九州大学学術情報リポジトリ
Kyushu University Institutional Repository
ペプチド基質を用いたミトコンドリアプロセシング ペプチダーゼの基質認識機構に関する研究
新留, 琢郎
Graduate School of Sciences, Kyushu University
https://doi.org/10.11501/3094234
a
CD
A Study on Substrate Recognition of Mitochondrial Processing Peptidase by Using Synthetic Peptides
T AKURO Nil DOME
CONTENTS
Page Summary * * * ** * * * * * * * * * * *** * * **** **** ** ** * 1 Abbreviations * * * ** * * ** * * ** * * * ** * ** ** ****** **** 2 Intrcxluction ** * *** ** * * * ********* *** ** * ***** ** 3 Materials and Methods ********************************* 6
Results ********************** *********** 10
Discussion * * * * * ** * * * * * * * * * ** ** * ** * ** * ** ** ** 27 References ***************************** **** 33 Acknowledgements ********************************* 36
SUMMARY
The mitochondrial processing peptidase \\'as purified from the n1atrix fraction or bovine liver mitochondria. The purified peptidase contained two protein subunits of 55kDa (a) and 52 kDa (f3) as determined by SDS-PAGE. To analyze the peptidase acti\'ity
quantitatively and identify the structure responsible for the recognition of the presequcnces by the peptidase, I used synthetic peptide, which corresponds to entire portion or presequence of mouse malate dehydrogenase together with its amino terminal mature portion, as a substrate. The peptide was cleaved at correct site by the peptidase, and the cleavage could be kinetically analyzed. For the effective cleavage of substrate by MPP, the peptide with 11 and 5 amino acid residues from the cleavage site to the amino- and carboxyl-terminal sides, respectively, was sufficient. Two of basic amino acids in the peptide, the distal arginine residue at position -10 and the proximal one at position -2 relative to the cleavage site, had an important role for the cleavage by the peptidase. The proximal arginine residue was not replaceable by lysine residues, therefore, the peptidase would specifically recognize the arginine residue not a simple basic amino acid. The position of the proximal arginine residue was strictly determined at -2 position. On the other hand, the distal arginine residue was replaceable by lysine residue, therefore, the peptidase would recognize the residue as a simple basic amino acid. The position of the distal arginine residue was not strictly determined. This result suggests the presence of a flexible region between the distal and proximal arginine residues of the substrate. The portion bet:v�'een the distal and proximal arginine residues of malate dehydrogenase presequence was replaceable by the glycine rich region, which expected to have a function as a flexible region, in adrenodoxin presequence. These results suggest that the flexible region contributes to give a appropriate distance between the arginine residues to fit substrate recognition site on the peptidase.
ABBREVIATIONS
MPP, mitochondrial processing peptidase MIP, mitochondrial intermediate peptidase MDH, malate dehydrogenase
pMDH, precursor of malate dehydrogenase Ad, adrenodoxin
pAd, precursor of adrenodoxin ALDH, aldehyde dehydrogenase
pALDH, precursor of aldehyde dehydrogenase HPLC, high performance liquid chromatography.
PMSF, phenylmethylsulfonyl fluoride
HBTU, 2-( 1H-benzotriazol-1-yl)-1, 1,3,3-tetramethyluronium hexafluorophosphate HOBt, 1-hydroxybenzotriazol hydrate
DIEA, N, N' -diisopropyl ethyl amine TFA, trifl uoroacetic acid
TMSBr, trimethylsilyl bromide
INTRODUCTION
The vast majority of mitochondrial proteins are encoded by nuclear genes and synthesized as larger precursors bearing amino-terminal presequences that contain sufficient information required for their targeting and localization to the mitochondria After the import of the precursors into mitochondria, the presequences are proteolytically removed from the precursors in the mitochondrial matrix (Rosenberg et al., 1987; Verner and Schatz, 1988; Pfaner and Neupert, 1990). The cleavage of the presequences in the matrix is catalyzed by two types of processing proteases. One is the general processing protease which catalyzes the processing of most precursor proteins and named mitochondrial processing peptidase, MPP (McAda and Douglas, 1982; Conboy et al. , 1982; Miura et al., 1982; Bohni eta!., 1983; Zwizinski and Neupert, 1983; Sagara et al., 1984). The other, mitochondrial intermediate peptidase (MIP), catalyzes the second step cleavage of the two-step processing of some precursor proteins (Kalousek et al. ,
1988).
MPP has been purified to homogeneity from the mitochondria of various biological sources, Neurospora crassa (Hawlitschek et al., 1988; Schneider et al., 1990), Saccharomyces cerevisiae (Yang et al., 1988), and rat liver mitochondria (Karousek et a!., 1988; Ou et al., 1989; Kleiber et al., 1990; Isaya et al., 1991). The purified enzymes are sensitive to metal chelators such as EDT A and 1, 10-phenanthroline, although the species of the metal bound to the enzyme is still unknown. The enzymes consist of two non-identical subunits of 55-59 kDa (a-MPP) and 52 kDa CP-MPP). The cDNAs for these subunits have been cloned and the deduced amino acid sequences show a high degree of similarity of amino acid sequence (Witte et al., 1988; Jensen and Yaffe, 1988; Pollock et al., 1988; Kleiber et al., 1990; Schneider et al., 1990; Kitada et al., 1993; Paces et al.,
1993).
The most striking functional feature of MPP is its unique substrate specificity. The acti\ ity of MPP is strictly specific to mitochondrial precursor proteins. Other proteins, including mitochondrial mature proteins and secretory protein precursors, arc not recognized by MPP. This indicates that the presequence of mitochondrial protein precursors have some information for the recognition by MPP. Analysis of a large number of sequences around the cleavage site revealed no consensus amino acid sequence for the processing, except that basic amino acid residues, usually arginine, arc often observed at - 2 or -3 position relative to the cleavage site (von Heijne, 1986; von Heijne et al., 1989;
Roise and Schatz, 1988; Pfanner and Neupert, 1990). However, since arginine residues are present at multiple positions in all proteins, it is obvious that "arginine -2 or -3 rule" is not sufficient for the specific cleavage of the presequence by MPP. Thus, MPP must recognize some specific structures common to all presequence of the precursors, besides the arginine residues at -2 or -3 position. This study was intended to answer the question how MPP recognizes a large variety of mitochondrial precursor proteins and cleaves off the presequences correctly.
