低温生物工学会誌〔Cryobiology and Cryotechnology〕, Vol. 63, No. 1, 1~10, 2017
トレハロースや
LEA タンパク質により誘導される生体の乾燥耐性
メカニズムに関する実験的・理論的研究
東京工業大学バイオ研究基盤支援総合センター
櫻井
実
Experimental and Theoretical Study on the Mechanism of Desiccation Tolerance of
Anhydrobiotic Organisms Induced by Trehalose and LEA Proteins
Minoru SAKURAI
Center for Biological Resources and Informatics, Tokyo Institute of Technology, B-62 Nagatsuta-cho, Midori-ku, Yokohama 226-8501, Japan
(Corresponding author, e-mail: [email protected])
In the anhydrobiosis state of desiccation-tolerant organisms such as sleeping chironomid (Polypedilum
vanderplanki), metabolic reactions are completely suspended but the increase of their entropy is
suppressed well unlike in the death (substrate) state. Probably there exists “entropy barrier” to be able to
suppress entropy increase in the body of anhydrobiotic organisms. The entropy barrier is thought to be
constructed from desiccation protectants such as trehalose and late embryogenesis abundant (LEA)
proteins. In the past three decades, we have investigated the structure and function of these protectants
from both experimental and theoretical viewpoints. Here, we summarize the main results of those studies.
In the first part, we summarize the solution and solid-state properties of trehalose and the underlying
mechanism of torehalose-induced desiccation tolerance in P. vanderplanki. In the second part, we
summarize the results for the structure and function of LEA peptides, which are model compounds
whose amino acid sequences consist of two or four tandem repeats of the 11-mer motif found commonly
in native group 3 LEA proteins.
(Received Dec. 8, 2016; Accepted Dec. 13, 2016)
1.緒 言
ネムリユスリカ幼虫などの乾燥耐性生物は,体内 第6 回低温生物工学会受賞講演
[Key words: Trehalose, LEA protein, Desiccation tolerance, Vitrification, Water replacement, Molecular shielding; トレハロース,LEA タンパク質,乾燥耐 性,ガラス化,水置換,分子シールディング] からほぼ完全に水が失われると代謝反応を停止し, 一見物質のような状態(塊)になる.しかし,給水 すると自発的に吸水し蘇生する.この物質のような 状態はアンヒドロビオシス(anhydrobiosis)状態と呼 ばれ,生物として活動している状態や死状態(本当 の物質状態)と区別される.それでは,これら3状 態を区別する要因は何であろうか. 量子力学の開拓者の一人であるシュレーディンガ -1- -1-
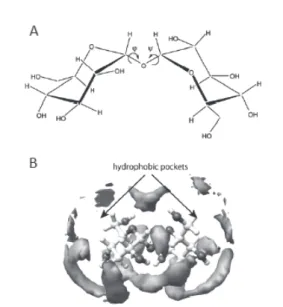
ーは,その著書「生命とは何か」の中で,生命とは“負 のエントロピーを摂取して秩序を維持している系” と述べている1).それならば対照的に,物質とは“崩 壊して平衡状態に至る系”と言えよう.こう考えると, アンヒドロビオシス状態の特徴も見えてくる.アン ヒドロビオシス状態では,代謝は停止しているので, 負のエントロピーを取り込むことはできない.しか し,“その内部にエントロピーの増大に抵抗する障壁 を擁して秩序を維持している(崩壊を著しく遅延さ せている)系”と言うことは可能であろう.こう考え ると,物質との違いも明確になり,第三の状態と位 置づけるのも無理はなくなる. それでは,乾燥耐性生物の中にあるエントロピー 障壁とは具体的にどのようなものであろうか.それ を解く鍵は,乾燥時に多量に産生される物質が握っ ているはずである.その一つは糖類であるが,中で もトレハロースは様々な生物界(植物,動物,キノ コ類,線虫,酵母,菌類,ブラインシュリンプ,昆 虫 な ど ) で 見 出 さ れ て い る 2). も う 一 つ は late embryogenesis abundant (LEA)タンパク質である. LEA タンパク質は,最初は植物の種子で見出された が,最近では線虫や昆虫などの動物でも見出されて いる3). では,なぜ自然はトレハロースやLEA タンパク質 を選択したのだろうか.また,前述したエントロピ ー障壁とは具体的にはどのようなものだろうか.物 理化学の教えるところによれば,すべての分子の性 質はその構造に起源がある.われわれは,トレハロ ースや LEA タンパク質の性質とその機能をその構 造特性から実験的・理論的に解明する研究を続けて きた. 2.トレハロースの構造・性質・機能 2-1.トレハロースの構造的特性 トレハロースは二つのグルコースがα,α-1,1 結合 した二糖である.何の変哲もない糖のように見える が,このグリコシド結合に仕掛けがある.二面角φ とψ は伴に-60°付近の値しかとり得ない.すなわち, 安定コンフォメーションが一つしかないのである. その結果,Fig. 1A に示すように,二枚貝が開いたよ うな外形をしている.さらに開いた側(bottom)にある OH 基はすべてエカトリアル配置をしていることも 特徴である. 2-2.トレハロースの水和特性 前述した構造特性によりトレハロースはユニー クな物性を示すはずである.われわれは,まず水和 特性に着目し,示差走査熱測定(DSC)から得られる 不凍水の量を比較した4).その結果,トレハロース の水和数はマルトースやスクロースの場合より多 い こ と が 判 明 し た . こ れ ら 二 糖 は 同 じ 分 子 式 (C12H22O11)で同じ分子量(342.3)をもつので,確 かに立体構造の違いにより異なる水和特性が生じ ることを示している.さらに,水和殻中の水分子の ダイナミクスの情報を 17O-NMR による水のスピン 格子緩和時間T1の測定により調べた4).その結果得 られた動的水和数nDHNは,やはりトレハロース>マ ルトース>スクロースの序列となった.さらに,回 転相関時間の評価から,トレハロース水和殻中の水 分子の運動は,他の二糖に比べて大きく拘束されて いることが判明した. われわれがこのような研究発表をしたのとほぼ 同時期に,世界的にもトレハロースの生理機能をそ の水和特性に関連付けて説明しようとする研究が 活発になり,水溶液の固有粘度,部分モル体積,等 エントロピー,部分モル圧縮率,部分モル熱容量な どのデータが発表された.これらのデータについて はいくつかの総説 5,6)の中で論じてきたのでここで は省略する.それら様々なデータを総合すると,ト
