修 士 学 位 論 文
題 名
Cultiv劉tionofmicrobialcommHnitiesfyomahotspring andinter負ctionamongmicrobesinvolvedinsulfurcycling
邦題:温 泉微生物群集の実験室内での培養 と硫黄循環に関与する微生物の相互関係(英 文)
指導教授 松浦 克美 教授
平 成23年1月7日 提 出
首都大学東京大学院
理工学研究科 生命科学専攻 学修番号09881333
氏 名 原 なつみ
温 泉 微 生 物 群 集 の 実 験 室 内 で の 培 養 と 硫 黄 循 環 に 関 与 す る 微 生 物 の 相 互 関 係
環 境微 生物 学研 究 室 原 な つ み 温泉地 では、 しば しば温泉水 の流 れに沿 って微 生物 が群集塊 を形成 して生育 して い る。 こ のよ うな温泉微生物群集 内で は、構成種間 の密 な関係が存在す ると考 え られて いる。長野県 中房温泉(弱 アルカ リ硫黄泉)に は、70℃‑80℃ の温泉水 の流水 中に無 色〜灰色〜 白色の微 生物群集塊が発達 してい る。 その集塊 の主要 な構成種 は、好熱性匂 π諺cα1ε5目の硫 黄酸化細 菌(3〃 のア1乃ッ4ア09ε刀∫ゐ∫z4〃z属)で、本菌 は好気的代謝 によって硫化 水素 また は硫 黄 を酸化 し、
硫 酸イオ ンを生成す る。 また同時 に7乃θ朋odθ5πの わαc'顔〃〃2sp.やC磁 伽10アoわ伽2sp.に 近縁 な 細菌 も共存 して お り、嫌気的代謝 に よ り硫酸 イオ ンまたは硫 黄 を還元 し硫化水素 を生成 す る ことが知 られて いる。 これ らの細菌 は硫黄化合物 の循環 に必要 であ るが、酸素 の要 求性 と感 受性 といった点で相容れ ない反応で あ る。 これ らの代謝が 同時 に起 ってい る事 を示 すのは環 境 にお ける硫 黄循 環 を考 える上 で重要 であ る。 そこで本研 究で は、温泉地 か ら採取 した微 生 物 群集塊 を実験 室内で安定 に培養 ・維持す る ことを試 み、集塊 を構成 してい る細 菌の代謝 が 同時に起 こる条件 を探 る事で、環境 中に発達 してい る微生物群集塊 内での硫黄代謝系 を明 ら かにす るこ とを 目的 とした。
長野 県中房温泉 で採 取 した微 生物 群集塊 を嫌気 条件 で実験 室 に持 ち帰 った。温泉水成 分 を 模 した人工温泉水 中で嫌気 的 に培養 す る と、培養液 中 に硫化水 素の生成 が確 認で きた。 この こ とか ら集塊 中の嫌気 的代謝 による硫酸 イオ ンまた は硫 黄の還元が あった と言 える。次 に、
培養瓶 に注射針 を刺 し、注射針 の穴(孔 径0.5mm)を 通 して培養瓶 の気相 が大気 とゆっ く り交換で きるような微好気条件 で培養 した ところ、 白色集塊 で は培養 開始後 す ぐに培養液 中 の硫化水素濃度 お よび硫酸 イオ ン濃度 の上 昇が観察 された。 この上昇 は約100時 間 に渡 って 続 いた。硫化水素濃度 の上昇 か ら、 このよ うな微好気条件で も嫌気的代謝が発揮 され た こと を示 してい る。 また、硫化水素濃度 が低下せ ず硫酸 イオ ン濃度 が上昇 したのは集塊 内に存在
した硫 黄 を硫 黄酸化細菌が酸化 し硫酸 イオ ンを生成 させ、 同時 に硫 黄 を還元す る細菌が硫 化 水 素 を生成 したた めだ と予想 さ る。一方、 白色 を呈す る硫黄 の蓄積量が少 ない灰色集塊 では 培 養開始後す ぐに硫 化水素濃度の低下が観察 され、 同時 に硫 酸 イオ ン濃度 の上昇が確認 され た。48時 間後 か らは硫 化水素濃度 は検 出限界以下の低 い濃度 を保 ち、硫酸 イオ ン濃度 も大 き な変化 は見 られず一定 を保 っていた。 この結果 よ り、 白色集塊 に は持 ち込 みの硫 黄 による細 菌への電子供 与体の供 給が あった ことが支持 され、灰色の集塊 内の硫 黄循環 を維持 す る培養 を行 うに は電 子供与体 を加 える必要 があ るこ とが示 唆 された。
さらに 白色集塊 の培養 を続 ける と、硫酸 イオ ン濃度 は上昇 を続 ける ものの硫化 水素濃度 は 低下 していった。 これ は硫 黄酸化細 菌の利用 で きる硫 黄が枯 渇 し、人工温泉 中の硫 化水素 を 消費 しは じめ るの と同時に、硫酸 イオ ン/硫 黄還 元細 菌の活性 も低 下 して きた ことが考 えら れ る。
今 回実験室 内において微生物群集塊 を培養 し、集塊 中の硫酸 イオン と硫化水素 を生成 す る 細菌 の活性 を同時に検 出で きる条件 を見 いだす ことがで きた。適切 な酸素供給条件下 では、
これ らの細菌 間でそれ ぞれが行 う好気的代謝 と嫌気 的代謝 に よる硫黄化合物 の授受が進行 し て い ることを強 く示唆す る結果で あ る。
Cultivation of microbial communities from a hot spring and interaction among microbes involved in sulfur cycling
Natsumi Hara 2011
Department of Biological Sciences Graduate School of Science and Engineering
Tokyo Metropolitan University
Contents
Abstract ...
Introduction...
Materials and Methods •
Results ...
Discussion ...
References ...
Figure captions • • • •
... 4
... 6
... 9 ...
13
...
17
... 22
...30
Abstruct
In the hot springs, microbes often forms the microbial communities along the flow of the hot spring water and they grow. It is thought that a close relation of the composition
interspecies exists in such a hot spring microbial communities. At Nakabusa hot spring (slight alkaline sulfur hot spring) located in Nagano Prefecture, clear-, gray-, and white- microbial communities develope in 70-80°C flowing water. The major component species of these communities is themophilic Aquilicales sulfur oxidizing bacteria (Sulfurihydrogenibium sp.), this bacteria produce sulfate to oxidize sulfide or elemental sulfur by aerobic metabolism. It was identified the coexistence of
Thermodesulfobacterium sp. and Caldimicrobium sp., they produce sulfide to reduce sulfate or elemental sulfur by anaerobic metabolism. These aerobic and anaerobic bacteria are neccesary for sulfur compounds cycling, however it is happend
contradictory reactions at the same time. It is important for thinking about the sulfur cycling in the environment to show that these metabolizing has happened at the same time. In this study, to clarify the sulfur cycling in microbial communities, we searched the conditions that microbes can metabolize under, in the way to maintain the stable- culture of the hot spring microbial communies in the laboratory.