Several attempts have been reported to elucidate the structural requirements of the presequences for the specific recognition by MPP. Mutations, including deletions and amino acid substitutions, of the precursor proteins and inhibition experiments with synthetic peptides revealed that regions at many residues upstream and downstream from the cleavage site is important for the cleavage of the presequences by MPP. However, the common structure recognized by MPP remained unknown. To identify the structure responsible for the recognition of the presequence by MPP, property of interactions between the precursors and the peptidase, and the rate of the proteolytic reaction should be quantitatively analyzed. The activity of MPP has so far been assayed by mixing the peptidase and the precursor proteins which were translated in vitro or expressed in E.
coli cell, and by detecting the products by fluorography. Since the amount of the
precursors synthesized in the in vitro translation system is Yery low and the data obtained from the fluorography often includes some error, it is extremely difficult to obtain kinetic parameters such as reaction velocity, affinity constants, etc ..
To overcome the weakness of the present assay method, I tried to establish a quantitative and rapid method using synthetic peptides as the substrate. This has allowed me to obtain various kinetic parameters of the cleavage reaction, to change easily the amino acid sequence of the presequence, and to discuss structural requirement for the recognition by MPP in more detail. For the purpose, I chose the prescquencc of malate dehydrogenase as a model precursor because it has short presequence of 16 amino acid residues. A peptide corresponding to the presequence of the malate dehydrogenase follo\ved by appropriate length of amino terminal portion of the mature protein and its derivatives \\'ere synthesized as substrate for MPP. In this study, I showed that arginine residues at portions both distal and proximal to the cleavage site and the flexible region between these arginine residues are necessary for the processing of the precursor proteins by MPP.
MATERIALS AND METHODS
Purification of mitochondrial matrix fraction
All manipulations of purification were carried out at 4°C. Fresh bovine liver ( 1 kg) was obtained from a local slaughterhouse and canied to the laboratory in ice. The liver was homogenized in the isolation buffer containing 10 mM NaPi, pH 7.5, 0.25 M sucrose and 200 mM KCl with a Warnig blender. The homogenate was centrifuged at .500 g for 10 min. The precipitate was homogenized and centrifuged as above. The supernatants were combined and centrifuged at 8,000 g for 10 min to obtain the pellet of mitochondria The mitochondria were suspended in a hypotonic buffer containing 20 mM NaPi, pH 7.2, 0.2 f.!g/ml each of leupeptin and pepstatin and 0.1 mM phenylmethanesulfonyl fluoride (PMSF), and homogenized with a glass-Teflon homogenizer, and sonicated. The sonicated mitochondrial suspension was centrifuged at 100,000 g for 60 min. The supernatant was used as the matrix fraction.
Purification of processing peptidase
DEAE-cellulose chronu1tography. The matrix fraction was loaded to a DEAE
cellulose column (Whatman DE-52; 5.0 x 65 em) which had been equilibrated with 10 mM NaPi buffer, pH 7.0. After the column was fully washed with 10 mM NaPi buffer, pH 7.0, containing 75 mM NaCl, MPP was eluted from the column with 10 mM NaPi buffer, pH 7.0, containing 150 mM NaCl.
Aminohexyl-Sepharose 4B chromatography. Aminohexyl-Sepharose 4B was prepared by conjugating diaminohexane to cyanogen bromide-activated Sepharose 4B (Pharmacia Chemical Co.). The active fractions from the DEAE-cellulose column were pooled and applied to an aminohexyl-Sepharose 4B column (2.6 x 58 em) equilibrated with 10 mM NaPi buffer, pH7.0, containing 200 mM NaCl. The column was washed
with the same buffer, and elution was carried out in a linear gradient of NaCl from 200 to 600 mM. MPP was eluted from the column at 400 mM NaCl.
Sephadex G-150 gel filtration. The fractions containing MPP activity were pooled, concentrated to 20 ml by ultrafilters (Diaflo ultrafiltration membranes, Amicon Co.), and then loaded to Sephadex G-150 column (Pharmacia Chemical Co.; 3.0 x 120 em). The gel filtration was performed in 20 mM NaPi buffer, pH7.0.
Hydroxlapatite chromatography. The active fractions from Sephadex G-150 were collected, and applied to a hydroxylapatite column (Bio-Rad Lab.; 1.5 x 13 em) equilibrated with 50 mM NaPi buffer, pH 7.0. The column was washed with the same buffer, and elution was carried out in a linear gradient of NaPi buffer, pH 7.0, from 50 to 200 mM. MPP was eluted at 100 mM NaPi.
FPLC Mono-Q chromatography. The active fraction from hydroxylapatite were loaded to a Mono-Q anion-exchange column (Pharmacia Chemical Co.; HRS/5), which had been equilibrated with 10 mM NaPi buffer, pH 7.0, containing 100 mM NaCl at a flow rate of 1 mllmin. The column was washed with the same buffer, and elution was carried out in a linear gradient of NaCl from 100 to 250 mM.
Preparation of adrenodoxin precursor
Precursors of adrenodoxin were synthesized in a rabbit reticulocyte lysate in the presence of [35s] methionine (Pelham and Jacson, 1976). Messenger RNAs of the precursors were prepared by in vitro transcription from the cDNAs cloned into SP6 transcription vectors (Ou et al., 1986; Furuya et al., 1987). In vitro transcription and translation was carried out as described previously (Ou et al., 1986).
Assay of MPP activity using precursor protein translated in vitro Translation products containing adrenodoxin precursor were incubated with MPP
fractions in 20 mM Hepes buffer, pH7.6, containing 1.0 mM MnCl2_, 2 �tg!ml each of leupeptin and pepstatin, 0.1 mM PMSF and 0.1% Tween 20 at 30 OC for 45 min. The processing reaction was stopped by addition of SDS to a final concentration of l %. The precursors and mature forms were analyzed by SDS-PAGE (Laemmli, 1970) followed by tluorography. The optical densities of the bands on the fiuorogram corresponding to precursors and mature forms were determined by a ·densitometer (Simadzu Chromatoscanner CS-930). The processing activities were estimated from the conversion of the precursors to mature forms. One unit of the enzyme activity was defined as the amount that produced 50% conversion of adrenodoxin precursor to the mature form under the assay condition described above.
