TH]T MEcaAIVIS7;4 ofIVRGIATrbB SLIG7VALIZVG in
SPERMA TOGOMAL PROLmaRA non of
inPANEsaRED‑BaLMD
7VEnv (CYZVOPS PYRRHO(ZASTER) Tll!S7:IS
by
OZLEM ORAL
A Dissertation
Submitted to the Graduate School ofthe Science and Technology of
Kumamoto University
in Panial Fulfi11ment of the Requirements fbr the Degree of
Doctor ofPhilosophy
< Kumameto University JAPAN, 2008
働㎎μ例5σ擁㎎伽伽
TABLE OF CONTENTS
Abstract...1...
ListofFigures..."....""m..m"....H.H.""..H...H.."H.m..."...""..."""."
LiStOfTableS‑e・‑・‑‑・‑"・‑‑・‑・・‑・・・‑・‑・・・・‑・‑‑・・・‑・・・・e・・・・・‑・・・・・・・・・・・‑・・・・・e・・・・・・・‑‑‑‑‑‑‑‑‑
ListofSymbols...・
Abbreviation..."""""."....‑..."""""""".""‑"""""""""...."""""""""""""".""・‑‑・
Chapter 1: General Introduction...・e.・・・・・・・・・・・・・・・・・・・・・・
1.1Spemiatogenesis...・・・・・.・・・・.・t・・・・・・・e・eee・・・・・・・・・・・・・・・・・・・・・・e・・・・・・・・・・・・・・・・・・・・・・・
1.1.1 Spermatogenesis in mammals...・・・・o・・・・・・・・・・・・・・・・・・・・・・
1.1.2 Spermatogenesis in amphibians (Newts)...・・・・
1.2 Structural and Functional Relationships of Somatic and Germ cells . . . ... . . . .
1.3 Hormonal Regulation ofSpermatogenesis...
1.4 Objectives of the thesis...・・・.・・・・・・・・・・・・・・・・・・・・・・・・・・e・e・・・e・・・・・・・・
1.5References...・・・・・・・・・・・・・・・・e・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・
Chapter 2: Identification of NRGI as a FSH‑upregulated lecal factor and elucidation of
its function on the spermatogonial proliferation in Newt Testis. . . ... . . . ...
2.1intrOdUCtiOn・...・・・・・・・・・・・・・・・・・・・e・・・・・・・・・・・・・・・・e・・・・・・・・・・・e・・・・・・・・・・・・・e''''''''''''''''''''''"""' 2.2Results...,...,...
2.2.I cDNA cloning and stmcture ofnNRG1 isoforms...
2.2.2 mRNA expression ofnNRGI isofbrms in newt testis...
2.2.3 Effect ofFSH on the expression ofnlg‑NRGI protein in different cell types...
2.2.4 Effect ofrecombinant nNRGI on the spermatogonial proliferation
inorganculture...・・・・・・・・・・・・・・・・・・・・・・・ee・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・
2.2.5 Effect ofrecombinant nNRGI on the spermatogonial proliferation
inreaggregateculture...,...,...
2.2.6 Effect of nNRG1 antibody on the spermatogonial proliferation. .. . .. .. . . ...
2.3 Discussion ..., 2.4 Experimental Procedures...・・・・..・・..・・・・・・・・・・・・・・・・・・・・・・・・・・・・・
2.4.1 Animals and Reagents...・・・e.・・...・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・
2.4.2 cDNA microarray analysis, cDNA cloning and RTdtPCR... ... ... ...
2.4.3 Sepailation ofcells at specific stage by laser microdissection (P.A.L.M)...
2.4.4 Real‑time quantitative PCR...
i
IV vi
vlii ix xi
1 2 2 4 6
11 14 16
21 22 24 24 27 29
31
33 35 37 41 41 41 42 42
2.4.5 Expression plasmids, poly(histidine) fusion protein and antibodies. . . ... . . . ... ...
2.4.6 SDS‑PAGE and immunoblotting...
2.4.7 Organ culture, fractionation of somatic and germ cells,
reaggregate culture of germ cells with or without somatic cells. . . ... . . ...
2.4.8 Histology and cell proliferation activity...
2.4.9Statistics...
2.5References...・・・・・・・・e・・・・・・・・・・・・・・・・・・・・・・・・・L・・・・・・・・・・・・・
Chapter 3: ErbB receptors on the NRGI‑induced spermatogonial proliferation... ... ... .. ...
3・1lntrOdUCtiOn・・・・・・・・・・・・・・'・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・e・・・・・・・e・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・
3.2 Results ...・.・b・・・・・・・・・・・・・・・eb・・・・・・・・・・・・・・・・'''e''''e'''''''''''''''''''''''''''''''''''''"""'''""
3.2. 1 Functional ErbB receptors executed in ANRG1‑induced proliferation. . . ... . . . . 3.2.2 Functional intracellular signaling pathways activated in
nNRG1‑inducedproliferation...・...・・..・.・・.・・...・・・・・
3.2.3 Effect of inhibitors on the purified spermatogonia...
3.2.4 Expression ofthe ErbB2 and ErbB4 in different cell types and
spermatogemcstages...,...
3.3Discussion...""...""""""""""."""""""".H""
3.4 Experimental Procedures...・・・・・・e・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・
3.4.I Animals and Reagents...・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・‑・̀・・・・・・・・・
3.4.2 Organ culture oftesticular fragments, reaggregate cqlture ofgerm cells with or wnhout somatic cells and BrdU incorporation assay...
3.4.3 Separation ofspermatogenic stage, fractionation of somatic and germ cells
immunoblotting...‑..."...".""".""""""".
3.5 Refierences...
Chapter 4: Identification ofthe proteins interacted with ErbB2 and ErbB4 receptors...
4.1lntroduction...・・...,...
4・2 ReSUItS ・・・・・・・・・・・・・・・・・e・・・・・・・・・・・・・・・・・・・・・・・・・・・;・・・・・・・・e・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・
4.2. 1 Screening ofthe ErbB2 or ErbB4‑associated proteins
wnh respect to FSH or PRL.."..".‑..""..‑."...".""..."...."..."...H."..."....‑.".."
4.2.2 Identification ofthe SAP155 as an ErbB4‑associated protein... ...
4.2.3 cDNA cloning ofnewt SAP155...