The microbial communities "streamer" were collected and brought to the laboratory under anaerobic. The cultivation under anaerobic condition in artificial hot spring water, the sulfide production was observed. This result indicated the there are the reduction of
sulfate or elemental sulfur in the streamers by anaerobic metabolism. Then, Under
microaerobic condition; through the injection needle(hole diameter 0.5 mm) to the
butyle cap; the sulfide and sulfate production were observed in white color streamers.
This production continued for 100 hours. This result indicated that anaerobic metabolism was exercised even under microaerobic. It was suggested that why the increasing of sulfate without the consumption of sulfide, is the existence of elemental sulfur which can be used sulfur oxidizing and sulfur reducing bacteria. On the other hand, in clear and gray color streamers which are not contained so elemental sulfur, the the sulfide concentration was decreased immediately and the sulfate was increased at the same time. After 48 h, the sulfide concentration was kept low level and the sulfide concentration was not changed dramatically. From these results we suggested that electron donor by elemental sulfur was supplied in white color streamer, and it is need to supply the electron donor to achieve the long time stable-culture in the clear and gray color streamers.
Moreover, to continue the cultivation of white color streamers, the sulfate concentration was increased by contrast the sulfide concentration was decreased. It was supported that sulfur oxidizing bacteria consumed the sulfide because of the lack of elemental sulfur, and sulfate/sulfur reducing bacteria was lost own activities.
In this experiment, we could find the conditions that sulfur oxidizing bacteria and sulfate/sulfur reducing bacteria can active under, at the same time in the streamers.
These results indicated the transfer of sulfur compounds were happened by aerobic and
anaerobic metabolism.
Introduction
Microorganisms play an important role in the groval cycle of various elements. Sulfur is an essential nutrient for organisms and it is cycling though a variety of sulfur
compounds on earth (12). In sulfide-rich environments, some microorganisms play a major role in energy production and the maintenance of the microbial community by utilization and cycling of sulfur (20). Sulfur-rich hot spring streams provide favorable conditions for the development of microbial communities, which contain
physiologically and phylogenetically different groups of bacteria, such as chemotrophic sulfur bacteria, cyanobacteria, and phototrophic bacteria (8).
At Nakabusa hot spring in Nagano prefecture, Japan, DGGE based phylogenetic analyses of sulfur metabolizing bacteria were previously reported at different
temperatures of the microbial communities . In the hot spring, the bacterial community structure dramatically changes with temperatures at the border at 70°C, because of no phototrophic bacteria above 70°C. In this high temperature environment, microbial
`streamer' develop in hot spring water flow. This microbial streamer was dominated by Aquificae-like microbes and Thermodesulfobacteria-like microbes (23). Phylogenetic
analysis of gene for sulfur metabolizing enzymes indicated the existence of bacteria
related to Thermodesulfobacteria (7,33), and Sulfurihydrogenibium (2,4,24,28,30) in
Nakabusa hot spring (Hase,2006 unpublished).
Sulfur oxidizing bacteria
(i)Chemolithotrophic sulfide oxidation
Obligate chemolithotrophs need an inorganic source (elemental sulfur, sulfite,
thiosulfate or sulfide) for energy, and CO2 as their carbon source. Oxygen is a universal electron acceptor used by the colorless sulfide oxidizers. The important reactors
involved in chemolithotrophic oxidation of sulfide and sulfur under aerobic conditions can be summarized as:
S0 + 3/202+ H20 ---* SO42- + 2H+
H2S + 202 —p SO42- + 2H+
(ii)Sulfurihydrogenibium sp.
A taxonomic study was reported on a novel thermophilic, chemolithoautotrophic, sulfur-oxidizing microaerophilic Sulfurihydrogenibium strains isolated from terrestrial hot springs. The strains can use elemental sulfur, sulfite, thiosulfate, ferrous iron or hydrogen as electron donors, and oxygen as electron acceptor. Some strains are also able to grow anaerobically, with elemental sulfur, arsenate and ferric iron as electron acceptors. The temperature growth range was 50-80°C and mildly alkaline.
Sulfate/sulfur reducing bacteria
(i)Chemoorganotrophic sulfate/sulfur reduction
A variety of compounds could serve as electron donors and often simultaneously as
carbon sources for sulfate/sulfur reducing bacteria. In addition to sulfate and sulfur,
most species of sulfate/sulfur reducing bacteria can utilize thiosulfate and sulfite as
electron acceptors.
(ii) The phylum Thermodesulfobacteria
The phylum Thermodesulfobacteria represents one of the deeply branching lineages within the domain Bacteria and comprises rod-shaped, anaerobic, thermophilic, sulfate reducing bacteria. The group of microorganisms belonging to the genus
Thermodesulfobacterium uses lactate and pyruvate as electron donors, and sulfate or thiosulfate as electron acceptors. In recent reports, it was clarified that some genus are not capable of sulfate reduction, and these bacteria can use elemental sulfur as electron acceptors.
These bacteria use sulfur compounds as electron donors or acceptors to obtain energy.
In a previous study from our laboratory, they suggested that in the microbial streamers over 70°C, sulfur oxidizing bacteria and sulfide reducing bacteria consume and produce sulfide in situ, and they establish sulfur cycling. To clarify these suggestions, they maintained the streamers and measured sulfide concentration in vitro for 100 minutes.
This in vitro sulfide-monitoring experiments indicated slight sulfide generation by sulfate reducing bacteria under anaerobic condition, and rapid consumption of sulfide by sulfur oxidizing bacteria under aerobic conditions. However, the relationship among sulfur oxidizing bacteria and sulfate reducing bacteria in situ has not been clear yet.
In this study, to find the conditions to maintain the microbial community structure to
metabolize sulfur compounds for a long term, we tried to maintain the stable co-
cultivation system of the aerobic bacteria and the anaerobic bacteria. In order to clarify
their interspecies relationship, experiments were performed from the aspect of the
suitable air conditions and the source of nutrition for the streamers.
Materials and Methods
Sampling site
The sampling site was Nakabusa (1400 m altitude, 36°23'10"N, 137°45'00"E), a slightly alkaline and sulfide containing hot spring, located in Nagano Prefecture, Japan.
Three types of streamers were collected at the site. Details of the sampling sites are summarized in Table 1 and Fig. 1. The microbial streamer samples were kept in a sterile plastic tube (50 ml) with spring water to prevent the oxidation of samples in anaerobic bags. Spring water for inorganic analyses was immediately fixed with the same volume of 90 mM zinc acetate solution for determination of sulfide concentration. All tubes capped were transferred to the laboratory within 1 day after sampling. Spring water samples for inorganic analyses were stored at 4°C until use.
The microbial mat and streamer samples for molecular analysis were kept in 1.5 ml centrifuge tubes and then frozen at -20°C until DNA extraction.
