年間に放出される DOCviral lysisを推定する際に,1 年を通じての表水層(0.5 m)と深水層
(60 m)のいくつかの数値が必要になる.しかし,本研究で得られたデータは 7~12 月まで のものである.そこで,本章では Table 4.2 中の結果と以下の設定をもとに,年間の DOCviral
lysisを推定した. i. 3.1 節での 17B における水温データから,成層期を 4~12 月,循環期を 1~3 月 とした. ii. 琵琶湖南湖の体積は琵琶湖全体の 1%未満であるため,計算対象を琵琶湖北湖 (623 km2)とした. iii. 表水層と深水層の境μ 界は,夏(6~9 月)に形成される水温躍層 15 m とした. iv. 細菌の含有炭素量と窒素量は Nagata(1986)による琵琶湖での値を用いた. v. データが得られている 7~12 月では,表水層も深水層も数値をそのまま利用し た. vi. 深水層での欠損データ(1~6 月)は,7~12 月の深水層のデータを平均した. vii. 成層期における表水層の欠損データ(4~6 月)は,7~12 月の表水層データの 平均値を利用した. viii. 循環期における表水層の欠損データ(1~3 月)は,深水層での平均データを利 用した.
推定したウイルス感染により放出される年間 DOC 量は,表水層で 3,770 tonC,深水層で 3,150 tocC であった.これらは表水層の細菌生産量の 15.8%,深水層の 36.1%である.琵琶 湖全体で考えると,内部生産 107,337 tocC/y(岡本ら,2012)のうち 30.4%(32,600 tocC/y) が細菌生産に消費されており,細菌生産量うちの 21.2%(6,920 tonC/y)がウイルス感染に より DOC に回帰している.この値は海洋で報告されている値よりも低いが(Wilhelm and Suttle, 1999),この理由としては,海洋(主に外洋)に比べて一次生産が活発である点が挙 げられる.また,琵琶湖では内部生産の約 20%が沈降フラックス(5,699 tonC/y)として深 水層へ移動し,深水層での重要な有機栄養源となっている.しかし,本章で推定されたウ イルス感染により放出される DOC 量は深水層で 3,150 tonC/y であり,沈降フラックスに匹 敵する重要な有機栄養源であることが分かる.DON(Dissolved organic nitrogen)も同様に ウ イ ル ス感 染 に よ っ て 放 出 さ れる . 放 出 さ れ る DON は細胞内でほとんどがアミノ酸や DNA など易分解性の有機物に含まれていることから,放出後は細菌によりすぐに利用され ると考えられる.さらに利用された易分解性の細胞由来の有機物は無機化されて窒素やリ ンなどの栄養塩として環境水中に放出される.このようにウイルスとその後に続く細胞破 壊プロセスは,細菌や藻類への重要な利用性の高い栄養源として琵琶湖水中で重要な役割 を果たしている.
5. 結論と今後の課題
本研究では,ウイルスとそれに続く溶菌プロセスの琵琶湖物質循環における寄与を推定 するため,毎月の現場観測(7~12 月)から細菌・ウイルス数やウイルスによる細菌の感 染率を調査した.琵琶湖北湖今津沖中央においては,細菌数は表水層で 0.8~3.2 × 106 cells mL-1,深水層で 0.4~0.8 × 106 cells mL-1であった.また,ウイルス数は表水層で 2.6~9.8 × 106 mL-1,深水層で 1.3~3.7 × 107 mL-1であった.表水層での細菌数とウイルス数は 7~12 月にかけて深水層での値に近づいた.また感染率は表水層で 0.9~2.7%,深水層で 1.4~3.0 であった.推定された感染成功率は深水層(4.8~20.4%)の方が表水層(1.7~11.8%)に比 べて高く,細菌数の少ない深水層において感染成功率を上げる生存戦略をウイルスは取っ ていることが示唆された.また,年間のウイルス感染により放出される DOC(Dissolved organic carbon)は,約 6,900 tonC であり,細菌炭素生産量の約 20%,一次生産の約 6%で あることが分かった.深水層に限れば,ウイルス感染により放出される DOC 量は,深水層 での細菌炭素生産量の約 36%を占めた.以上のことから,ウイルスが琵琶湖物質循環にお いて重要な役割を果たしていることが示された. ウイルス分流はウイルスが細菌に感染することで始まる.しかし,感染率,つまり衝突 頻度と感染成功率がどのような因子に支配されているかについてはよく分かっていない. 感染率を左右する要因として,宿主の多様性やウイルスとの組み合わせ,あるいは細胞へ の付着に関与する物質や水温などの物理因子などが考えられるが,解明には至っていない. 今後は,感染率の制御因子の探索が必要になると考えられる.また,実際に琵琶湖水中に 存在しているウイルスのうちシアノファージとバクテリオファージがどの程度の割合を占 めているか,あるいは,それらの不活化率を把握することは,より正確なウイルスの寄与 を評価する上で重要な知見となり得る.6. 参考文献
Binder B. (1999): Reconsidering the Relationship between Virally Induced Bacterial Mortality and Frequency of Infected Cells, Aquatic Microbial Ecology, 18, 207–215.
