ブラジル産ハリナシハナバチのアラタ体
22
0
0
全文
(2) Vol. 18, No. 1 Journal of Hokkaido University of Education (Section II B) September 1967. Corpora Allata in Brazilian Stingless Bees1. By Yukio AKAHIRA, Darvin BEIG25 and Warwick E. KERR35 Biological Laboratory, Hokkaido University of Education, Kushiro. ?!r^^S : 77 '^/^^^^ y -)-'y^^-^^-(D~7 =7 ^-^. There are many publications concerning Hymenopteran corpora allata, including anatomical descriptions, biometrical studies, and studies of endocrinological interaction with the other organs, especially with the reproductive system, or the development of worker ovaries (HanstrOm, 1939; Nabert, 1913 ; Palm, 1948 ; Williams, 1958, etc.). According to our studies, on the other hand, it has been verified that among colonies of the so-called stingless bees three types of development of worker ovaries exist under normal queen-right condition, i.e., 1) a type having well-developed worker ovaries, for example, Scaptotrigona postica, Melipona group, and others ; 2) a type in which worker ovaries never develop, for example, Frieseomelitta group, Hypotrigona mullerii, Dnckeola ghilianu, Partamona group, and others ; 3) a type having partially developed worker ovaries, for example, Plebeia droryana, Nannotrigona testaceicornis, and others (Sakagami et al., 1963). The present study has been carried out under the aim of making biometrical and histological comparisons of corpora allata among two tribes of stingless bee {Trigonini and JVteliponini) and Apis, attempting to decide the interrelations between activities of the corpora allata and development of worker ovaries. Our sincere thanks are also due to Prof. S. F. Sakagami, Zoological Institute, Faculty of Science, Hokkaido University, for his kind help and suggestions concerning this work.. Material and Method The materials for this investigation generally consist of five pairs of corpora allata of worker bees, principally foragers, per species (or ten corpora allata, when found not paired under microscopical observations). 1) The present study has been carried in the Laboratory of General Biology, Faculty of Philosophy, Science and Literature of Rio Claro, Brazil, where the first author had investigated in past two years, and it was performed with the financial support of Unived States Department of Agriculture Research Service, under Public Law 480, and of the following Brazilian organizations : Fundagao de Amparo a Pesquisa. do Estado de Sao Paulo ; Companha Nacional de Aperfeigoamento de Pessoal de Nivel Superior (CAPES). 2) Faculdade de Filosofia, Cienclas e Letras de Rio Clara, Sao Paulo, Brasil. 3) Faculdade de MecUcina de Riberao Preto, Universidade de Sao Paulo, Brasil.. (24).
(3) Table I. Measurements of corpora allata, see detailed explanation in text. Cephalotri, ;ona femorata. N. N/. w. 1 2 3 4 5 6 7 8 9 10. 85 69 68 59 46 76 65 67 71 72. 16 16 16 16 10 21 18 6 8 8. 78 82 90 85 71 78 58 71 79 51. x. 67.8. 13.5 74.3 19.9. •ies. _%_ c.v.. Trigona (T.) cilipes cilipes. H. D. v. p. N. 17 18 20 20 18 15. 49 49 49 35 35 49 42 49 42 35. 393 413 430 356 280 348 230 378 344 325. 1 1 2 2 3 4 5 6 7 8. 23 25 17 33 30 41 55. 17. 21 18 28. 19.2 43.4. 349.7 5.15 25.29. V.I.. T . (Frieseomelitta) flavicornis. Species. N. N'. w. H. D. 1 2 3 4 5 6 7 8 9 10. 33 33 22 28 31 34 38 28 23. 14. 22. 43. 32 26 23 27 28 25 21 26 18. 28 '. 39. 42 37 34 37 43 35 43 28 37. x. 30.9. 10.6 34.3. 37.9. %_. 11 12. 8 9. 11. 7 8 4. v. p. 32. 35 28 21 35 28 35 35 21. 42. 229 175 149 129 231 143 203 155 88. 2 2 3 3 4 4 5. 25.8. 30.8. 184.6. 344. 1. 1. 6. 5.97. c.v.. 101.70. V.I.. T. (Meliponula) bocandei. Species. N. N/. w. H. D. v. p. 1 2 3 4 5 6 7 8 9 10. 57 94 62 61 98 98 65 74 84 65. 3 2 2 3 4 2 14 5 20. 49 52 49 52 52 49 40 52 49 45. 28 35 33 33 31 36 36 28 38 27. 49 56 35 49 56 49 35 56 49 42. 396 660 346 420 511. 1 1 2 2 3 3 4 4 5 5. x. 75.8. %. c.v.. 11. 6.6 48.9 8.7. 32.5 47.6. 511. 283 356 509 261 425.3 5.61 23.20. V.I.. Partamona.(P.) pearsonn. Species. N. N'. w. H. D. v. p. 1 2 3 4 5 6 7 8 9 10. 28 51 50 22 45 54 57 33 60 29. 18 21 19 10 18 16 21 10 19. 49 50 47 40 45 43 40 31 53 43. 21 31 34 23 23 23 23 20 39 35. 28 28 35 21 43 42 56 35 35 14. 175 235 318 127 235 209 291 132 404 152. 1. x. 42.9. 16.6 44.1 38.6. 27.2. 33.6. 227.8. %. c.v. V.I.. 14. 5.31 29.34. 1. 2 2 3 3 4 4 5 6.