DNA extraction
About 0.1 g of the streamer was mixed in 1.5 ml tubes with 700111 of TPM buffer (50 mM Tris-HC1 pH7, 20 mM MgCI, 1.7% polyvinylpyrrolidone K25), about 100 mg of zilconic-silicon beads (0.1 mm dia.) and about 25 mg of beads (1.0 mm dia.). Cell were lysed by shaking with a bead beater at 2,500 rpm for 1 min (MINI-BEADBEATER, FULLTECH). After centrifugation at 15,000 rpm for 2 min at 4°C, the supernatant was discarded. 7001A of phenol based lysis buffer (5 mM Tris-HC1 pH7, 5 mM EDTA, 0.1%
SDS, 6% water-sat-phenol) was added to the precipitate, then beating at 2,500 rpm for
1 min and centrifuged 15,000 rpm for 2 min. 500111 of Tris-sat-phenol was added to the supernatant, then mixed for 3 min and centrifuged 15,000 rpm for 5 min. The aqua phase was moved into a new tube, and 100111 of 5 M NaCI and 80111 of 10% CTAB/0.7 M NaC1 was added and incubated at 65°C for 20 min. 600111 of CIAA (chloroform-
isoamyl alcohol; 24:1 v/v) was added, mixed for 3 min and centrifuged 15,000 rpm for 5 min. Then the aqua phase was purified with 600111 of PCI (phenol-chloroform-isoamyl alcohol; 25:24:1 v/v/v) and 600µ1 of isopropanol. After centrifugation at 15,000 rpm for 10 min, the supermatant was discarded, and pellets were washed with 70% ethanol and then dried by vacuum. DNA was resuspended in 30p1 TE and stored at 4°C.
PCR amplification of the 16S rRNA gene
DNA fragments encoding the 16S rRNA of domain Bacteria were amplified using the
following primers: 27F2-FAM (5'-AGAGTTTGATCMTGGCTCAG-3') (3)and 907R2
(5'-CCGTCAATTCCTTTRAGTTT 3') (4). PCR amplifications were performed with
50µ1 volumes containing 1-10 ng of template DNA, 1 xEX Taq buffer (Takara Shuzo,
Kyoto, Japan), 25 mM each of MgC12, 2.5 mM each of deoxynucleoside triphosphate,
25 pmol each oh the primers, 1.25U of EX Taq DNA polymerase (Takara Shuzo, Kyoto,
Japan), and super-pure distilled water. PCR amplifications were performed with initial
denaturation at 95°C for 1 min, followed by 25 amplification cycles of 94°C for 30 sec,
52°C for 45 sec, and 72°C for 1 min, and a final extension at 72°C for 4 min. PCR
products were analyzed by electrophoresis in 1% agarose (in 1XTAE) gels containing
ethidium bromide (1 µg m1-1). PCR products were stored at 4°C until use.
PCR purification
The PCR products were purified with the QlAquick PCR Purification Kit (QIAGEN).
Restriction enzyme treatment of 16S rRNA gene fragments Amplifide fragment were digested with the restriction enzyme MspI.
Restriction digests consisted of 4p1 enzyme mixture (1111 of l Oxbeffer [TOYOBO], 1111 of 0.1% BSA[Takara], 0.5111 volumes containing 5U of restriction enzyme
MspI[TOYOBO], and 1.5111 of super-pure distilled water) and 6111 volumes containing 20ng of amplified PCR product. DNA was digested at 37°C overnight under light interception and then stored at -20°C until use.
T RFLP analysis of 16S rDNA
The lengths of the T RFs from the amplified rDNA products were determined by ABI 3730 (Applied Biosystems 3130/3130x1 Genetic Analyzer ), as follows. 0.3µ1 of
digested DNA was mixed with 151A1 of the mixture (HIDI formamide-size standard mix;
40:1 v/v). Then, this mixture was denatured at 94°C for 3 min and immediately chilled
on ice and analysed. The lengths of fluorescently labeled T RFs were determined by
comparison with internal standards by using Peak Scanner Software v. 1.0 (Applied
Biosystems).
Analytical methods
In situ temperature and pH of hot spring water were measured with the electrode method using a thermometer (Check tempi by HANNA) and a pH sensor (Twin pH, AS ONE). Sulfide concentration was measured colorimetrically by the methylene blue formation method (Cline, 1969). Sulfate concentrations were analyzed by ion chromatography (LC-20AD SP, Liquid Chromatograph, SHIMADZU, Japan) and determined by using Chromatpac (SHIMADZU, Japan).
Cultivation conditions
An artificial hot spring water was prepared to simulated Nakabusa hot spring water.
The concentrations of sulfide, SO42-, Cl-, PO42-, and CO32- in the artificial hot spring water for cultivation were, 0.3 mM, 0.5 mM, 1 mM, 1 mM, and 1 respectively (Table 2). The microbial streamers were collected at Nakabusa hot spring and kept in hot spring water at room temperature until use. 2.0 g(wet weight) of the microbial streamers were incubated in 100 ml of artificial hot spring water in glass vial at 70°C with
reciprocal shaking (100 rpm). Air phases ware set up for anaerobic, aerobic, and
microaerobic conditions. Anaerobic condition was achieved by replacing the gas phase
with N2 gas and the vial was capped with a butyl rubber and an aluminium cap. Aerobic
condition was achieved by the air phase and the vial was capped by butyl rubber and
aluminium cap. For the microaerobic condition, the air phase was replaced with N2 gas
and capped, then the cap was stuck with a 00.5 x25 mm injection needle.
Results
Characteristics of microbial communities over 70°C
The T-RFLP fingerprintings of bacteria 16S rRNA gene fragments revealed 4 termina restriction fragments ^^^ from 6^-7^^C streamer samples. ^ tech basis ofDNA sequences detected from Nakabusa hot spring, T-RFs with sizes of 95 and 270 would be derived from ^u^^ nibium azorense, that of 117 would be derived from
Thermus kawarayensis, and that of 154 would be derived from Caldimicrobium sp..
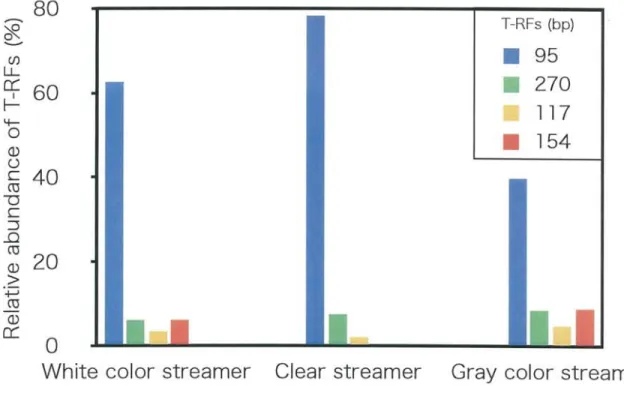
Three types of streamer were collected in 17 November 2010. From T-RFLP analysis, relative abundance of]6^r^7^/\ gene fragments of different types uf the streamer was revealed as shown in Fig. 2. The T-RFs with the sizes of 95, 270, and 117 were present in all samples. At this time, the T-RF of 154 was not detected in the clear streamer. ^^
FlFo^und27O were present in most of the streamers. In white streamer, sum of the
peak at 95 and 270 bp account for 69% in the total, 117 bp 4%, and 154 bp 6%. In clear streamer, the bands of 95 and 270 bp account for 86% in the total, and 117 bp 2%. In the gray streamer, the bands of 95 and 270 bp account for 48% in the total, 117 bp 5%, and l54bp0Y6.^^Hi^^ derived from sulfate reducing bacteria ^^enno^^un^^boc/^r/um sp. was not detected in this experiments. These streamers developed the different
sulfide concentration areas. White color streamer was observed in lower level sulfide concentration area than gray. And the difference was observed by the sampling date in the sulfide concentration. To be winter, to be higher level concentration in all areas.