Preparation of synthetic peptides
Solid-phase synthesis was carried out manually according to the principle of Dryland and Sheppard et al., (1986). Fmoc-Ala-Wang resin (Wang, 1986) (25 mg, amino acid content 6 �mol) was used for the synthesis. The following Fmoc amino acids were condensed by using of HBTU (3.0 eq) in the presence of HOBt (3.0 eq) and DIEA (6.0 eq); Gly, Ala, Leu, Val, Phe, Pro, Met, Set(tBu) Lys(Boc) and Arg(Mtr). Each condensation reaction was repeated until the resin became negative to the Kaiser test (Kaiser et al., 1970). Deprotection and removal of the peptides from resin were achieved as described by Fujii et al., 1987. The protected (MDH)-Wang resin was treated with TFA ( 1 rnl) in the presence of m-cresol (30 �1), ethandithiol (90 �l) and thioanisol ( 180
�l) at room temperature for one hour. The resin was removed by filtration and washed once with TFA (0.5 ml). TMSBr (200 �l) was added to the filtrate, and the mixture was allowed to stand for one hour on ice. The peptides were precipitated by ice-chilled dry ether (5 ml). After centrifugation, the ether was removed by decantation. After repeated washing with ice-chilled dry ether, the pell�t was dissolved in 10% acetic acid. The
.. - ----����---- - - ---
solution was applied to a column of Sephadex G-10 ( 1.5 x 5.0 em). The fractions which indicate positi\·e to the Kaiser test were combined and lyophilized. The product was purified by reverse phase HPLC on a COSMOSIL AR-5Cl8 colun1n (20 x 250 mm, NACALAI TESQUE, INC Kyoto), and characterized by amino acid analysis with PICO
T AGTM (Waters). Peptide concentrations \\'ere determined by ninhydrin method (Cocking and Yemm, 1955) after hydrolyzation of the peptide in 6N HCI at 105°C for 24 hours.
Acetylation of a-amino group of amino terminal amino acid was performed with acetic anhydride before cleaYage of the peptide from the resin.
Assay of cleavage activity of MPP using synthetic peptides
Synthetic peptides (50 �.M) were incubated with purified MPP in 100 �I of 20 mM Hepes-KOH buffer (pH 7.3) containing 0.1% of Tween 20 at 30°C. The reaction was terminated by addition of 1 mM EDT A after appropriate time. The hydrolyzed products were analyzed by HPLC with a Shimadzu LCA-lOA �pparatus on a COSMOSIL 3C18-P column (4.6 x 100 mm, NACALAI TESQUE, INC Kyoto). Elution was carried out with a linear gradient established between 0.05% trifluoroacetic acid and 0.05% trifluoroacetic acid in 48% acetonitrile for 25 min. The substrate and prcxluct peptides were detected by monitoring at 215 nm. Obtained peak fractions were collected and analyzed for determination of cleavage point by amino acid compositions with PICO-T AG™. To characterize the kinetic profiles of MPP, injtial velocities were examined at various peptide concentrations (2.0, 3.3, 5.0, 10, 16, 25 and 50 !J,M). Michaelis constant (Km) and maximum velocity (Vmax) were determined from double-reciprocal (Lineweaver-Burk) plots of 1/velocity versus 1/[peptide]. In the series of the experiments, from 0.6 to 2.5 units of MPP was used to observe accurate initial velocity of cleavage. Vmax in Tables were corrected for the value when 0.8 units of MPP used.
RESULTS
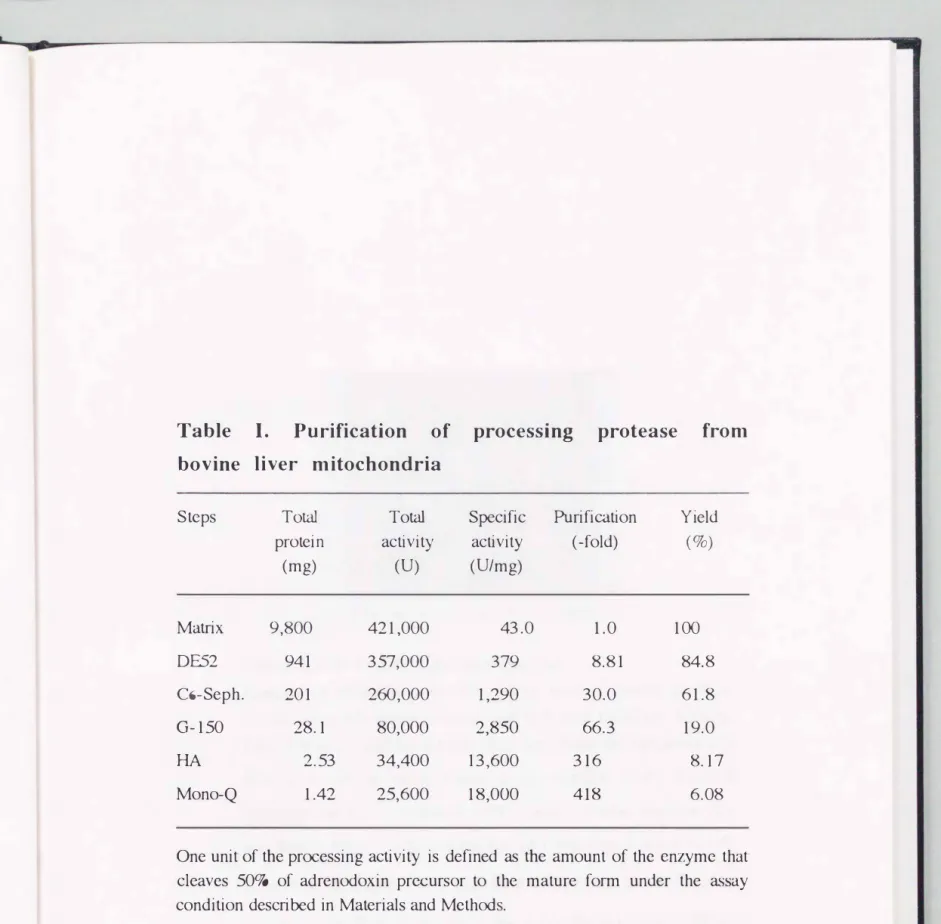
Purification of processing protease
The purification of MPP was carried out by using adrenodoxin precursor as the substrate according to the procedures as described in Materials and Methods. The result is summarized in Table I. I added leupeptin, pepstatin and PMSF to the buffer used in the preparation of the matrix fraction to protect the processing protease from degradation by lysozomal proteases. The processing activity was not inhibited by these inhibitors (Ou et
al., 1989). MPP was purified 418-fold and the recovery was about 6% of total activity in the matrix fraction.