(4) 34.3. _%_. 5.97. c.v.. 101.70. V.I.. T. (Meliponula) bocandei. N. N'. w. H. D. v. p. 1 2 3 4 5 6 7 8 9 10. 57 94 62 61 98 98 65 74 84 65. 3 2 2 3 4 2 14 5 20. 49 52 49 52 52 49 40 52 49 45. 28 35 33 33 31 36 36 28 38 27. 49 56 35 49 56 49 35 56 49 42. 396 660 346 420 511 511 283 356 509 261. 1 1 2 2 3 3 4 4 5 5. x. 75.8. 48.9. 32.5 47.6. _%_. c.v.. 11. 6.6 8.7. 425.3 5.61 23.20. V.I.. PartamonaI (P.) pearsonn >ecies. N. N/. vv. H. D. v. p. 2 3 4 5 6 7 8 9 10. 28 51 50 22 45 54 57 33 60 29. 18 21 19 10 18 16. 49 50 47 40 45 43 40 31 53 43. 21 31 34 23 23 23 23 20 39 35. 28 28 35 21 43 42 56 35 35 14. 175 235 318 127 235 209 291 132 404 152. 1. x. 42.9. 27.2. 33.6. 227.8. 1. %. c.v.. 21. 10 19 14. 16.6 44.1 38.6. 1. 2 2 3 3. 4. 4 5 6. 5.31 29.34. V.I.. Melipona <:ompressipes manaosensis Species. N. N/. w. H. D. v. p. 1 2 3 4 5 6 7 8 9 10. 122 121 128 134. 18 18. 70. 47. 42 42 56 63 63 63 42 42 35 49. 702 703 795 940 1199 1016 710 715 654 656. 1 1 2 2 3 3 4 4 5 5. x. %.. 126. 132 99. 112. 102 106. 27 31. 25 17 18 19 19 19. 67 90 85 71 59 85 85 70 64. 49 29 32 49 50 35 34 45 40. 118.2 21.1 74.6 41.0 17.8. 49.7. 809.0 6.84 38.47. c.v. V.I.. M. seminigra merrillae Species. N. N'. w. H. D. v. p. 1 2 3 4 5 6 7 8 9 10. 209 181 187 176 146 116 95 131 176 205. 32 34 25 28 28 24 34 26 27 20. 81 79 85 84 82 85 84 90 65 98. 51 47 31 42 31 26 28 35 49 53. 63 56 49 56 70 63 70 63 70 56. 1333 1171 805 886 928 736 929 919 1C 02 1404. 1 1 2 2. x. 162.2. 27.8 83.3 17.1. 39.3. 61.6 1011.3. %.. c.v. V.I.. 6.23 34.61.
(5) Yukio Akahira, Darvin Beig and Warwick E. Kerr. Fifteen species were collected on the expedition to Manaus in 1963, and the other seventeen species were reared in natural nests collected from neighbouring in Rio Claro, Brazil. With the Neotropical and Tropical stmgless bee species was included one Ethiopian Trigona. boccandei) and two subspecies of Apis {mellifera ligiistica and mellifera adansonii). The list below shows the materials examined, and in the following description we will use the abbreviations, instead of the full scientific name of each species. Genera & Subgenera. Species & Subspecies. Localities Abbreviations. Cephalotrigona. femorata Smith. Manaus. Trigona (Trigona). cilipes cilipes (Fabriclus). Manaus. crassipes (Fabricius). Manaus. hyalinata bruneri Cockerell. Manaus. spinipes (Fabricius). Rio Claro. T. (Tefragona). dorsalis Smith. Manaus. T. (Frieseomelitta). flavlcornis (Fabricius). IVIanaus. f-reiremaiai (Moure). Guarapari. CF TCC TCr. THB TS TDo. TF1 TF. TVG. van a geneoscantha Moure. Manaus. ghilianii (Spinola) jaty Smith. Manaus. T. {Ptilotrigona). lurida Smith. M.anaus. T. {Meliponula). bocandei (Spinola). Luanda, Angola. Scaura {Scaura). tenuis (Ducke). Manaus. Plebeia {Plebeia}. droryana Friese. Rio Claro. ST TD. P. (Friesella). schrottki Friese. Rio Claro. TSc. cnpira Smith. Rio Claro. nigrior (Cockerell). Cristalina. TC TN. T. (Duckeola) T. (Tetragonlscd). Partamona {Pwtomond). Rio Claro. pearsoni (Schwarz). Cristalina. testacea testacea Cockerell. Manaus. Nannotrigona {Nannotrigona). testaceicornis (Lepeletier). Rio Claro. N. {Scaptotrigonci). bipimctata (Lepeletier). Rio Claro. Melipona. postica Latreille. Piracicaba. xanthotricha (Moure). Cabreuva. compressipes manaosensis Schwarz. Manaus. marglnata amazonlca Schulz. Manaus. marginata marginata Lepeletier. M^anaus. pseudocentris psezidocentris Cockerell. Manaus. quadrifasciata anthidioides Lepeletier. Rio Claro. quinqnefasciata Lepeletier. Cristalina. semlnigra merrillae Cockerell. Manaus. TG. TJ. TL. MB. TPe. TTT TT TB TP TX. MCM. MMA MMM. MPP. MQA MQ MSM. mellifera adansonli Latreille Johanesburg, South Afric£ i AMA. Apis. mellifera ligusfica Spinola. Imported from U.S.A.. AML. The fresh heads were submerged and dissected in a \% Ringer's solution and the brains with corpora allata were immediately transferee! to Bouin's fixative. The materials obtained on the expedition were fixed in Kahle's solution, and the dissected brains were also refixed in Bouin's solution. They were cut sagittaly into 7 p. thick sections and stained with Delafield's haematoxylin and eosin. The volumes of corpora allata were calculated by following formula.. (25).
