Bull. Fish. Lab. Kinki Univ., No.14, 231 - 307 (2014)
Improvement of larval rearing techniques in bagrid catfish Mystus nemurus
Bagrid catfish Mystus nemurus
t�:tatt
01-fm!nJ1f;j1ffiO)cj(�
Sharifah Rahmah Binti Syed MuhammadChapterl Chapter2
Contents
General Introduction
Development of morphology, sensory organs and behavior of larval bagrid catfish
2.1 Introduction · · · · · 2.2 Materials and Methods · · · ·
2.2.1 Preparation of larvae • · · · · · 2.2.2 Experimental design
2.2.3 Statistical analysis
2.3 Results · · · · · · · 2.4 Discussion · · · • · · · · · ·
Chapter3 Manipulation of photoperiod and feeding schedule to improve survival and growth performances of larval bagrid catfish
3.1 Introduction · · · • · 3.2 Materials and Methods · · · • · •
3.2.1 Preparation of larvae 3.2.2 Experimental design 3.2.3 Statistical analysis 3.3 Results · · · · · 3.4 Discussion · · · • · · · · · ·
. .
. .
. .
. .
.
.
.
.
. . .
. .
.
.
.
.
. .
.
. .
. .
.
. .
.
.
. . .
.
232 235 235 240 240 241 242 243 253 258 258 261 261 262 264 265 270Chapter4 Manipulation of stocking density and shelter to · · · · · · · · · · · 275 improve survival and growth performances of
larval bagrid catfish
4.1 Introduction · · · · · · · 4.2 Materials and Methods · · · ·
4.2.1 Preparation of larvae 4.2.2 Experimental design 4.2.3 Statistical analysis 4.3 Results · · · · · 4.4 Discussion · · · · • · · • · · · · · · · · · · Summary · · • • • · · · · · · · · · · · • • • · · · Acknowledgment · · · · · · · · · · · · · · · · · · References · · • · · · · · · · · · • • · · · · · · · · · 231 -275 277 277 277 279 280 284 289 294 296
近大水研報
Bagrid catflsh 入lyst1ιS 11e111Ur1ιS (synonym He111ihasr1ιS 11e111Urus) is categorized into the family ofBagridae.1t is known loca11y as 'baung', tropical cat丘Sh (Khan et al.1996), green Cat丘Sh (A1110rnsakun et al.1997) or bagrid cat丘Sh (Ng et al.2000} Abidin et al.2006).1tis an Asian 丘eshwater catfish inhabiting the dvers, estuaries,1akes and upstream ofreseNoirs at a depth ranging 丘om 2 t040 m. ThiS 丘Sh is frequently found in large muddy rivers, with slow Cunent and s0丑 bottom (Kottelat et al.1993).1t natura11y spawns twice a year 丘om Nlay to July and 丘om september to November (Muchlisin et al.2004) in aooded forest (Rainboth 1996). Ba8rid catflsh can attain a total body length of 65 Cm (Baird et al.1999) but are more Common between 25 and 35 Cm e、11esomya et al.2002).1t can be classified as omnivorous Species but have the tendency to feed like carnivorous-scavengers (Alnornsakun et al.1998b) Bagrid catfish natura11y feed on C川Stacean (crabs, prawns), aquatic and tenestrialinsects, 丘Shes, detritus and vegetation (A1110rnsakun et al.1998C)
1 4 万' Chapter l ( 20 14 ) Genera11ntroduction H^ .{ !3 L、 町 1、_
The remarkable progress in the commercial culture ofbagrid catfish in southeast Asia is attributed ftom its exce11ent taste and non-bony st川Cture (Alnomsakun et al.1998a) Bagrid catfish were reported to contain higher vitamin E and omega 3 fatty acids especia11y eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) compared to sardine, a marine 丘Sh (Nlesomya et al.2002). This proves exce11ent aesh quality despite being a freshwater fish. Typicalfteshwater flsh contains higher omega 6 Compared to omega 3 fatty acids. on the other hand, marine fish typica11y contains higher omega 3 Compared to omega 6 fatty acids (sargent et al.2002). F0110wing its exceptionaltaste and aesh quality, bagrid Catfish has been a popular fish for aquaculture in southeast Asia and should be introduced as a global commodity in the 6.1ture. Atthis moment, its market price is about usD8 Which iS 3 t05 times higher than other commercial catfishes such as A丘ican catfish C1αrias garieP加1ιS and river catfish Pαπgasiα110d0π hypophthalnlus especia11y in Malaysia,1ndonesia and Thailand
In Malaysia and lndonesia, bagrid catfish is cultured in ponds, pens and aoating cages (Kama川din et al.20ID and usua11y marketed ftesh (Rainboth 1996). The growing interest in the commercial culture ofbagrid catfish has encouraged various studies on the nutrition and husbandw (Khan et al.1993; 1996; k,,omsakon et al.1996; 1997; 1998a,b,C; Ng et al.200の, breeding (1halathiah et al.1988; Muchlisin et al.2004; 2009), genetics and population
(chong et al.2000; Leesa、Nga et al.2000; usmani et al.2003). The production of bagrid
Catfish in Malaysia started in 1993 With only 29 tonnes.1t increased gradua11y over the years With the highest production of l,366 tonnes in 2003 before decreasing t0915 tomles in 2009(FA02010). The reduced production is mainly due to the inconsistent supply ofwild seeds
(Tantikitti and chimsung 2001) resulted f卜om overflshing and environmental deterioration
Iuthough adi負Cia11y produced seeds are available,it sufferS 丘om low larvalsuNival(Abidin
et al.2006). However, atthis moment,the main cause oflow larvalsuNivalremains unclearkifahRa1血ahBintisyedMuha如nad : Bagridcat負Shuyst加"e形UruS における仔魚飼育技術の改善
Therefore, the principle objectives ofthis study were to predictthe cause oflow larval SUNivalin chapter 2. The resultS 負'om chapter 2 Were used to improve laNal survival of bagrid catfish by providing suitable rearing conditions in chapter 3 and 4
AtifahRa1如ahBintisyedMuhan血ad : Bagridcat丘Shuyst細πe柳U地S における仔魚飼育技術の改善
Chapter 2
Development ofmorph010套y,sensory organs and behavi0Ⅱr oflaNalba晋rid catnsh
2.1 Introduction
