29
1
1.1
Shewanella
Shewanella
(Venkateswaran et al. 1999,
Hau and Gralnick 2007, Fredrickson et al. 2008, Rodionov et al. 2011) Shewanella
(
[DMSO] [TMAO] )
(Fe, Mn, Co, V )
(Nealson et al.1994, Liu et al. 2002, Hau et al. 2008, Carpentier et al. 2003) (Flynn et al. 2014)
(Hau and Gralnick 2007, Murphy and Saltikov 2009)
Shewanella ( )
(
; bioelectrochemical systems; BES)
Shewanella ( )
Shewanella
Shewanella PAS
(Fredrickson et al. 2008) Shewanella
Shewanella
1.2
Shewanella oneidensis MR-1
Shewanella oneidensis MR-1 Shewanella
MR-1
(Myers and Nealson 1988) (Heidelberg et al. 2002)
(extracellular electron transfer
pathway; EET )
(Ross et al. 2011) (Figure 1) MR-1 BES
1.3
MR-1
MR-1 EET (CymA,
STC/FccA, OmcA, MtrA, MtrB MtrC) (Shi et al. 2007) (Figure 1)
CymA CymA
(STC/FccA) Mtr
(OmcA, MtrA, MtrB, MtrC) OmcA MtrC
(Shi et al. 2006) MR-1 (Lower et al. 2007, Xiong et al. 2006) (Figure 2A)
(Marsili et al. 2008, Von et al. 2008) (Figure 2B) MR-1
(Pirbadian et al. 2014) (Figure 2C) 2
OmcA
(Coursolle et al. 2010) MtrC
(Mitchell et al. 2012)
1.4
mtr
MR-1 EET (OmcA, MtrA, MtrB, MtrC)
(omcA, mtrC, mtrA, mtrB; mtr ) MR-1
(Figure 3) mtr
(Myers and Myers 1997, Beliaev et al. 2005, Teal et al. 2006, Rosenbaum et al.
2012, Pirbadian et al. 2014) mtr cyclic AMP
(cAMP) receptor protein (CRP) cAMP (
, CyaC) (Saffarini et al. 2003, Charania et al. 2009)
in silico omcA CRP
cAMP MR-1 mtr
1.5
CRP/cAMP
CRP/cAMP CRP/cAMP
Escherichia coli
(Ullmann and Monod
1968, Zubay et al. 1970) MR-1 CRP
(Saffarini et al. 2003, Charania et al. 2009) CRP
cAMP CRP
cAMP E. coli cAMP
(Makman and Sutherland 1965)
Figure 2. MR-1 A. B. C. OM: PP: IM: Mox: Mred; CymA MtrB MtrA MtrC OmcA OmcA e -CymA MtrB MtrA MtrC OmcA OmcA e -Mred Mox MtrB MtrA MtrC OmcA MtrB MtrA MtrC OmcA MtrB MtrA MtrC OmcA Mt rB Mt rA Mt rC O m cA MtrB Mt rA MtrC OmcA Mt rB Mt rA Mt rC OmcA Mt rB Mt rA Mt rC O m cA MtrB MtrA MtrC OmcA
CymA CymA CymA CymA
Figure 3. mtr
omcA
mtrC
mtrA
mtrB
mtrF
2
mtr
2.1
mtr CRP mtr CRP2.2
2.2.1
Table 1 Table2 (Escherichia coli) LB (Luria-Bertani) 37ºC
WM6026 100 µg/ml
(DAP) BL21 (DE3) 2 × yeast
extrac-triptone (2 × YT) 37ºC 30ºC S. oneidensis LB
(Lactate medium; LM) (15 mM DL- , 9 mM (NH4)2SO4, 5.7 mM
K2HPO4, 3.3 mM KH2PO4, 5.0 g L-1 yeast extract, 30 mM HEPES-NaOH (pH 7.4)) 30ºC
600 nm DU800 spectrophotometer (Backman)
100 ml LM 300 mL OD600 = 0.05 S. oneidensis 180 rpm (OD600 > 2.0) 10 mM LM 80 ml 100 mL S. oneidensis OD600 = 0.01 30ºC (OD600 > 0.25) 100 µg/ml (Am), 50 µg/ml (Km), 15 µg/ml
(Gm) 1.5% Bacto agar (Difco)
2.2.2
RNA
S. oneidensis LM
5 ml 30 ml 5 4ºC 7,000 rpm
RNA Trizol (Invitrogen)
1 ml Trizol 0.2 ml
RNA RNeasy Mini kit (Qiagen) RNase-Free DNase set (Qiagen)
250 µl RNA 250 µl Buffer RLT (1% ) 250 µl
100% RNeasy Mini kit 350 µl
Buffer RW1 80 µl DNase RNA
DNA 350 µl Buffer RW1 500 µl Buffer
RPE 50 µl RNase-Free H2O RNA RNA Bioanalyzer
(Agilent Technologies)
2.2.3
5′ rapid amplification of cDNA ends
(5′-RACE) PCR 2
MR-1
10 µg total RNA SuperScriptIII reverse transcriptase (Invitrogen) IR800
PE-mtrC-100 (Table 2) complementally DNA (cDNA)
10 µg RNA, 770 µM dNTP, 4 pmol PE-mtrC-100 13 µl 65ºC
5 1 4 µl 5 × First-Stand buffer, 5 mM DTT,
40 U RNase OUT Recombinant RNase Inhibitor (Invitorgen), 200 U SuperScriptIII reverse
transciptase 20 µl
: 25ºC (5 ), 50ºC (60 ), 70 ºC (15
) cDNA /
cDNA
Li-Cor4200 automated DNA sequencer (Li-Cor)
5¢-RACE MR-1
total RNA 1.0 µg SMATer RACE cDNA amplification kit (Clontech)
cDNA omcA-RACE-out mtrC-RACE-out (Table 2)
cDNA 5¢ SMATer RACE cDNA Amplification kit
cDNA
omcA-RACE-out mtrC-RACE-out
omcA-RACE-in mtrC-RACE-in (Table 2) kit Universal Primer A mix
PCR PCR QIAquick PCR Purification kit (Qiagen)
PCR PCR 5 Buffer PB kit
750 µl Buffer PE 50 µl Buffer EB DNA PCR
T-vector pMD19 (TaKaRa) JM109
Am LB 2 ml 1 13,000 rpm
250 µl Buffer P1 (RNaseA ) 250 µl Buffer P2
350 µl Buffer N3 10 13,000 rpm
kit Buffer PB Buffer
PE 50 µl Buffer EB
5′
2.2.4
PCR (RT-PCR)
total RNA SuperScriptIII reverse transcriptase (Invitrogen) Rnadom Primers
(Invitrogen) cDNA 5.0 µg RNA, 770 µM dNTP, 250 µg
Random Primers (Invitrogen) 13 µl 65ºC 5
1 4 µl 5 × First-Stand buffer, 5 mM DTT, 40 U RNase OUT Recombinant
RNase Inhibitor (Invitorgen), 200 U SuperScriptIII reverse transciptase
20 µl : 25ºC (5 )
50ºC (60 ) 70ºC (15 )
cDNA DNA TaKaRa Ex Taq Polymerase (TaKaRa)
(Table 2) PCR PCR 5 µl 2.0%
σ ChemiDoc XRS (Bio Rad)
2.2.5
RT-PCR (qRT-PCR)
(16S rRNA , omcA, mtrC) DNA MR-1
total DNA PCR
PCR QIAEX II Gel Extraction kit (Qiagen)
1.5 ml
3 Buffer QX1 10 µl QIAEX II
QIAEX II DNA Buffer QX1 Buffer PE 20 µl TE
DNA Total RNA LM
15 ng total RNA, 1.3 µl 50 mM Mn(OAc)2, 7.5 µl LightCycler RNA Master SYBR
Green I (Roche), 0.15 µM (Table 2) LightCycler 1.5
instrument (Roche) qRT-PCR
61ºC (20 ), 95ºC (30 ), ; 95ºC (5 ), 60ºC (5
), 72ºC (25 ) 45 , 95ºC (0 ), 65ºC (15 ), 95ºC (0 ),
40ºC (30 ) PCR mRNA
3
2.2.6
MR-1 crp pSMV-10 2
(750 bp )
(750 bp ) DNA Table 2