(6) Corpora Allata in Brazilian Stingless Bees. V(^77rS(AnXBn) A and B ... The longest and shortest diameters, in p., of corpora allata presented in each serial section respectively. n... ..... Number of sections in each corpora allata.. The corpora allata are spherlcal or ellipsoid in general, therefore, the sections show circular or oval forms. But when the corpora allata are flattened into the extremely elongated or irregular ellipsoids (because of their natural characters or artifacts of fixation) the formula above used does not give the exact value, and we have applied it only for approxicate comparison in such cases.. The number of nuclei in each section of corpora allata was calculated under the oil immersion of microscope; the total number was obtained by summing for all sections, but when the one nucleus appeared in two sections, for example, we discounted one from the total number.. We found that the nuclei of corpora allata may be divided into two types of different sizes. We calculated the percentage of large nuclei {%). Based on the series of corpora allata for each species we calculated the mean cell volume (C. V.) appertalning to each nucleus by dividing the total volume by the total number of nuclei. To compare the volume of corpora allata with the size of insect we measured the width of thorax (in mm, distance between the bases of both fore-wings) and divided the mean volume by the cube of this distance. The quantity so obtained we denote V.I. in Table 1.. Results GENERAL REMARKS. The appearence of corpora allata is spherical or more or less elongated, situated at the dorso-lateral side of oesophagus. They are connected with the corpora cardiaca, which lie dorsally on the oesophagus as in the sections. The connections are at the posterior ends of corpora allata, and therefore sometimes sections of corpora allata appear comletely isolated from the corpora cardiaca. The corpora allata are covered by a very thin epithelial membrane with quite degenerate nuclei. The corpora allata contain fairly large cells of variable size, with the cell boundaries usually distinctly observable and polygonal. The nucleus contains several small nucleoli and the nucleoplasm is generally very clear, not rich in chromatin ; but degenerating nucleus has irregular shape and shows strong affinity for haematoxylin. When the corpora allata are extremely flattened, nuclei are also flattened and very rich in chromatin. The nuclei are distributed uniformly in the corpora allata, but sometimes they concentrate superficially or radially. The cytoplasm. is fine granular or fibrillar ; often it has many vacuoles of various sizes, especially in certain of the species examined. Though both corpora allata and corpora cardiaca are paired organs, we found an unpaired corpus allatum quite frequently in the sections. As we show later this phenomeon seems to be due to a degeneration in the course of worker bee life.. BIOMETRICAL OBSERVATIONS. Results are summarized in Table 1 and abbreviations. (2ff).
(7) Yukio Akahira, Darvin Beig and Warwick E. Kerr. are as follows : total number of nuclei in each corpus allatum (N); number of large nuclei (Nf) ; maximal width and height of serial sections (W) and (H) ; maxiaml distance antero-posterior (D); volume of each corpus allatum (V, Xl00/<3); serial number of the insect (P), indicating also the pairing or non-pairing of the corpora allata and the number of insects used ; the mean value (X); percentage of large nuclei {%}; cell volume (C.V.); and volume index (V.L). From the table we find the following. 1) In both N and V relatively large variations are observed throughout the all species examined ; specially in TCC, TCr, THB, TS, TF1, TVG, TJ, MB, TSc, TC, TN, TPe, TT, TB, TP, TX, MPP, MQA, AMA (queen), AM:L (queen and worker) the minimum volume of corpora allata was less than half of maximum. IVlaterials were obtained from the worker bees, and these variations may relate to the age of the worker in addition to individual variations. 2) Where N/ is the greater it might be suggested that the secretary activity of corpora allata is more marked. Though N/ shows also comparatively great variation in the same species, the following species give more than 30 % of {%}; TF1, ST, TC, TN, TPe, TTT, AMA (queen), AML (worker). On the other hand, in TS, MB, MMA, MQ, they are less than 10%, indicating a generally lesser development in M'elipona. 3) Cell volume appears to be correlated to the body size in the stingless bees ; a group of small body size, ST, TSc, TDo, and TCr, shows the average value less than 3 //3 per nucleus ; however, a group of large body size, CF, TG, MB, MCM, and MSIVt, it is more than 5 /J,3. But it is notable that some species having not such large body size show also great cell volume, more than 5 {^ : TF1, TF, TC, TN, TPe, TP, and TX. On the other hand the Melipona group which has relatively large body size shows small value less than 3 /J,3, MMA, MM1VT, MMP, MQA, and MQ. In the honey bee, the cell volume is extremely large compared to that in stingless bees. 4) V.I. indicates a correlation between volume of corpora allata and body size ; when the former is comparatively large V.I. becomes larger. In following species V.I. is less than 50 : CF, TCC, TCr, THE, TS, TL, MB, TPe, TT, TB, TP, MCM, MMA, MMM, MPP, MQA, MQ, and MSM. While the following species show more than 100 of V.I. : TF1, TF, TJ, AMA (queen), and AML (worker). Among them all, TF1 and TF, belonging to subgenus Frieseomelitta, have especially large values and MPP shows the minimum. value. In the comparison between queen and worker of two honeybee subspecies, a curious relation was found. The queen of AMA had a larger mean V and V.I. than the worker, but in AML the queen shows a lesser mean V and V.I. than the worker. 5) The tolumn of P indicates the individual bee and brain examined and the presence paired corpora allata. As mentioned above, sometimes corpora allata were not paired. For example, in CF, 10 corpora allata were observed ; the first 4 were paired and obtained from 2 brains, while the remaining 6 were singular ; in total we used 8 brains for 10 corpora allata. When one corpus allatum. was lacking in the partner, the pair of corpora cardiaca was very frequently perfect. Therefore it is unlikely that the missing corpus allatum had been lost due to dissection or in the process of microtechnique. We dissected and cut more than .10 brains in each species, excepting those collected in Manaus, of which number available was sometimes. (27).