Fig. 1. A: Chemical structure of trehalose,
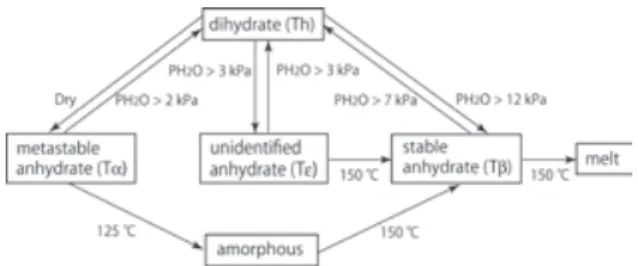
Fig. 2. Phase and state transitions of trehaslose in
the solid state. レハロースは周囲の水素結合ネットワークを破壊 し,それ自身の周りに秩序化された水和圏を形成す る能力(kosmotrope としての機能)の高い分子であ るという描像が得られた. トレハロースの機能を考えるときは,このような 溶液物性が共存するタンパク質,細胞膜,その他生 体材料の性質に本当に摂動を与えるのか否かが問 題である.そこで,われわれは糖の水和特性の違い を相互作用相手のマクロの性質に射影できる系と して,アガロースゲルの膨潤をとりあげた.その結 果,トレハロース添加系の膨潤度はスクロース添加 系の場合に比べて小さいことが判明した7).これは 次のように解釈される.トレハロース添加の場合は この糖の強い水和力により,膨潤の主要因であるゲ ル高分子鎖-水間の親和性が低下し,その結果,膨 潤の抑制に寄与するゴム弾性の効果が優位になり 膨潤度が低下した,と考えられる. 一般に実験データから得られる水のダイナミクス と水和構造の情報は,溶質の様々なコンフォメーシ ョンに関して平均化された分子全体に対する値であ る.したがって,実験のみでは溶質表面の各部位ご との水和特性の違いまでは識別することはできず, 糖の立体化学によって決まる水和の違いについて詳 細な情報を得ることは困難である.対照的に,コン ピュータシミュレーションはこのような問題の解決 に有効である.われわれは,いち早くMD シミュレ ーションを導入し,最初の研究ではトレハロースと マルトースの水和特性の比較に焦点を当てた 8).そ の結果,トレハロースではすべての水酸基が均等に 水と水素結合を形成し(マルトースでは欠損が見ら れた),水和殻の水分子の並進拡散がより効率的に抑 制されることが示され,前述した実験値の傾向を裏 付けるデータが得られた. 近年の目覚しいコンピュータ性能の向上によって, オリゴマー程度の糖水溶液に対するMD シミュレー ションならば迅速に且つ正確に行えるようになって きた.その結果,α,α-1,1 結合の剛直性から生じるト レハロースの水和特性が明らかとなった5).Fig. 1B に示したように,bottom 側では水の酸素原子は帯状 に連続した領域に規則的に分布している.これは水 分子が秩序だって配列していることを示している. このような規則的水和構造は,bottom 側に並ぶエカ トリアルOH 基の配置と水の水素結合ネットワーク 構造が上手く一致するために生じ,且つ立体構造が 剛直であるため安定に存在する.この結果は,トレ ハロースがよいkosmotrope であるという実験結果を 支持するものである.一方,二枚貝の蝶番のある側 (top)では,二つの疎水ポケット(第一水和殻が存 在しない領域)が存在する.これはグルコース環の 上下面は疎水性のためである. 以上述べた結果を総合すれば,トレハロースは分 子内に親水面と疎水面をもち異方的な水和殻を形 成していると言える.そのため,時には疎水性分子 として振舞うことも可能であり,弱いながらもベン ゼンやその誘導体と錯体を形成する9). 2-3.トレハロースの固体状態の性質 2-3-1 トレハロースの相・状態転移 トレハロースは通常二水和物(Th 相)として結晶 化する.日本分光(株)の赤尾賢一氏らとわれわれ は結晶水の HOH 変角振動の FTIR 測定から,この 結晶の融点は 90℃付近にあるが,結晶水は 70℃付 近でice-like 状態から liquid-like 状態へ転移すること を見出した 10).つまり結晶の融点より 20℃も低い 温度で結晶水の融解が先に起こる.そのため,昇温 条件によっては,二水和物の融解をジャンプして, 無水物結晶が直接生成されることがある.実際,ト レハロース二水和物結晶を温和な条件下で加熱乾 燥すると,130℃に融点を持つ準安定な結晶相(Tα 相)が得られる.われわれはこの準安定相がトレハ ロースの乾燥保護機能の鍵を握ると考え,その生成 条件や性質を熱測定 11,12),DSC-X 線同時測定13), FTIR 測定14,15)およびX 線結晶構造解析16)等を用い て詳細に調べた.その結果得られたトレハロースの 固相における状態・相転移をFig. 2 に示す.ここで, Tβ とラベルされた相は,通常の脱水過程で生成す る安定な無水物結晶である. -3- ーは,その著書「生命とは何か」の中で,生命とは“負 のエントロピーを摂取して秩序を維持している系” と述べている1).それならば対照的に,物質とは“崩 壊して平衡状態に至る系”と言えよう.こう考えると, アンヒドロビオシス状態の特徴も見えてくる.アン ヒドロビオシス状態では,代謝は停止しているので, 負のエントロピーを取り込むことはできない.しか し,“その内部にエントロピーの増大に抵抗する障壁 を擁して秩序を維持している(崩壊を著しく遅延さ せている)系”と言うことは可能であろう.こう考え ると,物質との違いも明確になり,第三の状態と位 置づけるのも無理はなくなる. それでは,乾燥耐性生物の中にあるエントロピー 障壁とは具体的にどのようなものであろうか.それ を解く鍵は,乾燥時に多量に産生される物質が握っ ているはずである.その一つは糖類であるが,中で もトレハロースは様々な生物界(植物,動物,キノ コ類,線虫,酵母,菌類,ブラインシュリンプ,昆 虫 な ど ) で 見 出 さ れ て い る 2). も う 一 つ は late embryogenesis abundant (LEA)タンパク質である. LEA タンパク質は,最初は植物の種子で見出された が,最近では線虫や昆虫などの動物でも見出されて いる3). では,なぜ自然はトレハロースやLEA タンパク質 を選択したのだろうか.また,前述したエントロピ ー障壁とは具体的にはどのようなものだろうか.物 理化学の教えるところによれば,すべての分子の性 質はその構造に起源がある.われわれは,トレハロ ースや LEA タンパク質の性質とその機能をその構 造特性から実験的・理論的に解明する研究を続けて きた. 2.トレハロースの構造・性質・機能 2-1.トレハロースの構造的特性 トレハロースは二つのグルコースがα,α-1,1 結合 した二糖である.何の変哲もない糖のように見える が,このグリコシド結合に仕掛けがある.二面角φ とψ は伴に-60°付近の値しかとり得ない.すなわち, 安定コンフォメーションが一つしかないのである. その結果,Fig. 1A に示すように,二枚貝が開いたよ うな外形をしている.さらに開いた側(bottom)にある OH 基はすべてエカトリアル配置をしていることも 特徴である. 2-2.トレハロースの水和特性 前述した構造特性によりトレハロースはユニー クな物性を示すはずである.われわれは,まず水和 特性に着目し,示差走査熱測定(DSC)から得られる 不凍水の量を比較した4).その結果,トレハロース の水和数はマルトースやスクロースの場合より多 い こ と が 判 明 し た . こ れ ら 二 糖 は 同 じ 分 子 式 (C12H22O11)で同じ分子量(342.3)をもつので,確 かに立体構造の違いにより異なる水和特性が生じ ることを示している.さらに,水和殻中の水分子の ダイナミクスの情報を 17O-NMR による水のスピン 格子緩和時間T1の測定により調べた4).その結果得 られた動的水和数nDHNは,やはりトレハロース>マ ルトース>スクロースの序列となった.さらに,回 転相関時間の評価から,トレハロース水和殻中の水 分子の運動は,他の二糖に比べて大きく拘束されて いることが判明した. われわれがこのような研究発表をしたのとほぼ 同時期に,世界的にもトレハロースの生理機能をそ の水和特性に関連付けて説明しようとする研究が 活発になり,水溶液の固有粘度,部分モル体積,等 エントロピー,部分モル圧縮率,部分モル熱容量な どのデータが発表された.これらのデータについて はいくつかの総説 5,6)の中で論じてきたのでここで は省略する.それら様々なデータを総合すると,ト
Fig. 1. A: Chemical structure of trehalose,
B: Hydration structure of trehalose.