The unsuccesS6.111arval rearing in captivity is partly due to the insufficient hlowledge Ofthe early life history and its ec010gical adaptation (Kawamura and Munekiy01989). Newly hatched laNae are fragile and immature.1n order to ensure their early suNival during the developmental stages, the ability for food detection and predator avoidance is crucial. This highly relates to the development of sensory organs (Blaxter 1986). previous studies on the development ofsensory organs focused on the eye for vision, taste bud for chemoreception of taste, olfactory organ for chemoreception ofodour,丘ee neuromast for mechanoreception and inner ear for balance and sound detection (Kawamura and lshida 1985; Kawamura et al
1989; 2003; senoo et al.1994; uyan et al.2006; Mukai et al.2008; 201の. These sensory Organs respond towards the environmental stimulation which is then exhibited in their behaviour.、Nith the hl0叉Vledge on its behavioural changes t11roughout the larval stages, Possible cause of low laNal suNival could be predicted in establishing suitable rearing teclmiques according to the environmental prefヒrences of the larvae (Blaxter 1986; Kawamuraand Munekiy01989)
Ihe primary 6.1nction offish eye is to perceive light for vision in feeding and predator avoidance (Nlatsuoka 20OD. The retina of the eye is a photoreceptor consists of pigment epithelium which controlsthe amount oflight received (Masuda et al.2003). The eye ofmost newly hatched larvae are non・pigmented and non・6、1nctional but gadua11y formed by flrst feeding (Blaxter 1986), indicating one ofthe last step to shoW 6.1nctionality (osman et al 2008). The retina typica11y consists of single cones, twin cones (formed by two single cones)
and rods. The cones are involved in c010ur vision and visual acuity while the twin cones Which is irregular in fish and rodS 丘.1nction in dim light condition (Kawamura and lamura
1973; Nlatsuoka 200り. single cones without rods are usua11y first to develop by the 負rst feeding in the early lawal stages ofmarble goby 0り,ele0かjs "1αrnlorams (senoo et al.1994), Sutchi catfish Pαπgasiαπod0元 hypophthalnlus (Mukai et al.2010), Aftican catfish C1αrias gariepiπUS (Mukai et al.2008), red seabream pagrus "1ql;or (Kawamura et al.1984) and japanese aounder parahcthys oliναCeus (Kawamura and lshida 1985), hence requires the Presence of light for vision. Twin cones and rods usua11y appear in later stages either immediately before or dudng metamorphosis (Nlatsuoka 20OD which increases the Sensitivity ofthe retina (senoo et al.1994). Development of twin cones and rods may also indicate the shi介 of habitat from pelagic to benthic environment (Kawamura et al.1984; Kawamura et al.1989).、Nith the eye development, suitable light condition may be provided to enhance the feeding efficiency and predator avoidance especiaⅡy during their early life
Stages
近大水研報 14号 ( 20 14 )
The primary sensory organ of gustatory system is the taste buds. Taste bud is a Chemoreceptor which is impodant in detecting and identifying different wpes of chemical Compounds(Reutrer and 叉Nitt 2004) as close・range receptors in mostfishes and vertebrates.1t is distnbuted on various parts of fish body and its density depends on the species and the environment they inhabit (Fishelson and Delarea 2004). Taste buds can be detected in the Oral cavity such as the tongue and lips, pharynx, giⅡS, in the skin near the mouth and along the body (Reutter and 、N'itt 1993; Grazia and Giacom020OD.1n addition to catfishes, taste buds primarily seNe as long・ran今e receptors in directing them to food (ova11e and shinn
197フ). singh and Kapoor (1967) emphasized the intense sensory characteristic oftaste buds Which are abundant at the distaltip ofthe barbelin contact W託h food. However, taste buds Were not present on the onset offirst feeding in marine species (Kawamura and lshida 1985)
and several fteshwater species (senoo et al.1994).1n fish such as largemouth bass ハ1icropter1ιS Sα1万10ides, Nile tilapia oreochro"1iS πilotiC1ιS and bluefin tuna 7h1ιnπ1ιS
Orieπtalis, the first taste bud appear several days a丑er the onset of feeding (Ka、Namura and XNashiyama 1989; Kawamura et al.2003). This indicates the insignificant importance of gustatory at least during their early laNal stage.\N'ith regards to their high sensitivity for 10calizing and di貸erentiating food, development of taste buds is important in succesS丘.11 feeding especia11y with adificial diets (sargent et al.2002)
The olfactory organ als06.1nctions as a chemoreceptor for odour with the ability to recognize specific odour molecules in the air or in the water (valentincic and capri0 1994) This includes, alarm pheromones as defensive behaviour to danger, amino acids as feeding Cues, bile acids as non-reproductive social cues for migration and sex steroids and Prostaglandins as sex pheromones. olfactory organ with receptor ce11S begins to develop at the early embryonic stages (zeiske et al.2003). At the time of hatching, a11 Ce11types in the Olfactory epitheHum are present (Kasumyan 20ID.1n the early laNal stage, the olfactory epithelium develops along the basal surface of the olfactory cups with only a few sensory epithelial ce11S and the olfactory neNe is unclear. The epithelium wi11then differentiate where it is composed of sensory ce11S, supporting ce11S and ci11iated ce11S. The olfactory nerve is detectable at the base ofthe epithelium that extends to the brain where it develops gradua11y as the laNae age. As the laNae develop, the olfactory organ wi11increase in size with increasing number and size ofthe olfactory folds. This wiⅡ increase the number of receptor Ce11S 工Nhich increase the sensitivity ofthe chemoreceptor. The sensitivity development ofthe Olfactory may vary in species. since aⅡ Ce11types ofthe olfactory epithelium are present even during embryonic development, newly hatched laNae already display non・specialized response towards olfactory stimulation even before the 丘lnctionality of taste system (taste buds). soon a丑er the initiation ofexogenous feeding, sensitivity for defensive behaviour and