4.2.4 Stage‑specific and cell‑type specific expression ofnewt SAP155... ... ...
4.2.5 Interaction between ErbB4 and SAP155 in different sperrnatogemc stages. . . ...
4.3 Discussion ...‑..."....".,.・‑"・・・・・‑‑・・・・‑・・・・・・・・・・・・・・・・・・・・・・・・‑・・・・・・・‑・・・・・・‑‑‑・
43 44
45 46 47 48 54 55 61 61
64 67
69 71 74 74
74
75 76 82 83 86
86 88 95 97 99 101
4.4Experirnenta1Procedures...
4.4.1 Animals and Reagents...
4.4.2 Co‑immunoprecipitation and Immunoblotting... ... ... ... ... ...
4.4.3 SYPRO RUby and Deep Purple Total Protein Gel Staining...
4.4.4MassSpectrometry...・
4.4.5 cDNA CIoning and Sequencing... ... ... ...
4.5References...・・・・・・・t・・・・・・・・・・・・e・・・・・・・:・・・・・・・・・・・・・・・・
Chapter 5: Summary and Conclusions...
Acknowledgements...・・・・e・・・・・e・・・・・・・・・・・・・・・・・・・
102 102
!02 l02 103 103 l05 108 l12
iii
ABSTRACT
Spermatogenesis is a complex process, consisting of highly organized steps controlled by co‑action of hormones, such as fbllicle‑stimulating hormone (FSH) and luteinizing hormone (LH), and testicular somatic cells. ln newt testis, FSH stimulates spermatogonial proliferation and differentiation into primary spermatocytes through the receptor expressed on its target somatic cells, namely Sertoli cells. Though it is considered that the FSH effects are necessary to be mediated by local paracrine factors, which are secreted from Sertoli cells and act directly on sperrnatogonia within the testis, such factofs have not been identified yet. This dissertation aimed to identify FSH‑upregulated local factors and their signaling mechanism exerted directly in spermatogonial proliferation in newt (Cynops,ru2mbogasteip testis.
Neuregulinl (NRGI) was identified as a novel FSH‑upregulated gene and panial cDNAs encoding two different clones (newt immunoglobulin (nlg)‑type and newt cysteine‑rich domain (nCRD)‑type) were isolated. Expression analysis indicated that both clones were highly expressed at spermatogonial stage than at spermatocyte stage. ln vitro FSH treatment increased nlg‑・NRGI mRNA expression markedly in somatic cells, whereas nCRD‑NRGI mRNA was only slightly increased by FSH. To elucidate the function of newt NRGI, recombinant EGF‑like domain ofNRGI (nNRGI‑EGF) was added to organ and reaggregated cultures with or without somatic cells. The data demonstrated that nNRGI promoted sperrnatogonial proliferation in all culture systems and treatment of the cultures wnh the antibody against EGF‑like domain caused remarkable suppression of spermatogonial proliferation activated by FSH, concluding that nNRGI plays a pivotal role in promoting spermatogonial proliferation by both direct efl}ect on spermatogonia and indirect effect via somatic cells in newt testes.
To identify tyrosine kinase receptors responsible fbr the NRGI signaling, several inhibitors were used in diffbrent culture systems and the results revealed that inhibitors for ErbB2 and ErbB4 significantly suppressed the number ofthe proliferated gemi cells induced by NRGI and '
FSH in both organ and reaggregate culture with or without somatic cells. lmmunoblotting of '
these receptors, ErbB2 and ErbB4, in different testicular cell types and spermatogenic stages showed their expression in both somatic and germ cells at all spermatogenic stages from early spermatogonia to primary spermatoyte stage. Furthermore, as the ligand binding to ErbB receptors induces fbrmation of homo‑ or hetero‑dimers, leading to the activation of distinct
intracellular pathways, including the mitogen‑activated protein kinase (MAPK) and the phosphatidylinositol‑3 kinase (PI3K) pathways, specific inhibitors fbr each pathway were used to reveal functional signaling transduction. The data indicated that PI3K inhibitor was more ethcient in suppression of nNRGI ‑induced spermatogonial proliferation than MAPK inhibitor.
'
Thus, nNRGI can induce spermatogonial proliferation by direct and indirect action on spermatogonia vid the ErbB2 and ErbB4 fecept6rs, which leads to the activation of mainly PBK pathway.
To funher understand the mechanism underlying activation of the spermatogonial proliferation by NRGIErbB, proteins interacting with ErbB2 or ErbB4 were screened. Using co‑
immunoprecipitation method, coupled with matrix assisted laser desorption‑time of fiight‑mass spectrometry (MALDI‑TOF‑MS) or electron spray ionization (ESD, spliceosome associated protein 155 (SAP 1551SF3BiS5), a component of the U2 small nuclear ribonucleoprotein, was detected as a protein interacting with ErbB4 in the testis. Cloning and sequencing revealed a partial cDNA approximately 2.7‑kb in length encoding 930 amino acid residue homologous to mouse SAPI55. Expression analysis in different spermatogenic stages and cell types showed that SAP155 was expressed in germ cells at all the spermatogenic stages examined. On the other hand, co‑immunoprecipitation of SAP155 with ErbB4 antibody as well as the reciprocal co‑
immunopreciphation demonstrated their interaction only in primary spermatocyte stage, suggesting that SAP155‑ErbB4 binding may be linked to the downregulation of the germ cell proliferation in primary spermatocyte stage,
V
Figure 1.1 Figure 1.2 Figure l.3 Figure l.4 Figure 1.5 Figure 2.1 Figure 2.2 Figure 2.3
Figure 2.4
Figure 2.5
Figure 2.6 Figure 3.1 Figure 3.2 Figure 3.3
Figure 3.4
Figure 3.5
Figure 3.6
Figure 3.7
Figure 3.8 Figure 3.9
LIST OF FIGURES
Spemiatogenesis in mammals...・・e・・・・・・・・‑・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・
Spermatogenesis in newts...・
A ctoss‑section ofthe seminiferous tubule...
Newt testicular structure...,...
Hormonal interactions involved in the regulation ofspermatogenesis...
cDNA cloning ofnlg‑NRGI and nCRD‑NRGI...
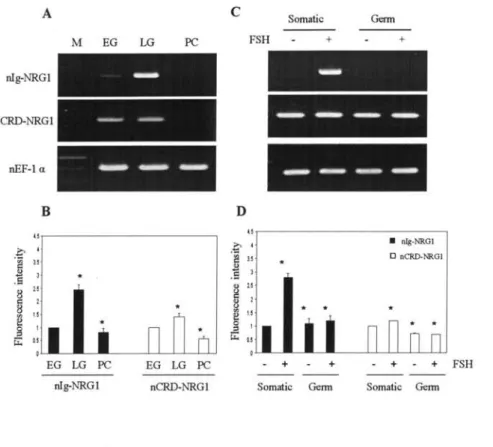
Stage‑specific and cell‑type specific mRNA expression of nNRGI isofbrms.