-2-
乾燥大気下でTα は,105℃において Th を脱水す ることで形成される.Tα は高吸湿性であり,低湿で も水を含む大気にさらされると容易に再水和して Th に戻る.一方,通常乾燥下で 170℃くらいの高温 にすると Tβ が得られるが,これは非常に安定で水 和には高い水蒸気圧が必要である. Tα の特異な性質を理解するためには結晶構造の 情報が必須である.そこで,われわれは,星薬科大 の長瀬弘昌博士のグループの研究に加えていただき, Tα の X 線解析を行った16).その結果,Tα 内の分子 配列や水素結合ネットワークは Th のものと極めて よく似ているが,前者では,一つの結晶軸に沿って 二つの異なる穴(hole-1 と hole-2)が存在することが見 出された.hole-1 はトレハロース二分子がそれらの 重心に対しらせん状に配置することによって構成さ れており,hole-1 の直径とおおよそ等しい直径のポ ケットが穴の側面に存在する.hole-1 と側面のポケ ットは,二水和物中の二つの水分子の位置に一致す る.したがって,hole-1 は側面のポケットを持つ一 次元水チャネルと考えられる.このチャンネルを通 して,Tα 結晶は水を取り込んで二水和物に戻ること が判明した. 結局トレハロースは,Fig. 2 に示した状態・相転 移を通じて,環境に十分水が存在するときは水溜 (sink)として,低湿度のときは水の供給源として働 くということになる.この特異な挙動によって,ト レハロースは脱水状態でも生物を概ね安定に保持 することができると考えられる. 2-3-2トレハロースのガラス状態 緒言で述べたエントロピー障壁という概念からす れば,実は最初に思いつく物性はガラス化である. 分子運動のほぼ凍結されたガラスの中にタンパク質 や細胞膜が閉じ込められると,これら生体分子の秩 序構造は保持されたままになるであろう(エントロ ピーの発散は防がれる).Table 1 は天然に存在する グルコ二糖(二分子のグルコースからなる二糖)と スクロースのガラス転移温度Tgをまとめた5).トレ ハロースのTg値は,天然のグルコ二糖の中では最も 高いが,特別に高いわけではない.それゆえ,乾燥 状態での生物試料の保存にトレハロースが優れた保 護剤として働くメカニズムには,Tg以外の性質も関 係しているはずである.その一つは,エンタルピー 緩和の活性化エネルギーΔErelaxである.これはガラ スを形成している分子の並進拡散に要する活性化エ ネルギーであり,この値は着目しているガラスマト リックスの化学的,物理的安定性を直接測る物差し といってよい.われわれはすべての天然グルコ二糖 に対して ΔErelaxを測定した 5).その結果,トレハロ ースは圧倒的に大きいΔErelax (401 kJ mol-1)をもつこ とが判明した.興味あることに,α,β-1,1 結合をもつ ネオトレハロースでは,Tgは 105℃でトレハロース の値に近かったが,ΔErelaxは223.4 kJ mol-1であり大 きく低下していた.したがって,トレハロースは他 の既知の二糖よりガラス化が容易で(高い Tg),且 つ生成したガラスは劣化し難いと結論できる.すな わち,トレハロースガラスは良質なエントロピー障 壁となりえる. では,なぜトレハロースは安定なガラスをつくの か.この疑問に答えるためには,やはりMD シミュ レーションによる原子レベルの解析が必要である. われわれは,トレハロースとα,β-1,1 結合をもつネオ トレハロースのガラスを比較した17).その結果,コ ンフォメーションの単一なトレハロースは,複数の コンフォメーションをもつネオトレハロースに比べ 密度が均一なガラスをつくるため安定性が高いので はないかとの結論を得た. 一般的に,水はガラス化したマトリックスに対す
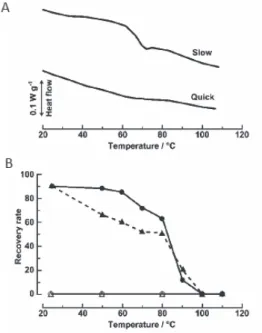
る良い可塑剤である.水の含量が増えれば,ガラス 転移温度は下がり,これは乾燥状態における生体材 料保護にとっては都合の悪い現象である.もしトレ ハロース共存下でガラス化した生体材料乾燥サン プル中に前述したTα のような準安定相が含まれる と,これが環境中の余剰水分を吸収し,ガラス部位 への直接の水の浸入を防ぐであろう.そのとき,ガ ラス状態は安定に維持され生体材料の保存に有利 となる.すなわち,トレハロースの場合は,アモル ファス状態(ガラス状態)と結晶状態が協同的に働 き優れた乾燥保護剤となっている. 2-4トレハロースとアンヒドロビオシス われわれは前節まで述べたトレハロースの基礎物 性の研究を遂行しつつ,トレハロースとリポソー ム18,19)あるいはトレハロースとDNA20)混合系の相互 作用を分光学的手法を用いて調べ,トレハロースの 作用機構を明らかにしてきた.さらに,この研究の 延長線上で in vivo での乾燥保護作用メカニズムの 解明にも挑戦したいと考えていたが,当時すでに三 つの作用メカニズム仮説が提案されていた 2).すな わち,水置換仮説 (water replacement hypothesis),水 閉じ込め仮説(water entrapment hypothesis),およびガ ラス状態仮説(vitrification hypothesis)である. 2-2節で述べたトレハロースの水和特性,特に そのkosmotrope 的性質から,この糖の存在下では水 置換仮説や水閉じ込め仮説が上手く機能するのでは ないかと予想されたが,実際,凍結乾燥酵母細胞に 対する初期の研究において,これらのメカニズムが 確かに機能しているという証拠を得た21,22). このような in vivo 系の実験をさらに発展させた いと考えていたとき,幸運にも,第48 回低温生物工 学会のセミナーで(独)農業生物資源研(現 農研 機構)の奥田隆博士のネムリユスリカに関する講演 を拝聴する機会に恵まれた.ネムリユスリカはアン ヒドロビオシスを乾燥耐性戦略にもつ生物の中で最 も複雑で大きい動物である.奥田グループでは実験 室条件下でネムリユスリカの幼虫をアンヒドロビオ シス状態に誘導・維持することに成功し,乾燥幼虫 の体内では高濃度のトレハロース(乾燥重量の約 20%)が蓄積されていることを見出していた23). われわれはすぐに共同研究を申し込み,トレハロ ース含量が大きく異なる二種類の乾燥幼虫を調製し ていただき,幼虫丸ごとの物理化学測定に着手した. その成果は文献 24 で発表した.α,α-1,1 結合という ユニークな結合のお陰で,トレハロースは992 cm-1 に特有の振動バンドを示す.それゆえ,FTIR 画像分 光法を用いれば幼虫体内におけるトレハロースの分 布を可視化できるのではないかと考えた.この実験 は成功し,その結果,トレハロースを多量に含むサ ンプル(Fig. 3 の slow)ではトレハロースは幼虫体 内にほぼ均一に分布していることが判明した.DSC 測定によると,このサンプルのサーモグラムはガラ ス転移(転移中点は65℃)の存在を示した (Fig. 3A). 別途行われた乾燥→再水和後の生存率の温度依存性 のデータ(Fig. 3B)とこのサーモグラムを並べると, 両者の間には非常に高い相関が見られた.すなわち, トレハロースがガラス状態のときだけネムリユスリ カは蘇生することが判明した.この結果は,トレハ ロースが体全体に行き渡ってガラス化し,それがエ ントロピー障壁となって細胞や組織を破壊から防ぐ というシナリオにぴったり合致したものであり,自 然の巧みさに驚きを禁じえなかった. なお,この研究では,水置換メカニズムも同時に 機能しており,これとガラス化メカニズムは互いに 排他的ではないことも示した.