(8) Corpora Allata in Brazilian Stingless Bees. dificient. The ratio of normal (paired) to degenerated (impaired) shown in Table 1 does not mean much because of the smallness of the samples but it does suggest an outline of the degeneration tendency in each species ; the species, in which more than 7 brains were necessary, are as follows: CF, TCr, TG, TC, TN, TP, TX, MPP, and MQA. The Partamona group shows a strong tendency to degeneration; in the Melipona group, MPP and MQA show a stronger tendency than the others. HISTOLOGICAL OBSERVATIONS. In Plates from I .to VI transverse sections of corpora allata are shown, all photographs being to the same enlargement for convenient of comparison (X 170 in low power, and X 1100 in high power). Often the maximal sections of the two corpora allata did not occur in the same cut; in these cases a montage has been made from the maximal left and right sections, but the two sides are always from the same brain (ex. Figures, 3-TCC, 5-TCr, 9-TS, 15-TF, 23-TL, 29-TD, 33-TC, 45-TP, 47-TX, 49-MCM, 53-. MMM, 57-MQA, and 61-MSM). With respect to the histological analysis, the following characters might relate to the functioning of corpora allata: a) the size of nucleus and its degenerating figure, b) quantity of chromatin and its distribution, c) cell boundery, d) the staining affinity of cytoplasm, and e) vacuolization of cytoplasm. We will now describe these characters in each species and abbreviate " corpora allata " by " c.a.". CF: Both c.a. and the large nuclei are flattened, often nuclei show polygonal forms, varying 11 to 13 p. in diameter. Nuclei are strongly stained and very rich in chromatin. Sell boundaries are not clear, staining affinity of cytoplasm is weak, and vacuoles appear rarely. TCC : C.a. are very flattened and irregular in form. Large nuclei are also elongated and very rich in chromatin and small degenerating ones stain quite darkly. The several nucleoli in large nuclei are mixed with much fine chromatin. Cell boundaries are obscure and no vacuoles exist.. TCr: C.a. are flattened. Chromatins are more or less rich and are distributed uniformly in the nucleoplasm. 2 to 3 sometimes 4 to 5 nucleoli are seen in the oval nuclei, the number is generally higher when the nuclei are smaller. The large nuclei are generally 5 to 7 p. in diameter, and the smalls less than 4//. Cytoplasm. stains weakly and cell boundaries and vacuoles are lacking. THB : C.a. are flattened. The large nuclei are remarkably numerous and measuring 6 to 8 p. in diameter, and chromatins are very abundant. Cell boundaries indistinct and vacuoles apparent.. TS : C.a. are oval. The nuclei vary in size extremely, large ones amount to 7 X 9 p. in oval form while the smalls are 2.5 {J. in diameter. Chromatin is rather less and well scattered in the clear nucleoplasm. Cell boundaries are indistinct, and several large vacuoles are observable in cytoplasm, of which the staining affinity is very weak. TDo: C.a. are oval, but nuclei are slightly elongated, measuring 9 to 11 p. in long axis, being large nuclei in high proportion. IVtuch chromatin and several nucleoli were contained inside the large nucleus. A few vacuoles appear in rare cases and c.ytoplasm has obscure cell boundaries.. (2S).
(9) Yukio Akahira, Darvin Beig and Warwick E. Kerr. TFl: C.a. are almost spherical. Oval and quite large nuclei, 7 X 9 p. in diameter, are extremely numerous. Their nucleoplasm are clear, somewhat poor in chromatin and include 2 or 3 nucleoli. The small nuclei are irregular and more or less rich in chromatin. Vacuolization proceeds strongly around the large nucleus and cell boundaries are distinct. Staining affinity of cytoplasm is weak. TF : Oval c.a. are always found as paired in the sections. Some large nuclei measuring 7 to 9 p. in diameter are rich in both nucleoli and chromatin, vacuolization is remarkable, cytoplasm hardly stains with eosin, and cell boundaries are quite clear; but the other large ones have contrary or negative features. The small nuclei tend to be irregular in their pycnosis. TVG: C.a. are almost flattened ; large nucleus with a few nucleoli is rich in chromatin, varying 7 to 9^ in diameter, while the small 3 to 5 p., Vacuolization does not occur and cell boundaries are obscure. Proportion of large nuclei is relatively high. TG: C.a. are spherical. Large nuclei show about 9 p. in diameter, having 2 to 3 nucleoli. The nucleoplasm is clear and not rich in chromatin. Small nuclei are slightly irregular in shape and show pycnotic figures. Vacuolization proceeds variable, some have much around the nucleus, while the others have not. Cell boundaries are not always distinct and cytoplasm shows moderate affinity to stain. TJ: C.a. are oval and the histological characters rather positive ; 2 to 4 large nucleoli and a considerable amount of chromatin; small vacuoles are recognized around the nuclei. The large nuclei vary in size 7 to 9 p. in diameter. Cell boundaries are quite distinct. But irregular, pycnotic, small nuclei are found scattered among the large nuclei. TL : C.a. and nuclei are flattened. The latter vary in size 7 to 9 p. in diameter. Nuclei stain strongly with haematoxylin and the small ones show completely pycnotic figures. There are no vacuoles, and cell boundaries are obscure.. MB : C.a. are oval, with large nuclei about 6 X 9 /-<, Often nuclei are elongated. They are rich in chromatin and have 2 to 4 nucleoli; small nuclei are darkly stained and show pycnotic figures. Active and large vacuoles are seen in the cytoplasm frequently; they are measured more than 10 p.. Cytoplasm is basophile, and cell boundaries are fairly clear. ST: Though samples are little, large nuclei are in high proportion and rich in chromatin. Vacuoles and cell boundaries are indistinct. TD : C.a. are small and round. Large nuclei are comparatively clear, measuring about 6 p. in diameter, while the small ones are darkly stained and irregular. A high proportion of large nuclei is seen. Affinity of cytoplasm for basOphile is strong, cell boundaries are distinct, and vacuolizatlon appears frequently around the nuclei. TSc: C.a. are oval, chromatin is more or less poor in nuclei, which vary 4 to 7 /-< in diameter. Vacuolizaton is rarely found. Cell boundaries are generally obscure but a few show distinctly. Staining affinity of cytoplasm is weak. TC : C.a. are spherical. Large oval nuclei are in high proportion measuring 9 to 12 p. in diameter. A few large nucleoli are distributed in each and chromatin is abundant. As regards cell boundaries and vacuolization a great variation is seen. Some (about half number examined) have positive characters, while the others are quite negative, these states being independent. (25).
