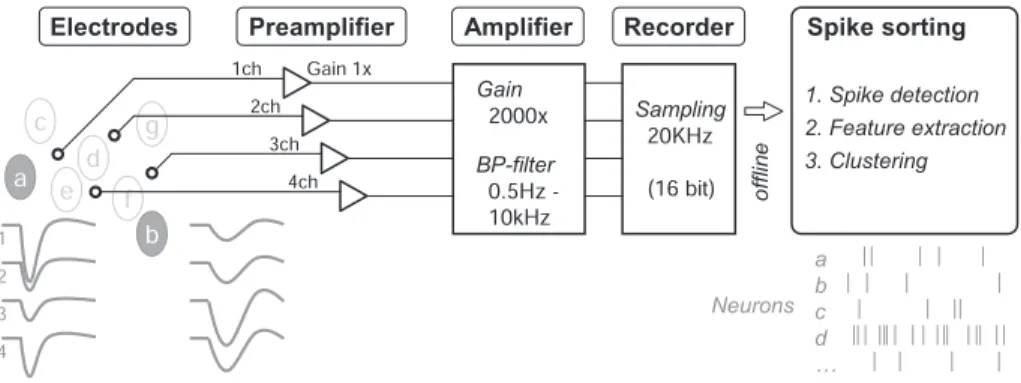
大脳皮質や海馬のような脳領域における局所回路内の信号処理の仕組みを調べるためには, 複数の電極をもちいて多数の神経細胞の発火活動を同時に検出できるマルチニューロン記録法 が有効である.ここでは同記録法の簡単な具体例を中心にして,動物の手術から,多点電極, プリアンプ,メインアンプ,記録装置,オフラインでの自動スパイク・ソーティングとスパイ ク・クラスターの修正と選択までの各段階を,順を追って詳細に解説する.
1.
は じ め に 脳の情報処理の仕組みを本質的に理解するためには, 多数の神経細胞の発火活動を同時に測定し個別に解析 することが必須である(マルチニューロン記録).欧米 ではマルチニューロン記録技術の発展と普及が目覚し く1),海馬の再活性化(reactivation/replay)現象の 発見など生物学的に重要な研究成果が次々に挙がりつ つある(例えば文献2)).一方,我が国でも,外山敬 介,小野武年,櫻井芳雄らにより独自の多チャンネル 記録実験の研究が報告されてきたが3∼5),ほとんどの 生理学研究者にとってマルチニューロン記録はまだま だ敷居が高いのが現実であろう.そのため,科研費特 定領域「統合脳」支援班(平成21年度終了)/マルチ ニューロン研究会や新学術領域研究「包括脳」支援活動 では,国内外の最新のマルチニューロン記録技術に関 する研究リソースを紹介する支援活動をおこなってい る6).それでも,電極から解析に至るまで選択肢があ まりに多過ぎるため,本格的なマルチニューロン記録 の導入例をここで詳細に解説することも有益であろう と考える.単一ユニット記録の電極本数をただ増やす よりも(1電極1細胞),近接した複数の電極から多数 の神経細胞の活動を個別に捉えるほうが(多電極多細 胞),局所回路内の細胞間相互作用を効率良く調べるこ とができる.その際,海外の既製記録システムは大変 高価なうえに研究目的に合わせて改変しにくく,ある 程度の電気生理学の知識と経験をもつ研究者には,低 予算で高性能の記録システムを自ら構築することを勧 めたい.そこで本稿では,マルチニューロン記録の具 体例として,頭部を固定した覚醒ラットの大脳皮質や ∗〒194–8610 東京都町田市玉川学園 6–1–1 海馬の細胞活動を,テトロードやシリコンプローブを 使って検出し,その信号をプリアンプとメインアンプ で増幅処理して,ハードディスク記録装置に保存した 記録データを,Mac(またはUNIX)コンピューター に組み込んだスパイク解析プログラムを使ってオフラ イン解析する,という記録実験と解析の手順を中心と して解説する(図1).2.