Fig. 3. A: DSC thermograms for trehalose-contaning
(slow) and –free (quick) samples of dehydrated larvae. B: Dependence of the recovery rate after rehydration on exposure to high temperatures in slowly (filled symbols) and quickly (open symbols) dehydrated larvae. Data from ref. 24. -5- 乾燥大気下でTα は,105℃において Th を脱水す ることで形成される.Tα は高吸湿性であり,低湿で も水を含む大気にさらされると容易に再水和して Th に戻る.一方,通常乾燥下で 170℃くらいの高温 にすると Tβ が得られるが,これは非常に安定で水 和には高い水蒸気圧が必要である. Tα の特異な性質を理解するためには結晶構造の 情報が必須である.そこで,われわれは,星薬科大 の長瀬弘昌博士のグループの研究に加えていただき, Tα の X 線解析を行った16).その結果,Tα 内の分子 配列や水素結合ネットワークは Th のものと極めて よく似ているが,前者では,一つの結晶軸に沿って 二つの異なる穴(hole-1 と hole-2)が存在することが見 出された.hole-1 はトレハロース二分子がそれらの 重心に対しらせん状に配置することによって構成さ れており,hole-1 の直径とおおよそ等しい直径のポ ケットが穴の側面に存在する.hole-1 と側面のポケ ットは,二水和物中の二つの水分子の位置に一致す る.したがって,hole-1 は側面のポケットを持つ一 次元水チャネルと考えられる.このチャンネルを通 して,Tα 結晶は水を取り込んで二水和物に戻ること が判明した. 結局トレハロースは,Fig. 2 に示した状態・相転 移を通じて,環境に十分水が存在するときは水溜 (sink)として,低湿度のときは水の供給源として働 くということになる.この特異な挙動によって,ト レハロースは脱水状態でも生物を概ね安定に保持 することができると考えられる. 2-3-2トレハロースのガラス状態 緒言で述べたエントロピー障壁という概念からす れば,実は最初に思いつく物性はガラス化である. 分子運動のほぼ凍結されたガラスの中にタンパク質 や細胞膜が閉じ込められると,これら生体分子の秩 序構造は保持されたままになるであろう(エントロ ピーの発散は防がれる).Table 1 は天然に存在する グルコ二糖(二分子のグルコースからなる二糖)と スクロースのガラス転移温度Tgをまとめた5).トレ ハロースのTg値は,天然のグルコ二糖の中では最も 高いが,特別に高いわけではない.それゆえ,乾燥 状態での生物試料の保存にトレハロースが優れた保 護剤として働くメカニズムには,Tg以外の性質も関 係しているはずである.その一つは,エンタルピー 緩和の活性化エネルギーΔErelaxである.これはガラ スを形成している分子の並進拡散に要する活性化エ ネルギーであり,この値は着目しているガラスマト リックスの化学的,物理的安定性を直接測る物差し といってよい.われわれはすべての天然グルコ二糖 に対して ΔErelaxを測定した 5).その結果,トレハロ ースは圧倒的に大きいΔErelax (401 kJ mol-1)をもつこ とが判明した.興味あることに,α,β-1,1 結合をもつ ネオトレハロースでは,Tgは 105℃でトレハロース の値に近かったが,ΔErelaxは223.4 kJ mol-1であり大 きく低下していた.したがって,トレハロースは他 の既知の二糖よりガラス化が容易で(高い Tg),且 つ生成したガラスは劣化し難いと結論できる.すな わち,トレハロースガラスは良質なエントロピー障 壁となりえる. では,なぜトレハロースは安定なガラスをつくの か.この疑問に答えるためには,やはりMD シミュ レーションによる原子レベルの解析が必要である. われわれは,トレハロースとα,β-1,1 結合をもつネオ トレハロースのガラスを比較した17).その結果,コ ンフォメーションの単一なトレハロースは,複数の コンフォメーションをもつネオトレハロースに比べ 密度が均一なガラスをつくるため安定性が高いので はないかとの結論を得た. 一般的に,水はガラス化したマトリックスに対す -4- -5-
以上の結果は,トレハロースがアンヒドロビオシ ス誘導・維持の主役であることを示している.しか し,皮肉なことに,ネムリユスリカの体内でガラス が形成されるとき,トレハロース以外の因子が関係 している可能性が少なからずあることもまたこの研 究で明らかとなった.これは一つには,幼虫のガラ ス転移温度は純粋なトレハロースと水の二成分混合 物から予想される理論値とは一致しなかったからで ある.当時,後期胚発生期に大量合成されるタンパ ク 質 (Late embryogenesis abundant protein, LEA protein)が,アンヒドロビオシスが可能な生物で発 現することが知られていたのだが 3),まことにタイ ムリーなことに,奥田グループの黄川田隆洋博士に より LEA 様タンパク質がネムリユスリカでも発見 された 25).ここから,われわれの LEA タンパク質 に関する研究が始動した.
3.