(10) Corpora Allata in Brazilian Stingless Bees. of the presence of chromatin. Vacuoles are always found surrounding the nuclei; cytoplasm stains weakly with eosin. TN : The characters are quite similar to those of TC above, but vacuolization proceeds more strongly and the amount of chromatin is relatively small. TPe: C.a. are oval and again there are similarities with TC. A remarkable high proportion of large nuclei is seen, measuring 10 to 12 fi, vacuolization are very variable but cell boundaries are always distinct. TTT: C.a. and nuclei are very flattened. Large nuclei are seen in high percentage, measuring 9 to 11 [X in long axis. In these dark and rich chromatins are contained ; the smalls are irregular and crushed, or polygonal. Cell boundaries are obscure and vacuoles often appear. TT: C.a. are oval. The small nuclei are dark stained and irregular, showing pycnosis, while the large ones have a few nucleoli at the center. Chromatin is not rich and cell boundaries fairly distinct. Around the large nuclei minor degree of vacuolization occurs. TB : C.a. are oval. The large oval nuclei, measured 6x8^, are clear, not rich in chro-. matin and contain 2 to 3 nucleoli. Cell boundaries are relatively distinct and vacuolization proceeds actively around the nuclei. TP : C.a. are quite spherical, and the large nuclei vary 7 to 9 p. in diameter. A few nucleoli are visible while chromatin is poor. In some cases cell boundaries are distinct and less vacuoles exist in the cytoplasm, but cells of opposite character appear quite often. The nuclei tend to be arranged near the surface of the c.a. The small nuclei are irregular and dark stained. TX : C.a. are oval. The large nuclei are measured 6 to 8 /^ in diameter and are not rich in chromatin. Generally vacuolization takes place actively at the periphery of nuclei, sometimes one of the vacuoles becoming very large, about 15 p. in diameter, while cell boundaries are distinct. The small nuclei are irregular in shape and dark stained, measuring 2 to 3 p.. MCM: C.a. are slightly elongated. The large nuclei contain a few nucleoli and a great amount of chromatin, measuring about 11 p. in diameter. Vacuolization proceeds actively around the nuclei. Cell boundaries are quite obscure and cytoplasm stains strongly with eosin. MMA: C.a. are oval. The nuclei at the peripheral vary 7 to 9 /-( in diameter, and are very rich in chromatin. Occurrence of vacuoles is rare and cell boundaries are indistinct. MMM : C.a. are oval. The large nuclei are more or less rich in chromatin, being 7 to 9 p. in diametr, but they are scarce; the smalls are irregular in form, about 5 p., and are darkly stained. Vacuolization takes place strongly around the nucleus. Cytoplasm stains weakly and cell boundaries are indistinct. MPP: C.a. are very flattened; the nuclei have a large amount of chromatin and are strongly stained. They vary in size 7 to 9 p. in diameter. Vacuolization weak and cell boundaries obscure.. JVIQA: C.a. are oval. The large nuclei are measured 9 to 11 /-< in diameter, not rich in chromatin, and distribute near the suface of c.a. The small nuclei are sometimes arranged radially. They are about 4 p. and irregular. Cell boundaries are obscure and vacuolization appears frequently.. (30).
(11) Yukio Akahlra, Darvln Beig and Warwick E. Kerr. IVIQ : C.a. are elongated. The large nuclei have rather clear nucleoplasm, poor in chromatin. They are oval, about 8 X 11 iU, and contain one or two nucleoli. Often their long axes arrange parallel to that of the corpus allatum. The small ones occur in great numbers, are irregular and darkly stained, measuring 3 /-< in diameter. No vacuolization takes place and cell boundaries are indistinct. MSM[: C.a. are somewhat elongated. The round nuclei, both the large and the small, are rich in chromatin and contain a few nucleoli. The large ones vary in size 9 to 11 p. in diameter. Vacuolization proceeds intensely. Cell boundaries are obscure and staining affinity of cytoplasm is relatively strong. AMA : In both worker and queen bees, c.a. are quite spherical. The large nuclei appear in high proportion and are 12 to 14 p. in long axis, having much chromatins. The small ones are also darkly stained. Cytoplasm stains weakly and shows fine gramilar structure. Cell boundaries are often indistinct. Large vacuoles are commonly observable around the nuclei, especially the tendency is stronger in queen bee than in the worker. AML : Also in both castes c.a. are almost spherical and the large nuclei occur in high proportion. Chromatin is very rich in the worker but not in the queen. Cell boundaries are commonly distinct but in some cases of the queen they are rather indistinct. Cytoplasm stains strongly with eosin. To the contrary of AMA active vacuolization proceeds in the worker.. Discussion and conclusion Among the material that collected at Manaus and prefixed in Kahle's solution, for example, CF, TCC, TCr, THE, TDo, TVG, TL, ST, MCM, MMA, MPP, and MSM, the corpora allata showed flattened shape in a greater or lesser degree. But with the fresh material flattened or oval forms sometimes occur, for example, TS, TF1, TJ, TPe, TP and MMM. Therefore the general flattening in the Amazonian species may be an artifact of the prefixation, but may to some extent also be a real phenomena.. Although it is quite difficult to determine the degree of physiological activity of corpora allata, we select the following characters as being relevant conveniently and divide each into four classes.. A) Percentage of large nuclei in the total number ; when % increases activity of nuclei seems to be high. The 4 classes are : 1=less than 10%, 2= between 10 and 20°^, 3= between 20 and 30%, 4= more than 30%. B) Cell volume (C.V.); when it is large activity of the corpora allata seems to be high. Classes: l=less than 3 ^t3, 2=between 3 and 5 /J,3, 3=between 5 and 7 ^3, 4=more than 7/^3.. C) Volume index (V.I.); when it is large, the corpora allata are probably more active. Classes: l=less than 50, 2==between 50'and 75, 3=between 75 and 100, 4==more than 100. D) Quantity of cKromatin in large nucleus ; in general when chromatin is scarcely contained the nuclear activity seems to be high. Subjectively we have distinguished 4 classes : 1 == very large amount of chromatin, 2 = large amount of chromatin, 3 = quantity is a little, 4 = poor chromatin.. E) Vacuolization in cytoplasm ; when it takes place strongly the activity of cytoplasm seems. (37).