対 象 動 物 体重150–250 gのLong-Evans系ラットをあらかじ め実験者や実験環境に馴化させておく.イソフルラン 吸入麻酔下でラットの頭部を脳定位固定装置(ナリシ ゲ・SR-8N)に固定し,スライド式の頭部固定具(ナリ シゲ,アルミニウム製13 g)を歯科用セメント(サン メディカル・スーパーボンドC&B,クラレ・パナビア F2.0,ジーシー・ユニファストII)とステンレス製ビス (M1規格,長さ2 mm)で頭蓋骨に定位的に設置する 一次手術を施す7).小脳の硬膜上に基準referenceお よび接地ground用電極となるテフロン銀線を留置す る.頭蓋骨の記録領域とゼロ点(bregmaやlambda) はスーパーボンドを薄く塗り,シリコン印象材(松風・ デントシリコーンV)で覆っておく.手術回復後,必要 に応じて,オペラント学習課題の訓練装置(小原医科産 業)を活用して頭部固定状態での前肢のレバー操作や 舌のリッキングなどの行動課題を訓練する.簡単なオ ペラント学習課題の訓練は数日から数週間程度で完了 する.記録前日,麻酔下で頭蓋骨の電極挿入部に小さ な孔を開けて硬膜も開窓し,シリコン印象材で再び封じ る二次手術を施す.このように小動物に脳定位固定下 head-restraintで特定の行動をさせる実験系には,(1) 記録電極を高い空間精度で配置できる,(2)二光子レー図1 マルチニューロン記録と解析の概略. 近接する4つの電極から複数の神経細胞の発火活動を同時に記録し,各スパイクを細胞別に分類するオフ ライン解析をおこなう. ザー走査型顕微鏡などの測定装置も導入できる,(3)視 覚や触覚などの感覚刺激を適切に呈示できる,(4)四肢 やヒゲなどの運動発現を適切に計測できる,(5)オペラ ント学習などの行動課題を効率よく訓練できる,とい う利点がある7).一方,自由行動freely-moving動物 をもちいた慢性記録の実験系には,(1)睡眠・覚醒を含 む数日間の連続記録が可能である,(2)心理学で確立さ れた行動試験を適用できる,(3)空間認知に関連する 神経活動を解析できる,という利点がある8).しかし, 自由行動といえども,あまりに多数の記録チャンネル を盛り込んでしまうと,動物は重いマイクロドライブ とプリアンプを頭部だけで支えることになり,頭上の 導出コードの束も太くなって,動物の行動が制約され てしまう.現在,これらの問題の解決策として記録シ ステムの無線化や集積回路技術を駆使したワンチップ マイコン化などがBrain-Machine Interface技術の流 行・発達に伴ってさかんに研究されているが,本格的 な実用化にはまだ至っていない.
3.
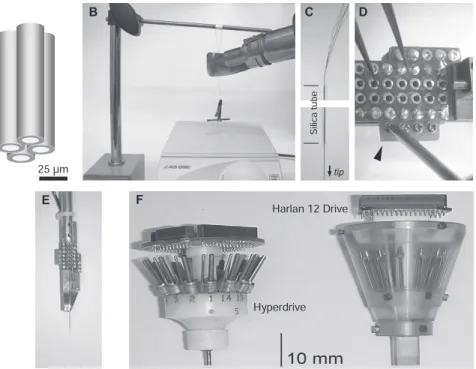
多点電極とプリアンプ 最も標準的な多点電極は,極細の被覆ニクロム線を4 本捻って作るテトロードtetrodeと呼ばれるワイヤ電極 である9)(図2A).被覆ニクロム線(A-M Systems・ポ リイミド被覆ニクロム線#761000,径25µm)約50 cm を四つ折りに吊り下げて金属クリップで留め,マグネッ トスターラーでゆっくりと捻ってから,ヒートガンで すばやく被覆を溶かしてそのまま冷ます(図2B).