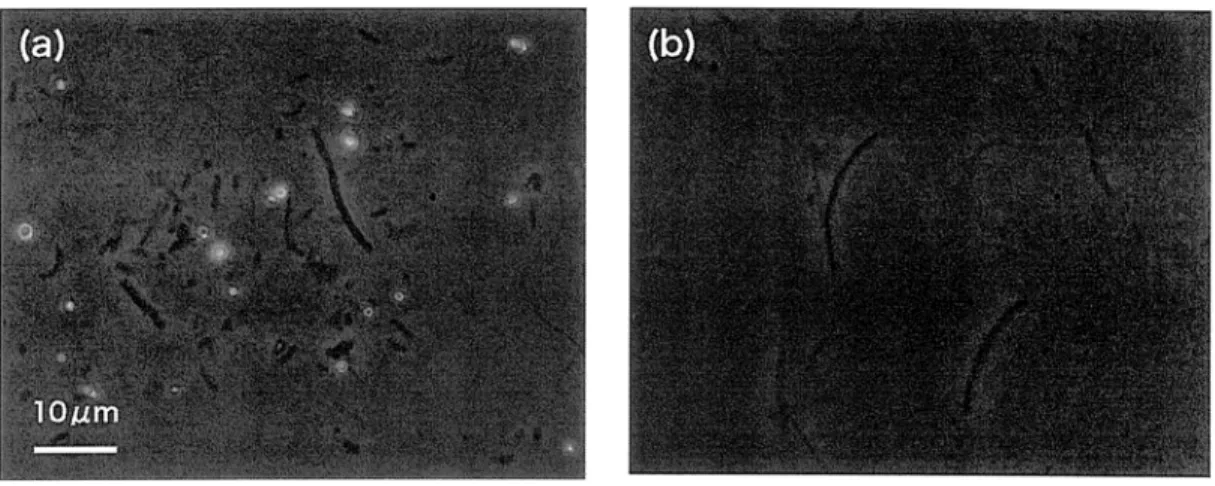
Then, microscopic observation indicated that white color streamer contained a lot of
sulfur globules in the streamer shown as Fig. 3.
Detection of sulfide producing ability
Nakabusa hot spring was a slightly alkaline, sulfide-containing water. In this experiment artificial hot spring water like real Nakabusa hot spring water was used.
Consumption and production of sulfide by the white color streamer in artificial hot spring are shown in Fig. 4. Under anaerobic conditions, the sulfide concentration increased immediately from the beginning of incubation, and after 336 h incubation gradually increased to 250 11M. On the other hand, under initially aerobic conditions, the sulfide concentration decreased below detection limit within 1 h after incubation and did not increase. Supplemental addition of 1 mM of lactic acid, 1 mM of Mg2+, 3 mM of N H4+, and trace elements, in to the artificial hot spring water did not affect the results under anaerobic condition (Fig. 5). Under initially aerobic conditions, the sulfide
concentration decreased below detection limit as observed in Fig. 5; however the sulfide concenration increased after 200 h of incubation.
02 effects during sulfide producing period
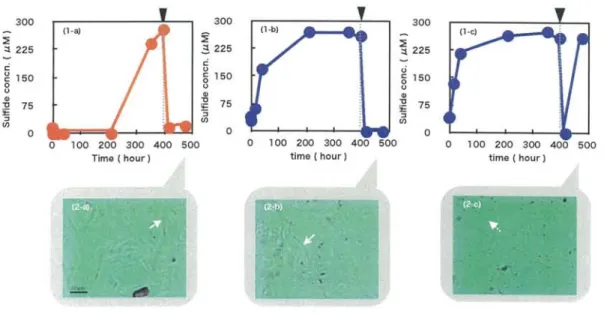
After 400 h cultivation under initially anaerobic and aerobic conditions, 02 was supplied through an injection needle. The sulfide concentration change was shown in
Fig. 6, and arrows indicate the time of injection. Under initially aerobic conditions, the sulfide production was observed after 330 h incubation. Supplementation of 02 at the time decreased the sulfide concentration. Microscopic observation after 500 h
incubation indicated the existence of a large number of long rod cells (over 101..tm long) which could be Sulfurihydrogenibium sp.. Under anaerobic conditions, the sulfide concentration was increased to 27511M until 400 h. After the supplementation of 02,
sulfide concentration decreased in two samples tested. After 500 h incubation, change of
the sulfide concentration were different between different samples. When there are more large shape bacteria like SOB (Fig. 6; 2-b) in the microscopic observation, the sulfide concentration was decreased. When there are more small shape bacteria like SRB (Fig.
6 ; 2-c), the sulfide concentration was transiently decreased by 02 supplementation.
These results suggested that the number of bacteria in microbial community effected the sulfide production under microaerobic condition.
Sulfide production and consumption under microaerobic conditions
Change in sulfide concentration under microaerobic conditions was shown in Fig.7.
Initial concentration of sulfide in the artificial hot spring water was 256 laM. The sulfide concentration was decreased at the beginning of incubation with 25 h, then increased.
Consumption of sulfide concentration was observed again after 75 h incubation. After that, small increase and decrease in sulfide concentration ware observed.
The relationship between Sulfide and Sulfate concentration
At Nakabusa hot spring over 70°C, microbial communities of three color types; white, clear and gray, were observed.
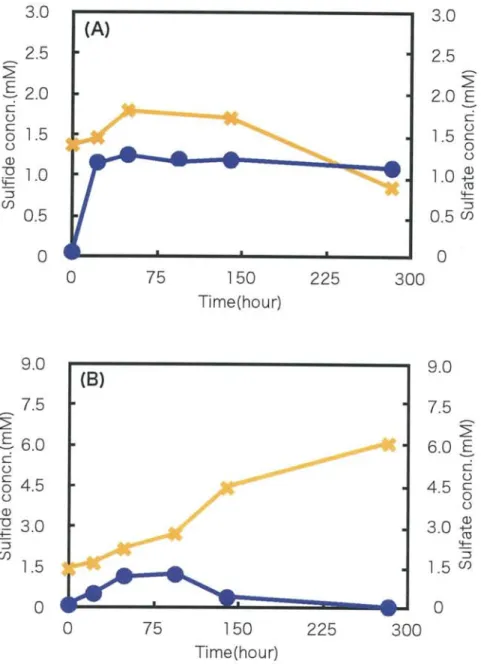
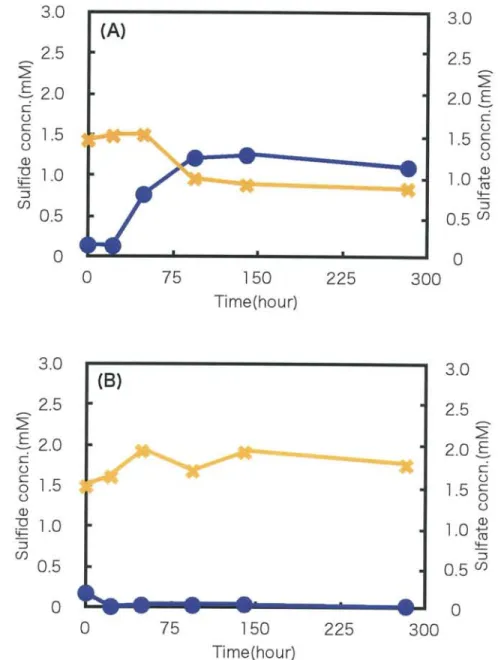
The sulfide and sulfate concentrations in 2 weeks cultivation of white streamer were
shown in Fig. 8. Changes of under anaerobic conditions, the sulfide concentration was
increased for 24 h and kept around 1.3 mM until 275 h. The sulfate concentration was
increased until 48 h, after that it was decreased. Under microaerobic conditions, the
sulfide concentration was increased to 1.3 mM until 48 h, then after 140 h cultivation it
was decreased. The sulfate concentration was increased from 1.5 mM to 6.0 mM for 275 h cultivation.