ArifahRa1ⅡnahBintisyedMuha1血ad : Bagridcat負Shuyst加πe抗川那における仔魚飼育技術の改善
food detection started to develop with the ability to differentiate similar odour (Kasumyan 20ID.1n most fish, the sensitivity of odour for food detection develops later that the defensive behaviour as obselved in sole solea solea (Knutsen 1992) and brown trout salnlo かUita (入learnS 1986). The sensitivity for food detection of odour depends on fbod searching methods. common carp cypriπUs calpio, a bottom feeder develops earlier olfactory Sensitivity during the larval stage compared to zebra丘Sh Dα11io reho, a visual pelagic-Plankton feeder(Kasumyan 20ID atthe end ofthe laNalstage
Free neuromasts also act as chemoreceptors as weⅡ as mechanoreceptors,(Kawamura and lshida 1985).1tfbrms the lateralsystem found on the head, around the eyes, between the nostrils, on the opercular and occipital regions, on the lowerjaw (Higgs and Fuiman 1998), tωnk (Mukai and Kobayashi1994; 1995) and the tail ofthe 丘Sh (seooo et al.1994). The Iateral system is related to prey detection (Janssen et al.1999), predator avoidance (Blaxter
1996), rheotaxis (Nlontgomery et al.1997) and intraspecific interaction (patridge and pitcher 1980).1t consists of sensory hair ce11S surrounded by supporting ce11S and a cupula, a je11y Iike rod protruding 丘om the body surface (Mukai and Kobayashi 1994). vibrations in the Water cause the cupula to move, therefore triggering the neNous impulses which enable the IaNae to detect nearby movements in the water. Distribution of 丘ee neuromasts may vary according to species. studies have been conducted on the relationshゆS between the distribution of 丘'ee neuromasts and their morphogenetic changes (入Iukai and Kobayashi 1991; 1995; Kawamura et al.2003). Free neuromastS 釘e distributed on the lower and upper Surface ofthe head for speciesthat search for food t11roughoutthe water column. on the other
hand,丘ee neuromasts are concentrated on the lower and lateral surfaces of the head for
Species that search for food on the bottom (入Iukai & Kobayashi 1994). The importance of 丘ee neuromasts indicatesthat excessive aeration, water cunent, vibration or other mechanical disturbances could lead to h喰h mortaⅡty
AtifahRa1血ahB血tisyedMuha1血ad : Bagridcat丘Sh地St細 11e"1UruS における仔魚会司育技術の改善
Inner ear provides balance and detection of sound in fish larvae (Mukai et al.2008)' 1t Consists of labyrinth which is divided into pars superior and pars inferior. pars superior Consist of three major canals and one sac・1ike st川Cture, the utriculus while the pars inferior Consists of two additional sac・1ike structures, the sacculus and the lagena (popper and
CoombS 1980).1n the pars superior, semicircular canals (anterior vertical, horizontal and
Posterior vertical) prot川de fiom the labyrinth and each connects at both ends with the
Utdculus, a gravistatic organ. Each semicircular canal has a sensoly st川dure blown as crista
and provides the ability to maintain an upright position and to initiate intentional swimming
(osman et al.2008). The sacculus and lagena are primary organs for sound reception (popper
and coombS 1980). The sac-1ike structures, the utriculus, SUCculus and lagena, possess areasOf sensory epithelium, known as maculae which contain sensory hair ce11S. Each sac・1ike
St川Cture has its own ot01辻h known as the lapi11US, sagitta and asteriscus respectively. sagittae are the major receptors of sound in fish. Fishes hear 工入lhen sound waves cause the sensoryepithelium and the sa即ttae to vibrate. Ne工Nly hatched larvae have only two pairs of otoliths
(Bever and Fekete 2002) indicating undeveloped inner ear and the inability to maintain an
Upright position until the development oft11ree semicircular canals with rudimentary cristae
The anterior of the t、入10 pairs becomes the lapi11US (utdcular otolith) and the posterior pairbecome the sagittae (saccular otolith)(Fuiman 2004). The astedscus (1agena otolith) is the
Iast otolith to form (Bever and Fekete 2002). The auditory development is completely 員lnctional when the semicircular canals are 6.1Ⅱy ossi負ed (Kawamura and lshida 1985). The inner ear is able to detect vibrations in the absence of the 丘ee neuromasts (Kawamura and Washiyama 1989)XNith the development ofthe sensory organs and its specifiC 丘lnctions, fish are able to
respond towards the environmental stimulation which is closely related to the changes of
behavioural development (Leis and Brooke 2003). since the ability for food detection and
Predator avoidance are C川Cialto ensure the early survival of ash laNae, sensory organs
developments can also be used to correlate with the bahaviour to determine the time for the Onsets of first feeding and cannibalism. This is important to mitigate the causes of mortalitthe soonest possible during their early laNal stage for successful production.1n this regards,
the relationship among the development of morph010gy, sensory organs and behaviour ofbagrid cat負Sh were investigated to understand the behavioural changes in orderto predictthe