テ トロード専用とした解剖用ハサミで切り出して,シリ カチューブ(Polymicro Technologies・TSP100170, 外径170µm)数cmに通して写真用クリアファイルに 保存しておく(図2C).このテトロードをプリアンプ のステンレス管に装填し,被覆をコネクターネジで擦 り切って導通させるか(図2D),鈍刃のメスで被覆を剥 いで導通性ペースト(GC electronics・Silver Print) でコネクターに接着して導通させ,長さを整えて瞬間 接着剤で固定する(図2E).急性・亜急性記録にはテ トロードを解剖用ハサミで直角にまたは斜めに切断し てそのまま使用できるが,慢性記録には必要に応じて 切断面に金めっきを施して電極抵抗を0.3–0.4 MΩに 下げるとよい.記録当日,まず記録領域を再び露出し てマニピュレーターで保持したテトロードの先端を脳 表面に接触させ,2%アガロースを流し込んでテトロー ドを埋め込む形にする.次にテトロードを脳内の目的 部位までゆっくりと挿入してしばらく静置する.乾燥 を避けるためにパラフィンを上塗りしておく.電極の 挿入直後には傷ついた細胞の連発ユニットが目立つが, 1∼数時間後にはさまざまな大きさのユニット(振幅約 0.2–0.5 mV)が安定して観察されるようになる10).一 方,個々のユニットを数時間から数日に渡って追跡す る慢性記録では,記録段階において,できる限り分離 した(S/N比の良い)クラスターが形成されると期待 される座標にいかにして電極先端をうまく留置できる かが重要なポイントとなり,その作業にはµm単位で の微調整が必要である.そのため現在の主流としては, 複数のテトロードをマイクロドライブ(David Kopf・ Neuro-Hyperdrive(図2F,左),Neuralynx・Harlan Drive(同右)など)に取り付けて埋め込み,手術後数 週間かけて個別に電極位置の微調整をおこなう方法が とられている8).特に安定した長期間記録をおこなう 場合や,100ユニットクラスの大規模なマルチニュー ロン記録を試みる場合は,計測領域付近での電極移動 量は数µm/日とする研究者も少なくない.そこで,微図2 テトロードの作製と装填.(A)テトロードの構造.(B)ワイヤを縒り合せて固める.(C)シリカチュー ブに通す.(D)プリアンプに装填してワイヤを各端子に接続する.(E)急性記録用テトロード・プリアン プの完成.(F)慢性記録用マイクロドライブ.左)David Kopf Instruments社Hyperdrive(製造終了). 直径約1.5 mmの範囲内に12本のテトロード電極と2本の基準電極を挿入できる.深さ方向の可動範囲は 約10 mm.右)Neuralynx社Harlan 12 Drive.合計12本の電極を挿入・調整できる.
調整の精度を向上させるために,電極の位置調整を電 動化したシステムなども提案されている11).多電極が 装填可能なマイクロドライブを研究室単位で量産する ことは少々難しい.このようなデバイスは現在のとこ ろ国内ではほとんど生産されておらず,日本の微細加 工技術を応用した製品開発が待たれる分野といえよう.
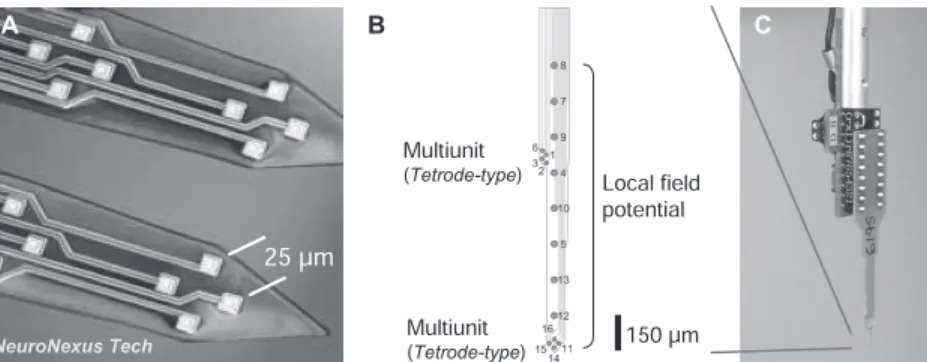