LEA タンパク質の構造・性質・機能
3-1 LEA ペプチドの構造と熱的性質 タンパク質の物性研究と言えば,その三次構造か ら出発するのが普通である.しかし,LEA タンパク 質はそのようなアプローチがとれない.というのも, このタンパク質は天然変性タンパク質(intrinsically disordered protein: IDP)だからである.先行研究では, LEA タンパク質は水中でランダムコイル(disordered) 状態,乾燥するとα-ヘリカルコイルドコイルに構造 化し,機能を発現するらしいことが分かっていた. これら先行研究では,主として線虫や植物のLEA タ ンパク質が対象となっていたが,同様な実験をネム リユスリカについてやってもオリジナリティーに欠 ける.そこで,われわれは多くのLEA タンパク質に 見られる共通モチーフの構造と性質を解明すること に焦点を絞った. LEA タンパク質はいくつかのグループに分かれ るが,高等生物の乾燥耐性に寄与しているのはグル ープ3に属するものである.グループ3の特徴は, アミノ酸一次配列の主要部分が 11 残基の繰返し単 位から構成されていることである.ただし,この繰 返し単位の配列はきっちり決まったものではなく,1 つの LEA タンパク質内の複数の繰返し単位間ある いは異なる生物種の LEA タンパク質間で若干の自 由度がある.さらに,一つの生物種(例えばネムリ ユスリカ)の中にLEA タンパク質は複数種類発現し ているので,繰返し単位をすべて個別に調べていく のは困難である.そこでわれわれは,バイオインフ ォマティクス解析により,植物,線虫,ネムリユス リカ各生物種ごとに 11 残基繰返し配列の共通モチ ーフを導いた26).ここまで配列情報を縮約すること により,以下に示す化学的アプローチが可能となっ た. われわれは,11 残基の繰返し部位が LEA タンパ ク質の活性構造や機能を担っているとすれば,共通 モ チ ー フ 部 分 のみ か ら なる ペ プ チ ド で も天 然 の LEA タンパク質と同様な構造や機能をもつのでは ないかと考えた.そこで,各生物種ごとに共通モチ ーフを2回乃至4回繰返したペプチド(以下LEA ペ プチド)を化学合成した26).例えば,ネムリユスリ カ の LEA タ ン パ ク 質 に 対 す る モ デ ル は , AKDGTKEKAGE を2回乃至4回繰返したものであ る. これら LEA ペプチドの水溶液中および乾燥状態 での構造を FTIR 測定によって調べたところ,水中 ではランダムコイル,乾燥状態ではα-ヘリカルコイ ルドコイル構造をとることが判明し,天然LEA タン パク質に対する先行研究に一致した26).共通モチー フと同じアミノ酸組成をもつが,配列をランダム化 したペプチド(コントロールペプチド)では乾燥状 態で構造化起こらなかったので,LEA タンパク質の IDP 的特質は,繰返し配列部分に起源があることが 明確になった.さらに,LEA ペプチドに対しトレハ ロースを5倍モル程度まで混合した系においても, 乾燥状態における構造化が観測された. さらに,LEA ペプチド自身が比較的高い温度でガ ラス化すること,そのガラス転移点はコントロール ペプチドのそれより高いこと,さらにエンタルピー 緩和の活性化エネルギーもコントロールペプチドの 場合より高いことが判明した26).すなわち,11 残基 の繰返し配列をもつことにより,ペプチドはよりガ ラス化し易く,安定なガラスを形成するようになっ たと言うことができる. アンヒドロビオシスに関係して最も重要な発見は, 実は,トレハロースにLEA ペプチドを混合してガラ ス化すると,トレハロース単独のときよりガラスが 強化されることである.FTIR 測定によると,LEA ペプチドを添加するとトレハロースのOH 基の伸縮振動は低波数側にシフトし,温度変動に対する応答 も小さくなることが示された26).これは,LEA ペプ チド添加により水素結合ネットワークが強化された ことを示している.言い換えると,LEA ペプチド添 加によって,トレハロースのエントロピー障壁とし ての性質がより頑健になったのである.トレハロー スをコンクリート,LEA ペプチドのつくる α-ヘリッ クスを鉄筋と考えると,乾燥耐性生物のアンヒドロ ビオシス状態では,生体内が鉄筋コンクリートのよ うな強固な支持体で充填されるため,細胞や組織が 破壊から護られているのかもしれない. 3-2 LEA ペプチドの機能とそのメカニズム 3-2-1 イオンスキャベンジャー機能 アンヒドロビオシスの誘導過程では,脱水と伴に 生体内のイオン濃度が高まり,浸透圧ストレスがか かってくるはずである.それゆえ,生体内にはイオ ンを捕捉するメカニズムがあるに違いない.LEA ペ プチドがこのような機能を有しているかどうかを調 べるため,NaCl, KCl, MgCl2, CaCl2などの塩共存状 態におけるコンフォメーション挙動を調べた27).そ の結果,いずれの場合も水溶液中ではランダムコイ ルであったが,乾燥状態では1価カチオン共存下で α-ヘリックス,2価カチオン存在下では β-シート構 造をとることが判明した.すなわち,相互作用する 相手に応じて最適な構造に変化し,それらを捕捉し ていることが分かった. 3-2-2 タンパク質凝集抑制機能 通常のタンパク質は水溶液中で固有の立体構造を とる.これは疎水性相互作用などの駆動力により安 定化されているので,結合水が失われるような極度 な乾燥状態に置かれると立体構造は破壊され凝集を 起こす.しかしながら,アンヒドロビオシス状態で は,そのような乾燥誘導凝集は抑制されているはず である.われわれは,LEA ペプチドがそのような抑 制機能をもつかどうかを調べた28).その結果,リゾ チームに対しては4 倍モル以上添加すると顕著な抑 制効果を示すことが判明した.両者の分子サイズを 考慮すると,4 倍モルというのはちょうど LEA ペプ チドがリゾチームの表面を一層カバーできる量であ る.このことから,LEA ペプチドがリゾチームの表 面をシールディングすることにより,乾燥状態にお いてリゾチーム分子同士の直接の接触が防がれ,結 果として凝集が抑制されたと考えられた.実際,MD シミュレーションを実行すると,水溶液状態におい てすでにリゾチームの表面に結合していく様子が観 察された29). 興味あることに,線虫由来の天然のLEA タンパク 質をリゾチーム溶液に添加すると,乾燥→再水和過 程でむしろ凝集が増加するという意外な結果が得ら れた.一つの原因として,タンパク質の電荷状態が 考えられた.すなわち,リゾチームのpI は 9 付近に あるが,線虫由来のLEA タンパク質の pI は 4 付近 であるため,中性の水溶液中では大きな静電的引力 が働き凝集を促進してしまうのではないかと解釈し た.一方,われわれの開発したLEA ペプチドは負電 荷の残基(Glu, Asp)と正電荷の残基(Lys)を同数含ん でいるためpI が 7 付近にあり,相手のタンパク質の 電荷に依らず,適度な相互作用をすると考えられる. この仮説の妥当性は,LEA ペプチド-タンパク質間 の会合定数の測定から裏付けられた30). LEA ペプチドは,タンパク質・酵素の乾燥誘導凝 集を抑制するだけでなく,酵素機能そのものも保持 する能力がある31).われわれは,ラクトースデヒド ロゲナーゼ(LDH)や β-D-ガラクトシダーゼ(BDG)に 表面をシールドするに十分な量の LEA ペプチドを 添加すると,乾燥→再水和後でもフレッシュサンプ ルの70%程度の酵素活性が保持されることを示した. なお,LDH と BDG はサイズが同程度で,pI がそれ ぞれ8.3,4.6 である.したがって,LEA ペプチドは 相手酵素の電荷状態に依らず,それらの機能を保護 できることが明らかとなった. 3-2-3 リポソーム凝集抑制機能 細胞膜もタンパク質と同様,乾燥ストレスにより 凝集・融合を引き起こすと考えられる.われわれは, リン脂質二重膜(リポソーム)を用いて,LEA ペプ チドの細胞膜に対する凝集・融合抑制効果を調べた. 最初の研究では,POPC からなる 100 nm 程度のサ イズの単一層リポソームを調製し,乾燥→再水和後 の粒径分布変化を動的光散乱測定により調べた 32). その結果,添加したLEA ペプチドの濃度依存的に凝 集・融合による粒径増大は抑制されることが分かっ た.特に,リポソームを構成する POPC に対し 0.7 倍モル程度添加すると,ほぼフレッシュサンプルと -7- 以上の結果は,トレハロースがアンヒドロビオシ ス誘導・維持の主役であることを示している.しか し,皮肉なことに,ネムリユスリカの体内でガラス が形成されるとき,トレハロース以外の因子が関係 している可能性が少なからずあることもまたこの研 究で明らかとなった.これは一つには,幼虫のガラ ス転移温度は純粋なトレハロースと水の二成分混合 物から予想される理論値とは一致しなかったからで ある.当時,後期胚発生期に大量合成されるタンパ ク 質 (Late embryogenesis abundant protein, LEA protein)が,アンヒドロビオシスが可能な生物で発 現することが知られていたのだが 3),まことにタイ ムリーなことに,奥田グループの黄川田隆洋博士に より LEA 様タンパク質がネムリユスリカでも発見 された 25).ここから,われわれの LEA タンパク質 に関する研究が始動した.
3.