Cause oflow larvalsuNival
近大水研報 14方
2.2 2.2.1
(20 14)
Materials and Methods
Preparation oflaNae
Artificial reproduction was conducted in Aug 2009 at the center of c011aborative
Research in Aquaculture, universiti Malaysia sabah、Kinki university in Nlalaysia. Final
Oocyte maturation of50o g fヒmale brood fish was initiated with two intraperitonealinjectionsOfovaprim (0.5 ml kg、1, synde11ntemationa11nc., canada) at 6 h inteNal. Nlale received one
hormonalinjection (025 ml kg'1) at the same time as the second injection for f註male. At
Ovulation time (about 8 h a介erthe second injection at 28 - 29゜C), eggS 丘om female brood 丘Sh were stripped into a dry bowl by applying a ge址le pressure on the abdomen. Milt 丘om male were stdpped and inhaled into a vinyltube (inner and outer diameter; 4 and 6 mm, respectively) connected to a 60 ml plastic vial before mixing with the eggs to initiate
fedilization. Fe此ilized eggs were spread onto a net with the mesh opening of l mm and
Placed in a l,ooo lincubation tank until hatching. The tank was kept under natura11ight
Condition a2h light:12h dark) with temperature, dissolved oxygen and pH measured at 28 29゜C,5 - 6 mg r and 6 -フ, respectively・ 240 ・
2.2.2 Experimentaldesign
ArifahRahmahBintisyedMuha如nad : Bagddcat丘Shuyst加πe形WaS における仔魚飼育技術の改善
Newly hatched laNae were divided into two sets of experimentaltanks; set l with
tdplicates of non-recirculatory transparent aquariums of 5 1 and set 2 With one recirculatory
transparent aquadum of 801.、入later exchange was performed twice daily at 0630 and 1830
for tanks in set 2. No water change was perfonned for the tank 血 Set l and recirculato System was only initiated at 18 hours a丘er hatching (hAH) when the larvae were able tomaintain a 3・dimensional swimming position. Both sets of experimentaltanks were stocked
With 30 larvae l、1. The laNae were fed with rotifer (βrachi0元1ιSphcatilis sp. complex) at 30
hA11 f0110wed by 冱rte"1iajシα11CisCαπα nauplius during 2 t0 4 days a丑er hatching (dAID,
U0加α(4 t0 7 dAH) and an途Cial feed (otohime B2 and cl, Biokyowa, Japan) containing
approximately 50% protein and lo% 1ipid (7 t014 dAH) at 07:00,11:00,15:oo and 19:00
Tanks fbr set l were used to monitor daily suNival, mortality and cannibalism rates
Surviving and dead laNae were counted in each tank daily and any injured or disa eared
IaNae were considered to be cannibalized. SUNival(S), m0此ality (M) and cannibalism (C)
Were calculated in % as s =(total suNived larvae /initial number oflarvae)*100, M =(total
dead laNae /initial number oflarvae)*10o and c =(total number of injured and disappeared
Iarvae/initial number oflarvae)*100
Tank for set 2 Was used fbr sampling to measure totalbody length (TL), mouth height
(入侃), yolk sac volume (YSV), hist010gical analysis, morph010gical and behavioural
developments. Ten laNae were sampled and anaesthetized 工Vith Transmore containing
a-methyl quinoline as active ingredient (Nika Trading, Malaysia) at 6 h inteNals from o t03
dA11 and at l d inteNalS 丑'om 4 t0 14 dA11. These samples were used for obseNation of
morph010gy conducted under a light microscope (Model EC1ゆSe E600, Nikon, Japan) and
measurements of TL, NⅡ'1 and Ysv were taken 丘'om digitalimage using lmage J ima e
Processing s0丑Ware (Rasband 1997-2008). Mouth height was calculated aS入層ji丁〒fjf、,
Where uJL = upperjaw length, LJL= 10werjaw length (Guma'a 1978). Yolk sac volume was
Calculated aS 4Bπ*(rl/2)2*r2/2, where rl
mlnor axls, r2 major axis (Fukuhara 198の Additiona1 306Sh were sampled, anaesthetized and fixed in Bouin's solution for hist010gical analyses ofthe eye, taste bud, olfactory organ, fiee neuromast and inner ear developments Haematoxylin、eosin staining of 6 μm cross and sagittal sections were obseNed under li合ht microscope (Nlodel EC1ゆSe E600, Nikon, Japan). The presence ofretinaltapetum in the eye Of bagrid catfish laNae was determined by i11Uminating the hist010gical section prepared earlier only with surface light 化S・L11A, Futaba) and obseNed under the light microscope This was conducted since the eyes ofadult bagrid catfish possess a retinaltapetum that shine Under a aash light at night indicating nocturnal characteristicsChanges in feeding and swimming behaviour were monitored 丘om the outside ofthe transparent readng tank at every feeding time for lo min.1mmediate phototaxis, rheotaxis and thigmotaxis responses were also observed on 50 laNae placed in a l,ooo ml beaker at every sampHng time (6 h intervalS 丘'om o t03 dAf王 and at l d inteNalS 丘'om 4 t0 14 dA11) Phototaxis behaviour was determined by obseNing the swimming response when an LED Iight was aashed at one side ofthe beaker. Rheotaxis behaviour was detennined by obseNing the swimming response to a weak current created by a glass pipette. Thigmotaxis behaviour 工Nas assessed by obseNing the swimming response towards a mechanicalstimulus by a glass
Plpette
近大水研報 1 4 万' ( 20 1 4 )
Statistical analysis 2.2.3
A11 data were statistica11y analyzed using spss version 11.0. Differences in mean Values were analyzed using one・way analysis of variance (ANOVA) f0110wed by post HOC Tukey'S HSD test u)<0.05). The normality ofvariance was analyzed using shapiro・xvilk test
'
゛底'一¥ー
ー゛'6 .ー︼ーー
2.3
Alifah Ra1血ah B血ti syed Muh如血ad
ReSⅡlts
Only prominent developments were presented. changes ofsuNival, m。rtl't d
Cannibalism
Were presented in Fi8.2.1 While TL, NⅡ・1, Ysv and m。r h。1。 1
Characteristicsofbagridcat6ShlaNaewerepresentedinFi s.2.2_ 2.5, d l