Effbct ofFSH on the expression ofnlg‑NRGI protein
in different cell types...
Effect of nNRGI ‑EGIT on spermatogonial proliferation
in organ culture oftesticular fragments...
Effect ofnNRGI ‑EGF on spermatogonial proliferation
in reaggregate culture...
Effect of the nNRGI ‑EGF antibody on spermatogonial proliferation.. . . .. . . . Schematic representation of the fbur ErbB receptor tyrosine kinases. ... .. . .. . .. . The ErbB signaling network...
The diffbrential abilities ofEGIF‑like growth factors to interact
with distinct ErbB receptor combinations...
Concentiation‑dependent effbcts ofErbB inhibitors on NRGI‑induced
spermatogonial proliferation in organ culture...
Effects ofErbB inhibitors on nNRGI‑induced spermatogonial proliferation
in reaggregate culture...
Dose‑dependent effbcts ofPI3K and MAPK inhibitors on
nNRGI ‑induced spermatogonial proliferation in organ culture. . . .
Effects of PBK and MAPK inhibitors on nNRGI ‑induced
spermermatogonial proliferation in reaggregate culture...
Effects ofthe inhibitors on purified spermatogonia...
Expression ofErbB2 and ErbB4 in diffbrent spermatogenic stages and
cell types.mm...Hm."...".‑・・・・・・・・・‑・・・‑・・・・・・・・・・‑・‑・・・‑・・・・‑・・・・‑‑・・‑・・・‑・・・・‑・・
3 5 8 10 13 26 28
30
32
34 36 55 57
59
62
63
65
66 68
70
Figure 4.1 Figure 4.2
Figure 4.3 Figure 4.4
Figure 4.5 Figure 4.6 Figure 4.7 Figure 4.8
Figure 4.9 Figure 4.1O
Proteolytic cleavage pathway fbr ErbB4...・・・・・・・・・・・・・・
Co・‑immunopreciphation with ErbB2 and ErbB4 antibodies
upon FSH or PRL treatment...・・・・・・・・・・・・・・・e
ErbB2‑ or ErbB4‑precipitated proteins from FSH‑treated newt testis...
MALDI mass spectrum of the peptides obtained from
ErbB4‑precipitated protein number 1...
Identification ofSAP155 as an ErbB4‑interacting protein...
MSIMS analysis forthe selected peptide...
Validation ofMS result...
The deduced amino acid sequence alignment ofnewt SAPI55
with those ofXenopus, mouse, rat and human...
Expression ofSAPI55 in diffk)rent spermatogenic stages and cell types...
Co‑immunoprecipitation with ErbB4 and SAP antibodies
in different spemiatogenic stages...
84
87 89
91 92 93 94 96
98
1OO
vli
LIST OF TABLES
Table 4.1 Table 4.2 Table 4.3
ErbB2‑binding proteins identified by MALDI‑TOF analysis...
ErbB‑4‑binding proteins identified by MALDI‑TOF and ESI analysis...
Primers used fbr cDNA cloning...
90 90 104
Symbols ofamino acids
A B C D E F G H I K L M N P Q R S T V W Y z
Symbols ofnucleic acid
A C G T U
LIST OF SYMBOLS
Ala
dsx Cys Asp Glu Phe Gly His
Ile
Lys Leu Met Asn
Pro Gln Arg Ser Thr Val Trp Tyr Glx
iX
Alanine
Asparagine or Asparatic acid Cysteine
Asparatic acid Glutamic acid Phenylalanine Glycine Histidine Isoleucine Lysine Leucine Methionine Asparagine
Proline Glutamine Arginine Serine Threonine Valine Tryptophan Tyrosine
Glutamine or Glutamic acid
Adenosine Cytidine Gaunosine Thymidine Uridine
BrdU ,
BSA cDNA
Cy3
DNA
DTT ECL
EGilFR
EGTA GSH GSSG HRP
IPTG
L‑15
LB
mRNA
NCBI
OVA
PBS
PMSF PRL
Tris
ABBR}i VIATIONS
Bromodeoxyuridine Bovine serum albumin complementary DNA
indocarbocyanine deoxyribonuclease Dithiothreitol
Enhanced chemi luminescence Epidermal growth factor receptor Ethylene glycol tetraacetic acid Glutathione reduced
Glutathione oxidized Horse radish peroxidase lsopropyl‑P‑D‑thiogalactoside Leibovitz‑ 1 5
Luria‑Bertrani
messenger Ribonucleic acid
National Center fbr Biotechnology infbrmation
Ovalbumin
Phosphate buffered saline Phenylmethyl sulfonyl fiuoride Prolactin
Tris (hydroxymethyl) aminometharie
CHAPTER l
Gθηθm11力 70伽α∫oη
(]hapter 1
1.1 SPERMATOGENESIS
Spermatogenesis is an essential process fbr sexual reproduction in development. lt takes place within several structures ofthe male reproductive system and includes sequential mitotic divisions and two meiotic divisions. Number of the mitotic divisions is species‑specific and stable (Roosen‑Runge, 1969). The entire process can be broken up into several distinct stages, each corresponding to a particular type of cell: spermatogonium, primary spermatocyte, secondary spermatocyte, spermatid and sperm.
1.1.1 ,Sipermatogenesis in mammals
Seminiferous tubules of the testes are the starting point for the spermatogenesis. Diploid type A spermatogonia divide mitotically to produce diploid intermediate gonial cells which then produce primary spermatocytes. Each preleptotene primary sperrnatocyte duplicates its DNA and subsequently undergoes two meiotic divisions to produce haploid spermatids. This division provides sources of genetic variation, such as random inclusion of either parenta1 chromosomes or chromosomal crossover, to increase the genetic variability ofthe gametes. Spermatids, which possess half the normal complement of genetic material, are then developed into spermatozoa, mature male gametes in many sexually reproducing organisms.