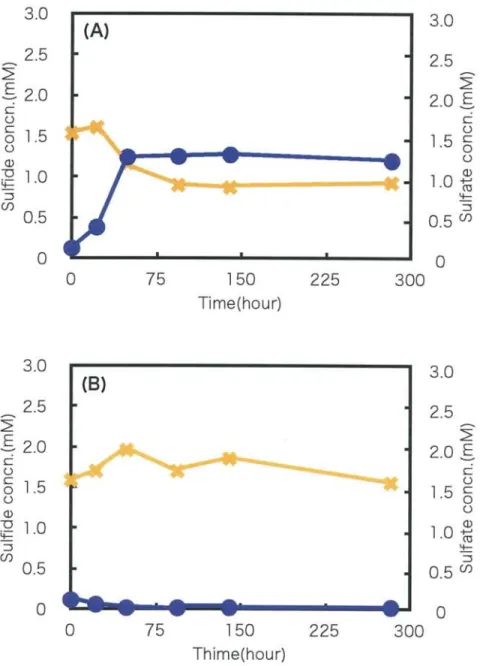
The sulfide and sulfate concentration for 2 weeks cultivation of the clear streamer was
shown in Fig. 9. Under anaerobic conditions, the sulfide concentration was increased
between 25 h and 100 h after the start of incubation, then kept 1.3 mM until 275 h. The
sulfate concentration was kept about 1.5 mM for 48 h cultivation, after that it was
decreased and kept 0.8 mM until 275 h. Under microaerobic conditions, the sulfide
concentration was decreased immediately below detection limit. The sulfide and sulfate
concentrations for 2 weeks cultivation of gray streamer was shown in Fig.10. Under
anaerobic conditions, the sulfide concentration was increased to 1.3 mM until 48 h and
kept its concentration until 275 h. The sulfate concentration was decreased after 24 h
cultivation, and it was kept at 0.8 mM until 275 h. Under microaerobic conditions, it
was the same tendency as the results in the clear streamer.
Discussion
I found conditions for maintaining active microbial communities from natural habitat with sulfur oxidizing bacteria and sulfate/sulfur reducing bacteria in vitro for several days. Microbial streamers in Nakabusa hot spring over 70°C were dominated by sulfur oxidizing bacteria (Sulfurihydrogenibium sp.) and sulfate/sulfur reducing bacteria (Thermodesulfobacterium sp., Caldimicrobium sp.). Sulfur oxidizing bacteria (SOB) can use elemental sulfur and sulfur compounds as electron donors, and oxygen as electron acceptor (2,24,28,30). On the other hand, sulfate/sulfur reducing bacteria (SRB) can use sulfate or elemental sulfur as electron acceptors, and some organic compounds as electron donors (19,11,29). Sulfurihydrogenibium sp. is microaerophilic bacteria, and Thermodesulfobacterium sp. and Caldimicrobium sp. are anaerophilic bacteria. Therefore, it might be happened both aerobic and anaerobic metabolisms are present at the same time in the microbial community. To clarify the ways to metabolize sulfur under such different conditions, we tried to find the conditions to maintain the microbial community with the native form of streamer, and searched the conditions that ware appropriate for growth of both bacteria in the in vitro communities.
Microbial composition and sulfur globules
In the samples around 70°C at Nakabusa hot spring, it was found that there were three main species; Sulfurihydrogenibium sp., Thermus sp., and Caldimicrobium sp.
Sulfurihydrogenibium sp. and Thermus sp. (3,14) known as sulfur oxidizing bacteria,
and Caldimicrobium sp. is a species of sulfur reducing bacteria. From the results of T-
RFLP in Fig. 2, the major components in all streamers investigated were sulfur oxidizing bacteria. By comparing between white and gray color streamers, it was
formed that the white streamer had more SOB than the gray one, on the other hands, the gray streamer had more SRB than the white one.
In a previous report (Hase,2006 master degree thesis, Tokyo Metropolitan University), DGGE results indicated the presence of Thermodesulfobacterium sp. However, in the T RFLP analysis, the T RF of Thermodesulfobacterium sp. was not observed in this study.
Anaerobic cultivation
Sulfurihydrogenibium sp. is a microaerophilic bacterium and Thermodesulfobacterium sp. and Caldiicrobium sp. are anaerophilic bacteria. Air conditions are important factors to maintain all those bacteria. Under anaerobic conditions, all samples produced sulfide
and kept high level sulfide concentration after about 150 hours of cultivation. It might be caused by sulfate/sulfur reducing bacteria (SRB), such as Thermodesulfobacterium sp. and Caldimicrobium sp. By comparing between white and gray color streamers (Fig.
8, 10(A)), white streamer produced sulfide faster than the gray one. It suggested that there were many sulfur globules in white color streamers, and therefore Caldimicrobium sp. may be used elemental sulfur as electron acceptor and produced sulfide. On the other hand, sulfate production was detected only in white streamer even if 02 was not present. It was reported that some anaerobic bacteria are able to catalyze the
disproportionation of elemental sulfur to sulfate and sulfide (10,16). Therefore, I
suggested that Caldimicrobium sp. produced sulfate by using elemental sulfur which
was contained in white streamer by the disproportionation.
Aerobic conditions
From Fig. 4 and 5, under aerobic conditions, by the addition of Mg2+, NH4+, lactic acid and trace element, sulfide production was detected after 200 hours of cultivation. An isolation medium of Sulfurihydrogenibium sp. contains magnesium and nitrogen source in the forms of Mg2+ and NH4+ . From the result of sulfide producing, I can suggest that initial sulfide was oxidized by sulfur oxidizing bacteria (SOB), such as
Sulfurihydrogenibium sp. by 02. This condition would be suitable for SOB, which consumed 02 as electron acceptors withi 200 hours after the start of cultivation, and then made the air phase anaerobic. After that, SRB in the streamer stayed alive, produced sulfide because of suitable anaerobic condition for SRB after the 02 consumption.