LEA タンパク質の構造・性質・機能
3-1 LEA ペプチドの構造と熱的性質 タンパク質の物性研究と言えば,その三次構造か ら出発するのが普通である.しかし,LEA タンパク 質はそのようなアプローチがとれない.というのも, このタンパク質は天然変性タンパク質(intrinsically disordered protein: IDP)だからである.先行研究では, LEA タンパク質は水中でランダムコイル(disordered) 状態,乾燥するとα-ヘリカルコイルドコイルに構造 化し,機能を発現するらしいことが分かっていた. これら先行研究では,主として線虫や植物のLEA タ ンパク質が対象となっていたが,同様な実験をネム リユスリカについてやってもオリジナリティーに欠 ける.そこで,われわれは多くのLEA タンパク質に 見られる共通モチーフの構造と性質を解明すること に焦点を絞った. LEA タンパク質はいくつかのグループに分かれ るが,高等生物の乾燥耐性に寄与しているのはグル ープ3に属するものである.グループ3の特徴は, アミノ酸一次配列の主要部分が 11 残基の繰返し単 位から構成されていることである.ただし,この繰 返し単位の配列はきっちり決まったものではなく,1 つの LEA タンパク質内の複数の繰返し単位間ある いは異なる生物種の LEA タンパク質間で若干の自 由度がある.さらに,一つの生物種(例えばネムリ ユスリカ)の中にLEA タンパク質は複数種類発現し ているので,繰返し単位をすべて個別に調べていく のは困難である.そこでわれわれは,バイオインフ ォマティクス解析により,植物,線虫,ネムリユス リカ各生物種ごとに 11 残基繰返し配列の共通モチ ーフを導いた26).ここまで配列情報を縮約すること により,以下に示す化学的アプローチが可能となっ た. われわれは,11 残基の繰返し部位が LEA タンパ ク質の活性構造や機能を担っているとすれば,共通 モ チ ー フ 部 分 のみ か ら なる ペ プ チ ド で も天 然 の LEA タンパク質と同様な構造や機能をもつのでは ないかと考えた.そこで,各生物種ごとに共通モチ ーフを2回乃至4回繰返したペプチド(以下LEA ペ プチド)を化学合成した26).例えば,ネムリユスリ カ の LEA タ ン パ ク 質 に 対 す る モ デ ル は , AKDGTKEKAGE を2回乃至4回繰返したものであ る. これら LEA ペプチドの水溶液中および乾燥状態 での構造を FTIR 測定によって調べたところ,水中 ではランダムコイル,乾燥状態ではα-ヘリカルコイ ルドコイル構造をとることが判明し,天然LEA タン パク質に対する先行研究に一致した26).共通モチー フと同じアミノ酸組成をもつが,配列をランダム化 したペプチド(コントロールペプチド)では乾燥状 態で構造化起こらなかったので,LEA タンパク質の IDP 的特質は,繰返し配列部分に起源があることが 明確になった.さらに,LEA ペプチドに対しトレハ ロースを5倍モル程度まで混合した系においても, 乾燥状態における構造化が観測された. さらに,LEA ペプチド自身が比較的高い温度でガ ラス化すること,そのガラス転移点はコントロール ペプチドのそれより高いこと,さらにエンタルピー 緩和の活性化エネルギーもコントロールペプチドの 場合より高いことが判明した26).すなわち,11 残基 の繰返し配列をもつことにより,ペプチドはよりガ ラス化し易く,安定なガラスを形成するようになっ たと言うことができる. アンヒドロビオシスに関係して最も重要な発見は, 実は,トレハロースにLEA ペプチドを混合してガラ ス化すると,トレハロース単独のときよりガラスが 強化されることである.FTIR 測定によると,LEA ペプチドを添加するとトレハロースのOH 基の伸縮 -6- -7-同一の粒径分布が保持できることが分かった.また, 蛍光物質の漏出試験でも良好な結果が得られ,0.7 モル倍添加系では60%の保持率を示した.幾何学的 計算によると,100 nm サイズのリポソーム表面全体 をLEA ペプチドがシールドするためには,0.5 倍モ ル程度の量が必要である.表面への結合は動的平衡 にあることを考慮すれば,やや過剰の 0.7 倍モルは 妥当な値と言える.すなわち,LEA ペプチドは,リ ポソーム表面をシールドすることにより,リポソー ム同士の直接的接触を防ぎ凝集・融合を抑制してい ると考えると合理的である.さらにFTIR 測定から, LEA ペプチドはリポソーム表面と相互作用する際, β-シートリッチのコンフォメーションをとっている ことも明らかにした. 次の研究では,より実際の細胞に近いリン脂質組 成やサイズをもつリポソームに対して,LEA ペプチ ド添加の効果が調べられた33).これは東大の白樫教 授との共同研究である.卵黄レシチンから調製され た4~5 μm のサイズをもつ巨大リポソームに対して LEA ペプチドを添加すると,やはり濃度依存的に保 護効果を示し,10 mM 濃度のとき約 70%の保護能に 達した. 以上述べた LEA タンパク質とリポソーム表面の 相互作用の存在は FTIR 測定によるリン脂質極性基 の振動バンドシフト等から裏付けられるが,原子レ ベルの相互作用メカニズムについては実験のみでは 分からない.われわれは,MD シミュレーションに より,その詳細を調べた33).その結果,LEA ペプチ ドの中のLys 残基の側鎖がアンカーを降ろしたよう な形で膜表面に突っ込み,その先端のNH3+基はリン 酸基の負の電荷領域と静電的相互作用をしているこ とが判明した (Fig. 4).
4.結 言
以上,トレハロースとLEA ペプチドに関するわれ われの研究を振り返った.実は,トレハロースに関 しては,今回の主題である乾燥保護という枠組みに 収まらない様々な機能が知られている.われわれが (株)林原の奥和之博士(当時)との共同研究で明 らかにした不飽和脂肪酸に対する抗酸化作用 34,35), 最近ではオートファジー誘導分子として熱い注目を 浴びている36).小さいのに何とまあミラクルな分子 であろうか. 緒言において,アンヒドロビオシス状態と物質状 態の本質的違いはエントロピー障壁が有るか無いか だ,と述べたが,結局,この障壁とはトレハロース や LEA タンパク質によってつくられる強化ガラス やこれらの分子シールディング機能のことであろう. この存在によって乾燥状態においても生体構成成分 はその構造と機能をnative な状態に維持できるので ある. われわれの今後の目標としては,トレハロースや LEA ペプチドを使って,哺乳類細胞のエネルギーフ リー常温乾燥保存法を開発することである.ネムリ ユスリカなどの耐乾燥生物では,これら保護物質だ けではなく,抗酸化や遺伝子修復を含む様々なメカ ニズムが相乗的に働いて乾燥耐性を獲得していると 思われる.したがって,一物理化学者が考えるほど この目標達成は簡単ではないと思われるが,定年ま での数年間チャレンジしてみたい.謝 辞
本研究は本文中で名前を出させていただいた方以 外にも多くの方々のご協力を得て可能となったもの です.特に,ほとんどの実験・計算のデータは,博 士研究員の古木隆生氏をはじめとする櫻井研究室の メンバーによるものです.ここに,深く感謝いたし ます. また,研究の多くの部分は,科研費(15H02378 その他)によって行われました.文 献
1) シュレーディンガー著,岡小天,鎮目恭夫訳, phosphate (POPC)ester oxygen (POPC) Lys side chain
Fig. 4. Close-up view of a region where a Lys side chain is interacting with the headgroups of nearby phospholipid molecules.