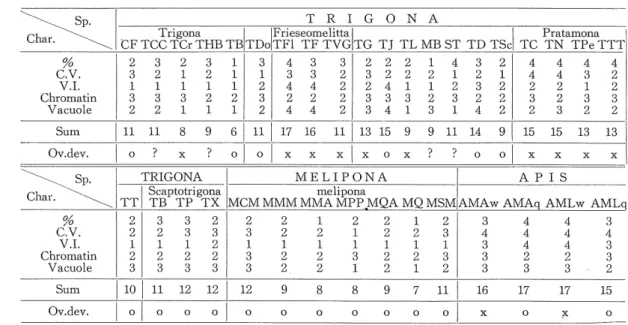
(12) Corpora Allata in Brazilian Stingless Bees. to be high. But according to Palm ('48) the occurrence of vacuoles might be an artifact, and in the same treatment of microtechnique and in the same species its occurrence frequently varies. Nevertheless, when we compare with the case of another gland, for example the pharyngeal gland, it seems the more possible that the vacuoles may have contained some secretion substance which has dissolved out in the solvent, such as alcohol or xylol. This character is divided into 4 classes subjectively: 1= no vacuoles, 2= a little amount of vacuoles, 3= active vacuoles, 4= very large amount of vacuoles.. In Table 2 these characters are summarized, however, it must be admitted that the classes are largely arbitrary and we do not even know which characters are the more important ; nevertheless we believe that the major tendencies in activity of the corpora allata are correctly indicated in this table. In addition the- table contains the another character that denotes various degree of ovary development in worker bees. Previously it has been shown that in some stingless bee species worker ovaries develop even in queenright colonies contrary to the case of Apis (Sakagami et al., '63); moreover, the curious fact has been noted that eggs laid by the workers serve as food for the mature queen (Sakagami & Zucchi, '63). On the other hand, in the other stingless bee species well developed ovaries in worker bees are never found. In Apis it has been observed that the worker ovarioles just after emergence are slightly more swollen than that in foragers (unpublished data). With respect to their development worker ovaries could be classified in three categories; a) worker ovaries do not develop at all (TCr, TF1, TF, TVG,. TG, TL, TN, TPe, and TTT, additionally Hypotrigona miillerii, examined by Sakagami); b) worker ovaries develop well (CF, TS, TDo, TJ, TD, TSc, TB, TP, TX and Melipona group,. in which are included of Melipona nigra schencki and Melipona rzifiventris); c) worker ovaries develop partially that condition is yet uncertain (TCC, THB, ST, and MB). These categories are denoted by " x ", " o ", and " ? " in Table 2 respectively. Table 2. Comparison between some histologlcal characters of corpora allata.. Char. %_. c.v. V.I.. Chromatin Vacuole. Sum Ov.dev.. la. Frieseomelitta. Pratamona. CF TCC TCr 1:HB 3 2 3 2 1 2 3 2 1 1 1 1 3 3 2 3 1 2 2 1. TDo TF1 TF TVGtTG TG TJ TL MB ST TD TSc TC TN TPeTTT FPe TTT rB|TDo|TFl. 1 1 1 2 1. 3 1 2 3 2. 4 3 4 2 4. 3 3 4 2 4. 3 2 2 2 2. 11. 6. 11. 17. 16. 11. 0. 11 ?. 8. 9. x. ?. 0. 0. x. FRIGONA. Sp.. TT|. LRF,. G 0 N A. T :R ]. Sp.. Scaptotrigona TB" TP TX. 2 3 2 2 3. 2 2 1 3 1. 1 2 1 2 3. 4 1 2 g 1. 3 2 3 2 4. 2 1 2 2 2. 4 4 2 3 2. 4 4 2 2 3. 4 3 1 3 2. 4 2 2 3 2. 13 15. 9. 9. 11. 14. 9. 15. 15. 13. 13. x. ?. ?. 0. 0. x. x. x. x. x. 0. MELIPONA MELI. AP IS. ma. MCM MMM MMA MPPMQA MQ MSM AMA^vA VlAq AMLw AMLq. Chromatin Vacuole. V.I.. 2.. 1 2 1 2 2. Sum. 10. 11. 12. 12. 12. 9. 8. ?. 9. 7. 11. 16. 17. 17. 15. Ov.dev.. 0. 0. 0. 0. 0. 0. 0. ). 0. 0. 0. x. 0. x. 0. _%.. 3 3 1 2 3. x. 2 2 4 3 4. 2 2 1 2 3. c.v.. 3 2 1 2 3. x. 2 3 2 3 3. 2 3 1 3 3. 2 2 1 2. (32). z L L 3 L. 2 2 1 2 2. 1 2 1 2 1. 2 3 1 3 2. 3 4 3 3 3. 4 4 4 2 3. 4 4 4 2 3. 3 4 3 3 2.