Børsheim K.Y. (1993): Native Marine Bacteriophage, FEMS Microbiology Ecology, 102, 141-159. Dubin S.B., Benedek G.B., Bancroft F.C. and Freifelder D. (1970): Molecular Weights of
Coliphages and Coliphage DNA II. Measurement of Diffusion Coefficients using Optical Mixing Spectroscopy, and Measurement of Sedimentation Coefficients, Journal of Molecular Biology, 54, 547-556.
Jover L.F., Effler T.C., Buchan A., Wilhelm S.W. and Weitz J.S. (2014): The Elemental Composition of Virus Particles: Implication for Marine Biogeochemical Cycles, Nature Reviews Microbiology, 12, 519-528.
Mitchell J.G., Pearson L Bonazinga A, Dillon S and Khouri H and Paxinos R (1995): Long Lag Time and High Velocities in the Motility of Natural Assemblages of Marine Bacteria, Applied and Environmental Microbiology, 61, 877-882.
Murray A.G. and Jackson G.A. (1992): Viral Dynamics: A Model of the Effects of Size, Shape, Motion and Abundance of Single-celled Planktonic Organism and Other Particles, Marine Ecology Progress Series, 89, 103-116.
Nagata T. (1986): Carbon and Nitrogen Content of Natural Planktonic Bacteria, Applied and Environmental Microbiology, 52(1), 28-32.
Noble R.T. and Fuhrman J.A. (1998): Use of SYBR Green I for Rapid Epifluorescence Counts of Marine Viruses and Bacteria, Aquatic Microbial Ecology, 14, 113-118.
Ogawa H., Amagai Y., Koike I., Kaiser K. and Benner R. (2001): Production of Refractory Dissolved Organic Matter by Bacteria, Science, 292, 917-920.
Okazaki Y., Hodoki Y. and Nakano S. (2013): Seasonal Dominance of CL500-11 Bacterioplankton (Phylum Chloroflexi) in the Oxigenated Hypolimnion of Lake Biwa, Japan, FEMS
Microbiology Ecology, 83, 82-92.
Patel A., Noble R.T., Steele J.A., Schwalbach M.S., Hewson I. and Fuhrman J.A. (2007): Virus and Prokaryote Enumeration from Planktonic Aquatic Environments by Epifluorescence Microscopy with SYBR Green I, Nature Protocols, 2 (2):, 269-276.
Pradeep Ram A.S., Nishimura Y., Tomaru Y., Nagasaki K. and Nagata T. (2010): Seasonal Variation in Viral-induced Mortality of Bacterioplankton in the Water Column of a Large Mesotrophic Lake (Lake Biwa, Japan), Aquatic Microbial Ecology, 58, 249-259.
Suttle C.A. (2007): Marine Viruses – Major Players in the Global Ecosystem, Nature Reviews Microbiology, 5, 801-812.
Weinbauer M.G., Winter C., Höfle M.G. (2002): Reconsidering Transmission Electron
Microscopy based Estimates of Viral Infection of Bacterioplankton using Conversion Factors derived from Natural Communities, Aquatic Microbial Ecology, 27, 103–110.
Wilhelm S.W. and Suttle C.A. (1999): Viruses and Nutrient Cycles in the Sea, BioScience, 49, 781-788.
岡本高弘,佐藤祐一,早川和秀,一瀬諭(2012):水質汚濁メカニズムの解明に関する政 策課題研究―難分解性を考慮した琵琶湖における有機物の現状と課題―.滋賀県琵 琶湖環境科学研究センター試験研究報告書,7,87-102. 岡本高弘,早川和秀 (2011):琵琶湖における溶存有機物の現状と課題.水環境学会誌, 34 (A),151-157. 今井章雄,福島武彦,松重一夫,井上隆信,石橋敏昌(1998):琵琶湖湖水および流入河 川水中の溶存有機物の分画.陸水学雑誌,59,53-68. 佐藤祐一,岡本高弘,早川和秀,大久保卓也,小松英司(2016):琵琶湖における難分解 性有機物の起源:発生源における生分解試験とボックスモデルによる推計.水環境 学会誌,39(1),17-28. 滋賀県琵琶湖環境部(2014 年):平成 25 年度公共用水域水質測定結果(琵琶湖・河川) 資料 1. 謝辞 琵琶湖調査および底質コアサンプルの採取では,それぞれ滋賀県琵琶湖環境科学研究セン ターおよび京都工芸繊維大学・布施泰朗助教の多大なるご支援を賜りました.本研究を遂 行するにあたり研究室の院生・学生諸氏のご協力をいただきました.ここに記して謝意を 表します. ・助成事業者紹介 清水芳久 現職:京都大学大学院工学研究科教授(Ph.D)