Phusion High-Fidelity DNA polymerase (New England Biolabs) PCR
DNA pSMV-10 SpeI pSMV-crp WM6026 MR-1 (single-crossover ) Km LB LB 17 10% (w/v) LB Km (double-corssover ) PCR
2.2.7
LacZ
omcA mtrC lacZ (omcA_F-54 omcA_F-150 mtrC_F-42 mtrC_F-203) (omcA_R+93 mtrC_R+117) (Table 2) PCR PCRQIAquick PCR Purification kit (Qiagen) 2.2.3
PCR pMElacZ EcoRI HindIII EcoRI BamHI
pMElacZ
2.2.8
S. oneidensis LB S. oneidensis 2 ml 16,000 × g, 2 4ºC 1 ml 300 mM 2 100 µl 300 mM 50 µl 1.0–3.0 µg 0.1 cm (Bio Rad)5 Micropulser (Bio Rad) 180 kV
500 µl LB 1.5 mL
30ºC 2 100 µl Gm LB
2.2.9
LM
OD600 = 0.1 0.5 ml 0.5 ml
Z-buffer (60 mM Na2HPO4, 40 mM NaH2PO4, 2 mM KCl, 200 µl MgSO4, 5 mM
2-) 10 µl 0.1%SDS 15 µl 0.2 ml 4 mg/ml
2- -β-D- (ONPG)
0.5 ml 1M Na2CO3 4 mg/ml ONPG
1M Na2CO3 12,000 × g, 2
420 nm
-Miller unit (Miller, 1972) 3
Miller unit = (1000 × Abs420) / ( (ml) × OD600 × ( ))
2.2.10
His
CRP
N His-tag-CRP MR-1 total DNA
crp_NdeI_F crp_BamHI_R (Table 2) PCR crp
PCR QIAquick PCR Purification kit (Qiagen) 2.2.3
PCR N His pET-28a (+)
(Novagen) NdeI BamHI His-tag-CRP
(pET-crp; Table 1) E. coli BL21 (DE3) 300 mL
100 ml Km 2 × YT OD600 = 0.05 30ºC
180 rpm OD600 = 0.5–0.8
isopropyl-β-D-galactopyranoside (IPTG) 0.1 mM His-tag-CRP
3 50 ml
300 µl IMAC wash buffer (Bio-Nobile) 40 µl 360 µL
IMAC wash buffer 10 400 µl
(Misonix) QuickPick IMAC Metal Affinity
kit for proteins (Bio-Nobile) His-tag-CRP His-tag-CRP
Micro BCA Protein Assay kit (Thermo Fisher Science)
2.2.11
Cy3
DNA
Cy3 DNA 5′ Cy3 (Table
2) PCR PCR
2.2.12
(EMSA)
2 nM Cy3 DNA 0–200 µg His-tag-CRP (10 mM
Tris-HCl (pH 7.6), 0.5 mM EDTA (pH 8.0), 100 mM KCl, 50 µg/ml , 50
µg/ml poly (deoxyinosinic-deoxycytidylic) acid [poly (dI-dC) ], 0–50 µM cAMP, 10%
) 20 µl 30 12.5%
0.5 × Tris-borate-EDTA
150 V 30 Typhoon FLA 9000 (GE Healthcare)
2.2.13
DNA
5′ DNA DNA- (Table
2) 2 nmol
(10 mM Tris-HCl (pH 7.5), 0.1 M NaCl, 1 mM EDTA)
100 ml 65ºC 15
4 DNA
Streptavidin Sepharose High Performance (GE Healthcare) (20 mM HEPES
(pH 7.4), 100 mM KCl, 0.5 mM EDTA (pH 8.0), 0.5 mM (DTT), 50 µg/ml
BSA, 10% ) 3
50% 100 µl 50% 100 µl
2 DNA 900 µl 2
DNA 3
1 ml 1 × Blocking Reagent (Roche)
16 DNA
3 100 µl
2.2.14
MR-1 10 mM LM
, ( ; 5 ml, ; 50 ml) 7,000 g, 5 4ºC
300 µl IMAC wash buffer (Bio-Nobile)
IMAC wash buffer 10 (Misonix) 10,000 g
Micro BCA Protein Assay kit (Thermo Fisher
10 µl (200 µg ) 100 µl DNA 900 µl
2 3
25 µl 2 × SDS-Buffer 65ºC 15
20 µl
12.5% (E-T12.5L, ATTO) 20 µl
SDS-PAGE buffer (AE-1410 EzRun, ATTO) 200 mA
AE-1360 EzStrain Silver (ATTO)
500 ml 100 ml (40 ml 50 ml 10 ml 1 ml S-1 ) 10 100 ml 30% 10 100 ml 10 100 ml (100 ml 1 ml S-2 ) 5 30 100 ml (100 ml 0.5 ml S-3 0.5 ml S-4 ) 30 100 ml 100 ml (100 ml 1 ml ) 10 100 ml 2 ChemiDoc XRS (BIORAD)
2.3
2.3.1
mtr 10 mM LM MR-1RNA RT-PCR (Figure 1) mtrF omcA
(Figure 1B, lane 1) omcA-mtrC mtrC-mtrB
omcA mtrB (Figure 1B, lane 3,
4).
2.3.2
omcA mtrC (transcription start site;
TSS) (Figure 2) (Shao et al. 2014, Beliaev et al. 2001). Shao
et al. (2014) 5′ end sequencing omcA
93 bp (TSSomcA; Figure 2A) Beliaev
et al. (2001) 5′-RACE mtrC ATG 119 bp TSS
Beliaev 10 mM MR-1 5′-RACE mtr TSS TSSomcA, TSSmtrC (Figure 2) mtr TSS TSSmtrC (Figure 3) mtrA mtrB 5′-RACE TSSmtrC
2.3.2
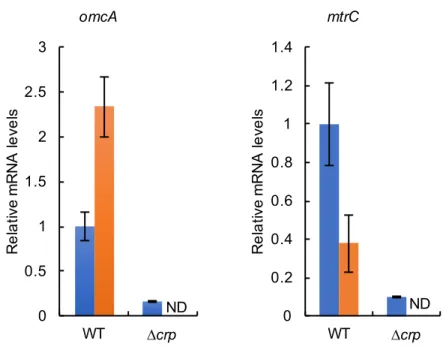
5′-RACE mtr 2 TSS (TSSomcA, TSSmtrC) omcA mtrCAB 10 mM MR-1 RNA omcA mtrC qRT-PCR (Figure 4) omcA 2.3 mtrC 2.7 omcA mtrC2.3.3
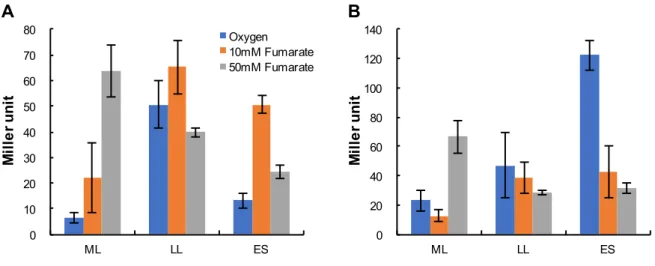
omcA mtrC (PomcA PmtrC) (Table 1; pMEomcA-150, pMEmtrC-203) MR-1 MR-1 10 mM [ (ML), (LL),(ES)] (Figure 5) PomcA
(Wang et al. 2013) 15 mM 10 mM , 50 mM PomcA PmtrC ML (Figure 5A, B) 10 mM ES 50 mM ES ES PomcA PmtrC PomcA PmtrC PomcA ES PomcA
2.3.4
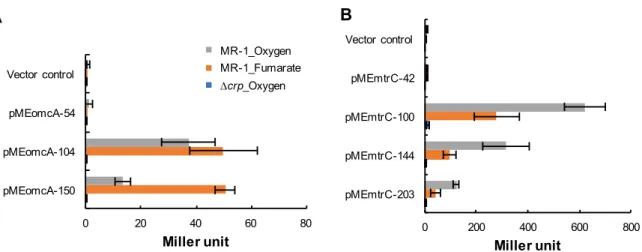
omcA mtrC 5′ (Table 1; pMEomcA-54,pMEomcA-104, pMEmtrC-42, pMEmtrC-100, pMEmtrC-144) MR-1
(Figure 6) 10 mM
LacZ
PomcA 5′ TSSomcA
-104 -54
(Figure 6A) -104 -54 PomcA
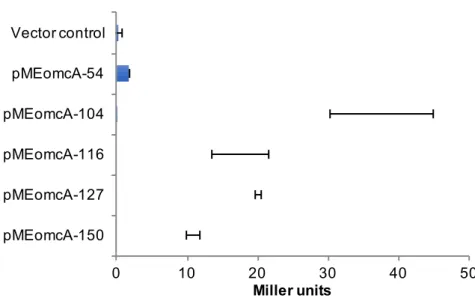
-150 -104 2 -150 -104 PomcA PomcA 5′ (Figure 7) 5′
(Table 1; pMEomcA-69, pMEomcA-84, pMEomcA-95, pMEomcA-116,
pMEomcA-127) MR-1
PomcA PomcA
(Figure 7) -104 -116 PomcA (5′-AAAAGGAACTCAGATGATGCTGTT-3′; TSSomcA -90 -113 ) (Figure 2A) PmtrC 5′ TSSmtrC -100 -42 (Figure 6B) -100 -42 PmtrC -203 -144, -144 -100 PmtrC TSSmtrC -203 -100 PmtrC
2.3.5
∆crp
omcA
mtrC
5′ PomcA PmtrC PomcA PmtrC (Figure 2) PomcACRP (Gao et al. 2010) PomcA
PmtrC CRP ∆crp (WT) (Figure 6) ∆crp (Saffarini et al. 2003) ∆crp ∆crp ∆crp omcA, mtrC ATG pMEomcA-150 pMEmtrC-203 WT ∆crp omcA mtrC qRT-PCR WT (Figure 3) PomcA PmtrC CRP
2.3.6
P
omcAP
mtrCCRP
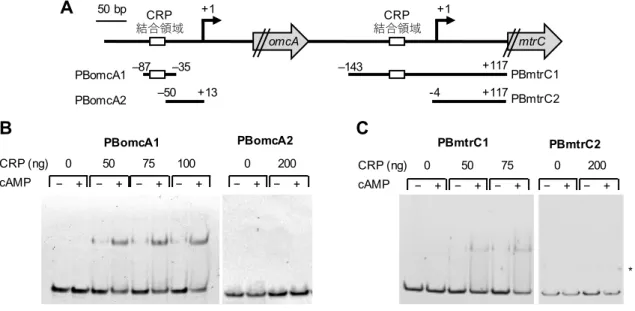
TSSomcA TSSmtrC CRP(electrophoretic mobility shift assay; EMSA) (Figure 8)
PomcA CRP TSSomcA -87 -35 (PBomcA1; Figure 8A)
PBomcA1 DNA CRP PBomcA2
(Figure 8B) PomcA CRP
CRP cAMP DNA
cAMP CRP PBomcA1 (Figure 8B)
CRP PomcA cAMP
PmtrC CRP CRP DNA
TSSmtrC -143 +117 (PBmtrC1; Figure 8A) -4 +117 (PBmtrC2; Figure 8A)
TSSmtrC PBmtrC1 PBmtrC2 2 DNA CRP CRP PBmtrC1 CRP PBmtrC2 (Figure 8C) cAMP DNA CRP PBmtrC1 cAMP (Figure 8C) CRP PomcA PmtrC CRP PomcA PmtrC cAMP PomcA PmtrC CRP