Ba師dcat丘Shuyst紹"e"1U地Sにおける仔魚飼育技術の改善
bagrid cat丘Sh 丘om o t0 14 一→'-survival ー.-Modality ^Cannibalism 75 70 0
Fig・ 2.1 Changes in suNival, mortality and cannibalism oflaNal
dA112 4 5 6 7 8 9 10
Days a丑er hatcMng (dAH)
・ 243 、 Ⅱ 12 B 14 、ER),日め=合一婁δで姦ご=昏0Σ (ゞ)一今一Ξめ ア3
0505009988
332211SO
505050
30 25 近大水研報 20 15 14号 10 5 ( 20 14 ) 0 0 0 ゛
Fi套.2.2 Changesin totalbody length oflaNalbagrid cat丘Sh 丘om o t014 dAH
2 0 口 3 口 4 重 5 6 7 8 9 10 Days a丑er hatcMng (dAH),゛
1-'ー^一π Ⅱ 12 B CP 口 14ぜP
臣 口 口 口 臣 0 0 口Fi套.2.3 Changesin mouth height oflaNalbagrid cat丘Sh 丘'om o t014 dA11
0 3 口 4 「一ーーーー^ーーーー了^T-ーー 口 5 6 7 8 9 10
Days a丑er hatcMng (dAH)
口 口 Ⅱ 12 B 14 (安Ξ)二晶忌]一巴0-i d (巨)三ぽ=占言Σ ﹁2 6 4 2 0 8 6 4 2 0
ーーーー00000
AtifahRa1血ahBintisyedM血a如nad
・ 245 、
Bagridcat丘Sh地St加πe形討地S における仔魚飼育技術の改善
0 6
゛
Fi曹・ 2・4 Changesin yolk sac volume oflarvalbagrid cat6Sh 丘om o t07211A11
12 18 24 30 36 42 48
Hoursa丑er hatcMng (hAH)
゛゛,
54 60 66 口 72 (MΞΞ)省三0>詩如"-0>42086420ーーー00000
a 近大水研報 b 1」 勺 1 卜↓ C 14号 ^ ( 20 14 ) d ^ h e ^ @ 1」 f ^ 1 ゆ g ) ^ @ 'j. φ ^ L 、.._ 1 」 k 嫉.邑 ^ '^
Fi套.2.5 Changes of m0印h010gical charactedstics of laNal bagrid catfish' a, o hAH; b,6
hAH; C,12 hA11; d,18 hA11; e,2411AH; f,3011AH; g,3611A11; h,5411A11; i,72 hA11;j,5
dAH; k,10 dA11;1, B dA11. Bar,1 mmノ ^ Φ 『、『_' ノ、巧之1う 、 'ニイ '゛*J-LL .、ー ^ t、"^'
、、司^
」1 命 ^ gト\ 、.jl^又゛,→、 \呉ト、ミミ ^ Z^__P-=セf 、ー:.'ニ、 』^ξΞ,ミ1J 、耳=』'、:コ之一、 T 、、"';、具、、、 ^ := 、 { 二 厶 . 一ι 戸一 一,、 , ' T1゛ 一゛一、一一一一 ←、 一、 ÷、ニ ﹃、士 、^、一一.一三、气ミ三、 才、 エー、 .1 , 一f .. L 4 一' .J 卜上 .デ 一,一' 風、 t、
都 =討. 軽 サー.主 一 1 ,、、、 一1 堅、 三、{ 三 '=ニ、 一子一 ユ1 +よ ﹃ト、 ' 二 ,,b、一 J 〆 '. 、=' こ 江、 一、 一睦一、 リ、一 一﹄ T 三之ト ユ: .一 '丁,一'
¥、L ﹁ .J 一 . 一"一一 ,' 、-i 一り,﹁工、, .一゛"一. =, 一.ー,.L﹂ ﹂ーー. 'T , ミ、, 一一一 一一、 "一' 一ノL 一升キ . .゛・Υ、 一一 1 , ..゛、 1、 ノ﹃、 h"In newly hatched larvae (mean士SD,4.61士0.09 mm in total body length,1L), pect。ral
丘n was unapparent. The digestive system was also undeveloped with unapparent mouth and
anus cavity as we11 as thin-wa11ed, narrow and straight intestinal tract. First pair of barbel
(maxi11ary) was apparent with adhesive characteristics at the tips. At this stage, sensory
Organs were undeveloped where the eye was unpigmented and the retinal ce11S were
Undifferentiated (F喰.2.6a).1n addition,1aNae had only sma11 nasal pit with cilia at its
Surface with no appearances oftaste bud and 丘ee neuromasts. The larvae also had onl oval_
Shaped otic vesicle with two otoliths, indicated as lapi11Us and sagi杜a while the
Sensoryepithelium ofthe inner ear was recognized as utricular and succular maculae (Fig.2.10a). At
this stage, the larvae lied at the b0札om of the ta11k either in upright, inverted or sidewa s
Position with 丘'eqU3nt movement of the tail. No phototaxis, rheotaxis or thi motaxis
responses were observed
At 6 hA壬1(4.76士0.07 mm), mouth gape and 川dimental pectoralfin were apparent
The intestinal wa11thickened with visible lumen. second pair of barbel (mandibular) was
Visible. The eye became slightly pigmented but only at a sma11 Pa此 of the outer layer (Fig
2・6b). Retinaltapetum (Fig.2.6C) and optic neNe (Fig.2.6d) was obseNed. At this stage, the
IaNae showed disoriented movement with twirling manner on yolk sac or sidewa s. some
IaNae showed negative phototaxis and swam away 丑'om the light source while aⅡ ofthem
had mild positive rheotaxisresponse towardsthe water cunent. No tMgmotaxisres onse was
ObseNed atthis stage
At 12 h細(5.03士0.10 mm),川dimentalhyP山al,third pair ofbarbel(mandibular) and
a pair ofnostril became apparent. Lowerjaw sh0工Ned slow movement and the intestinal waⅡ
thickened with increased lumen width. At this time, two pairs of 丘・ee neuromasts were
ObseNed at the anterior and postedor part towards the eye (Fig.2.9a). For the inner ear
development,the auditory vesicle enlarged and fbmled two prominent semicircular canals. At
ArifahRa1ⅡnahBintisyedMuha1血ad : Bagidcat6ShMst加11e1πUruS における仔魚飼育技術の改善
this stage, more larvae exhibited negative phototaxis response while stronger positive rheotaxis response was obseNed. AⅡ larvae showed mild negative thigmotaxis response When they were stimulated with the glass pipette
At 18 hAH (5.32士0.22 mm), ye110wish auid was obseNed along the intestinaltract The retina ofthe eye differentiated into pigmented epithelium, outer nudeus, outer plexiform, inner nucleus, inner plexiform and ganglion layer (Fig、 2.6e). At the same time, single cone Ce11S were also observed (Fig.2.60. sma11taste buds can also be seen on the lips and barbels Ofthe laNae. The protNsion ofthe semicircular canals ofthe inner ear waS 員.1rther divided into t11ree sections namely anterior vertical, horizontal and postedor vertical canals (Fig 2.10b).1n accordance to these developments, the laNae started to show aggregation behaviour by gathering atthe b0杜om darker corner ofthe tank. At 24 hAH (5.75士0.12 mm), 川dimental dorsalfln was apparent. N丘ld peristalsis was observed with discharged ye110wish auid which indicated the opening of the anus. At this sta8e, some of the larvae showed Vedicalswimming behaviour along the bottom comer waⅡ ofthetank
At 3011Afl(6.09士0.21 mm), thick folds ofintestinalwa11 With 丘equent peristalsis was Observed. The adhesive charactedstics at the tips of maxi11ary barbel disappeared. At the Same time, taste buds were observed inside the mouth and on gi11 arches, and their numbers On the lips and barbels increased (Figs.2.7a・d). The sensory epithelium ofthe olfactory organ thickened and had sunk deeper 丘om the nasal surface together with its cilia, forming deeper
