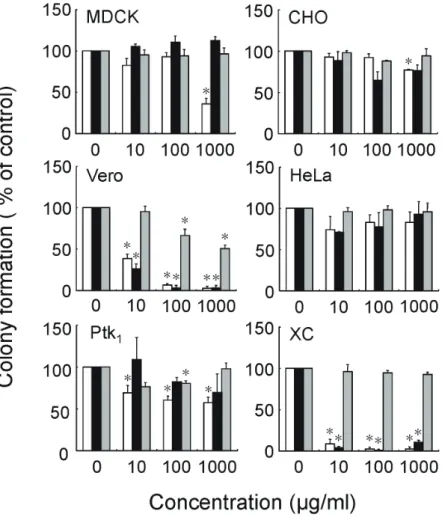
Running title: Cell Growth Promoting Activity of Ascophyllan
Effects of Sulfated Fucan, Ascophyllan, from the Brown Alga Ascophyllum nodosum on Various Cell Lines: A Comparative Study on Ascophyllan and Fucoidan
Zedong JIANG,
1Takasi OKIMURA,
2Takeshi YOKOSE,
1,3Yasuhiro YAMASAKI,
4Kenichi YAMAGUCHI,
1,4and Tatsuya ODA
1, 41
Graduate School of Science and Technology, Nagasaki University, Nagasaki 852-8521, Japan
2
Research and Development Division, Hayashikane Sangyo Co., Ltd., Shimonoseki, Yamaguchi 750-8608, Japan
3
Nagasaki Prefectural Institute for Environment Research and Public Health, 11306-11, Ikeda 2-chome, Omura, Nagasaki, 856-0026, Japan
4
Division of Biochemistry, Faculty of Fisheries, Nagasaki University, Nagasaki 852-8521, Japan.
Key words: Ascophyllum nodosum; ascophyllan; fucoidan; sulfated fucan; cytotoxicity;
growth promotion; MDCK cells