2.3.7
DNA
PomcA (Figure 7)(5′-AAAAGGAACTCAGATGATGCTGTT-3′; TSSomcA -90_-113) (Figure 2A)
PomcA PomcA
DNA
DNA CRP -54 -84
(PAomcA1) -85 -116 (PAomcA2) CRP
(PAomcA3) 3
(Figure 9A) DNA MR-1
SDS-PAGE (Figure 9B)
PAomcA3 20-23 kDa
(Figure 9B) PAomcA3 MR-1
MALDI TOF-MS PomcA PAomcA3 CRP PAomcA1 PAomcA2
2.4
mtr 2 (PomcA, PmtrC) CRP Arc Fur mtr(Saffarini et al. 2003, Yang et al. 2008) mtr
mtr 2
CRP mtr CRP
CRP
cAMP DNA (Brückner
et al. 2002, Stülke et al. 1999) S. oneidensis MR-1 CRP mtr
(Charania et al. 2009) Shewanella
CRP E. coli S. oneidensis MR-1
cAMP
qRT-PCR ∆crp PomcA PmtrC
WT (Figure 4, Figure 6) TSSomcA
TSSmtrC 5′ PomcA PmtrC EMSA CRP CRP PomcA PmtrC CRP E. coli K12 CRP cAMP DNA DNA CRP
(Kolb et al. 1993, Botsford and Harman 1992) CRP -10 box -35 box
(5′-TGTGA-6N-TCACAA-3′) RNA
(Kolb et al. 1993, Botsford and Harman 1992, Busby and Ebright
1997) TSSomcA TSSmtrC TSS -61.5 -60.5
CRP class I CRP (Joung
et al. 1993, Tebbutt et al. 2002) PomcA PmtrC -35 box
-10 box E. coli (5′-TATAAT-3′; Kolb et al. 1993, Raibaud and
Schwartz 1984) E. coli CRP -35 box S. oneidensis MR-1 CRP E. coli MR-1 cAMP (Saffarini et al. 2003) MR-1 cAMP E. coli cAMP S. oneidensis MR-1 cAMP PomcA PmtrC CRP cAMP 10 mM PomcA PmtrC (Figure 5) 10 mM 15 mM PomcA PmtrC (Figure 5) mtr 50 mM PomcA PmtrC mtr (Wang et al. 2013) PomcA PmtrC 50 mM 10 mM (Figure 5) PomcA PmtrC (Figure 5) mtr
(Beliaev et al. 2005, Teal et al. 2006) PmtrC
(Figure 5B) PmtrC 50 mM PmtrC PomcA (Figure 5A) PomcA PmtrC PomcA PmtrC CRP PomcA PmtrC PomcA TSSomcA -116 -104 5′ TSSomcA -116 CRP 39 E. coli CytR CRP
PomcA (Kristensen et al. 1997,
Valentin-Hansen et al. 1996) Shewanella
omcA CRP (Figure 9) CRP CRP DNA 5′ PmtrC mtrC 5′ PmtrC (Figure 6B) AT E. coli AT H-NS AT PmtrC mtr Figure 1A PomcA PmtrC CRP PomcA (Figure 9) Shewanella EET
PomcA PmtrC (Figure 4 ) Shewanella PAS
mtr
MR-1 OM-cyt
Figure 2. omcA (A) mtrC (B) TSSomcA TSSmtrC (+1) omcA
mtrC TSS
CRP -10 -35 GeSTer program (Mitra et al. 2011)
Figure 5. - PomcA (A) PmtrC (B)
( ) 10 mM ( ) 50 mM ( ) pMEomcA-150
pMEmtrC-203 MR-1 LacZ (ML)
(LL), (ES) 3
(pMElacZ)
MR-1 LacZ 3 Miller units
Table 1.
Strain or plasmid Relevant characteristic Source or reference
E. coli strains
DH5α F-, φ 80dlacZ∆M15, ∆(lacZYA-argF)U169, deoR, recA1, endA1,
hsdR17(rK-, mK+), phoA, supE44, λ-, thi-1, gyrA96, relA1
Takara
JM109 recA1. endAl, gyrA96, thi. hsdR17, supE44, relA1, λ-,
∆(lac-proAB), [F’, traD36, proAB, lacIq Z∆M15] Yanisch-Perron et al. 1985 JM109λpir JM109 lysogenized with λpir Penfold and
Pemberton, 1992 WM6026 lacIq, rrnB3, DElacZ4787, hsdR514, DE(araBAD)567,
E(rhaBAD)568, rph-1, att-lambda::pAE12-del(oriR6K-cat::frt5),
DE(endA)::frt, uidA(delMluI)::pir(wt), attHK::pJK1006-del1/2 (deloriR6K-cat::frt5, deltrfA::frt)
William Metcalf, University of Illinois BL21 (DE3) F ompT hsdR17 (rB mB+) gal dcm(DE3) F , ompT, hsdSB (rB
mB ), gal(λcI 857, ind1, sam7, nin5, lacUV5-T7gene1), dcm(DE3)
Novagen
S. oneidensis strains
MR-1 Wild type ATCC
∆crp SO_0624 (crp) disrupted This study
∆cpdA SO_3901 (cpdA) disrupted This study
∆cyaC SO_1329 (cyaC) disrupted This study
∆cpdA∆crp SO_3901 (cpdA), SO_0624 (crp) disrupted This study ∆cpdA∆cyaABC SO_3901 (cpdA), SO_4312 (cyaA), SO_3778 (cyaB), SO_1329
(cyaC) disrupted
This study
∆ldhA SO_ (ldhA) disrupted Nakagawa et
al. 2015
Plasmids
pET-28(a) Expression vector, T7 promoter Novagen pET-crp pET-28(a) containing N terminal His-tag-crp This study pSMV-10 9.1 kb mobilizable suicide vector; oriR6K, mobRP4, sacB, Kmr,
Gmr Chad Saltikov, California Inst.
of Tech. pSMV-crp 1.6 kb fusion PCR fragment containing ∆crp cloned into the SpeI
site of pSMV-10
This study
pSMV-cpdA 1.6 kb fusion PCR fragment containing ∆cpdA cloned into the SpeI site of pSMV-10
This study
pSMV-cyaA 1.6 kb fusion PCR fragment containing ∆cyaA cloned into the SpeI site of pSMV-10
This study
pSMV-cyaB 1.6 kb fusion PCR fragment containing ∆cyaB cloned into the SpeI site of pSMV-10
This study
pSMV-cyaC 1.6 kb fusion PCR fragment containing ∆cyaC cloned into the SpeI site of pSMV-10
This study
pMElacZ pME4510 derivative, lacZ Gmr Endoh et al. 2003
Table 1. ( )
Strain or plasmid Relevant characteristic Source or reference pMEmtrC-203 pMElacZ containing region from -203 to +116 relative to TSSmtrC This study pMElldP+1 pMElacZ containing region from +1 to +192 relative to TSSlldP This study pMElldP-60 pMElacZ containing region from -60 to +192 relative to TSSlldP This study pMElldP-182 pMElacZ containing region from -182 to +192 relative to TSSlldP This study pMElldP-541 pMElacZ containing region from -541 to +192 relative to TSSlldP This study pBBR1MCS-2 Broad-host-range vector, lacZ promoter, Kmr Kovach et al.
1995
pBBR1MCS-5 Broad-host-range vector, lacZ promoter, Gmr Kovach et al. 1995
pBBRcrp pBBR1MCS-2 containing crp This study
pBBRdld pBBR1MCS-2 containing dld This study
pBBRcyaC pBBR1MCS-2 containing cyaC This study
Table 2.