The final preparation of MPP contained two heterogeneous polypeptides of 55 kDa (a) and 52kDa (�),as shown in SDS-PAGE (Figure 1, lane 6). The a and � subunits formed stable complex through out the purification procedures, although two subunits of Neurospora enzyme were dissociated each other during the purification steps
(Hawlitschek et al., 1988).
Cleavage of synthetic peptide
As a substrate for MPP, I synthesized a peptide, MDHl-30 (Figure 2), which contains whole sequence of extension peptide of MDH and additional amino-terminal five residues of the mature portion. A cysteine residue which could be coupled to another peptides or proteins with cross-linking reagents was connected to the carboxyl terminus.
This peptide has two cleavage sites; one for MPP and another for mitochondrial intermediate peptidase (MIP) (Sztul et al., 1988). MDHl-30 was subjected to react with the purified MPP, and the hydrolyzed products were analyzed by HPLC. Two new peaks appeared and increased in time-dependent manner with concomitant decrease of the main peak. The amino acid compositions of the peak fractions showed that the peptide with
Table I. Purification of processing protease from bovine liver mitochondria
Steps Total Total Specific Purification Yield protein activity activity (-fold) (o/o)
(mg) (U) (U/mg)
Matrix 9,800 421,000 43.0 1.0 100
DE52 941 357,000 379 8.81 84.8
C6-Seph. 201 260,000 1,290 30.0 61.8
G-150 28.1 80,000 2,850 66.3 19.0
HA 2.53 34,400 13,600 316 8.17
Mono-Q 1.42 25,600 18,000 418 6.08
One unit of the processing activity is defined as the amount of the enzyme that cleaves 50o/a of adrenodoxin precursor to the mature form under the assay condition described in Materials and Methcxls.
-- ---
69k-
49k-
29k-
Figure 1, SDS-PAGE at purification steps.
Samples of different purification steps were subjected to SDS
PAGE using 8% polyacrylamide gel followed by silver staining.
BSA
(69
kDa), ovalbumin (45 kDa) and carbonic anhydrase (29
kDa) were used as the molecular weight markers (MW). a and
�
represent the two subunits of MPP. Lane 1, matrix fraction
(3.9
�-tg)� lane 2, DEAE-Cellulose fraction (
3
.1 �-tg); lane3,
aminohexyl- Sepharose 4B fraction (2.4 �-tgL lane 4 Sephadex G-150 fraction (1.2 �-tg)� lane 5, hydroxylapatite fraction(690
ngL lane 6, FPLCMono-Q fraction (520 ng).
MPP MIP
1 6�7
24�
�25 _____________________________ �3 38����� Mature Peptide
MIP
MLSALARPVGAALRRSFSTSAQNNAKVAVLGAS--
Synthetic Peptide (MOH1-30)
MLSALARPVGAALRRSFSTSAQNNAKVAVC
Figure 2, Sequences of amino-terminal portion of mouse malate dehydrogenase (MDH) precursor and synthetic peptide.
Presequence of MDH consists of 24 amino acid residues, and are processed in two steps. Initially, MPP cleaves between position 16 and 17, and second, MIP cleaves bet\veen position 24 and 25. The synthetic peptide (MDHI-30) corresponds to the whole presequence and amino-terminal five residues of the mature portion. A cystein residue was attached to the carboxyl-terminus.
retention time of 14.1 min, 16.5 min and 18.3 min were the carboxyl terminal fragment (Phc17- Cys30), the amino terminal fragment (Metl - Ser16), and the substrate (MDHI- 30), respectively (Figure 3). The result indicated that the cleavage occurred at a single site between Ser(l6) and Phe(17) of MDH1-30. The cleavage was not enhanced by addition of metal ion (0.1 and 1.0 mM Mn2+), but was completely inhibited by 1.0 mM EDTA (data not shown).
To detem1ine the kinetic parameters of MPP toward MDH1-30, initial velocitic of the cleavage of the peptide were determined at various concentrations of MDHI-30 (2.0, 3.3, 5.0 and 10 p.M) with 0.8 units of MPP for 0, 15 and 30 min. Km and Vmax values were calculated to be 0.73 f..tM and 7.8 pmollmin, respectively (Table II). These parameters obtained by repeated experiments varied at most 30% and could be reproducible.
Deletion of amino acid residues in the MDH 1-30
To investigate which portions in the MDH1-30 are important for the cleavage by MPP, I synthesized peptides with various length of the peptides which were deleted amino acid residues from both the amino- and carboxyl-terminus (Table II). MDH1-24 and MDH1-21 were deleted of 6 and 9 residues, respectively, from the carboxyl terminus of MDHI-30. MDH6-21, MDH9-21 and MDH12-21 were deleted of 5, 8 and 11 re idues, respectively, from the amino terminus of MDH1-21. I determined kinetic parameters for these peptides. As shown in Table I, the peptides deleted from carboxyl terminus (MDHl- 24, MDH1-21) were cleaved at the same efficiency as MDH1-30. The presence of five amino acid residues in the downstream of the cleavage site was sufficient for the cleavage by the peptidase. Km for MDH6-21 was increased by only about 2-fold over that for MDH l-21, while those for MDH9-21 and MDH12-21 were increasedby about 24-fold and 50-fold, respectively, without any concomitant changes in their maximum velocities.
1 16 17 30
MLSALARPVGAALRRSFSTSAQNNAKVAVC
I ® II CD I
I 0 I
®
...__ _ 12 _..__, __ __,__ __ 14 �•--�--16 ---'---------•-18 -___.v'
Retention time (min)
N 0
ci
II
Figure 3, HPLC profile for cleavage of MDHl-30 by MPP
MDHI-30 was incubated with purified MPP at 30°C. At each time point, 100 �I aliquot was withdrawn. EDTA added and the products were analyzed by HPLC. The circled numbers indicate the corresponding fragments.
Table II. Deletion of amino acid residues in the MDH 1-30
MDH1-30 MLSALARPVGAALRRS-FSTSAQNNAKVAVC MDHl-24 MLSALARPVGAALRRS-FSTSAQNN
MDH1-21 MLSALARPVGAALRRS-FSTSA MDH6-21 ARPVGAALRRS-FSTSA MDH9-21 VGAALRRS-FSTSA
MDH12-21 ALRRS-FSTSA
Km Vmax [mM] [pmol/minl
0.73 7.8 0.78 6.3 0.71 8.4
1.2 10
17 9.4
35 9.3
V max Km
11 8.1 12
8.3 0.55 0.27
The kinetic parameters of the processing peptidase toward these substrates were mesured as described under "Materials and Methods". The hyphen indicats the cleavage point.