CZIapter 1
Type A spermatogonia
[IYpe B . ‑ i,,, e .t. ‑ .."k.i",If!.!il'd """['T', i"' 'l d"̀vH'ljl. '"'L ,i,‑'t'i.f;:'i./s spermatogoma
'
P"slPi,IIi,S,[,illl/,atigiC,yt,e ,,,efi.Il2}}. ,,.e.",ts'' Cytopiasmicbridges
‑ " t‑ It tV. tt
se,co,,nd.a,ry,,ejgeam,a,!o,s,yte :.I.altr.as,pt.."S.w..Si‑h[es,sSin・:'gS.,,
;S;.!S,"'i,ptt:IXSinVS.'l":・Si'i$!.S'・!・(;ell":'SilptW
,..,, K","wtwtVWaS"g/kV
Residual bodies '#' ‑'‑ ...e. ‑m‑‑ ‑‑ . , 'k,,. .., . '"".. ‑eX
"e}ga,lb ' Maturesperm
Figure 1.1 Spermatogenesis in mammals Different types of germ cells during spermatogenesis are shown.
Cytoplasmic bridges form between differentiating cells facilitating synchrony of the wave of germ cell development. Adodij}ed.fiom Developmental Biology Sbott jFl Gilbert le(V, SinuaerAssociates, 7'h ed, Flig. 19.20, p 63O (Bloom and]Ecxwcett, 197Lsp
3
enapter 1
1.1.2 Siperntatngenesis in amphibians (7Vewtw
In newts, testicular function shows marked seasonal changes, as is the case with many species of amphibians living in temperate climates (Lofts, 1974; Pierantoni, 1998). During the relative warm period, spermatogenesis occurs within sperrnatocysts which are placed in the lobules, corresponding to the seminiferous tubules in mammals, and during the relative cool period, sexual characteristics develop (Tanaka and Iwasawa, 1979; Tanaka and Takikawa, 1984).
Primary spermatogonia proliferate synchronously through 7 mitotic divisions and then differentiate into primary sperrnatocytes in the 8th generation. in newt testis, mitosis of spermatogonia and meiosis of spermatdcytes start from spring to early autumn. Transfbrmation of spermatocytes to spermatids occurs in the end of summer; accordingly, a large number of spermatids can be observed in autumn. After morphogenetic processes of spermatids, mature male gametes, spermatozoa, are developed in late autumn.
C]lapter 1
A
Sperrnatogonia
Spermatocyte
Spermatid
3",'e.g....
/tt t tt t/ t/
gt i:
!kve"e':
ig!"egti
iiirs}S,,iitMS
o+X Mitosis
oo+
o 1S̀ Meiotic divison
ooi X>4 2nd Meiotic divison+x
oo
Elongated
t
SpermlogeneslsB
1 St generation
2nd 3rd 4th 5th 6th 7th
g
generatron generatson generatson generation generatson generation
g
8th generation
g
"・..,,....20
lst mitotic division
...,H21 '''''''''・・・.22 ''''''''・‑‑23
'"M'H'・‑24
・‑.,..,.,.,25 ,.,,","."26 7th mitotic division
'''''''・・..27
Meiosis Initiation (Preleptotene pdmary spermatocyte)
mature sperm
Figure 1.2 Spermatogenesis in newts A) After several round ofmitosis, sperrnatogonia differentiate into primary sperrnatocytes that undergo two meiotic divisions, giving rise to roimd spermtatids that eventually diflferentiate into mature sperm. B) After the 7th mitotic division, spermatogonia initiate meiosis in the 8th gerieration and differentiate into preleptotene spermatocytes ooodij7ed.from Yazawa et aL, 2000cO.
5
Chapter 1
1.2 STRUCTURAL and FUNCTIONAL RELATIONSHll?S of SOMATIC and GERM CELLS
The mammalian testis consists of two compartments: the interstitium and the seminiferous tubules. The composition of the interstitium difliers among species, but in general it contains Leydig cells, macrophages, endothelial cells, lymph space and blood vessels (Fawcett, 1973).
The seminiferous tubules are avascular and contain different types of germ cells embedded in Sertoli cells (de Kretser and Kerr, 1988). Peritubular myoid cells and a basal lamina are located between two compartments and surrounding the seminiferous tubules.
The function of the interstitial Leydig cells is the production and secretion of androgens.
Lutemizing hormone (LH) is the main regulator ofLeydig cell function, but the cells also take part in a complicated interplay between different cell types in the testis. Factors secreted by Sertoli cells have been reported to affect Leydig cell mitosis, the number ofLH, gonadotropin releasing hormone (GnRH) receptors and LH‑stimulated testosterone secretion in viro (Kerr and Sharpe, 1985). Peritubular myoid cells are in close contact with the Leydig cells in interstitium (Skinner et al., 1991) and together with the Sertoli cells they produce an extracellular matrix to provide structural support fbr the spermatogenic epithelium (Skinner et al., 1985). Peritubular myoid cells produce a potential regulator of Sertoli cell function, termed PmodS (Reritubular myoid cell factor which modulates Sertoli cell function; Skinner and Fritz, 1985; 1986). in cell cultures, this paracrine factor has a marked impact on a number of Sertoli cell functions: it increases the production of androgen binding protein (Tung and Fritz, 1980; Hudson and Stocco, 1981) and transfenin (Holmes et al., 1984; Skinner and Fritz, 1985). Sertoli cells are the only somatic cell type of the spermatogenic epithelium. They form functional blood‑testis barriers consisting oftightjunctions located in between neighbouring Sertoli cells (Sharpe, 1983; Saez et al., 1987; Bardin et al., 1988). The conuibution of Sertoli cells to these barders is important because junctional contacts between adjacent Sertoli cells restrict the passage of many growth factors, nutrients and hormones from the basal to the adluminal compartment of the seminiferous tubule. Germ cell development takes place in the protected environment of the seminiferous tubules, as a defined series ofdevelopmenta1 events (Clermont and Harvey, 1967).
C7laapter 1
Spermatogenesis does not seem to rely on direct dontrol of the germ cells by the endocrine system, but rather is dependent on.hormone action on Sertoli cells and cell‑cell interactions between germ cells and Sertoli cells (Griswold et al., 1988; Skinner, 1991). In this respect, Sertoli cells are also referred to as nursing and supporting cells of the germinal epithelium. For example, Sertoli cells provide an energy‑yielding substrate fbr the developing germ cells in the form of lactate (Jutte et al., 1983), which play an important role in detoxification of antisperrnatogenic compounds (Grootegoed J.A., et al., 1989). The most important hormonal regulator of spermatogenesis, FSH, exerts its actions by regulating activities of Sertoli cells directly, because this is the only oell type in males that expresses the FSH receptor (Heckert and Griswold et al., 1991). Androgen action is involved in effects not only on the Sertoli cells but also on the androgen receptor expressing peritubular myoid cells (Ruizeveld de winter et al.,
1991). Direct action of androgens on germ cells is not very likely, because these cells do not express androgen receptors (Anthony et al., 1989)
7
(]hapter 1
vas
1 deferens Epididymis
/ T;estis
1
1Xt"ii..