Primer Sequence (5′–3′) For use
cpdA_FO CGTTTCTACTTTTATTGGGC cpdA disruption
cpdA_5_OUT GATCACTAGTGATTCATCACAGGCACAAGG cpdA disruption
cpdA_5_IN AACTTTGGTACCGATCACTTTCAGCACATTGAATACCC cpdA disruption
cpdA_3_IN GTGATCGGTACCAAAGTTGGTTATTAACGCGAATGACC cpdA disruption
cpdA_3_OUT GATCACTAGTCCTAAATGGTTAGGTCGGG cpdA disruption
cpdA_RO GCTTCATCGACACTGTTATC cpdA disruption
cyaA_FO CTAATAATGCTTTAACGCGC cyaA disruption
cyaA_5_OUT GATCACTAGTGAGATAAAGCTGGCGATACG cyaA disruption
cyaA_5_IN GGTAGCGTTGCTCACGCTATCCATCCTAGAACACAACAC cyaA disruption
cyaA_3_IN AGCGTGAGCAACGCTACCTTTTAAGCGGGAGCTACCTC cyaA disruption
cyaA_3_OUT GATCACTAGTGTACAGTGGCGTACCATAGG cyaA disruption
cyaA_RO CGCATTATTAAAGGCATGCC cyaA disruption
cyaB_FO GATCTTAAGGCGATCTCAGG cyaB disruption
cyaB_5_OUT GATCACTAGTACTGCAGATCCACGATATCC cyaB disruption
cyaB_5_IN CTGAGCCACGATTGAGTTATTCATCAACAATCTCCATTTTTAC cyaB disruption
cyaB_3_IN AACTCAATCGTGGCTCAGCAATAATCGCTTTGCTGGG cyaB disruption
cyaB_3_OUT GGGATCACTAGTCGTAATCGCCACCATTCC cyaB disruption
cyaB_RO AACCTTATTTGCACAGGTCG cyaB disruption
cyaC_FO CCCCTCAAAATCTGAGCG cyaC disruption
cyaC_5_OUT GATCACTAGTGCGCTAATATGATGGCAGG cyaC disruption
cyaC_5_IN AGCGCCGTTGGTACCTGATGCTCTCACAACGGGTTTTG cyaC disruption
cyaC_3_IN TCAGGTACCAACGGCGCTCCTTTATAAGACTCACAACAGG cyaC disruption
cyaC_3_OUT GATCACTAGTCGGTGAAGAAGTCGTACAG cyaC disruption
cyaC_RO CAGTTGATTGAACGTGACC cyaC disruption
crp_5_OUT GGACTAGTGGTCACGTTTAATCAACTGGCC crp disruption
crp_5_IN GGTAGCGTTGCTCACGCTCAGAGCCATGTCGATGTTCCTCG crp disruption
crp_3_IN AGCGTGAGCAACGCTACCCGTTAAGTTAGACTTCAGCTTG crp disruption
crp_3_OUT GGACTAGTGGGTTTACCTAAATAATCATCGG crp disruption
crp_RO GGCCATGGGCACGATGAATAAGTGC crp disruption
dld_F_BamHI CGCGGATCCACCATCTTGTTCGAAGTCAC dld complementation
dld_F_XbaI CTAGTCTAGACTTCTAAAGAAAAACGGGGC dld complementation
qRT-16S-F AGCGCAACCCCTATCCTTAT qRT-PCR for 16S rRNA
gene
qRT-16S-R CGTAAGGGCCATGATGACTT qRT-PCR for 16S rRNA
gene
qRT-omcA-F GGATACGGCGTTGAAGATGT qRT-PCR for omcA
qRT-omcA-R TGGTATCCGTTCCATTCCAT qRT-PCR for omcA
Table 2. ( )
Primer Sequence (5′–3′) For use
qRT-mtrC-R GCTTCGTTAGTGGCGAAAAC qRT-PCR for mtrC qRT-lldP-F GGCGTTTCATGCAAGTC qRT-PCR for lldP qRT-lldP-R ATCGACGCAAGACCTGC qRT-PCR for lldP qRT-dld-F CATCGGCACTCAACTTCTCA qRT-PCR for dld qRT-dld-R CGCAGGTATCAATCACATCG qRT-PCR for dld qRT-lldF-F CGACCTACAGCGCCTTCTAC qRT-PCR for lldF qRT-lldF-R AGTGTTAAGGCAGCCACCAC qRT-PCR for lldF
qRT-ldhA-F GACCCCTACCCTAATCCAGC qRT-PCR for ldhA
qRTldhA-R GACCCCTACCCTAATCCAGC qRT-PCR for ldhA
crp_NdeI_F CATGCATATGATGGCTCTGATTGGTAAGCC pET-crp
crp_BamHI_R GATCGGATCCTTAACGGGTACCATATACCAC pET-crp
PE-mtrC-100 CCATCACTACCATCATTGCC Primer extension
RT-omcA-mtrF-F TAACAATGGCACCTTCACCA RT-PCR RT-omcA-mtrF-R TCGCCACCTTTATGGATAGC RT-PCR RT-omcA-F CCATCAGGCGTTGATAACCT RT-PCR RT-omcA-R ATACCCAAATTACGGCACCA RT-PCR RT-omcA-mtrC-F GAAAGCCCACGAAAGTGAAG RT-PCR RT-omcA-mtrC-R CCTTCTACGTGGCAAGAAGC RT-PCR RT-mtrCAB-F AAGAAGAAAGGCGCATTGAA RT-PCR RT-mtrCAB-F GAGTGCGGTAAACCAACGAT RT-PCR RT_lldP_F CGCCGCCGTGTTTATTTGGC RT-PCR RT_lldP_R GGATTGCTTAATCGCAGGGC RT-PCR RT_lldP-dld_F CGCCACTCAAAGCTTGGC RT-PCR RT_lldP-dld_R CCAATAACGGCCGCGCC RT-PCR RT_dld_F CCGTTTTGCTTGGTCTACCG RT-PCR RT_dld_R GCCCTTTACGCATCGCCC RT-PCR RT_dld-lldE_F GAGCCCAAGATGCAAGCG RT-PCR RT_dld-lldE_R CGGCTCCTTCATCGACGGC RT-PCR RT_lldEF_F CAGCCTTGCTACGCCAAGCG RT-PCR RT_lldEF_R GGTTAGCGCCCATATCGGC RT-PCR RT_lldFG_F CTAAGGCTGGCGAGTCCG RT-PCR RT_lldFG_R GCAGTGCCGCAAGGCCGC RT-PCR
omcA_R+93 GTAGAAGCTTGATATTTCCCTGCAATAGTTTTAATCA HindIII, LacZ reporter
assay
omcA_F-54 GATGGAATTCTTACCCGCTTAAAGTGACTG EcoRI,LacZ reporter assay
omcA_F-69 GTACGAATTCTCAGATCTCAATCACATTACC EcoRI, LacZ reporter assay
omcA_F-84 GTACGAATTCCCTCAAAAGAAATAGTCAG EcoRI, LacZ reporter assay
omcA_F-95 GTACGAATTCGCTGTTATCTACCTCAAAAG EcoRI, LacZ reporter assay
Table 2. ( )
Primer Sequence (5′–3′) For use
omcA_F-116 GTACGAATTCTATAAAAGGAACTCAGATGATGC EcoRI, LacZ reporter assay
omcA_F-127 GTACGAATTCGCAACTGCCAATATAAAAGG EcoRI, LacZ reporter assay
omcA_F-150 GATCGAATTCATGATGCAGGCCCAAAAG EcoRI, LacZ reporter assay
mtrC_R+117 GATGAAGCTTTTTTCCCTGCATAGGTTTGG HindIII, LacZ reporter
assay
mtrC_F-42 CTCGGGATCCGCTTAGAAGATTTTAACGGCATGT BamHI, LacZ reporter
assay
mtrC_F-100 TGACGGATCCCCTTGTGGTTTAACTACCTCTTTAGAA BamHI, LacZ reporter
assay
mtrC_F-144 CTACGGATCCTCCACCTCACTATCTGTTGTTTTTTTGCTCTC BamHI, LacZ reporter
assay
mtrC_F-203 CTACGGATCCAATTATCTGAATCGAGAGACGAAA BamHI, LacZ reporter
assay
lldP_+192_R GATCGTCGACGCTTGCCTCTACATAAAGTGG SalI, LacZ reporter assay
lldP_+1_F GATCGAATTCTCATACAGCAGCCAAAATG EcoRI, LacZ reporter assay
lldP_-60_F GATCGAATTCCATGTTTTTTAAAAAAATATTTG EcoRI, LacZ reporter assay
lldP_-182_F GATCGAATTCGGATCAGTTAATTGCAATAC EcoRI, LacZ reporter assay
lldP_-360_F GATCGAATTCCAAAGCGTGACCAGAATCAC EcoRI, LacZ reporter assay
lldP_-541_F GATCGAATTCGCCGTATCGCAGCAAAAGCC EcoRI, LacZ reporter assay
omcA_RACE_out CCGCGGTCAGCTTCTGTTTCTCCCAC 5 -RACE PCR
omcA_RACE_in CGGGAGTTAATTGCGCAATACCAAATCGC 5 -RACE PCR
mtrC_RACE_out CGCCCCTTCCGGTATTAATTGCAGTGC 5 -RACE PCR
mtrC_RACE_in GCGAGACCAATCACTGGCATGTCGGC 5 -RACE PCR
lldP_race_out GCGCCGAAGATGATGGTTAAGGGGG 5 -RACE PCR
lldP_race_in ACAACGGATGCGGCCAATAGAGTGG 5 -RACE PCR
omcA-50_F CCCGCTTAAAGTGACTGATAAATACC 5 -Cy3, PBomcA2
omcA-87_F CTACCTCAAAAGAAATAGTCAG 5 -Cy3, PBomcA1
omcA+13_R GGAATTAGATCCCACCTGTAAGGC 5 -Cy3, PBomcA2
omcA-35_R CAGTCACTTTAAGCGGGTAATG 5 -Cy3, PBomcA1
mtrC-4_F CCTTGGGGAATTCTATTTCC 5 -Cy3, PBmtrC2
mtrC-143_F TCCACCTCACTATCTGTTGTTTTTTTGCTCTC 5 -Cy3, PBmtrC1
mtrC+117_R TTTTCCCTGCATAGGTTTGG 5 -Cy3, PBmtrC1,
PBmtrC2
lldP_EMSA_F GGATCAGTTAATTGCAATAC 5 -Cy3, PBlldP1, PBlldP2
lldP_EMSA_R_1 TCAGATAGTTATAAAAAAGG PBlldP1
lldP_EMSA_R_2 TGTCACTTAAAGAGTGGGG PBlldP2
lldP_EMSA-84-35_F ATATACCCCACTCTTTAAGTGACACCGATCACAGTTA AGAAAATCCCATG
Table 2. ( )
Primer Sequence (5′–3′) For use
lldP_EMSA-84-35_ R CATGGGATTTTCTTAACTGTGATCGGTGTCACTTAAA GAGTGGGGTATAT PBlldP3 lldP_EMSA-84-35m _F ATATACCCCACTCTTTACCCCCCACCGAGGGGGGTT AAGAAAATCCCATG 5′-Cy3, PBlldP3m lldP_EMSA-84-35m _R CATGGGATTTTCTTAACCCCCCTCGGTGGGGGGTAA AGAGTGGGGTATAT PBlldP3m