These results suggested that the peptide with 11 and 5 amino acid residues from the cleavage site to the amino- and carboxyl-terminal sides, respectively, was sufficient for the cleavage of the peptide substrate. MDHl-21 was, then, used as the standard substrate in the later experiments.
Substitution of arginine residues by alanine residues
Since earlier studies indicated that some basic amino acid residues in the presequence arc essential for the import of the precursors to mitochondria and the cleavage of the precursors by MPP (Kumamoto et al., 1987 and 1989; Chu et al., 1987), I studied the effect of four positively charged groups, a-amino group and three arginine residues, in MDH1-21. I replaced one or two of three arginine residues of the MDH peptide by alanine residue, and a-amino group of methionine was blocked by acetylation.
Table III shows that substitution of Arg(7) by Ala (MDH7A) produced 2-fold increases in Km, 3-fold reduction in Vmax and then 6-fold reduction in peptide catalytic efficiency (Vmax!Km). Amino terminal acetylation of MDH7A gave 4-fold increases in Km. This change of the kinetic parameters were remarkable when compared to those of MDH1-21, indicating that the positively charged group in the position distal to the cleavage site are necessary for effective cleavage of the peptide.
Substitution of Arg(l4) by Ala (MDH14A) did not gave a significant change in any kinetic parameter. In contrast, substitution of the Arg( 15) by Ala (MDH 15A) gave 4-f old reduction in Vmax. This suggests that MDH15A could be recognized but was slowly cleaved by MPP. Substitution of both arginine residues by Ala (MDH14, 15A) gave 50- fold increase in Km, two-fold reduction in Vmax and therefore 100-fold reduction in Vmax!Km. The cleavage was occurred at the correct site, Ser-Phe (16-17) in these peptides. These results suggested that out of two proximal arginine residues, only Arg(l 5) are sufficient for the efficient cleavage by MPP. Since the cleavage occurcd at the same site
Table III. Substitution of argtntne residues by alanine residues
Km Vmax Vmax [mM] lpmol/min] Km
MDH1-21 MLSALASPVGAALBBS-FSTSA 0.7 1 8.4 12 MDH7A MLSALAAPVGAALRRS-FSTSA 1.4 2.8 2.0 MDH7A-Ac Ac�LSALAAPVGAALR RS-FSTSA 6.2 4.0 0.64 MDH14A MLSALARPVGAALARS-FSTSA 1.3 12 9.2 MDH15A MLSALASPVGAALRAS-FSTSA 1.3 2.3 1 .8 MDH14,15A MLSALARPVGAALAAS-FSTSA 34 3.8 0.1 1
The kinetic parameters of the processing peptidase toward these substrates were mcsurcd as described under "Materials and Methods". Basic amino acids are indicated by underline. The hyphen indicats the cleavage point.
in spite of the substitution of the pro\..imal arginine residues, other amino acid residuec except for the proximal arginine residues would be also recognized by MPP.
Substitution of arginine residues by lysine residues
To examine if arginine residues in the presequence were replaceable by lysine residues, I synthesized three peptide which contained lysine residue in the place of arginine residue (Table IV). Substitution of the distal Arg(7) by Lys (MDHKR) did not ga\'e a significant change in any kinetic parameters. In contrast, substitution of the proximal Arg(l5) by Lys (MDHRK) caused a dramatic decrease in affinity between the peptide and MPP, and in cleavage efficiency. MDHKK which was substituted for both distal and proximal arginine residues, was cleaved only with a low efficiency same as MDHRK. This results indicated that the distal arginine residue was recognized simply as a basic amino acid, however, the proximal arginine residue should be itself.
Position of arginine residues
I next examined the effect of the positions of two arginine residues, at the proximal and distal to the cleavage site, on the cleavage by MPP. I synthesized peptides of which distal and proximal arginine residues were shifted to upstream or downstream in the presequence (Table V). Shifting the position of the proximal arginine to the just before the cleavage site (MDH16R) produced 2-fold increases in Km and 8-fold reduction in Vmax compared with MDH14A. Shifting the position to amino-terminus for one residue (MDH16R), which was showed in Table III, produced 5-fold reduction in Vmax though
with the same Km value. Further shift to amino-terminus for two residues (MDH13R) produced 7-fold increases and 5-fold reduction in Vmax. It is concluded from these results that position of the proximal arginine residue should be at -2 position and that shift of this residue to -3 or -1 position was resulted in a remarkably decrease in the cleavage efficiency. On the other hand, shifting the position of the distal Arg(7) by two residues
-- OJ p
Table IV. Substitution of arginine residues
MDH14A MDHKR MDHRK MDHKK
MLSAL�PVGAAL�S-FSTSA MLSALA!PVGAALARS-FSTSA MLSALARPVGAALAKS-FSTSA - - MLSAL�PVGAALA!S-FSTSA
by lysine residues
Km Vma.'\ Vma\
[mM] [pmol/min] Km
1.3 12 9.2
2.2 17 7.7
9.1 4.2 0.46
10 2.3 0.23
The kinetic parameters of the processing peptidase toward these substrates were mesurcd as described under "Materials and Methods". Basic amino acids are indicated by underline. The hyphen indicats the cleavage point.
Table V. Position of argrn1ne residues
MDH16R MDH14A MDH15A MDH13R
MDH9R MDH14A MDH5R
MLSALAgPVGAALAA�-FSTSA MLSALAgPVGAALA�S-FSTSA MLSAL�PVGAAL�AS-FSTSA MLSALAgPVGAAgAAS-FSTSA
MLSALAAP�GAALA�S-FSTSA MLSALAgPVGAALA�S-FSTSA MLS�AAPVGAALARS-FSTSA
Km
lmM]
3.0 1.3
1.3 9.2
2.6 1.3 3.8
Vma.�
lpmol/mi n]
1.5 12
2.3 2.6
11 12 16
Vrnax
Km
0.50 9.2
1.8 0.28
4.2 9.2 4.2
The kinetic parameters of the processing peptidase toward these substrates were mesured as
described under "Materials and Methods". Ba<;ic amino acids are indicated by underline. The hyphen indicats the cleavage point.