シ リ コ ン プ ロ ー ブ silicon probe( 例 え ば Neu-roNexus社「ミシガンプローブ」)は半導体製造技術を もちいて長さ数mm,幅70–100µm,厚さ15µm程 度の板状のシャンクに多チャンネルの電極パターンを 配置したものであり,挿入時の組織へのダメージが比 較的少なく,多数の細胞活動の時空間分布を正確に捉 えることができる.多数の近接する神経細胞のスパイ クを正確に分離するためには25µm間隔のテトロード 型配列が適している(図3A).100µm間隔で直列16 チャンネルのシリコンプローブは局所フィールド電位 の測定には最適であるが10),1チャンネルごとにスパ イク・ソーティングするために多数の近接細胞のスパ イク解析にはやや不向きである.最近,礒村らがデザ インしたシリコンプローブは,先端0µmと800µmの 位置に皮質深層と浅層の発火活動を検出するテトロー ド型配列を有し,先端から150µmおきに1,200µm まで局所フィールド電位を検出する電極8点を配し ており(図 3B),大脳皮質の複数の細胞活動と脳波 活動を同時に解析することができる7)(NeuroNexus・ LFP8+TetrodeSD1).急性記録用シリコンプローブ はプリアンプのICソケット部に差し込んで接続する (図3C).実験後はSCAT希釈液やコンタクトレンズ 洗浄液に一晩浸けて洗浄すると繰り返し使用可能であ る(超音波洗浄はしてはならない).また,記録後にシ リコンプローブに直接電流を流すことにより脳内の記 録位置のマーキングも可能である12). これらの多点電極は近接するプリアンプ(市販または 自作)に接続し,各測定点で検出した微弱な電位変化を メインアンプへ伝える.プリアンプを自作する場合は, 4チャンネルのオペアンプ(図4;Texas Instruments・ TLC2274CNまたはCD)の各IN(+)端子に電極から の入力を接続し,IN(−)端子と短絡させたOUT端子か らリッツワイヤ(New England Wire・ N28-36E-400-2)を通じてメインアンプへ導出する(図4B;voltage
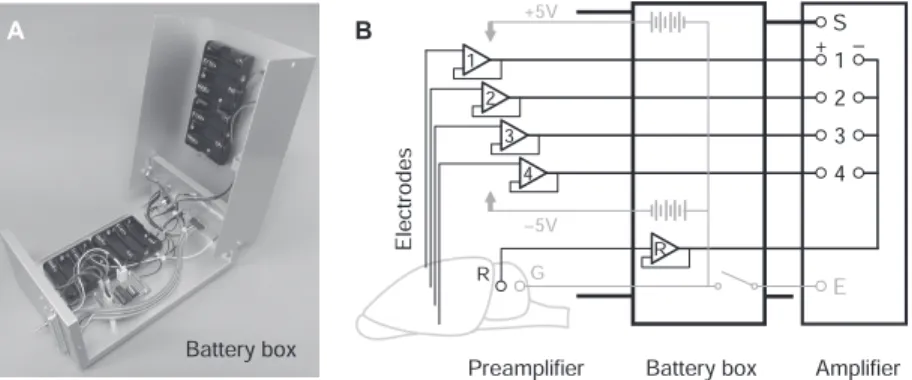
図3 シリコンプローブ.(A)NeuroNexus社16chシリコンプローブの一例.2シャンクに4つのテトロード型 配列を有する(同社ホームページ:転載許諾済).(B)1シャンクに2つのテトロード型配列と8つの局所 フィールド電位用電極を有する特別設計プローブ(同社).(C)急性記録用シリコンプローブ・プリアンプ. 図4 自作プリアンプ.(A)オペアンプTLC2274の入出力信号(4チャンネル)と電源の配線.IN(−)端子と OUT端子は短絡させる.(B)組み立て例.16chシリコンプローブをICソケットで受け,オペアンプ4 個に配線する.基準電極と接地電極からのリード線はそれぞれRソケットとGソケットで受ける. follower回路).電極からオペアンプの入力までの配線 の長さは極力短くし,ハンダ付けにはフラックスは使 わない.バッテリーボックス内の充電式ニッケル水素 電池で±4.8 V(4本直列×2系統)の電圧をオペアン プに供給すると(図5A),ゲイン1倍のDC信号がメ インアンプへ出力される.動物に埋め込んだ基準およ び接地用電極は,多点電極の挿入前に短い導線でプリ アンプのコネクターに接続しておく(図5B).必要に 応じて基準電極の信号にも同型のオペアンプをかます とよい.