Seminiferous tubule
r・
Cross section of seminiferous tubule
'"i>,,,t,,,,..,';i' il・・,t .Y.,.i,.・g・"ti"" , ,.is,Slty...・''
../1 t.
's'i'1'1'/''1'11'1'11 ' 1't '1/
.' ‑, ,".. I‑1yrnph space ' ,.1.I'‑s 'f"'1'" Ti"1 , i'+' '' '/' ,4. ,.' r.... ,,.3'pti. '''
ik"K"kipt:
Seminiferous tubule
Y
Sertoli cell tight junctions Basal lamina
Interstitium Peritubular myoid cells and endothelial cells
Blood vessel
Leydig cell
Spermatogonium Spermatocyte Round spemiatid Elongating spemiatid
Figure 1.3 A cross section of the seminifk}rous tubule The relationship between Sertoli and germ cells (Z]lym, 197rp.
Ghupter 1
Newt testis consists ofthe lobules which is ensheathed by basement membranes. Clonally derived germ cells are enclosed in a cyst, the smallest unit ofthe testis, and surrounded by Sertoli cells within the lobules. Leydig‑like cells, also called pericystic cells or lobule boundary cells, blood vessels and red blood cells are located outside the lobules. The testis displays well‑marked zones of spermatogenic cell types because the lobules formed at the cephalic region gradually acquire more caudal positions as the cells mature (Callard et al., 1978). All spermatogenic stages from spermatogonia to the most advanced stage can be observed in longitudinal sections, which permit to isolate germ cells in specific stages ofspermatogenesis.
The functions of pericystic and Sertoli cells are not well known in newt testis. A5‑3P‑
hydroxysteroid dehydrogenase (3P‑HSD) reaction is detected strongly in pericystic cells in glandular tissue, but barely in germ cells or Sertoli cells (Imai and Tanaka 1978). Sertoli cells proliferate along with the proliferation of spermatogonia and they are degenerated after spermiation (Abe, 2004). In each cyst, germ cells, which are in the same stage of diflierentiation, rnake a close contact with Sertoli cells. The blood‑testis banier, formed by Sertoli cells, shows size selectivity in newt testis. It allows small molecules (‑‑500 Da) to penetrate into the cyst, but not larger ones (>1.9 kDa) (Jin et al., 2008). Binding experiments for fractionated germ and somatic cells indicated that FSH binds to somatic cells (mostly Sertoli cells) but not to gerrn cells (Ji et al., 1995). Funhermore, a cDNA encoding a FSH receptor (FSH‑R) is isolated from the Sertoli cells (Nakayama et al., 2000) and FSH‑activated Sertoli cells are shown to mediate proliferation of spermatogonia and their differentiation into primary spemiatocytes (Abe and Ji,
1994; Abe, 2004).
9
(]hapter 1
A
‑‑‑‑>
B
Cyst
L‑‑‑v‑."/
Lobule
Figure 1.4 Newt testicular structure A) Newt testis consists ofseveral zones each consisting ofthe same stage of spermatogenesis; sperrnatogonial stage (SG), primary spermatocyte stage (PC), round spermatid (RT) and elongated sperrnatid stage (ET), B) A cyst, smallest unit of the testis, consists of a germ cell clone derived from a single sperrnatogonium, and Sertoli cells. Cysts are packed in a lobule that is surrounded by basement membrane. Some Leydig‑1ike cells, called pericystic cells, are placed outside the lobules (Abe, 2004).
enupter 1
1.3 HORMONAL REGULATION of SPERMATOGENESIS
Hormonal control ofspermatogenesis varies among species. in most species, however, it is initiated by the interaction of the hypothalamus, pituitary gland and testicular somatic cells at puberty. The gonadotropins, luteinizing hormone (LH) and fbllicle stimulating homione (FSH) are secreted by the pituitary gland which is stimulated by the gonadotropin releasing hormone (GnRH) from the hypothalamus.
In mammals, LH acts upon Leydig cell and it is responsible fbr the production of testosterone, the male hormone that exerts both endocrine activity and intratesticular activity such as spermatogenesis. FSH stimulates both the production of androgen binding protein by Sertoli cells and the fbrmation ofthe blood‑testis banier. Androgen binding protein is essential fbr concentrating testosterone in levels high enough to initiate and maintain spermatogenesis.
Therefbre, Sertoli cells themselves mediate parts of spermatogenesis through hormone production. They are a!so capable of producing inhibin hormone, which inhibits pimitary gonadotropin production, preferentially that of FSH (Burger and Igarashi, 1988) and thus it is considered to be a part ofthe pituitarylgonad feedback mechanism.
ln amphibians, development of the gonads is affected by the ambient temperature (Tanaka and Takikawa, 1984; Rastogi et al., 1987). Comparison of the biological propenies of newt gonadotropins showed FSH is excreted by the pituitary only in July, while the activity ofFSH 'and LH is very similar in February (Tanaka and Takikawa, 1984). In newts, presence of both '
FSH and LH in pituitary was demonstrated by isolation of newt FSHB and LH6 cDNAs (Saito et al., 2002) but there has been no report on the biological action of newt LH and FSH, because these proteins in purified fbrm are not available (Kano et. al., 2005). Bullfrog LH stimulates testosterone production and spermiation, while bullfrog FSH maintains testicular weight and stimulates spermatogenesis (Tanaka et al., 2004). Ovine FSH also promotes spermatogenesis, while LH has poteney in testosterone production in newt testis (Tanaka and Takikawa, 1984).
Secretion of estrogen is primarily stimulated by ovine FSH (Callard et al., 1978; Bolaffi and Callard, 1981) but the role of estrogen in newt sperrnatogenesis has not been clarified yet. Direct evidence is fbund fbr gonadal negative feedback control ofplasma gonadotropins in amphibians
11
Chapter 1
by using iadioimmunoassay (McCreery and Lichg 1984; Pavgi and Licht, 1989). ln newt testis, gonadoectomy increased the expression levels of LHP and FSHB, and testosterone replacement inhibited the expression ofLHP, but not FSHP, indicating that FSHP expression is regulated by a non‑steroid factor (Kano et al., 2005). The negative feedback system by andfogen, namely testosterone is weakly regulated in newts (Kano et al., 2005).