- --- -
toYvard carboxyl- or amino-terminus (MDH9R and MDH5R) ga\'e only 2-fold increases in Km and little change in Vmax. Thus, the position of the basic amino acid rei due(s) m
the distal portion of to the cleavage site is not so critical for recognized by MPP.
Distance between distal and proximal arginine
Our previous data showed that the basic amino acid residues at 20 to 35 residues upstream of the cleavage site were necessary for the recognition of the adrenodoxin precursor (pAd) by MPP. As shown above, the position of the distal arginine residue of the pMDH is only 7 amino acid residues apart from the cleavage site. The portion between the distal and proximal arginine residues is expected to have some function that allow the arginine residues at different position to be recognized by MPP. In this portion of pAd, I noticed the presence of an unique glycine rich region. I examined that MPP can recognize a chimeric presequence (MDHAdl4) in which the glycine rich region, from Gly(35) to Thr(48), of Ad presequence was inserted into the portion between distal and proximal arginine residues of MDH presequence (Figure 4). The amino-terminal a-amino group of MDHAd14 was acetylated in order to eliminate positively charged group except for the distal and proximal arginine residues. This exchange gave a little reduction in Km, and only 2-fold reduction in Vmax (Table VI). To examine which of two distal arg1mne residues of MDHAd14 was recognized by MPP, I substituted one of these arg1mne residues by alanine residue (MDHAdAR and MDHAdRA). When both of the distal arg1mne residues (MDHAdAA) were replaced to alanine, the Km for the peptide remarkably increased, as observed in the experiments used the original MDH peptidcs (Table III). These results indicated that the distal arginine residues in the MDHAd series peptides was recognized by MPP, and the portion between distal and proximal arginine residues had an important function for recognition of distal arginine residue by MPP.
pAd:
1 5 10 15 20 25MAARllRVASAAlGDTAGRWRllVR-
30 35 40 45 50 55
PRAGAGGLR(tt§itij;tij!§§if4'fJiJTLSVSGRAQ-SSS---
MDH14A: "' Cleava !e site
1 5 10 15
·;
20MlSALA.B}PVGAALAflS-FSTSA
MDHAd14:
1 5 1o 15 2o 25Ac-MlSALAR(d#)itij:Zij!ijij§t;)!IJJ:Js-FsTsA
Figure 4, Schematic structure of MDHAd14.
Presequence of pAd has a long glycine rich region indicated by closed box. By this region, the portion present between distal and proximal arginine residues of MDH14A indicated by opened box was replaced.
The amino-tenninal a-amino group of the resulting peptide was acetylated (MDHAd14).
Table VI. Exchange of the portion present between distal and proximal arginine in MDH presequence for glycine rich region of Ad presequence
MDH14A MLSAL�PVGAAL�S-FSTSA
MDH-Adl4 Ac�LSAL�GSRGPGLGGGAVATRS-FSTSA MDH-AdAR Ac�LSALAAGSRGPGLGGGAVATRS-FSTSA MDH-AdRA Ac�LSAL�GSAGPGLGGGAVAT�S-FSTSA MDH-AdAA Ac�LSALAAGSAGPGLGGGAVATRS-FSTSA
Km Vmax [mM] [pmollmin]
1.3 12
0.98 5.7
1.7 5.8
2.0 5.4
34 13
Vmax Km
9.2 5.8 3.4 2.7 0.38
The kinetic parameters of the processing peptidase toward these substrates were mesured as described under "Materials and Methods". Basic amino acids are indicated by underline. The hyphen indicats the cleavage point.
Importance of proline and glycine residues
The presequence of MDH haYe a glycine and proline residues, \\'hich \\'Ould break secondary structure of peptide chain. To examine if these amino acid residues arc important for the cleavage by MPP, I replaced these residues by alanine residues
(MDHa). As the result of the replacement, the cleavage efficiency was remarkably reduced (Table VII). This result suggested that the flexibility of the portion \\'ould be important for the recognition by MPP.
-- ::g:t p
Table VII. Substitution of proline and glycine residues by alanine residues
MDHI-21 ML SALARPVGAALRRS-FSTSA MDHa ML SALARAVAAALRRS-FSTSA
Km
[roM]
0.71 6.0
Vmax [pmol/min]
8.4 5.0
Vmax Km
12 0.83
The kinetic parameters of the processing peptidase toward these substrates were mesured as described under "Materials and Methods". Substituted alanine residues are indicated by underline. The hyphen indicats the cleavage point.
DISCUSSION
In eukaryotic cells, mitochondria contain se,·eral components required for mitochondrial protein translocation. Out of them, MPP is the first c\.ample of clearly identified component involved in the process. MPP from Neurospora crassa contained two protein components, a and B subunits. Although both of them were required for the processing activity, they did not form a complex. I have shown that the processing peptidase purified from bovine li\'cr mitochondria was also composed of two a and f1 subunits, which '''ere very close in molecular weight to those from Neurospora crassa.
Unlike the fungi enzyme, however, a and B subunits obtained from bovine were not separated from each other at any chromatography same as MPP from rat liver mitochondria(Ouetal., 1989). Yeast a and B subunits were dissociated in gel filtration, but remained associated in sucrose gradient centrifugation (Yang et al., 1988). It seems indicating that there is a week interaction between them. The interaction of the two subunits was different between the mammalian and lower eukaryotes' enzymes.
MPP specifically recognizes most of presequences of mitochondrial precursor proteins. I intended to clear specific structure common to all presequences. The study of structural feature of the presequence on MPP is one approach to reveal recognition mechanism of MPP, and also give a clue to that of import machinery on mitochondrion.
To clear the structural feature, it is necessary to analyze the processing reaction quantitatively, and to investigate what portion and/or amino acids in the presequence is responsible for recognition by MPP. For this purpose, I used synthetic peptides, instead of whole precursor molecules translated in vitro, as the substrate. First of all, I synthesized a peptide, MDH 1-30, which corresponds to entire portion of presequence of mouse malate dehydrogenase together with the amino-terminal portion of the mature protein. It was cleaved at a single site by the purified MPP. The amount of resulting
fragments \�as quantitatiYely analyzed by HPLC. Kinetic data obtained from this assay methcxi seem comparable to those from the other method using precur or proteins expressed in E. coli: the Km for MDHl-30 (0.73 �tM) was close to that for a pCOXIV
DHFR fusion protein (0.17 f.!M) (Yang et al., 1991). The result suggest.� that the peptide was cleaved b) MPP in the same manner as the precursor protein. Furthermore, the fact that the peptide could be cleaved indicates that mature portion of the prccur. or was not necessary for the recognition by MPP. For the effective cleavage of substrate by MPP, the peptide with 11 and 5 amino acid residues from the cleavage site to the amino- and carboxyl-terminal sides, respectively, was sufficient.