4.
信号増幅とデータ保存 プリアンプから出力される信号は,ゲインを1,000– 2,000倍,ローカットおよびハイカット・フィルター を0.5 Hzと10 kHzに設定した多チャンネル・メイン アンプ(日本光電・MEG-6116)をもちいて基準電極 の信号との間で差動増幅する.各チャンネルの信号を (+)端子に,基準電極の信号を(−)端子に,動物の接地 用電極をE端子に,プリアンプ本体や各導線のシール ド部をS端子に接続する(図5B).この幅広いフィル ター設定はユニット活動(1 kHz周辺)と局所フィール ド電位(1∼250 Hz)を同時に記録するためである10). 動物本体,行動実験装置,記録測定装置,実験台それぞ れの配線や接地を工夫して,ユニットがオシロスコー プ上で明瞭に目視できる程度までノイズを極力減らす (約100–150µV以下).各チャンネルの増幅信号は多 チャンネル記録装置(TEAC・LX-120)を使って,サ ンプリング周波数20 kHz・信号分解能16 bitでオー バーレンジしないように(±5 V程度)コンピューター のハードディスクに連続記録する.16チャンネルで1 時間記録すると約2 GBのバイナリーデータ(datファ イル)として保存される.図5 メインアンプへの信号入力.(A)電池ボックス.プリアンプ(オペアンプ)に電力を供給するとともに, プリアンプからの各信号をメインアンプへ中継する.予備電源付.(B)電極からプリアンプ,電池ボック ス,メインアンプへの配線図.各チャンネルの信号はメインアンプの+端子へ,基準電極(R)の信号は− 端子へ入力する.接地電極(G)はアース(E)端子に落とすか浮かせるか選択可能とする.プリアンプか らの配線経路を覆うようにシールドしてS端子につなげる. 図6 自動スパイク・ソーティング.(1)スパイク検出.各トレースにバンドパス・フィルターをかけ,一定の閾 値を超える振幅の大きな電位変化をスパイク候補として検出する.(2)特徴抽出.検出したスパイクの波形 の特徴をチャンネルごとに算出する.(3)クラスタリング.自動クラスタリング用プログラムKlustaKwik やEToSを使用し,多次元の特徴空間におけるスパイクの分布に基づいて,個々のスパイクをクラスター に分類していく.
5.
自動スパイク・ソーティング 近接する複数の電極から記録される多数の神経細胞 のスパイク情報をオフラインで取り出す.一般には, 各スパイクの検出spike detection,特徴抽出feature extraction,クラスタリングclusteringの3段階を経 て個別の神経細胞の発火活動を分離していく13)(図6). 第一段階では,近接する電極の各チャンネルのトレー スに800 Hz∼5 kHz程度のバンドパス・フィルターを かけたうえで,ある閾値を超える電位変化をスパイク 候補として検出する.この閾値の設定は大変重要であ り,予備解析においてS/N比に応じた最適な値を決定 しておくことが望ましい.第二段階では,検出したス パイクの各チャンネルにおける振幅や波形の特徴を数 値化する.例えば,主成分分析principal component analysis(PCA)をもちいてスパイクごとに各チャン ネルの第一主成分,第二主成分,第三主成分を算出す る.テトロードの場合,スパイク振幅とPCA3成分をスタリングのアルゴリズムとしてClassification EM
(CEM)法を採用し,処理速度を上げるために数々の
工夫を凝らした自動スパイク・クラスタリング用のソ フトウェアであり13),Rutgers大学のBuzs´aki研究室 のホームページから無償で入手できる14).最近,理化 学研究所の竹川高志は,スパイクの特徴抽出にウェー ブレット変換をもちいて,クラスタリングにt分布を 仮定した(ロバスト)変分ベイズ法を採用することに より,さらに高精度のスパイク・クラスタリングを実 現できることを示した15).この研究の過程で竹川が開 発したEToSは,スパイク検出の段階でFIRフィル ターかメキシカンハット・フィルターかを,特徴抽出の 段階でPCAかウェーブレット変換かを,クラスタリ ングの段階でEM法か変分ベイズ法かを選択すること ができ,処理速度を下げないように多くの改良を加え た高性能の無償ソフトウェアである16).これらのソフ トウェアを活用するためには,できる限り多くの速い コア(CPU)と十分なメモリを備えたコンピューター が必要であり,竹川はエントリーマシンとしてアップ ルのMacPro(例 2.93 GHz Quad-Core Intel Xeon 2個,2 GBメモリ8個搭載)を推奨している.もちろ んUNIXマシンでも問題なく動作する.目安として, 数時間分の記録データからのスパイク・クラスタリン グに数時間から1晩ほど要すると考えておいたほうが よい.