Pit with laNal growth which began to fold at 30 IL、11(Fig.2.8a). At this stage, the laNae
Started to show foraging behaviour atthe bottom ofthe tank. Atthis time, rotifer was offered to the laNae. The average mouth height and absorbed yolk at this time waS 500 μm and65.4%, respectively. However, no food was detected in the digestive tract ofthe laNae attMS
近大水研報 14 万' ( 20 1 4 )
ArifahRahmahB血tisyedMuha11如ad : Bagtidcat丘Shuyst加11e形UルS におけるイ子魚飼育技術の改善
At 36 hA11(6.35士0.06 mm),川dimental analfin was apparent. Forthe olfactory organ, One lame11a with increased cilia was recognized and continued to fold to four lame11ae by 4 dA11. Rod ce11S were also identified at the pigmented epithelium ofthe retina (Fig.2.6g). At this stage, feeding activity was 0丑en observed atthe bottom ofthe tank with rotifer detected in the intestine. Atthe 負rstintake offood, the mouth height and absorbed yolk were 550 μm and 66.1%, respectively. At 481し、H (6.79士0.19 mm), a pair ofnasal barbelwas evident
At 54 hA11(6.90士0.22 mm), teeth started to develop and aexion of notochord was ObseNed. The utricular and saccular maculae of the inner ear were also differentiated (Fig 2.10C). Atthis stage,2% ofcannibalism was obseNed with biting the postedor body oftheir Conspecifics representing loo% ofthe total mortality. At 72 hA11(8.24士027 mm), rays of dorsal, anal and caudal fins started to develop. Yolk sac absorption waS 99.29る at this time and could not be visibly measured at 4 dAH onwards. However, from the hist010gical Sections, smaⅡ traces ofyolk can sti11 be obseNed at 4 dA王1' T11roughout daytime, the laNae aggregated at the bottom darker area of the tank when they were not feeding' The larvae exhibited active swimming and fbraging behaviour in the water column at night or when food Was provided. some ofthe larvae swam against water current 丘'om the waterinlet
At 5 dA11(11.38士0.76 mm), dorsal and anal fin started to separate 負'om the 丘n・fold and caudal fin started to fork. pars superior and pars inferior ofthe inner ear developed and are 6.1Ⅱy 6.1nctional by 5 dA1王(F喰.2.10d). As the laNae grow, the appearance of 丘ee neuromasts had increased especia11y on the head and tNnk ofthe larvae (Fig.2.9b). However, the exact type, numbers and distribution of the 負'ee neuromasts were not able to be determined t11rough the use ofonly hist010gicalsections in this study. At 6 dAH a2.01士0.69) mm, the nostdlfbrmed two pits and became recognizable as antedor nasal pit and posterior nasal P北(Fig' 2.8b). At the same time, seven lame11ae with increased cilia were observed in the nasal pit (Fig.2.8の. At B dA11(23.07士1.86 mm), adゆOse 丘n waS 血11y separated 丘om
the fin・fold and the lalvae metamorphosized into juvenile stage. At 14 dA11 (23.1士1.99), m0此ality and cannibalism waS 26 and 259/o respectively. XNith this,96% ofthe m0此ality in Iarval bagrid cat負Sh was due to calmibalism
近大水研報 撫.ミ叺 ゛十遍、、 ー^^ 此 げ ず゛, f "'・e '邑"、・'゛ニ 、、、・ 1, "モエ↓'" 禅、'、美・"、瀞 14号 イ ,]'1レ〒 e 1 (2014) 了イ 宇 a 、・ 1
.、゛、y、一硝、、畿声j_
'*ゞ・',・",,ニー髪"、カ、、
盆赳、ミ、'遭'畴,,、,'.゛逢、t傘'、い・、
、 1悼殉 J'1ξ一.ノ.气,釜;;_1、、
松?油キ4'、'1都売;ゞ'、゛,1i
戸〆'、"!y"ー"、、、、'、.1.",'
d '、ー:1.捷ご・ゞ'二f'富!蛭'ー;
、,、吋^一畔゛^^ C ユFi套.2.6 Cross sections of laNal bagrid catfish eye. a, o hA11 (baF50 μm); b・d,6 hA11 (ba炉50 μm); e・t 18 hA11(e, ba仟50 μ皿 t baFI0 μm); g,36 hA11(baF50 μm). gd, ganglion ce111ayer} inl, inner nucle雛 layer;ゆ1, inner plexiform layer; 1,1ens; n, nudeus of Visual ce1上 onl, outer nudear layer; OPI, outer plexiform layer; on, optic neNe; pe, pigmented epithelium; r, rod ce1上 rt, retinaltapetum; SC, single cone ce11
号 f e 十 キ壯 1 ゛ ^ r 凡 亀 "T 1一、 ﹃1 斗. . :ゞ,'舳f .、 箪 凱一 ン、ゞ,、、、'金 ニ,顧為 ^,談ご、,"、、佑篭゛ゞ岬 一 ,".挑拠 姦t-,一 、一亨一 'ゞ 宇 ,、、、.'一 斎小', .、 '.ー、 =、、、 b ﹁ 式導 、、ξ一・ 舮'キ". 、、 ^二L ゛ι tぞ、連: 琳亀、ξ、, 1 風唇生." J ,、,亀、.玉 .、 、鶴, ゛ 二、 ゛ ゛ξ、一二," 江゛'、.'ー" 一4 :、、ハ 、' 、ゞ 一、 M 〒、、 1 '窪亀瀞豊蒸
11
﹂Ξ '气 一 、 、.玉 J 气 J、 、轟 厶 、ぜ﹂ 、T .、 0ごAtifahRa1血ahB血tisyedMuhalmnad : Bagridcat丘Sh珍S加 11el?1UruS における仔魚飼育技術の改善 皇毛、
\
ノ遷 ^ a\
"喜\
\
' 、〒 、,怠 孤一、Fig.2.7 Cross sections of larval bagrid catfish taste buds at 3011A11. a, HPS; b, inside ofthe mouth; C, barbe11; d, gi11 arches. Arrow indicates taste buds. Bar,100 μm
C ^ 賦\ b ゛ 郁 ^、て 、 ひ 1゛ ・; 買郡L冒 7 0 . ,戸 、 、 等 d 1 ^ ' Jル 式 ゛'ケ十 C a Y 、 0 ', 、
Fig.2.8 Cross sections oflarval bagrid catfish olfactory organ dA11(baFI00 μm); C, ciliated lame11a at 6 dA11(bar=20 μm) Im,1ame11a; pn, posterior nasal pit; se, sensory epithelium
叩\
抑X長、
^ ιt:コ『 d皇 、、1.'J ゛=^玉島:T," 1 、 ' ^ , b i '"壷1?',ー"*、
'、^'誤 . 、゛毒、、 ^ C J ^a,30 hAH (baFI00 μm); b,6 an, anterior nasal pit; ci, cilia;
' 逢 争 ・ 251 警争 夢亮 k\ ' ざ' r二 島, 巨,
冥\一
、、" """キ 、倉 、、{
ー\'塵1
A
1 ﹂﹂ ゞ 俄 ;玉" ' ゛、 1. L-.、一一式配甲王 ゛.一一,し、 .ー. 怜. ' ゛ ノ戸拠 1、弓二.'亡 "李卦 1ノ ー、、 一゛ 、'、. ヨー、近大水研報
司、
凡J
14 万'
一
Fi琴.2.9 Cross sections oflaNal bagrid catfish 丘'ee neuromasts. a,121Lヘ、fl(baF50 μm); b,5 dA1王(bar=10 μm)(t川nk). Arrow indicateS 丘ee neuromasts
a ( 20 1 4 ) E '晤 ιモ' 5JI、,ゞ宝'1'.6 \、、 3 '、 コ門卑. ' キ 、 瘻 、 "゛1 畷 ・手、 、. ' ^ ^ 1 1 嘆鷲 3、、._、 、. , b 智'y ゛先 含. . '{ Ⅱnl . ^ 司一 la C ξ'U avc
Fi晉.2.10 sagi杜al (a) and cross (b・d) sedions of laNal bagrid catfish inner ear. a, o hAI{; b,
18 11AH; C,54 11A11; d,5 dA王1} as, asteriscus; avc, anterior vedical canal; hc, horizontal
Canal; 1a,1api11US; 1g,1agena; PVC, postedor vertical cana上 S, saccule; sa, sa部tta; sm, sacculus macula; U, utdcle; um, utdcular macula. Bars=50 μm
、、 、、 hC 〒 b hC Snl ,, ,、、、 S ι U PVC d ^ Sa^、 S Snl "ン 一『^乢盆乃^ 五 1g 、 、.^ aS ワ 、、、^ '一 工 '、 ゛ 4' 1一 、社 き謡、ゞ 、゛ ︹イ ﹃、/ Y 、气 て 、﹃气 "1 毒含ぎ ' 一り a S .... 舞 呉一} . , 1 4 茎 卓 ゛ ゛ 運 a 1 r r 当﹂ ゛ . 、 、、