Two sets of basic amino acids, which are important for affinity with MPP, \\'ere found: one is a pair of the proximal Arg(l4 and 15) and the other is the distal Arg(7) of the presequence. Out of two proximal arginines, Arg( 15) are more important than Arg ( 14), because replacement of Arg(15) by Ala lead Vmax of the activity cleaving the peptide to 114 while that of Arg(l4) by Ala rather increased the Vmax value. Replacement of
Arg(l5) by Lys, as the positive-charged amino acid, the efficiency of cleavage of the peptide was reduced remarkably. The proximal basic amino acid should be arginine residue. The position was strictly determined at -2 position relative to cleavage site. MPP would have a substrate binding pocket at S2 site of which structure fit for side chain of arginine residue. A specific structure which facilitates cleavage between Ser(l6) and Phe( 17) could be formed in the substrate-enzyme complex in combination with Arg( 15), Phe( 17) and subsites of the enzyme.
Replacement of the distal Arg(7) by Ala decreased Vmax!Km to 116. Thi. result indicated that Arg(7) is also important for the cleavage by MPP. This is also sup}X)rted by accordance with the result that the deletion of eight amino acid residues from amino
terminal of the presequence gave drastic reduction in cleavage efficiency. Since the distal Arg(7) was replaceable with Lys, MPP would recognize distal basic amino acids in the substrate. Acetylation of the a-amino group of Met( 1) of MDH7 A also reduced
--- :at .
significantly cleaYage efficiency. This suggest that a-amino group could ubstitutc for the distal basic amino group. I also confirmed that the pcptides of which the distal arginine residue could be shifted for ±2 residues could be cleaved by MPP. These observations indicate that the position of the distal basic amino group is not � trictly determined for the recognition of the prcsequence by MPP.
Previous studies showed that little homology of the position of the distal arginine residues among the various presequences exists (von Heijne, 1986; von Hcijne ct al., 1989; Roise and Schatz, 1988� Pfanner and Neupert, 1991). As described above, the distal arginine of MDH presequence is 9 residues upstream of the cleavage site. Whereas, in the case of adrenodoxin precursor (pAd), arginine residues at 20 to 35 residues upstream of cleavage site were reported to be important for the cleavage by MPP (Ou et al., 1994 ). Thus, the distance between these distal arginine residues of pAd and cleavage site was much longer than that of pMDH. However, both of the precursors were correctly recognized and processed by MPP. How can MPP correctly recognize the distal arginine residues that exist at various positions in presequences? The presequences of many precursor proteins, including pMDH and pAd, have several glycine and proline residues between the distal and proximal basic amino acid residues. The fact suggests that the distal basic amino acid residues could be functionally and structuraly connected to the proximal arginine residue and cleavage site via a flexible linker containing proline and glycine residues. In the case of pMDH, the flexible region may be from Pro(8) to Gly( 10) bending the presequence either by Pro(8) or Gly(lO) was also important. Really, substitution of both the Pro(8) and Gly(l O) of MDH1-21 by Ala largely reduced cleavage efficiency.
In the presequence of pAd, a long glycine rich region between the distal and proximal arginine residues could serve as the flexible linker and bend the presequence allowing the distal arginine residues to approach to the binding site on MPP. The portion between the distal and proximal arginine residues of MDH1-21 could be replaceable by the longer
glycine rich region. Really, MDHAdl4 that replaced by the glycine rich regton 1n presequence of pAd was effectively cleaved. Out of two distal arginine residues of MD HAd 14, either arginine residue could be sufficient to be cleaved by MPP. This re. ult suggests that the distal arginine residues in different position could be recognized by MPP and also supports the 1lexibility of the glycine rich region.
Karslake et al. (1990) found the existence of structurele s Arg-Gly-Pro in the presequence of rat liver aldehyde dehydrogenase, and have confirmed its function a a flexible linker connecting two a-helices in the presequcnce by NMR studic . Subsequently, Thornton et al. (1993) have recently shown that deletion of the linker abolished cleavage of the presequence without altering the import ability of the precur or.
Our studies and the study for aldehyde dehydrogenase prompt me to propo e a ba<;ic amino acid-flexible linker-arginine model for pMDH processing by MPP (Figure 5). In this model, the presequences are largely random coil in solution (from circular dichroism study of MDHl-21, this has been confirmed, data not shown) (Figure SA). The distal Arg(7) and proximal Arg(l5) are recognized by MPP, while the flexible region contributes to give an appropriate distance between the two arginine residues to fit substrate recognition site on MPP. Accompanying the recognition of the arginine residues, cleavage site would accommodated to the active site on MPP, and the substrate was consequently cleaved (Figure 5B). In the case of pAd, the long flexible region bends the prescquence allowing the distal arginine residues to approach to the binding site on MPP.
In the k:inetical studies for synthetic peptides and its derivatives a') a substrate, I could indicate the primary structural feature of the presequences of precursor proteins that bind on MPP, and a clue to the mechanism that recognizes various presequences without sequence homology. However, the definite structure could not be clear. To clear this, NMR study, especially transferred NOEs, would become powerful. To analyze the structure, the peptide that binds but not be cleaved is now under investigation. On the other side, study on structure of MPP is also necessary for clarification of substrate
A
Jt
B Flexible region
I 1
a
�-- pAd ,_.�������� pM DH
Figure 5, Model for binding of precursor proteins to MPP.
A, ramdom coil state. B, specific binding of the precursor
proteins to MPP. The flexible region contributes to give a
appropriate distance between the arginine residues to fit substrate
recognition site on MPP.
recognition mechanism. cDNAs of both a and� subunits of rat MPP have recently cloned (Kleiber et al., 1990� Kitada et al., 1993� Paces et al., 1993). A series of such approaches will provide us the insight into how MPP recognizes and cleaves hundreds of different precursor proteins.