Chupter 1
1lestis
lvpothalamus
. 3.t'
‑1‑t/‑ ・・1, 'tt't"'l t'"
ttitt t;/tttt , ・l.g,‑ ,as‑ ",‑.',,.. k' itt
'i't.:"III'".t.1/',/.ll"a Yv/,'1' s・{ .,"・,
, ,.t..1.tt.tl/ ....1 ..t.. . tt''tf'fil・;ii//.l‑'‑,,‑,‑.1.,/.,
'1,1 /・l;E・ /'/ '.,‑ / )n
r.,. .., .‑k'・/E ',;'・,/i,c{'k[.',‑',・?y';,sit/,'te'
Pimimry LH,i,・/・' ia ll.,/ll]1,; '・si 4"#' ",‑ f.
FSH
1'i;/
‑.,;'.g・ ,, lt.,1 l.,ttlj,"1,l,l;'
'4mu
t"
.‑,t・' T
・ ・/'imp, , ep,dig eeU
:,.vSat,1・...
.,.,・1E
1
Peiitnbular eeU
tt
‑modS
't'" 't" ' ''St'.,",,.h.
Inhibin tt t//t
ttttt
F .‑pt'a・ t'i・ ‑a.T,/i. v.. ./
Sertoli eeU
,/t
t
x;,
'k.
k GenuceU
'kltetw. ‑..,t‑..,..
‑ ‑hA‑
,‑‑"‑,e
' t, ''e
't'!
,t t/,/g
,..n,.
wo‑..:,.
Figure 15 Hormonal interactions involved in th,e regulation of spermatogenesis GnRH, gonadotropin‑
releasing hormone; FSH, fbllicle stimulating hormone; LH, luteinizing hormone; T, testosterone; PmodS, peritubular myoid cell factor which modulates Sertoli cell function.
13
Chapter 1
1.4 OBJECTIVES OF THE 1llESIS
In adult organism, reproduction is one of the most intriguing aspects of development.
Spermatogenesis produces mature male gametes, which are able to fenilize the 'counterpart female gamete to produce a single‑celled individual known as a zygote. This is a cornerstone of sexual reproduction and involves two gametes each contributing half the normal set of chromosomes (haploid) to result in a chrornosomally normal (diploid) zygote. The abnormalities during the process cause male infenility or abnormal zygotes which would not survive fbr long time after conception. The cooperation with several hormones and the factors locally secreted from gonadal somatic cells are now generally accepted to play a crucial role in regulating spermatogenesis. The mechanisms behind this phenomenon are the great interest to understand the treatments arid cures for male infenility or to provide targets for contraceptive development.
The first aim ofthe current study was to identify the local factor expressed highly upon FSH treatment in newt testis. The second aim was to investigate experimentally the role ofthe factor on the spermatogonial proliferation and the last one was to study the signaling mechanism ofthe factor in newt spermatogenesis.
ln the general introduction of this dissertation (Chapter 1) an overview is given about the process of the spermatogenesis, including stages and location in both marmials and urodels.
Attention is then focused on the role of somatic cells and the factors that infiuence gamete development. This overview is completed with a description of the hormonal regulation of spermatogenesls.
The fbllowing chapter fbcuses on the isolation of the factor locally expressed in testicular cells in response to FSH treatment. Emphasis is put on the expression and the role ofthe factor in spermatogonial proliferation using diflierent culture systems (Chapter 2).
In chapter 3, the dissertation is concerned with the receptors of the factor, which play a role during spermatogenesis. Specific inhibitors fbr each receptor were used in diflierent culture systems to explore the functional receptors in spermatogonial proliferation. The expression pattern ofthe receptors is presented in diflierent spermatogenic stages and cell types.
C7laapter 1
Since ligand‑induced phosphorylation ofthe receptors serve as docking sites for a variety of signaling molecules whose recruitment stimulates numerous intracellular signaling cascades, the proteins interacting with the receptors were analyzed by immunoprecipitation coupled with matrix assisted laser desorption‑time of flight‑mass spectrometry (MALDI‑TOF‑MS) or electron spray ionization (ESD in chapter4.
15
CZtapter 1
1.5 REFEI{ENCES
e Abe, S.‑I., Ji, Z.‑S., 1994. Initiation and stimulation of sperrnatogenesis in vitro by mammalian fbllicle‑stimulating hormone in the Japanese newt, Cyiops nyrnhogaster. int 1 Dev. Biol. 38, 201‑208.
e Abe, S.‑I., 2004. Hormonal control ofmeiosis initiation in the testis from Japanese newt, Cynops,nyrthogtister. Zool. Sbi. 21, 691‑704.
e Anthony C.T., Kovacs W.J., and Skinner M.K, 1989. Analysis ofthe androgen receptor in isolated testicular cell types with a microassay that uses an affinity ligand. Ehdocrinology 125, 2628‑2635.
e Bardin, C.W., Cheng, C.Y., Musto, NA., and Gunsalus, G.L., 1988. WThe Sertoli cell;' in:
The Physiology of Reproduction (E. Knobil and J.D. Neil, eds.). Vol. 1, pp. 933‑974.
Raven Press New York.
'
'
e Bloom W., Fawcett D.W., 1975. A textbook of histology. WB Saunders Company, '
Philadelphia London Toronto, Tenth edition
e Bolaflri J.L., Callard I.P., 1981. ln vivo regulation of steroidogenesis by ovine gonadotropins in male and female mudpuppies, ATecturus maculosus (Rafrmesque). Gen.
Comp. Eleidocrinol. 44, 108‑116.
e Burger, H.G. and Igarashi, M, 1988. lnhibin: definition and nomenclature, including related substances. X CIin. EleidocrinoL Metab. 66 885‑886.
'
e Callard G.V., Petro Z., and Ryan K. J., 1978. Phylogenetic distribution of aromatase and other androgen‑converting enzymes in the central nervous system, Endocrinology 103, 2283‑2290.
'
e Callard I.P., Callard GV., Lance V., Bolafli J.L., Rosset J.S., 1978. Testicular regulation in nonmammalian vertebrates. BioL Reprod. 18, 16‑43.