ldhA_EMSA_F TTTGTGCAATAGATGATAAAGCTCACTCTTTTTACGGGGTAA 5′-Cy3, PBldhA
ldhA_EMSA_R TTACCCCGTAAAAAGAGTGAGCTTTATCATCTATTGCACAAA PBldhA
ldhA_EMSA_mut_F TTTGTGCAATAGACCCCCAAGCGGGGGCTTTTTACGGGGTAA 5′-Cy3, PBldhAm
ldhA_EMSA_mut_R TTACCCCGTAAAAAGCCCCCGCTTGGGGGTCTATTGCACAAA PBldhAm
PA_omcA_-85_-116 _F
TATAAAAGGAACTCAGATGATGCTGTTATCTA 5′-biotin, PAomcA1
PA_omcA_-85_-116 _R
TAGATAACAGCATCATCTGAGTTCCTTTTATA PAomcA1
PA_omcA_-54_-84_ F
CCTCAAAAGAAATAGTCAGATCTCAATCACA 5′-biotin, PAomcA2
PA_omcA_-54_-84_ R TGTGATTGAGATCTGACTATTTCTTTTGAGG PAomcA2 PA_omcA_-54_-116 _F TATAAAAGGAACTCAGATGATGCTGTTATCTACCTCAAAAG AAATAGTCAGATCTCAATCACA 5′-biotin, PAomcA3 PA_omcA_-54_-116 _R TGTGATTGAGATCTGACTATTTCTTTTGAGGTAGATAACAG CATCATCTGAGTTCCTTTTATA PAomcA3
qRT-pta-F TAGGTGCCCTGCTGCTTACT qRT-PCR for pta
qRT-pta-R GAGGTTTGCCAAGTGTTGGT qRT-PCR for pta
qRT-pflB-F CCCAATGGTTGTGGGTAAAC qRT-PCR for pflB
qRT-pflB-R AGTGATTGGGTCTGCTTTGG qRT-PCR for pflB
qRT-pykA-F CAAAATCGTCACCACACTGG qRT-PCR for pykA
qRT-pykA-R TGAGTAGCGCGTTTGAGATG qRT-PCR for pykA
qRT-metK-F ATGCCAGCAACGAAACTGAC qRT-PCR for metK
qRT-metK-R ACTTGGCTTTTCGCATCTGG qRT-PCR for metK
qRT-hisC-F AGCAAAATTGTCGCCAGTCG qRT-PCR for hisC
qRT-hisC-R AAAGGTTTGCGCGCTAATGG qRT-PCR for hisC
qRT-acs-F TAATCCAAAGGGCGTATTGC qRT-PCR for acs
qRT-acs-R AACCTACATCGGCCGTACAC qRT-PCR for acs
qRT-metR-F TCAGCTAAGGGTTGGGATTG qRT-PCR for metR
qRT-metR-R TTCTAGGGCATTGAGCCGAGT qRT-PCR for metR
qRT-metE-F GAAGGTGTGGGCTTTACCAA qRT-PCR for metE
Table 2. ( )
Primer Sequence (5′–3′) For use
ScpdA_F_EcoRI GGCCGGAATTCTGGCAGTCAACAACGTTTGG S. oneidensis cpdA
complementation
ScpdA_R_BamHI GGCGCGGATCCTTCCTCAAGCTGCGGTCATT S. oneidensis cpdA
complementation
EcpdA_F_KpnI GGGGGGTACCGCGATTCCGGTTTATTAGCG E. coli cpdA
complementation
EcpdA_R_BamHI GGGGGGGATCCCGAGCTGTTGAAACCGTG E. coli cpdA
complementation
cyaC_F_EcoRI GGGGGAATTCCTATAAACTACGGCATCCTG cyaC constitutive
expression
cyaC_R_SacI GGGGGAGCTCTGAGTATTTTGCCTGTTGTG cyaC constitutive
expression
crp_F_EcoRI GGGGGAATTCTTTCGTTTTAATCAATCGAGG crp constitutive expression
3
CRP
D
-3.1
S. oneidensis MR-1 EET mtr CRP MR-1 (lactate dehydrogenase; LDH) (EET ) ( ) MR-1 CRP3.2
3.2.1
2 Table 1BL21 (DE3) LB 2 × yeast extrac-triptone (2 × YT) 30ºC 37ºC
S. oneidensis 10 mM DL- D- L (9 mM (NH4)2SO4, 5.7 mM K2HPO4, 3.3 mM KH2PO4, 30 mM HEPES-NaOH (pH 7.4)) (DL-LMM, D-LMM, L-LMM, PMM ) 5 ml 30 mL S. oneidensis OD600 = 0.05 180 rpm 5 ml 13 mL 30 mM S. oneidensis OD600 = 0.01 OD600 Miniphoto518R (Taitech) 100 µg/ml
Am, 50 µg/ml Km, 15 µg/ml Gm 1.5% Bacto agar (Difco)
3.2.2
DLS. oneidensis DL-LMM 1 ml
0.2 µm D-/L
(F-kit D-/L-lactate, JK ) 340 nm
Abs340 1200lab (Hitachi) 96 90 µl H2O, 100 µl
5 Abs340 (E0) 2 µl Solution 4
30 Abs340 (E1) 2 µl Solution 5
30 Abs340 (E2) E1 E0 E2 E1
D- L- 3
3.2.3
RNA
RNA DL-LMM S. oneidensis 2.2.2
RNA Bioanalyzer (Agilent Technologies)
3.2.4
qRT-PCR
(16S rRNA, lldP, dld, lldF, ldhA) DNA MR-1 total DNA PCR
(2.2.5 ) Total RNA DL-LMM
(2.2.2 ) mRNA
(16S rRNA) 3
3.2.5
crp
dld
crp dld (pBBRcrp, pBBRdld) crp
dld Phusion High-Fidelity DNA polymerase (New England Biolabs)
crp_F_EcoRI, crp_R_BamHI dld_F_BamHI, dld_R_XbaI ( 2 Table 2)
PCR PCR QIAquick PCR Purification kit (Qiagen)
(2.2.3 ) dld pBBR1MCS-2 EcoRI BamHI BamHI XbaI (pBBRcrp, pBBRdld) crp dld
3.2.6
2.2.3 5′-RACE RNA MR-1 total RNA 1.0 µg 2 lldP_race_out lldP_race_in3.2.7
RT-PCR
MR-1 total RNA 5.0 µg SuperScriptIII reverse transcriptase (Invitrogen) Rnadom Primers
(Invitrogen) cDNA cDNA 2
DNA 2.2.4
3.2.8
LacZ
lldP lacZ
(lldP_+1_F lldP_-541_F) (lldP_+192_R) ( 2 Table 2) PCR
PCR QIAquick PCR Purification kit (Qiagen) 2.2.3
PCR pMElacZ EcoRI SalI
MR-1 ∆crp
3.2.9
DL-LMM 2.2.9 β- 3
3.2.10
His
CRP
His-tag-CRP pET-crp ( 2 Table 1)
BL21 (DE3) 300 mL 100 ml Km 2 × YT
OD600 = 0.05 30ºC 180 rpm OD600 = 0.5–0.8
Isopropyl-β-D-galactopyranoside (IPTG) 0.1 mM
His-tag-CRP 3 50 ml
His-tag-CRP QuickPick IMAC Metal Affinity kit for proteins (Bio-Nobile)
His-tag-CRP Micro BCA Protein Assay kit (Thermo Fisher Science)
3.2.11
EMSA
2 nM Cy3 DNA 0–200 µg His-tag-CRP (10 mM Tris-HCl
(pH 7.6), 0.5 mM EDTA (pH 8.0), 100 mM KCl, 50 µg/ml , 50 µg/ml poly
(deoxyinosinic-deoxycytidylic) acid [poly (di-dc) ], 0 - 50 µM cAMP, 10% )
20 µl 30 12.5%
0.5 × Tris-borate-EDTA
3.3
3.3.1
CRP
MR-1 CRP MR-1 (WT) CRP (∆crp) DL- ( ) D- L (DL-LMM, D-LMM, L-LMM, PMM, ) (Figure 1) DL-LMM ∆crp WT (Figure 1A) D-/L -∆crp D- (Figure 2) ∆crp D-LMM (Figure 1B) ∆crp crp D-LMM (Figure 3) CRP D -CRP D -∆crp L-LMM PMM (Figure 1C, D) CRP D -L-3.3.2
D-
CRP
CRP D- (D- ; D-LDH) MR-1 2D-LDH (Dld LdhA) LdhA NADH D
-D-LDH (Pinchuk et al. 2009) Dld D
-D-LDH (Pinchuk et al. 2009,
Brutinel and Gralnick, 2012) ∆crp D- dld
MR-1 dld lldP L-LDH lldEFG (LDH ) (Figure 4A) CRP RT-PCR qRT-PCR WT ∆crp MR-1 DL-LMM RNA RT-PCR lldP–dld
(Figure 4B, lane 2) mRNA
lldE–lldF (Figure 4B, lane 5) lldF–lldG (Figure 4B, lane 6)
dld-lldE (Figure 4B, lane 4)
LDH 5 lldP dld lldEFG 2
lldP dld lldF qRT-PCR ∆crp WT lldP dld 20% CRP lldP–dld lldF WT ∆crp CRP lldEFG ∆crp lldP dld TMAO (Figure 5) CRP
3.3.3
crp
D∆crp D- dld dld (pBBRdld) ∆crp ∆crp(pBBRdld) D-LMM MR-1(pBBR1MCS-2) ( ) (Figure 6A) ∆crp(pBBR1MCS-2) ( ∆crp) ∆crp(pBBRdld) TMAO MR-1(pBBR1MCS-2) D-LMM (Figure 6B) TMAO ∆crp(pBBRdld) ∆crp TMAO ∆crp D- dld
3.3.4
lldP-dld
DMR-1 D-LDH L-LDH L- (Brutinel and Gralnick 2012) D- L- lldP-dld lldEFG D-lactate L- MR-1 qRT-PCR lldP lldF (Figure 7) MR-1 L- lldF lldEFG L- ( . LlpR)
(Brutinel and Gralnick 2012) lldP
lldP-dld lldEFG
lldP-dld D
lldP–dld lldP–dld lldP 5 -RACE lldP 192 bp (TSSlldP) (Figure 8) 70 (-10 box, -35 box) CRP (Gao et al. 2010) CRP lldP–dld lldP-dld lacZ β-pMElacZ lacZ 5′ lldP
( 2 Table 1; pMElldP+1, pMElldP-60, pMElldP-182,
pMElldP-541) MR-1 DL-LMM CRP pMElldP-182 pMElldP-541 (Figure 9) TSSlldP 60 bp 182 bp lldP-dld ∆crp (Figure 9) CRP
3.3.6
CRP
lacZ lldP-dld CRP EMSA TSSlldP -60 bp -182 bp CRP (PBlldP1; Figure 10A) TSSlldP -182 bpCRP (PBlldP2; Figure 10A) 2 Cy3 DNA
(PBlldP3m; Figure 10A) EMSA
CRP PBlldP3 PBlldP3m (Figure