生命とは何か,岩波書店,2008
2) Crowe JH, Hoekstra FA, Crowe L: Anhydrobiosis, Annu Rev Physiol, 54, 579-599 (1992)
3) Tunnacliffe A, Wise MJ: The continuing conundrum of the LEA proteins, Naturwissenschaften, 94, 791-812 (2007)
4) Kawai H, Sakurai M, Inoue Y, Chûjô R, Kobayashi S: Hydration of oligosaccharides: anomalous hydration ability of trehalose, Cryobiology, 29, 599-606 (1992)
5) Sakurai M: Biological functions of trehalose as a substitute for water, Water and Biomolecules: Physical Chemistry of Life Phenomena (eds. Kuwajima K, Goto Y, Hirata F, Kataoka M, Terazima M), Springer-Verlag, Berlin and Heidelberg, Germany, pp.219-240 (2009)
6) 櫻井実,トレハロースと水の相互作用が織りな す細胞保護機能,低温生物工学会誌,58, 41-51 (2012)
7) Furuki T, Ito T, Asakawa N, Inoue Y, Sakurai M: Effects of trehalose on the swelling behavior of hydrogel - visualization of the preferential hydration of disaccharides, Chem Lett, 38, 264-265 (2009) 8) Sakurai M, Murata M, Inoue Y, Hino A, Kobayashi
S: Molecular-dynamics study of aqueous solution of trehalose and maltose: Implication for the biological function of trehalose, Bull Chem Soc Jpn, 70, 847-858 (1997)
9) Sakakura K, Okabe A, Oku K, Sakurai M: Experimental and theoretical study on the intermolecular complex formation between trehalose and benzene compounds in aqueous solution, J Phys Chem B, 115, 9823-9830 (2011).
10) Akao K, Okubo Y, Ikeda T, Inoue Y, Sakurai M: Infrared spectroscopic study on the structural property of a trehalose-water complex, Chem Lett, 759-760 (1998)
11) Furuki T, Abe R, Kawaji H, Atake T, Sakurai M: Thermodynamic functions of α,α-trehalose dihydrate and of α,β-trehalose monohydrate at temperatures from 13 K to 300 K, J Chem Thermodynamics, 38, 1612-1619 (2006)
12) Furuki T, Abe R, Kawaji H, Atake T, Sakurai M:
Effect of atmospheric pressure on the phase transitions of α,α-trehalose dehydrate, DTA study of the dehydration behavior in open systems: J Therm Analy Calorimetry, 93, 561-567 (2008)
13) Furuki T, Kishi A, Sakurai M: De- and rehydration behavior of α,α-trehalose dehydrate under humidity-controlled atmospheres, Carbohydr Res, 340, 429-438 (2005)
14) Akao K, Okubo Y, Inoue Y, Sakurai M: Supercritical CO2 fluid extraction of crystal water from trehalose dihydrate. Efficient production of form II (Tα) phase, Carbohydr Res, 337, 1729-1735 (2002)
15) Akao K, Okubo Y, Asakawa N, Inoue Y, Sakurai M: Infrared spectroscopic study on the properties of the anhydrous form II of trehalose. Implications for the functional mechanism of trehalose as a biostabilizer, Carbohydr Res, 334, 233-241 (2001)
16) Nagase H, Ogawa N, Endo T, Shiro M, Ueda H, Sakurai M: Crystal structure of an anhydrous form of trehalose: structure of water channels of trehalose polymorphism, J Phys Chem B, 112, 9105-9111 (2008)
17) Kawasaki N, Furuki T, Sakurai M: Molecular dynamics simulation on the glassy states of trehalose and neotrehalose, 低 温 生 物 工 学 会 誌 , 52, 121-124 (2006)
18) Nishiwaki T, Sakurai M, Inoue Y, Chujo R, Kobayashi S: Increasing packing density of hydrated dipalmityol-phosphatidylcholine unilammelar vesicles induced by trehalose, Chem Lett, 1841-1844 (1990)
19) Hanamura T, Asakawa N, Inoue Y, Sakurai M: A solid-state 31P NMR study of the interaction between trehalose and DPPC bilayer, Chem Lett, 713-714 (1998)
20) Zhu B, Furuki T, Okuda T, Sakurai M: Natural DNA mixed with trehalose persists in B-form double-stranding even in dry state, J Phys Chem B, 111, 5542-5544 (2007)
21) Sakurai M, Kawai H, Inoue Y, Hino A, Kobayashi S: Effects of trehalose on the water structure in yeast cells as studied by in vivo 1H NMR spectroscopy, -9- 同一の粒径分布が保持できることが分かった.また, 蛍光物質の漏出試験でも良好な結果が得られ,0.7 モル倍添加系では60%の保持率を示した.幾何学的 計算によると,100 nm サイズのリポソーム表面全体 をLEA ペプチドがシールドするためには,0.5 倍モ ル程度の量が必要である.表面への結合は動的平衡 にあることを考慮すれば,やや過剰の 0.7 倍モルは 妥当な値と言える.すなわち,LEA ペプチドは,リ ポソーム表面をシールドすることにより,リポソー ム同士の直接的接触を防ぎ凝集・融合を抑制してい ると考えると合理的である.さらにFTIR 測定から, LEA ペプチドはリポソーム表面と相互作用する際, β-シートリッチのコンフォメーションをとっている ことも明らかにした. 次の研究では,より実際の細胞に近いリン脂質組 成やサイズをもつリポソームに対して,LEA ペプチ ド添加の効果が調べられた33).これは東大の白樫教 授との共同研究である.卵黄レシチンから調製され た4~5 μm のサイズをもつ巨大リポソームに対して LEA ペプチドを添加すると,やはり濃度依存的に保 護効果を示し,10 mM 濃度のとき約 70%の保護能に 達した. 以上述べた LEA タンパク質とリポソーム表面の 相互作用の存在は FTIR 測定によるリン脂質極性基 の振動バンドシフト等から裏付けられるが,原子レ ベルの相互作用メカニズムについては実験のみでは 分からない.われわれは,MD シミュレーションに より,その詳細を調べた33).その結果,LEA ペプチ ドの中のLys 残基の側鎖がアンカーを降ろしたよう な形で膜表面に突っ込み,その先端のNH3+基はリン 酸基の負の電荷領域と静電的相互作用をしているこ とが判明した (Fig. 4).
4.結 言
以上,トレハロースとLEA ペプチドに関するわれ われの研究を振り返った.実は,トレハロースに関 しては,今回の主題である乾燥保護という枠組みに 収まらない様々な機能が知られている.われわれが (株)林原の奥和之博士(当時)との共同研究で明 らかにした不飽和脂肪酸に対する抗酸化作用 34,35), 最近ではオートファジー誘導分子として熱い注目を 浴びている36).小さいのに何とまあミラクルな分子 であろうか. 緒言において,アンヒドロビオシス状態と物質状 態の本質的違いはエントロピー障壁が有るか無いか だ,と述べたが,結局,この障壁とはトレハロース や LEA タンパク質によってつくられる強化ガラス やこれらの分子シールディング機能のことであろう. この存在によって乾燥状態においても生体構成成分 はその構造と機能をnative な状態に維持できるので ある. われわれの今後の目標としては,トレハロースや LEA ペプチドを使って,哺乳類細胞のエネルギーフ リー常温乾燥保存法を開発することである.ネムリ ユスリカなどの耐乾燥生物では,これら保護物質だ けではなく,抗酸化や遺伝子修復を含む様々なメカ ニズムが相乗的に働いて乾燥耐性を獲得していると 思われる.したがって,一物理化学者が考えるほど この目標達成は簡単ではないと思われるが,定年ま での数年間チャレンジしてみたい.謝 辞
本研究は本文中で名前を出させていただいた方以 外にも多くの方々のご協力を得て可能となったもの です.特に,ほとんどの実験・計算のデータは,博 士研究員の古木隆生氏をはじめとする櫻井研究室の メンバーによるものです.ここに,深く感謝いたし ます. また,研究の多くの部分は,科研費(15H02378 その他)によって行われました.文 献
1) シュレーディンガー著,岡小天,鎮目恭夫訳, phosphate (POPC)ester oxygen (POPC) Lys side chain
Fig. 4. Close-up view of a region where a Lys side chain is interacting with the headgroups of nearby phospholipid molecules.
