[5
t曹
ム
﹁,
1︑﹁ヤ.
︑
雫﹁
'
︑戸
‑一
1
ヒ﹁
臆
﹁サ
}
虞・
F7
㍉﹁
■q
整
▲鳶
̀
■
F
監〜
論 文 名
感 覚 一運 動系 の随意運動制御 にお ける交感神 経皮膚反応 と 事 象関連電位 の関係(英 文)
著 者
審査担 当者
主 査
委 員
委 員
委 員
i田 政 冑
/努 守 司 潅こ
ナ じ 一?,i
41ψ ・ 芳 ヌ誕
\ わ 手 努/15
上 記 の 論 文 を 合 格 と判 定 す る
平成 之 年3月/6日
東京都立大学大学院理学研究科教授会
聯 長桑r冷 禍
DISSERTATION FOR A DEGREE OF DOCTOR OF SCIENCE
TOKYO METROPOLITAN UNIVERSITY
TITLE: The relationship between the sympathetic skin response and event-related brain potentials in sensorimotor
control of human voluntary movements
AUTHOR:
r t (-
EXAMINED BY
Examiner in chief
Examiner ;9 / 7 /
Examiner
Examiner
„X ,
QUALIFIED BY THE GRADUATE SCHOOL OF SCIENCE TOKYO METROPOLITAN UNIVERSITY
Dean
Date /41(eLk 2....c,c5o
HUMAN VOL UNTARYMOVEMENTS
2000
Masahiro Shimoda M. P.
Department of Ecoregion Science, Faculty of Agriculture,
Tokyo University of Agriculture and Technology
3-5-8 Saiwai-Cho, Fuchu-City 183-8509,
Japan
This thesis describes original research carried out by the author whilst enrolled in the doctoral program in the Graduate School of Science at the Tokyo Metropolitan University from April 1996 to May 1998. The experiments reported in this thesis were conducted by the author at the Institute of Health and Sport Science of the University of Tsukuba.
M Shimo da
I would first like to express many thanks to my supervisor, Professor Kuniyasu Imanaka, Department of Kinesiology, Graduate School of Science, Tokyo Metropolitan University, for his encouragement, detailed guidance and comprehensive suggestions throughout the course of this dissertation. I would also like to express my gratitude to Dr.
Yoshiaki Nishihira, Institute of Sport Sciences, University of Tsukuba, for his orientation and encouragement in the beginning of my course of study. My thanks go to Dr. Ichiro Kita, Department of Kinesiology, and Dr. Yoshio Sugiura, Department of Urban and Human Geography, Graduate School of Science, Tokyo Metropolitan University, for their helpful comments on this thesis.
A special thanks is also extended to my colleagues at Tokyo University of Agriculture and Technology, particularly to Dr. Teruo Uetake, Department of Ecoregion Science, Faculty of Agriculture, for his support during the preparation of my dissertation at T.U.A.T. Without him this dissertation would not have been possible.
I also wish to extend my thanks to my colleagues at University of Tsukuba for
their assistance, encouragement and suggestions throughout the experiments reported in
this dissertation.
Table of Contents
TABLE OF CONTENTS
ACKNOWLEDGMENTS TABLE OF CONTENTS . LIST OF TABLES
11
LIST OF FIGURES ABSTRACT
.. VI
viii .. ix
CHAPTER 1 INTRODUCTION...