10B) CRP lldP-dld
3.4
Shewanella
(Serres
and
Riley 2006, Fredrickson et al. 2008) S.oneidensis MR-1 MR-1 DL -Shewanella MR-1 D -D-LDH dld CRP CRP MR-1 dld CRP Corynebacterium glutamicum CRP/Fnr GlxR L-LDH ldhA narKGHJI cAMP
(Kohl et al. 2008) cAMP
NADH LDH (iLDH)
(Garvie 1980, Jiang et al. 2014) C. glutamicum D
-D-iLDH (Dym et al. 2000, Kato et al.
2010) MR-1 D- D-iLDH (Dld) (Pinchuk
et al. 2009) CymA
( DMSO
) (Beliaev et al. 2001, Myers and Myers 2002, Gralnick et al. 2006, Shi et al.
2006, Hartshorne et al. 2009, Schuetz et al. 2009, Fonseca et al. 2013) D
L-LDH lldEFG CRP
(Figure 4C) lldEFG L- LlpR
(Pinchuk et al. 2009, Brutinel and Gralnick 2012)
(Figure 7) lldEFG L
Figure 3. crp (pBBR1MCS-2) MR-1 (
) ∆crp ( ) crp (pBBRcrp) ∆crp (
) 50 µg/ml Km D-LMM
Figure S1. Growth of crp-complemented ∆crp on D-lactate. WT harboring the control vector
pBBR1MCS-5 (closed circle), ∆crp harboring pBBR1MCS-5 (open circle), and ∆crp harboring pBBRcrp (open squire) were aerobically grown in MM containing 10 mM D-lactate. Error bars represent standard deviations calculated from at least three independent experiments.
Figure 5. TMAO WT ∆crp lldP dld qRT-PCR WT ∆crp 16S rRNA 0 0.2 0.4 0.6 0.8 1 1.2 1.4 lldP dld
Relative mRNA levels
Figure 6. ∆crp(pBBRdld) D-LMM A. B. TMAO
Figure S3. Growth of dld-complemented ∆crp on D-lactate under TMAO-reducing conditions. WT harboring the control vector pBBR1MCS-2 (closed circle), ∆crp harboring pBBR1MCS-2 (open circle), and ∆crp harboring pBBRdld (open squire) were anaerobically grown in MM containing 30 mM D-lactate and 30 mM TMAO. Error bars represent standard deviations calculated from at least three independent experiments.
Figure 9. β- LacZ WT ( ) ∆crp ( ) DL-LMM CRP 0 50 100 150 Miller units WT ∆crp lldP TSS (+1) pMElldP-541 pMElldP-182 pMElldP+1
Figure 10. EMSA. A. EMSA DNA (PBlldP1, PBlldP2, PBlldP3, PBlldP3m)
CRP B. PBlldP1, PBlldP2, PBlldP3
PBlldP3m CRP CRP cAMP (+) (-)
DNA CRP
fmicb-08-00869 May 13, 2017 Time: 16:27 # 8
Kasai et al. CRP RegulatesD-Lactate Oxidation
FIGURE 9 | Electrophoretic mobility shift assay (EMSA) using CRP and upstream regions of lldP. (A) Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE) of N-his-CRP protein samples. Protein samples (5 µg) were analyzed on 12.5% SDS-polyacrylamide gels. Lane 1, Escherichia coli BL21 (DE3) (pET-crp) crude extract; lane 2, purified N-his-CRP; lane 3, molecular weight marker.(B) DNA fragments used as probes. Positions of 50and 30ends of
the fragments relative to TSSlldP(+1) are shown. White boxes represent the putative CRP-binding motif. The mutated sequences in PBlldP3m are shown in bold.
(C) Binding of CRP to each probe. DNA-binding reactions were performed in the presence (+) or absence (–) of CRP, cAMP, and specific competitor (2 µM unlabeled PBlldP3 probe).
DISCUSSION
Like many other anaerobic respiratory bacteria, Shewanella spp. preferentially catabolize low-molecular-weight organic
acids (Serres and Riley, 2006; Fredrickson et al., 2008),
and studies have used lactate (mostly racemic DL-lactate) as
a substrate for cultivating S. oneidensis MR-1. Despite the importance of lactate as a catabolic substrate for Shewanella spp., however, little is known about how these bacteria regulate catabolic pathways for this compound. Here, we demonstrated that CRP plays a critical role in the transcriptional
regulation of the dld gene encoding D-LDH in MR-1. As
CRP is also involved in the expression of many anaerobic
respiratory genes (Saffarini et al., 2003; Kouzuma et al.,
2015), we suggest that MR-1 uses this transcriptional regulator
to coordinately regulate D-lactate metabolism and anaerobic
respiration. A previous study reported that, in Corynebacterium glutamicum, a CRP/Fnr-type global transcriptional regulator, GlxR, binds to promoter regions of the ldhA gene encoding
a fermentative L-LDH and the narKGHJI operon encoding
nitrate respiratory enzymes in a cAMP-dependent manner (Kohl et al., 2008). It is therefore likely that a broad range
of bacteria utilizes cAMP-dependent regulatory mechanisms for the coordinated expression of catabolic and respiratory pathways.
In many bacteria, lactate is oxidized to pyruvate by
NADH-independent LDHs (iLDHs) (Garvie, 1980;Jiang et al.,
2014). For example, in E. coli and C. glutamicum, D-lactate
is catabolized through membrane-bound D-iLDHs that utilize
membrane-associated quinones as electron-accepting cofactors (Dym et al., 2000; Kato et al., 2010). S. oneidensis MR-1 also
oxidizes D-lactate by D-iLDH (Dld) (Pinchuk et al., 2009),
and is thereby likely to reduce membrane quinones. In this strain, electrons accumulated in the membrane quinone pool are transferred to anaerobic electron acceptors, such as fumarate, DMSO, and metal oxides, via an inner membrane-anchored cytochrome, CymA, and periplasm- and outer membrane-localized proteins, such as FccA, DmsABEF, MtrCAB, and
OmcA (Beliaev et al., 2001; Myers and Myers, 2002; Gralnick
et al., 2006; Shi et al., 2006; Hartshorne et al., 2009; Schuetz et al., 2009; Fonseca et al., 2012). It is therefore suggested
that D-lactate oxidation in MR-1 is metabolically linked
to anaerobic respiratory pathways via membrane-associated quinones.
Frontiers in Microbiology | www.frontiersin.org 8 May 2017 | Volume 8 | Article 869
fmicb-08-00869 May 13, 2017 Time: 16:27 # 8
Kasai et al. CRP RegulatesD-Lactate Oxidation
FIGURE 9 | Electrophoretic mobility shift assay (EMSA) using CRP and upstream regions of lldP. (A) Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE) of N-his-CRP protein samples. Protein samples (5 µg) were analyzed on 12.5% SDS-polyacrylamide gels. Lane 1, Escherichia coli BL21 (DE3) (pET-crp) crude extract; lane 2, purified N-his-CRP; lane 3, molecular weight marker.(B) DNA fragments used as probes. Positions of 50and 30ends of
the fragments relative to TSSlldP(+1) are shown. White boxes represent the putative CRP-binding motif. The mutated sequences in PBlldP3m are shown in bold.