'
enapter 1
e Clermont Y., and Harvey S.C, 1967. Effects of hormones on spermatogenesis in the rat, Ciba Iloundotion Conbquious Ehdocn'nology 57, 173‑179.
e De Kretser D., Kerr J., 1988. The cytology of the testis. in: Knobil E, Neill J (eds) 7Jhe Pbysiology ofReproduction. Raven Press, New York, pp 837‑932.
e Dym, M., 1977. The male reproductive system. in L. Weiss and R.O. Greep (eds), Histology, 4th Ed. McGraw‑Hill, New York, pp. 979‑1038.
e Fawcett, D.W., 1973. 0bservations on the organization ofthe interstitial tissue ofthe testis and on the occluding cell junctions in the seminiferous epithelium. ln: Advances in biosciences, 10: The masculine gender (Raspe, G, Bernhard, S., eds.), p. 83‑99. 0xfbrd:
Pergamon Press
e Gilbert, S., 2000. Developmenta1 Biology, Seventh Ed. Sinauer, Massachussets, USA e Griswold M.D., Morales C., Sylvester S.R, 1988. Molecular biology of the Sertoli cell.
Oof Rev. Reproa Biol. 10, 124 ‑l61.
e Grootegoed J.A., Den Boer P.J., 1989. Energy metabolism of sperrnatids: a review. ln:
Hamilton DW, Wanes GMH (eds.), Cellular and Molecular Events in Sperrniogenesis as Targets fbr Fertility Regulation. London: Cambridge University Press pp 193 ‑215.
e Heckert L.L., (lriswold MD., 1991. Expression of fbllicle‑stimulating hormone receptor mRNA in rat testes and Sertoli cells. Mol. Elridocrinol. 5 670‑677.
'
e Hudson I.C., Stocco D.M, 1981. Peritubular cell infiuence on the efficiency of androgen binding protein secretion by Sertoli cells in culture. Ilhdocrinology 108, 1362‑1368.
e lmai K., Tanaka S., 1978. Histochemical and electron microscopic observations on the steroid hormone‑secreting cells in the testis of the Japanese red‑bellied newt, CjJuaops nyrnhogasterlzyrnhogasten Dev. Growth DtfiZir. 20, 151 ‑167.
17
(]hapter 1
. Ji, Z‑S, Kubokawa K., Abe S‑I., 1995. Promotion of differentiation of newt primary sperrnatocytes into spermatids by mammalian FSH via Sertoli cells. X Ex/p. ZboL 272, 374‑
383.
e Jin, Y., Uchida, I., Eto, K, Kitano, T., Abe, S.‑I., 2008. Size‑selective junctional banier and Ca2+‑independent cell adhesion in the testis of Cbenops wrrkagaster: expression and function ofoccludin. MoL Rezproa Dev. 75, 202‑216.
e Jutte, N.H.P.M, Jansen, R., Grootegoed, J.A., Rommerts, F.F.F.G. and Van Der Molen, H.J. 1983. FSH stimulation of the production of pymvate and lactate by rat Sertoli cells may be involved in hormonal regulation ofspermatogensis. X Reprod Eert. 68, 219‑226.
e Kano Y., Nakano T., Kumakura M, Wasa T., Suziki M, Yamauchi K., Tanaka S., 2005.
Seasonal expression ofLHP and FSHB in the male newt pimitary gonadotrophs. Gen. Com.
EhdocrinoL 141 248‑258.
'
e Kerr J.B., and Sharpe RM, 1985. Follicle‑stimulating hormone induction ofLeydig cell maturation. Eleidocn'nology 1 16, 2592‑2604.
e Lofts B., 1974. Reproduction. Iri PItysiology ojCthe Amphibia (ed.Lofts B), pp. 107‑218.
New York: Academic Press
e McCreery, B.R., Licht P., 1984. Effects of gonadectomy and sex steroids on pituhary gonadotrophin release and response to gonadotrophin‑releasing hormone (GnRH) agonist in the bullfrog, Rana catesbeiana. Gen. Cbmp. iindocrinoL 54, 283‑296.
e Nakayama Y., Yamamoto T., Oba Y., Nagahama Y., Abe S‑I., 2000. Molecular cloning, functional characterization and gene expression of a fbllicle‑stimulating hormone receptor in the testis ofnewt, Cjymopspyrrhognster. Biochem. Biopas. Res. Commun. 275, 121‑128.
e Pavgi S., Licht P., 1989. Efli2cts of gonadectomy and steroids on pituitary gonadotropin secretion in a frog, Rana pipiens. BioL Reprod. 40, 40‑48.
Chapter 1
e Pierantoni, R., 1998. R. Pieiantoni , Male reproduction system, amphibians. in: E. Knobil and J.D. Neill, Editors, E)zq)rcZQpedia ofReproduction, Academic Press, San Diego, pp. 1O‑
15.
e Rastogi R.K., Lela L., Delrio G., di Meglio M., Russo A., Chieffi G., 1987. Environmenta1 infiuence on testicular activity in the green frog, Rana esculenta, X Ebep. ZboL 206, 49‑64.
e Roosen‑RungeEC., 1969. Comperative aspects ofsperrnatogenesis. BioL Reprod i, 24‑39.
e Ruizeveld de Winter J.A, Trapman J., Vermey M., Mulder E., Zegers N.D., Van der Kwast T.H., 1991. Androgen receptor expfession in human tissues: An immunohistochemical study. X Histochem. dytochem. 39, 927‑936.
e Saez, J.M, Perrard‑Sapori, MH, Chatelain, PG, Tabone E., and Rivarola MA., 1987.
Paracrine regulation oftesticular function. X Steroid Biochem. 27, 3 1 7‑329.
e Saito A., Kano Y, Suzuki M, Tomura H., Takeda J., Tanaka S., 2002. Sequence analysis and expressional regulation of mRNAs encoding P‑subunits of fbllicle‑stimulating hormone and luteinizing hormone in the red‑bellied newt, Cjynops ,nvrr:hognsteL BioL Reprod. 66, 1299‑1309.
e Sharpe R.M., 1983. Local control oftesticular function. e. X lixp. Ph)nsioL 68, 265‑287.
e Skinner MK, Tung P.S., Fritz I.B., 1985. Cooperativity between Sertoli cells and testicular peritubular cells in the production and deposition of extracellular matrix components. .L CellBioL 100, 1941‑1947.
e Skinner M.K., Fritz I.B., 1985. Testicular peritubular cells secrete a protein under androgen control that modulates SertDli cell function. Proc. AlicitL Acad. Sbi. USZt 82 l 1̀Fl18.
'
e Skinner M.K., Fritz I.B., 1986. Identification ofa nonmitogenic paracrine factor involved in mesenchymal epithelial cell interactions between testicular peritubular cells and Sertoli cells. MoL CenEleidocrinol. 44, 85‑97.