(13) Yukio Akahira, Darvin Beig and Warwick E. Kerr There are five subgenera each having three or more species. In Apis two subspecies are. contained : Trigona (TCC, TCr, THB, TS), Frieseomelitta (TF1, TF, TVG), Partamona (TC, TN, TPe, TTT), Scaptotrigona (TB, TP, TX), Melipona (MCM, MMM, MMA, MPP, MQA, MQ, MSM), and Apis (AMA, AML). 1. In general the members of each subgenus group tend to be similar, for example, in the Melipona group similarity seems closely through each of the characters studied, although MCM and M.SM are slightly excentric. 2. Among the groups, Apis and Frieseomelitta groups (TVG is exceptional) seem to have extremely high activities of corpora allata; following them Partamona group shows fairly high, and Scaptotrigona ensues. On the other hand, M-elipona and Trigona groups appear to have low activities of corpora allata. 3. When we compare the characters of corpora allata with the development of worker ovaries, the following hypothesis seems to be supported. There is inverse correlation between activity of corpora allata and development of worker o varies. If a species has a high activity of corpora allata worker ovaries do not develop, conversely in the case of low activity worker ovaries are found well developed. Cases conforming with the above hypothesis are as follows : Frieseomelitta group, Partamona group, Melipona group, Apis group, TS, TSc, and TT, amounting to 19 species. Contrary are the three species, TCr, TJ, and TD. There remains the group showing moderate activity of corpora allata, Scaptotrigona group, CF, TDo, and TG, amounting to 6 species, nevertheless those indicate quite typical development of worker ovarles, with exception of TG. It is interesting that in the Trigona group the relation between activities of corpora allata and worker ovaries is indistinct. Thus TCr and TS are opposite in ovary development although both have low levels of activity of corpora allata. Among the species whose ovary development is as yet unknown, the four speciea, TCC, THB, ST, and MB, have lesser activity of corpora allata than those Scaptotrigona group. On the basis of our hypothesis therefore we might risk the prediction that these will have developed ovaries. This will be resolved in future researches. It was established that the development of worker ovaries followed a mountain curve, that is, maximal development of worker ovaries occurs in the stage of nurse bee as in the case of Scaptotrigona postica (Sakagami et al., '63). Furthermore it has been observed that in this species showing typical development the ovaries begin to grow at 5 days after emergence and maximal development comes 10 and 14 days. Subsequently ovaries undergo gradual degeneration which corresponds closely to the differentiation of labour, i.e., nurse bees becoming foragers (Sakagami and Akahira, unpublished data). We used only foragers in present investigations of corpora allata and how the growth cycle of corpora allata corresponds to that of the ovaries remains for subsequent study. But this may turn out to be a correlation between corpora allata activity and worker ovary development. It is well known experimentally in other insects that the corpora allata produce a " juvenile hormone " tending to inhibit the realization of imaginal characters during development, while in the last immature stage the corpora allata become inactive and a newly hormonal balance influences metamorphosis (Wigglesworth, '56 ; Williams, '58 ; Thomsen, '40). In this field for. (33).
(14) Corpora Allata in Brazilian Stingless Bees the stingless bees nothing has yet been established experimentally, but in those species belonging to the type having undeveloped worker ovaries it seems to be possible 'that the active corpora allata may continue to secrete something like " juvenile hormone " and that the ovary development of workers may be inhibited by it. On the other hand, the so-called " ectohormone " or queen substance of Apis, which may be an inhibiting substance to ovary development of adult honey workers, has special effects on the development of worker ovaries of stingless bees (Akahira, Sakagami and Kerr, in preparation). Thus though the hormonal controls on ovary development have not received much attention in stingless bees, they will certainly prove one of the important factors in the complex interaction of social life in this group of Hymenoptera. Summary 1. A comparative study of corpora allata in thirty-two species of stingless bees and two subspecies of honey bees has been made in the fields of biometry (size, number of nuclei, volume, etc.) and of histology (quantity of chromatin, staining affinity, vacuolization, etc.). 2. These are found to be common characters within each subgenus, while considerable differences are found between them. In general, Apis, Frieseomelitta and Partamona groups shows positive qualities, probably indicating secretary activity; the Scaptotrigona group occupies a moderate position; and the Melipona and the T-rigona groups shows extremely low or negative characters. 3. The correlations between activities of corpora allata and the states of ovary development are discussed.. Literature cited Akahira, Y. Development of worker ovaries in some Amazonian stingless bees (in preparation). Akahira, Y., S. F. Sakagami & W. E. Ken- Effects of queen-substance of Apis onto the development of worker-ovaries in Trigona {Scaptotrigond) postlca (in preparation). Brandenburg, J. 1956. Das Endokrine System des Kopfes von Andrena vaga pz. (Ins. Hymenopt.) und Wirkung der Stylopisation (Stylops, Ins. Strepsipt). Z. Morph. u. Oekol. Tiere 45: 343-364. Hanstrom, B.1942. Die Corpora cardiaca und Corpora allata der Insekten. Biol. gen. 15 : 485-531. Moure, J.S. 1961. A preliminary supra-specific classification of the old world Meliponine bees (Hym., Apoidea). Studia Ent. 4 : 181-242. Mussbichler, A. 1952. Die Bedeutung ausserer Einflusse und der Corpora allata bei der Afterweiselentstehung van Apis mellifica, Z. vergl. Physiol. 34: 207-221. Nabert, A. 1913. Die Corpora allata der Insekten. Z. wiss.Zool. 104 : 181-358. Novak, V. J. A. 1954. Growth of the corpora allata during the postembryonal development in insects. Acta Soc. Zool. Boheneoslovenicae 18 : 98-133. Palm, N. B. 1948. Normal and pathological histology of the ovaries in Bombus Latr. Opusc. Ent.Suppl. 7 : 1-1,01.. Pflugfelder, 0. 1948. Volumetrische Untersuchungen an den Corpora allata der Honigbiene, Apis mellifica L. Biol. Zbl. 67 : 223-241. Sakagami, S. F., D. Beig, R.Zucchi & Y.Akahira 1963. Occurrence of ovary-developed workers in queenright colonies of stingless bees. Rev. Brasil.Biol. 23 : 115-129. Sakagami, S. F. & R. Zucchi 1963. Oviposition process in a stingless bee, Trlgona (Scaptotngond) postica Latr. (Hym.). Stadia Ent. 6 : 497-510. Thomsen, E. 1940. Relation between corpus allatum and ovaries in adult flies. Nature, Lond. 145 : 28-29. Wigglesworth, V. B. 1954. The physiology of insect metamorphosis. Cambridge Univ. Press. Williams, C. M. 1958. The juvenile hormone I. Endocrine activity of the corpora allata of the adult Cecropia silkworm. Biol.Bull.H6: 323-338.. (34).