These results indicate that the supplements, Mg2+ and NH4+, improved the growth of SOB, resulted in the sulfide production in the streamer. Therefore, I suggested that supplements(Mg2+, NH4+, lactic acid and trace element) were need to maintain the SOB and SRB activities.
From Fig. 6, I suggested that the community structure in the streamer was changed during 400 hours cultivation, and this change affected the sulfide production. In the streamer which has more SOB (Fig. 6:2-a, 2-b), supplemented 02 and sulfide were consumed immediately by SOB. On the other hand, in the SRB dominant streamer (Fig.
6:2-c), the effect of supplemental 02 was transient on the sulfide concentration. These
results suggested that the community structure and the present of 02 were important for
maintaining the sulfur cycling in the streamer.
Microaerobic conditions
Fig. 7 shows that microaerobic condition was important for forming the microbial community containing both aerophilic and anaerophilic bacteria. This condition would make both aerobic and anaerobic parts in the streamer. Therefore we were able to observe the sulfide production and consumption due to the sulfate oxidizing and sulfide reducing activities by SOB and SRB. Changes of sulfate and sulfide concentrations
indicated the presence of the sulfur cycling in the streamer in vitro. Fig. 8, 9 and 10(B), show the sulfur cycling in white, clear and gray color streamer, respectively under the microaerobic conditions. All three streamers produced sulfate in 48 hours. However after 48 hours, only the white color streamer continued to produce sulfate,
accompanying the production of sulfide. I suggested that elemental sulfur was used by SOB as an electron donor and SRB as an electron acceptor. Microscopic observation indicated that the white color streamer had more elemental sulfur in the form of sulfur globules within the microbial community. Moreover, it was reported that some sulfur oxidizing bacterial species can store sulfur intracellular as an electron donor and oxidize
it (17,27,31). SOB in the white color streamer stored sulfur in the natural habitat and oxidized it to sulfate in this cultivation system. It is not clear in Sulfurihydroganibiur
sp. of this system yet, but some thermophilic sulfur bacteria have the system that oxidize intracellular sulfur (5,26). At the same time, Caldimicrobium sp. produced
sulfide by reducing elemental sulfur. The other streamers might contain sulfide too,
however there were less amount of elemental sulfur to be able to be used by
Caldimicrobium sp. During the additional 150 hour cultivation, the sulfide was
consumed in the white color streamer, and sulfate was produced continuously. This result suggested that SOB depleted with intracellular sulfur first and then begun to oxidize sulfide. It might be one of the reasons why there were originally a lot of SOB in the white color streamer than the gray color streamer. In natural environment, the gray color streamer would keep its community structure by continuous supply of sulfide from the spring water. During the long time cultivation such as about 300 hours, under microaerobic conditions, sulfide production did not continue because of the
consumption and lack of electron donors as organic compounds.
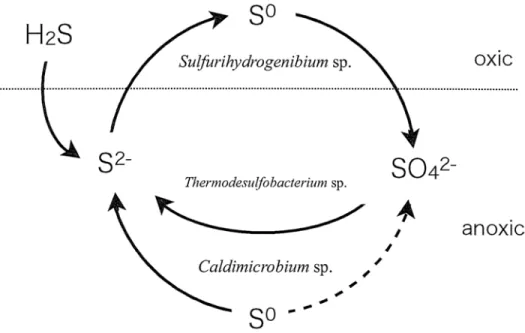
In natural environment, microbial communities developed with interaction among constitutive species. Over 70°C, chemotrophic bacteria interact through sulfur cycling to oxidize and reduce sulfur compounds. A model of microbial sulfur cycling in this part of the streamer in Nakabusa hot spring is shown in Fig. 11. To metabolize sulfur
compounds properly, air conditions and supply of electron donors are very important.
In this study, we showed that the external microaerobic conditions were a major factor to maintain both SOB and SRB activities. Under the flowing water, the streamer structure would make the outside of the streamer microaerobic and inside anaerobic.
However, even if the air phase was suitable, both activities did not continue because of
the lack of electron donors. At low sulfide concentration, SOB accumulated elemental
sulfur by oxidizing of sulfide. Therefore, it may be the reason why we could observe
three types of streamers at the same temperature with the same dominant species. The
electron donor's continuous supply is necessary to achieve the long term cultivation
more than this study period.
References
(1)Andrea S. Pott and Christiane Dahl. Sirohaemsulfite reductase and other proteins encoded by genes at the dsr locus of Chromatium vinosu are involved in the oxidation of intracellular sulfur. Microbiology. 1998. 144:1881-1894
(2)Andrew H. O'Neill, Yitai Liu, Isabel Ferrera, Terry J. Beveridge and Anna- Louise Reysenbach. Sulfurihydrogenibium rodmanii sp. nov., a sulfur-oxidizing chemolithoautotroph from the Uzon Caldera, Kamchatka Peninsula, Russia, and emended description of the genus Sulfurihydrogenibium. International Journal of Systematic and Evolutionary Microbiology. 2008. 58:1147-1152
(3)Bjornsdottir, S. H., Petursdottir, S.K., Hreggvidsson, G.O., Skirnisdottir, S., Kristjansson, J.K. Thermus islandicus sp. nov., a mixotrophic sulfur-oxidizing bacterium isolated from the Torfajokull geothermal area. International Journal of Systematic and Evokutionary Microbiology. 2009. 59:2962-2966
(4)Flores, G.E., Liu, Y., Ferrera, I., Beveridge, T.J., Reysenbach, A-L.
Sulfurihydrogenibium kristjanssonii sp. nov., a hydrogen- and sulfur-oxidizing
thermophile isolated from a terrestrial Icelandic hot spring. International Journal of
Systematic and Evolutionary Microbiology. 2008. 58:1153-1158
(5)Grimm, F., Dobler, N., Dahl, C. Regulation of dsr genes encording proteins responsible for the oxidation od stored sulfur in Allochromatium vinosum. Journal of Bacteriology. 2010. 156:764-773
(6)Guiral, M., Tron, P., Aubert, C., Gloter, A., Iobbi-Nivol, C., Gludici-Orticni, M- T. A membrane-boud multienzyme, hydrogen-oxidizing, and sulfur-reducing complex from the hyperthermophilic bacterium Aquifex aeolicus. Journal of Biological
Chemistry. 2005. 280(51),pp. 42004-42015
(7)Henry, L.A., R. Devereux, J.S. Maki, C.C. Gilmour, C.R. Woese, L. Mandelco, R. Schauder, C.C. Remsen and R. Mitchell. Characterization of a new thermophilic
sulfate-reducing bacterium Thermodesulfovibrio yellowstonii, gen. nov. and their origins deep within the bacterial domain. Arch. Microbial. 1994a. 161:62-69
(8)Hiraishi. A, Umezawa. T, Yamamoto. H, Kato. K, Maki. Y. Change in Quinone Profiles of Hot Spring Microbial Mats with a Thermal Gradient. 1999. Applied and Environmental Microbiology. p.198-205
(9)Hugenholtz, P., Pitulle, C., Hershberger, K.L., Pace, N.R. Novel division level
bacterial diversity in a Yellowstone hot spring. J. Bacteriol. 1998. 180:366-376
(10)Janssnen, P.H., Schuhmann, A., Bak, F., Liesack, W.
disproportionation of inorganic sulfur compounds by the sulfate-reducing bacterium Desulfocapsa thiozymogenes gen. nov., sp. nov. Archives of Microbiology. 1996.