(C) Binding of CRP to each probe. DNA-binding reactions were performed in the presence (+) or absence (–) of CRP, cAMP, and specific competitor (2 µM unlabeled PBlldP3 probe).
DISCUSSION
Like many other anaerobic respiratory bacteria, Shewanella spp. preferentially catabolize low-molecular-weight organic acids (Serres and Riley, 2006; Fredrickson et al., 2008), and studies have used lactate (mostly racemic DL-lactate) as a substrate for cultivating S. oneidensis MR-1. Despite the importance of lactate as a catabolic substrate for Shewanella spp., however, little is known about how these bacteria regulate catabolic pathways for this compound. Here, we demonstrated that CRP plays a critical role in the transcriptional regulation of the dld gene encoding D-LDH in MR-1. As
CRP is also involved in the expression of many anaerobic respiratory genes (Saffarini et al., 2003; Kouzuma et al., 2015), we suggest that MR-1 uses this transcriptional regulator to coordinately regulate D-lactate metabolism and anaerobic respiration. A previous study reported that, in Corynebacterium glutamicum, a CRP/Fnr-type global transcriptional regulator, GlxR, binds to promoter regions of the ldhA gene encoding a fermentative L-LDH and the narKGHJI operon encoding
nitrate respiratory enzymes in a cAMP-dependent manner (Kohl et al., 2008). It is therefore likely that a broad range
of bacteria utilizes cAMP-dependent regulatory mechanisms for the coordinated expression of catabolic and respiratory pathways.
In many bacteria, lactate is oxidized to pyruvate by NADH-independent LDHs (iLDHs) (Garvie, 1980;Jiang et al., 2014). For example, in E. coli and C. glutamicum,D-lactate is catabolized through membrane-boundD-iLDHs that utilize membrane-associated quinones as electron-accepting cofactors (Dym et al., 2000;Kato et al., 2010). S. oneidensis MR-1 also oxidizes D-lactate by D-iLDH (Dld) (Pinchuk et al., 2009), and is thereby likely to reduce membrane quinones. In this strain, electrons accumulated in the membrane quinone pool are transferred to anaerobic electron acceptors, such as fumarate, DMSO, and metal oxides, via an inner membrane-anchored cytochrome, CymA, and periplasm- and outer membrane-localized proteins, such as FccA, DmsABEF, MtrCAB, and OmcA (Beliaev et al., 2001;Myers and Myers, 2002;Gralnick et al., 2006;Shi et al., 2006;Hartshorne et al., 2009;Schuetz et al., 2009; Fonseca et al., 2012). It is therefore suggested that D-lactate oxidation in MR-1 is metabolically linked
to anaerobic respiratory pathways via membrane-associated quinones.
Frontiers in Microbiology | www.frontiersin.org 8 May 2017 | Volume 8 | Article 869
A
7
7.1
CRP
(Brückner and Titgemeyer 2002, Stülke and
Hillen 1999) E. coli K-12 CRP cAMP
cAMP
(phosphotransferase system; PTS) PTS
PTS PTS
Cya cAMP
cAMP (Green et al. 2014)
Shewanella CRP
Shewanella sp. ANA-3 CRP
(Murphy and Saltikov 2009) S. oneidensis MR-1
CRP Fe(III) Mn(IV) DMSO
(mtr, fccA, nap, dms )
(Saffarini et al. 2003, Charania et al. 2009)
CRP CRP MR-1 cAMP/CRP 2 MR-1 Mtr mtr CRP 3 MR-1 CRP D- (Dld) (dld) Dld D -D -MR-1 D- CRP 5 CRP cAMP CpdA CpdA (∆cpdA) SAM ∆cpdA
Beliaev AS, Klingeman DM, Klappenbach JA, Wu L, Romine MF, Tiedje JM et al. (2005). Global transcriptome analysis of Shewanella oneidensis MR-1 exposed to different terminal electron acceptors. J Bacteriol. 187(20):7138-7145.
Beliaev AS, Saffarini DA, McLaughlin JL, Hunnicutt D. (2001). MtrC, an outer membrane decahaem c cytochrome required for metal reduction in Shewanella putrefaciens MR-1. Mol Microbiol. 39(3):722-730.
Botsford JL and Harman JG. (1992). Cyclic AMP in prokaryotes. Microbiol Rev. 56(1):100-122. Brückner R and Titgemeyer F. (2002) Carbon catabolite repression in bacteria: choice of the carbon source and autoregulatory limitation of sugar utilization. FEMS Microbiol Lett. 209(2):141-148.
Brutinel ED, Gralnick JA. (2012). Preferential utilization of D-lactate by Shewanella oneidensis.
Appl Environ Microbiol. 78(23):8474-8476.
Busby S and Ebright RH. (1997) Transcription activation at class II CAP-dependent promoters. Mol Microbiol. 23(5):853-859.
Carpentier W, Sandra K, De Smet I, Brigé A, De Smet L, Van Beeumen J. (2003). Microbial reduction and precipitation of vanadium by Shewanella oneidensis. Appl Environ Microbiol. 69:3636-3639.
Charania MA, Brockman KL, Zhang Y, Banerjee A, Pinchuk GE, Fredrickson JK et al. (2009). Involvement of a membrane-bound class III adenylate cyclase in regulation of anaerobic respiration in Shewanella oneidensis MR-1. J Bacteriol. 191(13):4298-4306.
Coursolle D, Baron DB, Bond DR, Gralnick JA. (2010). The Mtr respiratory pathway is essential for reducing flavins and electrodes in Shewanella oneidensis. J Bacteriol. 192(2):467-474. doi: 10.1128/JB.00925-09.
Dym O, Pratt EA, Ho C, Eisenberg D. (2000). The crystal structure of D-lactate dehydrogenase, a
Flynn TM, O'Loughlin EJ, Mishra B, DiChristina TJ, Kemner KM. (2014). Sulfur-mediated electron shuttling during bacterial iron reduction. Science. 344:1039-1042. doi: 10.1126/science.1252066. Fonseca BM, Paquete CM, Neto SE, Pacheco I, Soares CM, Louro RO. (2013). Mind the gap: cytochrome interactions reveal electron pathways across the periplasm of Shewanella oneidensis MR-1. Biochem J. 449(1):101-108.
Fredrickson JK, Romine MF, Beliaev AS, Auchtung JM, Driscoll ME, Gardner TS, et al. (2008). Towards environmental systems biology of Shewanella. Nat Rev Microbiol. 6:592-603. doi: 10.1038/nrmicro1947.
Fuchs EL, Brutinel ED, Klem ER, Fehr AR, Yahr TL, Wolfgang MC. (2010). In vitro and in vivo characterization of the Pseudomonas aeruginosa cyclic AMP (cAMP) phosphodiesterase CpdA, required for cAMP homeostasis and virulence factor regulation. J Bacteriol. 192(11):2779-2790. Gao H, Wang X, Yang ZK, Chen J, Liang Y, Chen H et al. (2010). Physiological roles of ArcA, Crp, and EtrA and their interactive control on aerobic and anaerobic respiration in Shewanella oneidensis. PLoS One. 5(12):e15295. doi: 10.1371/journal.pone.0015295.
Garvie EI. (1980). Bacterial lactate dehydrogenases. Microbiol Rev. 44(1):106-139.
Gralnick JA, Vali H, Lies DP, Newman DK. (2006). Extracellular respiration of dimethyl sulfoxide by Shewanella oneidensis strain MR-1. Proc Natl Acad Sci U S A. 103(12):4669-4674.
Green J, Stapleton MR, Smith LJ, Artymiuk PJ, Kahramanoglou C, Hunt DM, Buxton RS. (2014). Cyclic-AMP and bacterial cyclic-AMP receptor proteins revisited: adaptation for different ecological niches. Curr Opin Microbiol. 18:1-7.
Hartshorne RS, Reardon CL, Ross D, Nuester J, Clarke TA, Gates AJ et al. (2009). Characterization of an electron conduit between bacteria and the extracellular environment. Proc Natl Acad Sci U S A. 106(52):22169-22174.
Hau HH, Gilbert A, Coursolle D, Gralnick JA. (2008). Mechanism and consequences of anaerobic respiration of cobalt by Shewanella oneidensis strain MR-1. Appl Environ Microbiol. 74:6880-6886. doi: 10.1128/AEM.00840-08.
Heidelberg JF, Paulsen IT, Nelson KE, Gaidos EJ, Nelson WC, Read TD, et al. (2002). Genome sequence of the dissimilatory metal ion-reducing bacterium Shewanella oneidensis. Nat Biotechnol. 20:1118-1123.
Jiang T, Gao C, Ma C, Xu P. (2014). Microbial lactate utilization: enzymes, pathogenesis, and regulation. Trends Microbiol. 22(10):589-599.
Joung JK, Le LU, Hochschild A. (1993). Synergistic activation of transcription by Escherichia coli cAMP receptor protein. Proc Natl Acad Sci U S A. 90(7):3083-3087.
Kasai T, Kouzuma A, Nojiri H, Watanabe K. (2015). Transcriptional mechanisms for differential expression of outer membrane cytochrome genes omcA and mtrC in Shewanella oneidensis MR-1. BMC Microbiol. 15:68.