武
﹃ι、、 ,武 ,、 " '輪゛ ﹂ι、 司亀 N ゛ 4 一一 C 出 噂 、i a2.4
ArifahRa111nahBintisyedMuha如nad : Bagridcat丘Shuyst伽 11e抗討ruS における仔魚飼育技術の改善
DisCⅡSsion
Genera11y, bagrid catflsh showed rapid development ofmorph010gy and 6.1nctionality Of sensory organs, simHar to other catfishes such as African catfish (1Vlukai et al.2008) and Sutchi catfish (Mukai et al.2010). These developments are strongly related to the changes of behaviour in order to improve the rearing techniques particularly in feeding capability during IaNalstages repoded in marble goby oxyeleob'is "1αr1πorams (senoo et al.1994)
In most laNae, vision is primarily important in feeding which depends on light availability obseNed in Japanese sardine sardiπOPS 111e1απOstictus (Nlatsuoka 20OD and red Porgy pagrus pagrus (Roo et al.1999). This is indicated by the appearance of eye Pigmentation, retinaltapetum, optic neNe and cone ce11S that was not obseNed in newly hatched larvae including bagrid cat丘Sh in the present study. However, rod ce11 Was also ObseNed in bagrid catfish laNae during the first feeding indicating nocturnal behaviour and the ability to see and fヒed in dim condition.1n most laNae, with some exceptions, rod ce11S appears later dudng metamorphosis for improved vision in dim light indicating a shi丑 ftom Pelagic to benthic behaviour (senoo et al.1994) or deeper water stratum (Fang et al.2004) These suggest that manゆUlation of light and dark condition or lower light intensity could improve the we11-being ofthe larvae, f0110wing its naturalnocturnalbehaviour
Taste buds in most fisheS 員.1nctions as dose・range receptor for fbod identification.1n Contrasts, taste buds in catfishes primarily serve as long・range receptors in directing them to food (ova11e and shinn 197フ). Taste buds on the barbels particularly show notable sensitivity Which develops early in larval stages ofcatflshes (ova11e and shinn 197フ; Mukai et al.2008) that were not present on the onset of first feeding in marine species (Kawamura and lshida
198S) and severa1 丘eshwater species (senoo et al.1994). This could explain the capability of bagrid cat負Sh laNae to feed on powdered artificial feed even during their first feeding and Under dark condition in our preliminary experiment. Although taste buds in bagrid cat6Sh
IaNae may contdbute in improved feeding capability in addition to visual sense, its remarkable sensitivity may pose an increased tendency ofearly cannibalism especia11y during nighttime as obseNed at 5411AI{ in the present study. Therefore, possible factors that might Iead to cannibalism such as fbod availability,1aNal density, shelter etc. should be considered in improving rearing teclmiques fbr higher suNival(Hecht and pienaar 1993)
Free neuromasts act as chemoreceptors and mechanoreceptors(Kawamura and lshida 1985). Functionality of 丘'ee neuromasts is indicated by the development of cupula (Kawamura and lshida 1985), evidence 丘om Pゆette avoidance ofbagrid catfish laNae at 12 hAH in this study.1Wai(1972) demonstrated that cupulae form in newly hatched larvae such as Japanese seabasS ιαte01αみrι裁jap0πicas, black seabream 豆Cα11thopagrus schlegeh, stone aounder καrei1ιS みic010rams, rockskゆPer Bleππius yatαみei, goby rhde元tiger ohscures, Chameleon goby Z かog0110cephalus and kusa員lgu Fug1ιπiph0みles.1n contrast, cupulae were ObseNed one or a fヒW days a丑er hatching in Japanese smelt Hyp01πesuS かαπδPaC加CUS πipp0πensis, big・scaled redfin rガh010d0π hak0πeπSis,丘eshwater minnow zacco P1αり,PUS, Common carp cypriπUs calpio, gold丘Sh carrassius auratus and japanese rice fish olyzias 1αtipes. studies have shown that neuromasts dependent laNal species showed lower feed intake when the 6.1nction of mechanoreceptors were chemica11y disabled or fed with 負'ozen Iive food. However, in the presence oflight, vision exceeds mechanical sense which enables feeding compared to dark condition (Mukai 2006). Although the 丘lnctionalimportance of 丘'ee neuromastS工刃ere not examined in this study, the ability to feed under dark condition may Iargely depend on taste buds or olfactory organs as also observed in sutchi catfish (Mukai et al.2010) and African cat丘Sh (Nlukai and Lim 20ID. Functional importance of 負'ee neuromasts in bagrid cat負Sh should be 会.1rther evaluated fbr better understanding of behaviouralresponsestowardsthe culture environment
Inner ear is associated with balancing abilities (Mukai et al.2008), sound and Vibration detection (Love11 et al.2007).1ncapability ofmaintainin83・dimensional S工刃'imming behaviour in newly hatched 1且Nae could also be due to the weipht of the volk qえC
ー^ーワ
Undeveloped pectoral fin and dyS且lnctional gas bladder as reported in characid fish salP1加1ιS 6rasilieπSis (santos and Godinh02002). Detection ofsound and vibration were more intense With the development ofotoliths where bagrid catfish larvae were easily sta此led by a sudden thump or movement. Tlis suggests minimal mechanical or physical disturbance to reduce Iarval stress