166:184-192
(11)Jeanthon, C., L'Haridon, S., Cueff, V., Banta, A., Reysenbach, A-L., Prieur, D.
Thermodesulfobacterium hydrogeniphilum sp. nov., a thermophilic,
chemmolithoautotrophic, sulfate-reducing bacterium isolated from a deep-sea hydrothermal vent at Guaymas Basin, and emendation of the genus
Thermodesulfobacterium. International Journal of Systematic and Evolutionary Microbiology. 2002. 52:765-772
(12)Kimberley Tang, Vikrama Baskaran, Mehdi Nemati. Bacteria of the sulphur cycle: An overview of microbiology, biokinetics and their role in petroleum and mining industries. 2009. Biochemical Engineering Journal. 44:73-94
(13)Kimura. H, Mori. K, Nashimoto. H, Hanada. S, and Kato. K. Short
Communication: In Situ Biomass Production of a Hot Spring Sulfur-Turf Microbial
Mat. Microbes Environ. 2010. 25:140-143
(14)Kurosawa, N., Itoh, Y.H., Itoh, T. Thermus kawarayensis sp. nov., a new member of the genus Thermus, isolated from Jaoaese hot springs. Extremophiles. 2005. 9:81-84
(15)Liu, W.-T., Marsh, T.L., Cheng, H., Forney, L.J. Characterization of microbial doversity by determing terminal restriction fragment length polymorphisms of genes encording 16S rRNA. Applied and Environmental Microbiology. 1997. 63:4516-4522
(1 6)Lovley, D. R., Phillips, E.J.P. Novel processes for anaerobic sulfate production from elemental sulfur by sulfate-reducing bacteria. Applied and Environmental Microbiology. 1994. 60:2394-2399
(17)Maki, growing in
Y. Biological oxidation of sulfide and elemental sulfur by A-type sulfur-turf hot springs effluents. J. Gen. Appl. Microbiol. 1987. 33:123-134
(1 8)Maki, Y. Factors in habit preference in situ of sulfur-turfs growing in hot springs effluents: dissolved oxygen and current velocities. J. Gen. Appl. Microbial. 1986.
32:203-313
(1 9)Miroshnichenko, M.L., Lebedinsky, A.V., Chernyh, N.A., Tourova, T.P., Kolganova, T.V., Spring, S., Bonch-Osmolovskaya, E.A. Caldimicrobium rimae gen.
nov., sp, nov., an extremely thermophilic, facultatively lithoautotorophic, anaerobic bacterium from the Uzon Caldera, Kamchatka. Internationnal Journal of Systematic and
Evolutionary Microbiology. 2009. 59:1040-1044
(20)Mostafa S. Elshahed, John M. Senko, Fares Z. Najar, Stephen M. Kenton, Bruce A. Roe, Thomas A. Dewers, John R. Spear, and Lee R. Krumholz.
Bacterial Diversity and Sulfur Cycling in Mesophilic Sulfide-Rich Spring.
2003. APPLIED AND ENVIRONMENTAL MICROBIOLOGY. p5609-5621.
(21)M.T Suzuki and S.J. Giovannoni. Bias caused by template annealing in the amplification of mixtures of 16S rRNA genes by PCR. Appl. Environ. MicrobioL 1996.
62:625-630
(22)Muyzer, G., A. Teske, C. O. Wirsen, and H. W. Jannasch. Phylogenetic relationships of Thiornicrospira species and their identification in deep-sea
hydrothermal vent samples by denaturing gradient gel-electrophoresis of 16S rDNA fragments. Arch. Microbiol. 1995. 164:165-172
(23)Nakagawa T. and M. Fukui. Phylogenetic characterization of microbial mats and streamers from a Japanese alkaline hot spring with a thermal gradient. J. Gen. Appl.
Microbial. 2002. 48:211-222
(24)P. Aguiar, T. J. Beveridge and A.-L. Reysenbach. Sulfurihydrogenibium azorense, sp. nov., a thermophilic hydrogen-oxidizing microaerophile from terrestrial hot springs in the Azores. International Journal of Systematic and Evolutionary Microbiology. 2004.
54:33-39
(25)Pickering, I. J., George, G. N., Yu, E. Y., Brune, D. C., Tuschak, C., Overmann, J., Beatty, J. T. & Prince, R. C. Analysis of sulfur biochemistry of sulfur bacteria using X-ray absorption spectroscopy. Biochemistry. 2001.40:8138-8145
(26)Pott, A. S., Dahl, C. Sirohaem sulfite reductase and other proteins encoded by genes at the dsr locus of Chromatium vinosum are involved in the oxidation of intracellular sulfur. Microbiology. 1998. 144:1881-1894
(27)Prange, A., Chauvistre, R., Modrow, H., Hormes, J., Truper, H.G., Dahl, C.
Quantitative speciation of sulfur in bacterial sulfur globules: X-ray absorption spectroscopy reveals st least three different species of sulfur. Microbiology. 2002.
148:267-276
(28)S. Nakagawa, Z. Shtaih, A. Banta, T. J. Beveridge, Y. Sako and A,-L.
Reysenbach. Sulfurihydrogenibium yellowstonese sp. nov., an extremely thermophilic,
facultatively heterotrophic, sulfur-oxidizing bacterium from Yellowstone National Park,
and emended descriptions of the genus Sulfurihygrogenibium, Sulfurihydrogenibium
subterraneum and Sulfurihydrogenibium azorense. International Journal of Systematic
and Evolutionary Microbiology. 2005. 55:2263-2268
(29)Sonne-Hansen, J., Ahring, B.K. Thermodesulfobacterium hverandense sp. nov., and Thermodesu fovibrio islandicus sp. nov., two thermophilic sulfate reducing bacteria
isolated from a icelandic hot spring. Systematic and Applied Microbiology. 1999.
22:559-564
(30)Takai. K, Kobayashi. H, Kenneth H. Nealson and Horikoshi.K.
Sulfurihydrogenibium subterraneum gen. nov., sp. nov., from a subsurface hot aquifer.
International Journal of Systematic and Evolutionary Microbiology. 2003. 53:823-827
(31)Yamamoto, H., A. Hiraishi, K. Kato, H.X. Chiura, Y. Maki, and A. Shimizu.
Phylogenetic evidence for the existence of novel thermophilic bacteria in hot spring sulfur-turf microbial mats in Japan. Appl. Environ. Microbiol. 1998. 64:1680-1687
(32)Yong-Jin Lee, Alexander Prange, Henning Lichtenberg, Manfred Rohde, Mona Dashti, and Juergen Wiegel. In situ analysis of sulfur species in sulfur globules produced from thiosulfate by Thermoanaerobacter sulfurigignens and
Thermoanaerobacterum thermosulfurigenes. Journal of Bacteriology. 2007. 189(20), pp. 7525-7529
(33)Zeikus, J.G., M.A. Dawson, T.E. Thompson, K. Ingvorsen and E.C.