(15) Yukio Akahira, Darvin Beig and Warwick E. Kerr. Explanation of plates All photography are transversal sections of corpora allata and abbreviations in parenthesis correspond to the text. Plate I. Fig.. Cephalotrlgona femorata (CF), x 170.. Fig.. Ditto, x 1100.. Fig.. Trigona cilipes (TCC), x 170.. Fig.. Ditto, x 1100.. Fig.. Trigona crassipes (TCr), x 170.. Fig.. Ditto, x 1100.. Fig.. Trigona hyallnata bmneri (THB), x 170.. Fig. 8.. Ditto, x 1100.. Fig. 9.. Trigona spinipes (TS), x 170.. Fig. 10.. Ditto, x 1100.. Fig. 11.. Tetragona dorsalis (TDo), x 170.. Fig. 12.. Ditto, x 1100.. Plate II. Fig. 13.. Frieseomelitta flavicornis (TF1), x 170,. Fig. 14.. Ditto, x 1100.. Fig. 15.. Fnesecnnelitta freiremaiai (TF), x 170.. Fig. 16.. Ditto, x 1100.. Fig. 17.. Frieseoinelitta varia geneoscantha (TVG), x 170.. Fig. 18.. Ditto, x 1100,. Fig. 19.. Duckeola ghilianii (TG), x 170,. Fig. 20.. Ditto, x 110,0.. Fig. 21.. Tetragonisca jaty (TJ), x 170.. Fig. 22.. Ditto, x 1100.. Fig. 23.. Ptilotrigona lunda (TL), x 170.. Fig. 24.. Ditto, x 1100.. Fig. 25.. Meliponula bocandei (MB), x 170.. Fig. 26.. Ditto, x 1100.. Plate III. Fig. 27.. Scaura tennis (ST), x 170.. Fig. 28.. Ditto, x 1100.. Fig. 29.. Plebeia droryana (TD), x 170.. Fig. 30.. Ditto, x 1100.. Fig. 31.. Friesella schrottki (TSc), x 170.. Fig. 32.. Ditto, x 1100.. Fig. 33.. Pwtamona cupira (TC), x 170.. Fig. 34.. Ditto, x 1100.. Fig. 35,. Partamona nigrior (TN), x 170.. Fig. 36.. Ditto, x 1100.. Fig. 37.. Partamona pearsonii (TPe), x 170.. Fig. 38.. Ditto, x 1100.. Fig. 39.. Partamona testacea testacea (TTT), x 170,. Fig. 40.. Ditto, x 1100.. Plate IV. Fig, 41,. Nannotngona testaceicornis (TT), x 170.. Fig. 42.. Ditto, x 1100,. (35).
(16) Corpora Allata in Brazilian Stingless Bees Fig. 43. Scaptotrigona blpunctata (TB), x 170. Fig. 44. Ditto, x 1100.. Fig. 45. Scaptotrigona postica (TP), x 170. Fig. 46. Ditto, x 1100.. Fig. 47. Scaptotngona xanthotricha (TX), x 170. Fig. 48. Ditto, x 1100. Fig. 49. Melipona compressipes manaoensis (MCM.), x 170. Fig. 50. Ditto, x 1100.. Fig. 51. Melipona marginata amazonica (MMA), x 170. Fig. 52. Ditto, x 1100. Plate V.. Fig. 53. Melipona marginata marginata (MMM), x 170. Fig. 54. Ditto. X 1100.. Fig. 55. Melipona pseudocentris pseudocentris (MPP), x 170. Fig. 56. Ditto, x 1100.. Fig. 57. Melipona quadnfasclata anthidioides (MQA); x 170. Fig. 58. Ditto, x 1100.. Fig. 59. Melipona quinquefasciata (MQ), x 170. Fig. 60. Ditto. X 1100.. Fig. 61. Melipona seminigra merrillae (MSM), x 170. Fig. 62. Ditto, x 1100.. Plate VI. Fig. 63. Apis mellifera adansonii (AMA), worker, x 170. Fig. 64. Ditto, x 430. Fig. 65. Ditto, x 1100. Fig. 66. Apis wiellifera aclansonu (AMA), queen, x 170. Fig. 67. Ditto, x 430. Fig. 68. Ditto, x 1100.. Fig. 69. Apis mellifera lignstica (AML), worker, x 170. Fig. 70. Ditto, x 430. Fig. 71. Ditto. X 1100.. Fig. 72. Apis mellifera ligustica (AML), queen, x 170. Fig. 73. Ditto, x 430. Fig. 74. Ditto, X 1100.. (35).
(17) Yukio Akahira, Darvin Beig and Warwick E. Kerr. Plate I. 12. (37).
(18) Corpora Allata in Brazilian Stingless Bees. Plate II ..---y-r^ r%.^. v:'.» "-^«. (3S).
(19) Yuklo Akahira, Darvln Beig and Warwick E. Kerr. Plate III. G 39).
(20) Corpora Allata in Brazilian Stingless Bees. Plate IV. 52. 57. (40).
(21) Yukio Akahira, Darvin Belg and Warwick E. Kerr. Plate V. (4i).
(22) Corpora Allata in Brazilian Stingless Bees. Plate VI. £tH^-^'yv:-"'. ''^^4:-^^. l^^^(t.:^.^;. 'ff ' -^1f. £-S^^.^"% r ,lr'y/^w:^..' ^: •SS£-ff'^/^'. -5. ^' Hi. •f-<.^'?' ^ '<• ^JJL''-^^.. (42).
(23)
図
関連したドキュメント
The properties of limit periodic homoge- neous linear difference systems with respect to their almost periodic solutions are mentioned, e.g., in [9, 24].. This paper is divided
We present sufficient conditions for the existence of solutions to Neu- mann and periodic boundary-value problems for some class of quasilinear ordinary differential equations.. We
(The Elliott-Halberstam conjecture does allow one to take B = 2 in (1.39), and therefore leads to small improve- ments in Huxley’s results, which for r ≥ 2 are weaker than the result
An integral inequality is deduced from the negation of the geometrical condition in the bounded mountain pass theorem of Schechter, in a situation where this theorem does not
This article is devoted to establishing the global existence and uniqueness of a mild solution of the modified Navier-Stokes equations with a small initial data in the critical
Wro ´nski’s construction replaced by phase semantic completion. ASubL3, Crakow 06/11/06
It is known that quasi-continuity implies somewhat continuity but there exist somewhat continuous functions which are not quasi-continuous [4].. Thus from Theorem 1 it follows that
A real matrix with nonnegative entries and having a largest eigenvalue of multiplicity just one has a strictly positive left eigenvector (see the Appendix) if and only if there