6.
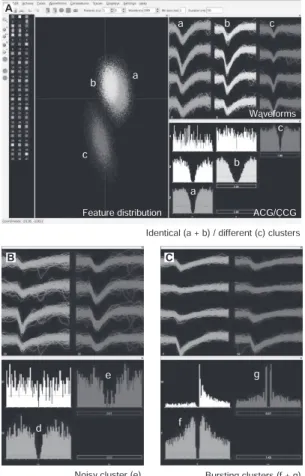
スパイク・クラスターの修正と選択 自動スパイク・クラスタリングの結果は完璧ではな く,解析者の目によりクラスターを修正し選択するこ とが必要である.ここではBuzs´aki研究室で開発され たソフトウェアKlustersを使って,自己相関図 auto-correlogram(ACG)や相互相関図cross-correlogram (CCG)を指標にクラスターを修正・選択する14, 17). まずKlustersに全スパイクの特徴パラメーターの情 報と自動スパイク・クラスタリングの結果を読み込む. スパイクはクラスターごとに色分けされ,各クラスター られるものは同一細胞由来のクラスターとみなして結 合する(図7A,a + b).竹川のEToSでは,ノイジー なクラスターの除外や類似クラスターの結合を客観的 に提案する情報がログファイル内に記載される.また, 波形が異なっていても,相互相関図の基準付近に間隙 が存在し,一方にのみ短潜時のピークがみられるもの は同一細胞のバースト発火と考えて結合する(図7C, f + g).当然,結合したクラスターの自己相関図にも 不応期を示す間隙がみられるはずである.スパイク数 が少なく判断のつかないクラスターは単独では解析に 使わないほうが無難であろう.ちなみに,相互相関図 において不応期がなく短潜時で一方に偏った単一ピー クは,2細胞間の単シナプス性興奮応答monosynaptic excitationか,第3の興奮性細胞common driverによる2細胞の時間差のある同期的発火であることを示唆 している3, 7).このような操作を繰り返すと,通常,1 テトロード配列あたり5∼15細胞の発火活動を分離す ることが可能である.ただし,どの解析にも常に偽陽 性スパイク(誤ったスパイクの混入)と偽陰性スパイク (正しいスパイクの欠落)が生じることには留意すべき である.最後に,得られたスパイク情報はMATLAB などをもちいて目的に応じて各種の解析を進めていく (他書参考).なお,スパイク情報に局所フィールド電 位や動物行動などの関連情報を重ねて表示するために は,Buzs´aki研究室で開発されたNeuroScopeという ビューアーを活用すると大変便利である14, 17).
7.