REFERENCES
Bohni, P., Daum, G., and Schatz, G. (1983) J. Bioi. Chenl. 258, 4937-4943
Chu, T. W., Grant, P.M., and Strauss, A. W. (1987) J. Bioi. Chern. 262, 12806- 12811
Cocking, E. C., and Y emm, F. W. ( 1955) Analyst 8 0, 209
Conboy, J. G., Fenton, W. A., and Rosenberg, L. E. (1982) Biochetn. Biophys. Res.
Commun. 105, 1-7
Dryland, A., and Sheppard, R. C. ( 1986) J. Chern. Soc., Perkin Trans. 1, 125 Fujii, N., Otaka, A., Sugiyama, N., Hatano, M., Yajima, H. (1987) Chern. Phann.
Bull. (Tokyo) 3 5, 3880-3883
Furuya, S., Okada, M., Ito, A., Aoyagi, H., Kanmera, T., Kato, T., Sagara, Y., Horiuch, T., and Omura, T. (1987) J. Biochem. 102, 821-832
Hawlitschek, G., Schneider, H., Schmidt, B., Tropschug, M., Hartl, F. -U., and Neupert, W. (1988) Cell 53,795-806
Isaya, G., Kalousek, F., Fenton, W. A., and Rosenberg, L. E. (1991) J. Cell Bioi.
113,65-76
Jensen, R. E., and Yaffe, M.P. (1988) EMBO J. 7, 3863-3871
Joh, T., Takeshima, H., Tsuzuki, T., Shimada, K., Tanase, S., and Morino, Y. (1987) Biochemistry 2 6, 2515-2520
Kaiser, E., Colescott, R. L., Bossinger, C. D., and Cook, P. I. (1970) Anal. Biochem.
34, 595
Kalousek, F., Hendrick, J.P., and Rosenberg, L. E. (1988) Proc. Natl. Acad. Sci. U.
S. A. 85, 7536-7540
Karslake, C., Piotto, M. E., Pak, Y. K., Weiner, H., and Gorenstein, D. G. (1990) Biochemistry 2 9, 9872-9878
Kumamoto, T., Morohashi, K., Ito, A., and Omura, T. (1987) J. Biochenz. l 02. 833- 838
Kumamoto, T., Ito, A., and Omura, T. ( 1989) J. Biochenz. 105, 7'2-78
Kitada, S., Niidome, T., Nagano, T., Ogishima, T, and Ito, A. (1993) Biochem.
Biophys. Res. Conzmun. 190, 289-293
Kleiber, J., Kalousek, F., Swaroop, Manju., and Rosenberg, L. E. (1990) Proc. Natl.
A cad. Sci. U. S. A. 8 7, 7978-7982 Laemmli, U.K. (1970) Nature 227, 680-685
McAda, P. S., and Douglas, M.G. (1982) J. Biol. Chem. 257, 3177-3182
Miura, S., Mori, M., Amaya, Y., and Tatibana, M. (1982) Eur. J. Biochem. 122, 641- 647
Ou, W. -J., Ito, A., Morohashi, K., Fujii-Kuriyama, Y., and Omura, T. (1986) J.
Biochem. 100, 1287-1296
Ou, W. -J., Ito, A., Okazaki, H., and Omura, T. (1989) EMBO J. 8 , 2605-2612
Ou, W. -J., Kumamoto, T., Mihara, K., Kitada, S., Niidome, T., Okazaki, H., Ito, A., and Omura, T. unpublished data
Paces, V., Rosenberg, L. E., Fenton, W. A., and Kalousek, F. (1993) Proc. Natl.
Acad. Sci. U. S. A. 9 0, 5355-5358
Pelham, H. R B., and Jackson, R. J. (1976) Eur. J. Biochem. 67, 247-2.56 Pfanner, N., and Neupert, W. (1990) Annu. Rev. Biochem. 59, 331-353
Pollock, R. A., Hartl, F. -U., Cheng, M. Y ., Ostermann, Joachim., Horwich, A., and Neupert, W. ( 1988) EMBO J. 7, 3493-3500
Roise, D., and Schatz, G. (1988) J. Bioi. Chern. 263, 4509-4511
Rosenberg, L. E., Fenton, W. A., Horwich, A. L., Kalousek, F., and Kraus, J. P.
(1987) Ann. NY Acad. Sci. 488, 99-108
Sagara, Y., Ito, A., and Omura, T. (1984) J. Biochem. 9 6, 1743-1752
Schneider, H. S., Arretz, M., Wachter, Elmar., and Neupert, W. (1990) 1. Bioi. Chem.
265, 9881-9887
Sztul, E. S., Chu, T. W., Struss, A. W., and Rosenberg, L. E. (1988) 1. Bioi. Chem.
262, 12085-12091
Thornton, K., Wang, Y., Weiners, H., and Gorenstcin, D. G. (1993) 1. Bioi. Chem.
268, 19906-19914
Verner, K., and Schatz, G. (1988) Science 241, 1307-1313 von Heijne, G. (1986) EMBO 1. 5, 1335-1342
von Heijnc, G., Steppuhn, J., and Hermann, R. G. (1989) Eur. 1. Biochenl.180. 535- 545
Wang, S.-S. (1973) 1. Am. Chem. Soc. 95, 1328
Witte, C., Jensen, R. E., Yaffe, M.P., and Schatz, G. (1988) EMBO J. 7, 1439-1447 Yang, M., Geli, V., Oppliger, W., Suda, K., James, P., and Schatz, G. (1991) 1. Bioi.
Chern. 266, 6416-6423
Yang, M., Jensen, R. E., Yaffe, M.P., Oppliger, W., and Schatz, G. (1988) EMBO J.
7' 3857-3862
ACKNOWLEDGEMENTS
I am deeply grateful to Professor Akio Ito and Dr. Tadashi Ogishima for their supports and discussions throughout this study. I am also thanksful to Professor Sadaaki Iwanaga and Dr. Shun-ichro Kawabata, Department of Biology, for analyzing amino acids composition. In addition, I would also express my heartful thanks to all the members in this laboratory for their kindly helps during the course of the study.








![Table VII. Substitution of proline and glycine residues by alanine residues MDHI-21 ML SALARPVGAALRRS-FSTSA MDHa ML SALARAVAAALRRS-FSTSA Km [roM] 0.71 6.0 Vmax [pmol/min] 8.4 5.0 Vmax Km 12 0.83](https://thumb-ap.123doks.com/thumbv2/123deta/9809741.1885660/30.996.18.941.40.1343/substitution-proline-glycine-residues-alanine-residues-salarpvgaalrrs-salaravaaalrrs.webp)