19
caupter 1
e Skinner M.K., 1991. Cell‑cell interactions in the testis. E>2docr. Rev. 12, 45‑77.
e Skinner M.K., Norton J.N., Mullaney, BP., Rosselli M, Whaley P.D., and Anthony C.T., 1991. Cell‑cell interactions and the regulation oftestis function, Ann. IV: Z Acad Sci. 637, 354‑363.
e Tanaka S., Iwasawa H., 1979. Annual changes in testicular structure and sexual character of the Japanese red‑bellied newt, (Jlynops evrnhogaster evrnhogaster. ZboL Mag (Tokyo) 88 295‑305.
'
e Tanaka S., Takikawa H., 1984. Amphibian and reptilian gonadotropin: biological activity.
Gunma Slymp. on Eleidocrinol. 21, 37‑61.
e Tanaka S., Sakai M., Hattori M., Kikuyama S., Wakabayashi K., Hanaoka Y., 2004. Effect of bullfrog LH and FSH on newt testes under diflierent temperatures. Gen. Comp.
EhdocrinoL 138, 1‑7.
e Tung PS, Fritz IB. 1980. Interactions of Sertoli cells with myoid cells in vitro. BioL Reprod. 23, 207 ‑217.
e Yazawa T., Yamamoto T., Nakayama Y., Hamada S., Abe S‑I., 2000a. Conversion from mitosis to meiosis: Morphology and expression of proliferating cell nuclear antigen (PCNA) and Dmcl during newt spermatogenesis. Dev. Growth DtOZir. 42, 603‑61 1 .
CHAPTER2
ldkintijication ofALRGI as a EgHLupregulated local.factor and eluciciation ofits,titnction
on the spermatogonialprolijZiration
enapter 2
2.1 INTRODUCTION
Spemiatogenesis is an essential process fbr sexual reproduction in development. It is uiggered by the sequential mitotic divisions of spermatogonia, fbllowed by their diffbrentiation into spermatocytes. After two meiotic divisions, spermatocytes develop into spemiatids, which possess halfthe norrnal complement ofgenetic material, and then into spermatozoa, mature male gametes in many sexually reproducing organisms. This complex process is controlled by cooperation with several hormones and testicular somatic cells. Follicle‑stimulating hormone (FSH) is one of the most important homiones required for early gonadal development, maturity and function (Sairam and Krishnamunhy, 2001). It is secreted by the pituitary gland and act on testicular somatic cells (Means et al., 1980; Ritzen et al., 1981; Hodgson et al., 1983; (lriswold et al., 1988), mainly Sertoli cells, through the specific receptor. Sertoli cells have essential roles in the regulation of spermatogenesis. They not only represent the only cellular component of the blood‑testis banier but also produce and secrete local factors to germ cells (Dym et. al., 1997).
In mammalian testis, Sertoli cells have been shown to synthesize stem cell factor (SCF) (Motro et al., 1991; Tajima et al., 1991), which plays a pivotal role in survival (Packer et al., 1995; Yan et al., 2000), proliferation and differentiation (Yoshinaga et al., 1991; Sette et al., 2000) of germ cells expressing the receptor c‑kit (Manova et al., 1990; Sorrentino et al., 1991).
insulin‑like growth factor‑I aGF‑I) is another factor shown to act specifically on spermatogonia and primary spermatocytes ofrainbow trout (Loir and Le Gac, 1994). The mRNAs fbr IGIT‑I and its receptors are present to a greater extent in Sertoli cell‑enriched populations and in those containing spermatogonia and primary spermatocytes (Le Gac et al., 1996). These findings may define SCF and IGF‑I as paracrine factors derived from Sertoli cells in the regulation of spermatogenesls.
in newt testis, FSH alone is suflicient to stimulate spermatogonial proliferation and differentiation into primary spermatocytes through the receptor expressed on Sertoli cells in both organ and reaggregate cultures (Abe and Ji, 1994; Abe, 2004), but not in the culture of spermatogonia alone ato and Abe, 1999). In addition, SCF and IGF‑I, both of which are upregulated in response to FSH (unpublished; Yamamoto et al., 2001), induce spermatogonial
CZeapter 2
proliferation (Abe et al., 2002) and differentiation (]Nakayama et al., 1999), respectively, in the organ culture. Recently, it was demonstrated that the blood‑testis barrier fbrrned by Sertoli cells shows size selectivity, which allows small molecules (‑500 Da) to penetrate into the cyst, but not larger ones (>1.9 kDa) (Jin et al., 2008). Therefore, SCF and IGF‑I induce spermatogonial proliferation and differentiation indirectly in organ culture. Though it is thus considered that the FSH effects are necessary to be mediated by local paracrine factors, which are secreted from Sertoli cells and act directly on spermatogonia within the testis, such factors have not been identified yet.
Because of the complex anatomical organization of the germinal compartment in mammalian testis (the Sertoli cells are normally associated with fbur or five gerrn cell generations), it is difllcult to examine the stage‑specific effects of the regulatory factors in the organ culture (Yamamoto et al., 2001). in contrasg clonally derived germ cells are enclosed in a cysg the smallest unit of the testis, and surrounded by Sertoli cells within the lobules in newt testis (Callard 1991; Abe, 2004). Therefbre, newt testis is an apprppriate model to isolate germ cells at a specific stage and study the efli2ct of various factors on specific stage of spermatogenesls.
To identify FSH‑upregulated local factors exerted directly in spermatogonial proliferation and differentiation, an EST (Expression Sequence Tag) library containing 5321 clones was prepared from newt testes and differentially screened by microarray analysis. Neuregulin 1, termed as newt neuregulin 1 (nNRGI), was identified as one of the novel FSH‑upregulated clones. First, partial cDNAs encoding two different clones (newt immunoglobulin (nfg)‑type and newt cysteine‑rich domain (nCRD)‑type) were isolated and then their expressions were examined in spemiatogenic stages and testicular cell types by semi‑quantitative and quanthative RT‑PCR. Next, the effect of nNRGI on sperrnatogonial proliferation was tested with recombinant epidermal growth factor (EGF)‑like domain of nNRGI (nNRGI‑EGF) in organ culture and reaggregated culture with or without somatic cells. The data indicate that nNRGI plays a pivotal role in promoting spermatogonial proliferation by acting both directly and indirectly via somatic cells on spermatogonia in newt testis.
23