お わ り に 本稿で紹介した記録解析システムは,国内メーカー の普及製品を活用するように考慮して構成した.多点 記録電極は国内でもいくつかの研究室で開発中であり 国産化が待たれる.メインアンプや記録装置などは同 等品を使用すると大幅なコストダウンも可能であろう. 実験装置や材料はバイオリサーチセンター(株)より 入手可能である(プリアンプやバッテリーボックス完 成品を含む).図7 スパイク・クラスターの修正と選択.(A)マニュアル・クラスタリング用プログラムKlusters.自動スパ イク・ソーティングにより分類された数十個のクラスター(左端)のうち,ここではクラスターa,b,c に注目する.スパイクの特徴分布図(中央)は,クラスターaとbの分布が重複し,クラスターcの分布 とは異なることを示している.各チャンネルのスパイク波形(右上)は,やはりクラスターaとbの波形 (振幅や形状)が似ていることを示している.各クラスターの自己相関図(ACG)(右下カラー)には基準 付近(±1 ms)に不応期に相当する間隙がみられ,それぞれ単一細胞由来のクラスターであることを示し ている.クラスターaとc(またはbとc)の間の相互相関図(CCG)(右下白色)には基準付近の間隙が みられず,両者が独立していることを示している(基準点の1ビンは検出不能のため除外).一方,クラス ターaとbの相互相関図には基準付近の間隙が存在するため,両者は同一細胞由来と解釈して一つのクラ スターに結合する.(B)ノイジーなクラスターの除去.クラスターeの自己相関図の基準付近には少なく ないスパイク活動がみられるため,異なる細胞由来のスパイクが混入していると解釈して解析対象から外 す.(C)バースト発火するクラスターの結合.クラスターfとgの相互相関図は,基準付近に間隙があり, 一方(右側)に短潜時のピークがみられるため,両者はバースト発火してスパイク波形がわずかに変化す る同一細胞由来のクラスターであると解釈して結合する.両クラスターのスパイク波形は,振幅が若干異 なるものの,形状や振幅比(4ch> 1ch > 3ch > 2ch)が一致することにも注意. なお,過去に本誌で紹介されたマルチニューロン記 録に関連する日本語解説を付記しておくので併せて参 考にされたい18, 19). 謝 辞 本稿の執筆にあたり,マルチニューロン記録法の導 入と改良を支えてくださったG. Buzs´aki,K. Harris, 深井朋樹,竹川高志,相澤秀紀,塚田稔,酒井裕各氏
ronal connectivity in cat visual cortex, J. Neu-rophysiol., Vol.46, No.2, pp.191–201
4) Kobayashi, T. et al. (1997): Task-dependent representations in rat hippocampal place neu-rons, J. Neurophysiol., Vol.78, No.2, pp.597– 613
5) Sakurai, Y. (1994): Involvement of auditory cortical and hippocampal neurons in auditory working memory and reference memory in the rat, J. Neurosci., Vol.14, No.5, pp.2606–2623 6) http://www2.bpe.es.osaka-u.ac.jp/multineuron/
multineuron resource/index.html
7) Isomura, Y. et al. (2009): Microcircuitry coor-dination of cortical motor information in self-initiation of voluntary movements, Nat. Neu-rosci., Vol.12, No.12, pp.1586–1593
8) Takahashi, M. et al. (2009): A code for spatial alternation during fixation in rat hippocampal CA1 neuron, J. Neurophysiol., Vol.102, No.1, pp.556–567
9) Gray, C.M. et al. (1995): Tetrodes markedly improve the reliability and yield of multiple single-unit isolation from multi-unit record-ings in cat striate cortex, J. Neurosci.
Meth-Brain Res. Meth-Brain Res. Protoc., Vol.9, No.2, pp.122–129
13) Harris, K.D. et al. (2000): Accuracy of tetrode spike separation as determined by simultaneous intracellular and extracellular measurements, J. Neurophysiol., Vol.84, No.1, pp.401–414
14) http://osiris.rutgers.edu/frontmid/indexmid. html
15) Takekawa, T. et al. (2010): Accurate spike sorting for multi-unit recordings, Eur. J. Neu-rosci., Vol.31, No.2, pp.263–272
16) http://etos.sourceforge.net/
17) Hazan, L. et al. (2006): Klusters, NeuroScope, NDManager: a free software suite for neuro-physiological data processing and visualiza-tion, J. Neurosci. Methods, Vol.155, No.2, pp.207–216 18) 石金浩史(2008):多細胞同時記録実験と解析,日 本神経回路学会誌, Vol.15, No.3, pp.203–208 19) 龍野正実(2010):自由行動中の動物からの多電 極同時記録とそのデータ解析—理論から実験に フィールドを広げた一研究者のケーススタディ—, 日本神経回路学会誌, Vol.17, No.3, pp.124–138