Kasai T, Kouzuma A, Watanabe K. (2017). CRP regulates D-lactate oxidation in Shewanella
oneidensis MR-1. Front Microbiol. 8:869.
Kato O, Youn JW, Stansen KC, Matsui D, Oikawa T, Wendisch VF. (2010). Quinone-dependent
D-lactate dehydrogenase Dld (Cg1027) is essential for growth of Corynebacterium glutamicum on
D-lactate. BMC Microbiol. 10:321.
Kim BH, Kim HJ, Hyun MS, Park DH. (1999). Direct electrode reaction of Fe(III)-reducing bacterium, Shewanella putrefaciens. J. Microbiol. Biotechnol. 9:127–131.
Kohl TA, Baumbach J, Jungwirth B, Pühler A, Tauch A. (2008). The GlxR regulon of the amino acid producer Corynebacterium glutamicum: in silico and in vitro detection of DNA binding sites of a global transcription regulator. J Biotechnol. 135(4):340-350.
Kouzuma A, Kasai T, Hirose A, Watanabe K. (2015). Catabolic and regulatory systems in Shewanella oneidensis MR-1 involved in electricity generation in microbial fuel cells. Front Microbiol. 6:609.
Kovach ME, Elzer PH, Hill DS, Robertson GT, Farris MA, Roop RM II, et al. (1995). Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene. 166(1):175-176.
Kristensen HH, Valentin-Hansen P, Søgaard-Andersen L. (1997). Design of CytR regulated, cAMP-CRP dependent class II promoters in Escherichia coli: RNA polymerase-promoter interactions modulate the efficiency of CytR repression. J Mol Biol. 266(5):866-876.
Liu C, Gorby YA, Zachara JM, Fredrickson JK, Brown CF. (2002). Reduction kinetics of Fe(III), Co(III), U(VI), Cr(VI), and Tc(VII) in cultures of dissimilatory metal-reducing bacteria. Biotechnol Bioeng. 80:637-649.
Lower BH, Shi L, Yongsunthon R, Droubay TC, McCready DE, Lower SK. (2007). Specific bonds between an iron oxide surface and outer membrane cytochromes MtrC and OmcA from Shewanella oneidensis MR-1. J. Bacteriol. 189(13):4944-4952.
Makman RS, Sutherland EW. (1965). Adenosine 3',5'-phosphate in Escherichia coli. J Biol Chem. 240:1309-1314.
Marsili E, Baron DB, Shikhare ID, Coursolle D, Gralnick JA, Bond DR. (2008). Shewanella secretes flavins that mediate extracellular electron transfer. Proc Natl Acad Sci U S A. 105(10):3968-3973. doi: 10.1073/pnas.0710525105.
Miller JH. (1972). Experiments in molecular genetics. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
Mitra A, Kesarwani AK, Pal D, Nagaraja V. (2011). WebGeSTer DB--a transcription terminator database. Nucleic Acids Res. 39:D129-135.
Murphy JN, Saltikov CW. (2009). The ArsR repressor mediates arsenite-dependent regulation of arsenate respiration and detoxification operons of Shewanella sp. strain ANA-3. J Bacteriol. 191:6722-6731. doi: 10.1128/JB.00801-09.
Myers CR, Myers JM. (1997). Outer membrane cytochromes of Shewanella putrefaciens MR-1: spectral analysis, and purification of the 83-kDa c-type cytochrome. Biochim Biophys Acta.1326(2):307-318.
Myers CR, Myers JM. (2002). MtrB is required for proper incorporation of the cytochromes OmcA and OmcB into the outer membrane of Shewanella putrefaciens MR-1. Appl Environ Microbiol. 68(11):5585-5594.
Myers CR, Nealson KH. (1988). Bacterial manganese reduction and growth with manganese oxide as the sole electron acceptor. Science. 240:1319-1321.
Nakagawa G, Kouzuma A, Hirose A, Kasai T, Yoshida G, Watanabe K. (2015). Metabolic Characteristics of a Glucose-Utilizing Shewanella oneidensis Strain Grown under Electrode-Respiring Conditions. PLoS One. 10(9):e0138813.
Nealson KH, Saffarini D. (1994). Iron and manganese in anaerobic respiration: environmental significance, physiology, and regulation. Annu Rev Microbiol. 48:311-343.
Penfold RJ, Pemberton JM. (1992). An improved suicide vector for construction of chromosomal insertion mutations in bacteria. Gene. 1992 Sep 1;118(1):145-146.
Pinchuk GE, Rodionov DA, Yang C, Li X, Osterman AL, Dervyn E, et al. (2009). Genomic reconstruction of Shewanella oneidensis MR-1 metabolism reveals a previously uncharacterized machinery for lactate utilization. Proc Natl Acad Sci U S A. 106(8):2874-2879.
10.1073/pnas.1410551111.
Raibaud O and Schwartz M. (1984). Positive control of transcription initiation in bacteria. Annu Rev Genet. 18:173-206.
Rodionov DA, Novichkov PS, Stavrovskaya ED, Rodionova IA, Li X, Kazanov MD, et al. (2011). Comparative genomic reconstruction of transcriptional networks controlling central metabolism in the Shewanella genus. BMC Genomics. 15;12 Suppl 1:S3. doi: 10.1186/1471-2164-12-S1-S3. Rosenbaum MA, Bar HY, Beg QK, Segrè D, Booth J, Cotta MA, Angenent LT. (2012.) Transcriptional analysis of Shewanella oneidensis MR-1 with an electrode compared to Fe(III)citrate or oxygen as terminal electron acceptor. PLoS One. 7(2):e30827. doi: 10.1371/journal.pone.0030827.
Ross DE, Flynn JM, Baron DB, Gralnick JA, Bond DR. (2011). Towards electrosynthesis in Shewanella: energetics of reversing the Mtr pathway for reductive metabolism. PLoS One. 6(2):e16649. doi: 10.137
1/journal.pone.0016649.
Saffarini DA, Schultz R, Beliaev A. (2003). Involvement of cyclic AMP (cAMP) and cAMP receptor protein in anaerobic respiration of Shewanella oneidensis. J Bacteriol. 185(12):3668-3671. Schuetz B, Schicklberger M, Kuermann J, Spormann AM, Gescher J. (2009). Periplasmic electron transfer via the c-type cytochromes MtrA and FccA of Shewanella oneidensis MR-1. Appl Environ Microbiol. 75(24):7789-7796.
Serres MH, Riley M. (2006). Genomic analysis of carbon source metabolism of Shewanella oneidensis MR-1: Predictions versus experiments. J Bacteriol. 188(13):4601-4609.
Shao W, Price MN, Deutschbauer AM, Romine MF, Arkin AP. (2014). Conservation of transcription start sites within genes across a bacterial genus. MBio. 5(4):e01398-14.
Shi L, Squier TC, Zachara JM, Fredrickson JK. (2007). Respiration of metal (hydr)oxides by Shewanella and Geobacter: a key role for multihaem c-type cytochromes. Mol Microbiol. 65(1):12-20.
Stülke J and Hillen W. (1999). Carbon catabolite repression in bacteria. Curr Opin Microbiol. 2(2):195-201.
Teal TK, Lies DP, Wold BJ, Newman DK. (2006). Spatiometabolic stratification of Shewanella oneidensis biofilms. Appl Environ Microbiol. 72(11):7324-7330.
Tebbutt J, Rhodius VA, Webster CL, Busby SJ. (2002). Architectural requirements for optimal activation by tandem CRP molecules at a class I CRP-dependent promoter. FEMS Microbiol Lett. 210:55-60.
Ullmann A, Monod J. (1968). Cyclic AMP as an antagonist of catabolite repression in Escherichia coli. FEBS Lett. 2(1):57-60.
Valentin-Hansen P, Søgaard-Andersen L, Pedersen H. (1996). A flexible partnership: the CytR anti-activator and the cAMP-CRP activator protein, comrades in transcription control. Mol Microbiol. 20(3):461-466.
Venkateswaran K, Moser DP, Dollhopf ME, Lies DP, Saffarini DA, MacGregor BJ, et al. (1999). Polyphasic taxonomy of the genus Shewanella and description of Shewanella oneidensis sp. nov. Int J Syst Bacteriol. 49:705-724.
Von Canstein H, Ogawa J, Shimizu S, Lloyd JR. (2008). Secretion of flavins by Shewanella species and their role in extracellular electron transfer. Appl Environ Microbiol. 74(3):615-623.
Wang H, Correa E, Dunn WB, Winder CL, Goodacre R, Lloyd JR. (2013). Metabolomic analyses show that electron donor and acceptor ratios control anaerobic electron transfer pathways in Shewanella oneidensis. Metabolomics, 9(3): 642-656.
cytochrome OmcA. J Am Chem Soc. 128(43):13978-13979.
Yang Y, Harris DP, Luo F, Wu L, Parsons AB, Palumbo AV, Zhou J. (2008). Characterization of the Shewanella oneidensis Fur gene: roles in iron and acid tolerance response. BMC Genomics. 9 Suppl 1:S11.
Yanisch-Perron C, Vieira J, Messing J. (1985). Improved M13 phage cloning vectors and host strains: nucleotide sequences of the M13mp18 and pUC19 vectors. Gene. 33(1):103-119.
Zhou G, Yin J, Chen H, Hua Y, Sun L, Gao H. (2013). Combined effect of loss of the caa3 oxidase and Crp regulation drives Shewanella to thrive in redox-stratified environments. ISME J. 7(9):1752-1763.