-8-
Bull Chem Soc Jpn, 68, 3621-3627 (1995)
22) Sano F, Asakawa N, Inoue Y, Sakurai M: A dual role for intracellular trehalose in the resistance of yeast cells to water stress, Cryobiology, 39, 80-87 (1999) 23) Watanabe M, Kikawada T, Okuda T: Increase of
internal ion concentration triggers trehalose synthesis associated with cryptobiosis in larvae of Polypedilum vanderplanki, J Exp Biol, 206, 2281-2286 (2003)
24) Sakurai M, Furuki T, Akao K, Tanaka D, Nakahara Y, Kikawada T, Watanabe M, Okuda T: Vitrification is essential for anhydrobiosis in an African chironomid,
Polypedilum vanderplanki, Proc Natl Acad Sci USA,
105, 5093-5098 (2008)
25) Kikawada T, Nakahara Y, Kanamori Y, Iwata K, Watanabe M, McGee B, Tunnacliffe A, Okuda T: Dehydration-induced expression of LEA proteins in an anhydrobiotic chironomid, Biochem Biophys Res Comm, 348, 56-61 (2006)
26) Shimizu T, Kanamori Y, Furuki T, Kikawada T, Okuda T, Takahashi T, Mihara H, Sakurai M: Desiccation-induced structuralization and glass formation of Group3 LEA protein model peptides, Biochemistry, 49, 1093-1104 (2010)
27) Furuki T, Shimizu T, Kikawada T, Okuda T, Sakurai M: Salt effects on the structural and thermodynamic properties of a Group 3 LEA protein model peptide, Biochemistry, 50, 7093-7103 (2011)
28) Furuki T, Shimizu T, Chakrabortee S, Yamakawa Y, Hatanaka R, Takahashi T, Kikawada T, Okuda T, Mihara H, Tunnacliffe A, Sakurai M: Effect of Group3 LEA protein model peptides on desiccation-induced protein aggregation, Biochim Biophys Acta, 1824, 891-897 (2012) 29) 山川賢太郎, 古木隆生, 古田忠臣, 櫻井実, Group3LEA ペプチドのタンパク質凝集抑制メカ ニズムに関する計算化学的研究,低温生物工学 会誌, 59, 101-105 (2013) 30) 山川賢太郎, 古木隆生, 古田忠臣, 畑中理恵, 黄 川田隆洋, 丹羽達也, 田口英樹, 古澤宏幸, 岡畑 恵雄, 櫻井実,Group3 LEA ペプチドのタンパク 質凝集抑制メカニズムに関する実験的研究,低 温生物工学会誌, 59, 95-99 (2013)
31) Furuki T, Sakurai M: Group 3 LEA protein model peptides protect enzymes against desiccation stress, Biochim Biophys Acta, 1864, 1237-1243 (2016). 32) Furuki, T, Sakurai, M: Group 3 LEA protein model
peptides protect liposomes during desiccation, Biochim Biophys Acta, 1838, 2757-2766 (2014) 33) Furuki T, Watanabe T, Furuta, T, Takano K,
Shirakashi R, Sakurai M: The dry preservation of giant vesicles using a Group 3 LEA protein model peptide and its molecular mechanism, Bull Chem Soc Jpn, 89, 1493-1499 (2016)
34) Oku K,Watanabe H, Kubota M,Fukuda S, Kurimoto M, Tsujisaka Y, Komori M, Inoue Y, Sakurai M: NMR and quantum chemical study on the OH∙∙∙π and CH∙∙∙O interactions between trehalose and unsaturated fatty acids. Implication for the mechanism of antioxidant function of trehalose, J Am Chem Soc, 125, 12739-12748 (2003)
35) Oku K,Kurose M,Kubota M,FukudaS, Kurimoto M, Tujisaka Y, Okabe A, Sakurai M: Combined NMR and quantum chemical study on the interaction between trehalose and diene relevant to the antioxidant function of trehalose, J Phys Chem B, 109, 3032-3040 (2005)
36) Sarkar S, Davies JE, Huang Z, Tunnacliffe A, Rubinsztein DC: Trehalose, a novel mTOR-independent autophagy enhancer, accelerates the clearance of mutant Huntingtin and α-synuclein, J Biol Chem, 282, 5641-5652 (2007)
低温生物工学会誌〔Cryobiology and Cryotechnology〕,Vol. 63, No. 1, 11~20, 2017
酵母に見出したプロリン
/アルギニン代謝に基づく
冷凍・乾燥ストレス耐性機構とパン酵母の育種への応用
奈良先端科学技術大学院大学バイオサイエンス研究科
高木
博史
Freezing and Air-drying Stress Tolerant Mechanisms Mediated by Proline/Arginine
Metabolism of Yeast and its Application to Breeding of Baker’s Yeast
Hiroshi TAKAGI
Graduate School of Biological Sciences, Nara Institute of Science and Technology, 8916-5 Takayama-cho, Ikoma, Nara 630-0192, Japan
(Corresponding author, e-mail: [email protected])
During the fermentation of dough and the production of baker’s yeast, yeast cells are exposed to numerous
and multiple environmental stresses including freezing, air-drying, and high-sucrose, so-called
baking-associated stresses. In addition, such stress conditions could induce oxidative stress in yeast cells
with an increase in reactive oxygen species level due to the denaturation of proteins including antioxidant
enzymes and the severe damage to mitochondrial membrane or respiratory chain. To avoid lethal damage,
baker’s yeast cells need to acquire a variety of stress-tolerant mechanisms. For example, proline and arginine
are important amino acids involved in the stress tolerance of baker’s yeast. In fact, the engineering of proline
and arginine metabolism is a promising approach for the development of stress-tolerant baker’s yeast. We
believe that not only baker’s yeast but also other important industrial yeasts with higher tolerance to various
stresses could contribute to yeast-based industry for the effective production of bread doughs and alcoholic
beverages or the breakthrough of bioethanol production.
(Received Nov. 25, 2016; Accepted Dec. 2, 2016)
緒 言
パン酵母(多くはSaccharomyces cerevisiae の二倍 体株)は発酵によって製造されるパン類に欠かせな い微生物であり,世界中で年間約200 万トンのパン 酵母が生産されている 1).製パン過程におけるパン 酵母の機能としては,1)発酵中に炭酸ガスの生成 によって生地(ドウ)の重量を増加させること,2) 生地の構造や質感を適切に形成すること,3)生地 に特徴的な味や風味を付与することなどが挙げられ る 2).パン酵母は主にクリームイースト,生イース ト(圧搾酵母),ドライイースト(乾燥酵母)の形態 で生産される.日本ではほとんどのパン酵母がクリ ームイーストか生イーストとして生産されており, セミナー総説[Key words: Baker’s yeast, Proline, Arginine, Freezing/air-drying stress; パン酵母,プロリン,ア ルギニン,冷凍/乾燥ストレス]