Hatshikian. Microbial ecology of volcanic sulphidogenesis: isolation and
characterization of Thermodesulfobacterium commune gen. nov. and sp. nov. J. Gen.
Microbiol. 1983. 129:1159-1169
Acknowledgements
I especially grateful to K. Matsuura and S. Haruta for their advice and valuable
discussion. I would like to thank S. Nagashima and S. Hanada for thier suggestion. I
thank to H. Otaki and K. Yamamoto for their help and technical suggestions. Finally, I
thank to Environmental Microbiology laboratory members for their strong support.
Tablel. Characteristics of spring water under which three kinds of streamers
ware collected at Nakabusa hot spring
Sample
Temp. (°C)
pHSulfide concentration(iiM)
White color streamer
(I) (II) (III) (IV) (VI) Clear streamer
(I) (II) (III) (IV) (VI) Gray color streamer
(I) (II) (III) (IV) (VI)
76 70 75.3 79.3 70
65
71 69.8
71
70 74.3
71 78.1
73
6.9 6.8 7.5 7.2 7
8.5 8.2 8.5 8.7 8.3
6.8 8.4 8.7 8.7 8.4
39.2 79.1 122.9 156.1 243.1
53.8 108.5
150 121.6 429.2
333.2 427.4 303.4 182.5
(I) (IV)
12 June 2010, (II ); 10 July 2010, (III); 19 August 2010, 29 October 2010, (VI); 17 November 2010
no tested
Table 2. Composition of artificial hot spring water.
1 mM NaCI
1 mM NaH2PO4 - 2H20 0.5 mM Na2SO4
(Sulfur source)
0.3 mM Na2S • 9H20 1 mM NaHCO3
Adjusted at pH 8.0
.--,
(A)
.;"4 ,;;c•..
•,;.; ,
if
J:::',- '` ..'' .;
. ,
:1
a i4 I., ...“ '
lirtYY • , ... 1 -'t r'
t :);, ,ie,'.• . '''.h. w
. ! • • ,... ..,-,.,,,,,„ , r : ;•',.,—.41..L.t: •;,,,, °
t, cp..;.7,,,,,..:,,..- •,1-.J
...i.,,,- .4..:.•
(C)
Fig. 1. Photographs of the streamer at Nakabusa hot spring.
Arrows respectively indicated White color streamer(A), Gray color streamer(B), and Clear streamer (C).
The dashed circles in small photo were sampling sites of gray color streamer and clear streamer.
32
80 ---
T-RFs(bp)
cn
^95
60 - • 270 ~
117
o• 154
a) U
a40 -
-0 C D
C 20 -
IF cu
Tb cc 0 II II
White color streamer Clear streamer Gray color streamer
Fig. 2. Relative abundance of 16S rRNA gene fragments retrieved from three
types of the microbial communities streamers on T RFLP analysis. The T RFs of
95(blue bar) and 270 (green bar) represented Sulfurihydrogenibium sp., 117(yellow
bar) represented Thermus sp., and 154(red bar) represented Caldimicrobium sp.
(a)
mum
(b)
Fig. 3. Microscopic images of microbial streamers. (a);Images of white color
streamer containing sulfur globules. (b);Images of gray color streamer.
300 ---
225 -
c 150 0 CD
75
cn
0 100 200 300 400
Time ( hour )
Fig. 4. Sulfide production and consumption by the white color streamer in artificial hot spring water. •(circle), the sulfide concentration under anaerobic cultivation. ^ ( diamond ), the sulfide concentration under aerobic cultivation.
35
300 ---
225 - C
c 150 -
O 0
0
75 - cn
0
0 100 200 300400
Time (hour )
Fig. 5. Sulfide production and consumption with the white color streamer in artificial hot spring water. .(circle), the sulfide concentration under anaerobic cultivation. • ( diamond ), the sulfide concentration under initially aerobic cultivation.
36
2 4
C 0 c 0 0
E cn
300
225
150
75
0
0 100 200 300 400 500
Time (hour )
300
225 c U
0 150
0 01 mi 75 N
0
0 100 200 300 400 500
time (hour )
2 4
0 C 0 0 m E c U)
300
225
150
75
0
0 100 200 300 400 500
time (hour )
Fig. 6. Effect of Oz supplementation changes of the sulfide concentration after supplementation of 02.
Air was supplied through an injection needle after the time indicated by arrow in graph(').
(1-a) Sulfide production and consumption under initially aerobic condition. Microscopic observation after 475 h cultivation was indicated (2-a).
(1-b,c) Sulfide production and consumption of different two samples under initially anaerobic condition. Microscopic observation after 475 h cultivation was indicated (2-b,c).
White solid arrow indicates long rod cells (2-a,b).
500
i 375
U c 250 O V N
to 125
0 100 200 300 400
Time ( hour )
Fig. 7. Sulfide production and consumption with the white color streamer under microaerobic conditions.
38
2.5 I- 2.5
ff
2.0- 2.0 E
UC
O 1.5. 1.5c
O0
0
1.0 -1.0
4- c 0
.5 -. 0.5 (1)
0--- 0
0 75 150 225 300
Time(hour)
9.0
(B)--- 9.0
7.5 --7 .5
g 6.0 --6 .0 E
o4.5 --4.50
UC 0733.0 --3.0
=4-co
u 1
.5 •.1.5 w
00
0 75 150 225 300
Time(hour)
Fig. 8. Sulfide and sulfate concentration with "white color" streamer for long term cultivation under anaerobic condition(A), and microaerobic condition(B) .
•(circle),data for the sulfide concentration. , the sulfate concentration .
39
3.0 2.5 2
g 2.0
E U
o 1.5
U N 1.0 U) 0
.5 0
3.0 2.5 2
E 2.0
U
0 1.5
U a) 1.0 n U)
0.5 0
0 75
150
Time(hour)
225
300
0 75 150
Time(hour)
225 300
3.0 2.5
2.0 g
0
1.5 0
0
1.0 1 0.5w
0
3.0 2.5 2.0 1.5 1.0
E
0 C 0 0 a) 4--(II
0.5 u) 0
Fig. 9. Sulfide and sulfate concentration with " clear" streamer for long term cultivation under anaerobic condition(A), and microaerobic condition(B).
•(circic), the sulfide concentration. , the sulfate concentration .
3.0 2.5 2
2.0 E U
o 1.5
U a)
0 1.0
(f) 0.5
0
3.0 2.5 2
2.0 E U
0 1.5
U N
0 1.0
Cl) 0.5
0
0
75 150
Time(hour)
225 300
0 75 150
Thime(hour)
225 300
3.0 2.5
2.0 g
0
1.5 0
0
1.0 0.5U'
0
3.0 2.5
2.0 E
0
1.5 0
U
1.0 %
4-
0.5 (I) 0
Fig. 10. Sulfide and sulfate concentration with " gray color" streamer for long term cultivation under anaerobic condition(A), and microaerobic condition(B).
•(circle), the sulfide concentration. , the sulfate concentration .
H2S
S2-
So
lfurihydrogenibiun'i sp.
Thermodesulfobacterium sp.
imicrobium sp .
So - -
41 ...
.
^
..,
SO42-
,
4
oxic
anoxic