1.1 THE AUTONOMIC AND CENTRAL NERVOUS SYSTEMS 1.2 TERMINOLOGY...
1.2.1 Emotional Sweating
1.2.2 Electrodermal Activity (EDA) 1.2.3 Orienting Response (OR)
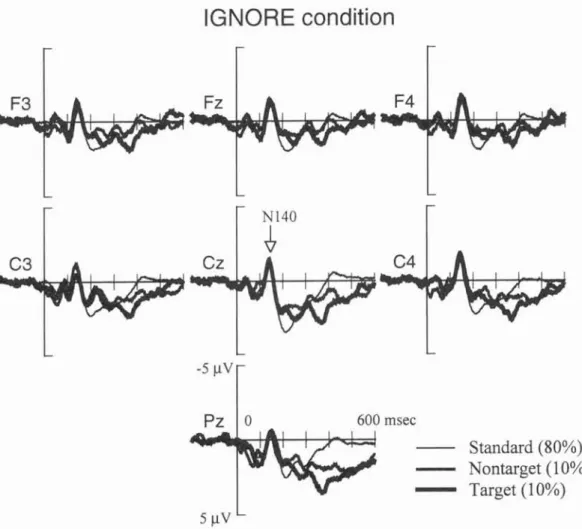
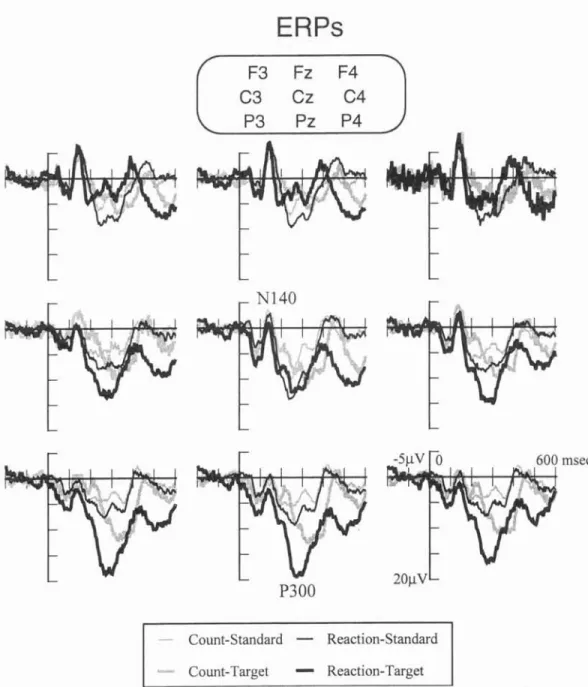
1.2.4 Sympathetic Skin Response (SSR) 1.2.5 Event-Related Brain Potentials (ERPs) 1.2.6 N140...
1.2.7P300...
1.3 LIMITATIONS OF THIS STUDY 1.3.1 Sympathetic Innervation
1.3.2 Averaging Method 1.3.3 Subjects
1.3.4 Emotions of Subjects...
1.4 SIGNIFICANCE OF THIS STUDY
1 1 6 6 6 7 7 7 7 7 8
8 8 9 10 10
CHAPTER 2 REVIEW OF LITERATURE...
2.1 SYMPATHETIC NERVOUS SYSTEM AND ELECTRODERMAL ACTIVITY...
2.1.1 Emotional Sweating and Electrodermal Activity...
2.1.2 Neural Control of the Electrodermal Activity...
2.1.2.1 Spinal and reticular control of the electrodermal activity...
2.1.2.2 Hypothalamo-limbic control of the electrodermal activity...
2.1.2.3 Cortical control of the electrodermal activity ...
2.1.3 Behavioral Correlates of the Electrodermal Activity...
2.1.3.1 Orienting response (OR)...
2.1.3.2 2.1.3.3
Emotions
Voluntary movements
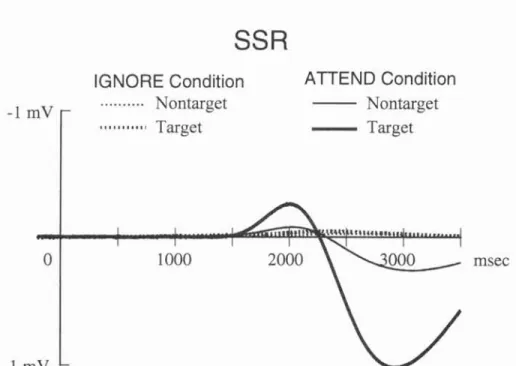
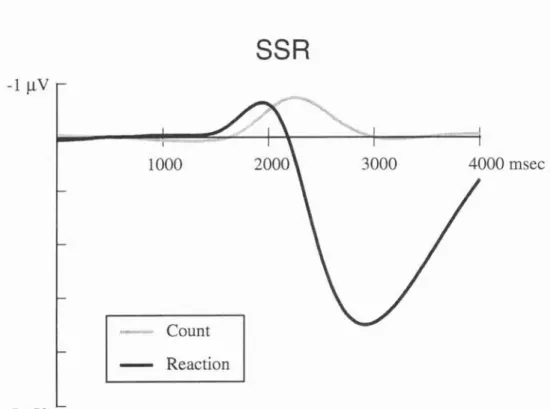
2.1.4 Sympathetic Skin Response (SSR)...
2.2 INFORMATION PROCESSING IN THE SENSORY-MOTOR SYSTEM OF THE BRAIN...
11 11 11 12 13 14 16 19 20 25 26 27
2.2.1 Reaction Time (RT) and Electroencephalogram (EEG) 2.2.2 Event-Related Potentials (ERPs) .
2.2.2.1 N140 component of somat 2.2.2.2 P300 components of ERPs 2.2.2.3 Significance of P300...
2.2.2.4
N140 component of osensory ERPs
Significance of P300 Go- and NoGo-P300
28
29
30
31
33
41
42
2.3 SYMPATHETIC SKIN RESPONSE AND P300: INTEGRATION BETWEEN THE SYMPATHETIC NERVOUS SYSTEM AND THE BRAIN...
2.3.1 EDA and P300 as OR
2.3.2 SSR and Automatic Processing.
2.3.3 SSR and Controlled Processing
2.3.4 Does SSR Have the Relations with Voluntary Movements?
44 45 46 47 49
CHAPTER 3 ISSUES TO BE INVESTIGATED, PARAMETERS, AND
METHODS IN THIS STUDY...54
3.1 AIMS, ISSUES AND RATIONALES IN THIS STUDY...54
... 54 3.1.1 Experiment 1
3.1.2 Experiment 2 3.1.3 Experiment 3 3.1.4 Experiment 4 3.2 PARAMETERS 3.3 METHODS AN
METHODS D DATA ANALYSES
3.3.1 Subjects and Apparatus 3.3.2 Stimulation
3.3.3 Recording Analogue Data for Each Parameter
3.3.3.1 3.3.3.2 3.3.3.3 3.3.3.4 3.3.3.5
3.3.4 Data Storage
Electroencephalogram (EEG) Electro-oculogam (EOG) Electromyogram (EMG)
Sympathetic skin response (SSR).
intervals (only Experiment 3)
3.3.5 Data Analyses 3.3.5.1
J ..