In newly hatched bagrid catfish laNae, adhesive charactedstiC 飢 the tips of the Pectoral barbel was observed unti1 24 hAH where it disappeared at 30 11AH. similar Characteristic was observed in A丘ican catfish larvae at the ventral part of the yolk sac.1n Catfish pseudohagl'us ishikqレル,aithe adhesiveness ofthe barbe1 丘_1nctions as a holding device On the substrate in the shelter for 2 t03 days a合er hatching (、刃atanabe 1994). Adhesive organ On the head of laNal characid fish 丘lnctioned as a holding device to the water surface in aoodplains during rainy season (santos and Godinh02002). similar breeding environmentin bagrid catfish may 血dicate a holding device ofitslaNae towardsthe bottom ofthe water bed This can be obseNed t11rough their benthic behaviour especia11y at the early laNal stage Which nlight suggest sinMng food, slower aeration and water cunentrate to be adopted in the Culture condition. Disappearance of the adhesiveness at 30 hAH conelated we11 With the Commencement offoraging and swimming behaviourin the工Vater column
RelationsMp ofmorph010今y, sensory organs and behaviour in this study indicated the Possibility of feeding capability, thus food was provided to the laNae at 3011AH. However, food were only obseNed in the intestinal tract six hours later suggesting that feeding Commencement requires acclimatization time (Kamler 1992). Nevertheless, the onset of丘rst feeding obseNed here was earlierthan that reported by A1110msakun et al.(1996) at 52 hA1王
AtifahRa1血ahBitltisyedMuha1血ad : Ba部idcat丘Shuyst加"e抗鉾地S における仔魚飼育技術の改善
The diaerence could be due to different developmental stages of the larvae resulted 丘om Variability of rearin8 temperatures; 28-29゜C (in the present study) and 2S-30゜C (Atnornsakun et al.1996), hence emphasized the importance of suitable temperature for Optimized growth. Different fbod sizes between the two studies may also be a {h,ctor despite the similar mouth size of the laNae (550 and 553 μm, respectively); rotifer a50-230 μm) Was provided in the present study compared t0 入10加α SP.(225 μm in width) in the latter Although shir0仇(1970) stated that optimalfbod size waS 50% ofthe larval mouth height, feeding behaviour and capability ofthe larvae towards the nature ofthe food should also be Considered fbr feeding success (senoo et al.1994)
The expression of cannibalistic behaviour in bagdd catfish was first obseNed at 54 hA1王 With chasing and biting ofthe postedor body part(but rarely consuming whole) which accounted for 96% ofthe total modality. This behaviour corresponded with the development Of teeth and increased mouth height. catmibalism in this study could also be due to the increased variation in laNal size with growth as reported in Japanese aounder parahchth),S OliναCeus (Dou et al.2000).1n addition, cannibalistic behaviour was 0介en obseNed at night as the appearance oftaste buds on barbels,1ips, inside ofthe mouth and 即11 arches increased its fbra今ing sensitivity. A1110rnsakun a998C) revealed higher入10加α SP.intake in 3 t015 dAI'1 bagrid catfish larvae durin今 the nighttime compared to the daytime but did not differ Significantly.1n A金'ican catfish, about 70% ofthe feed intake occurs at night when in excess Offood.XNhen they are exposed t024 h light condition, feed intake wasreduced by morethan 30%(Hossain et al.1999). These results indicated the existence of diel rhythm which could also be displayed in bagrid catfish lawae. However, common aquaculture practices 0介en Conduct feeding during daytime, thus leading to starvation during 伽e nighttime which might increase the dsk of cannibalism. Therefore, nigh犹ime feeding should be considered to improve the readng teC11tliques as also suggested in noctumal species (Mukai et al.200&
ArifahRa1血ahBintisyedMuha1血ad : Bagridcat丘Shuyst加11e形UruS における仔魚飼育技術の改善
2010).1herefore, suitable stocMng density, rearing conditions such as lighting, water turbidity, shelter as we11 as size sorting intervals should be evaluated to reduce the risk of 0牙""{、牙1{Qtn
^^ιι'ι鳳、ノ^"^ιι'
In conclusion, relationship among the development of morph010gy, sensory organs and behaviour could 6.1ndamenta11y be used to understand the m0印hogenesis and its Shown on the behaviour. This infbrmation can be used to provide the specific responses
requirement in establishing adificial rearing techniques for bagdd cat丘Sh laNae. From this experiment, itis suggested that bagrid catfish laNae should not be fヒd laterthan the onset of their flrst feeding at 36 hA11. soon a丘er the first feeding time, care6,11 Consideration should be focused on the cannibalism W11ich was the main cause of low larval suNivalin bagrid Cat丘Sh laNae
Manipulation ofphotoperiod and feeding schedⅡle to improve suNival and 晉rowth Performances oflaNalba晉rid catnsh
近大水研報
3.1 夏ntroducuon
In chapter 2, cafmibalism was one ofthe main factors causing low laNal suNivalin bagrid catfish which representS 96% of the total modality. cannibalism is defined as the activity of an atlimalthat consumes the aesh of its own species (Baras and Jobling 2002).1t OCCU犯 more intensely 血 fast growing laNae and eadyjuvenile stagesthan the adult because Of higher growth rate and feeding tendency in relation to its body size (Baras et al.1998) Tropical species showed higher cannibalism compared to temperate species (Baras et al 2000; puvanendran et al.2008) because it is tdggered by appetite and metabolism which is 員lrther enhanced by unsuitable rearing condition (Hecht and pienaar 1993; Baras et al.2000; 1998; Mukai et al.2008). This criteria correlates we11 With bagrid catfish larvae which is a fast growing tropical species attaining juvenile stage at 13 dA11 (28-29゜C) as reported in Chapter 2
Cannibalism that occurs in n飢Ure is 0丑en regarded as an ec010gical contr01 0r life Compensation in time offood scarcity to enhance the suNival ofthe 6ttest (Kawai1987).1n Contrast, occunence of cannibalism in captivity especia11y in intensive cultured 6Sh resulted in economica110st. since total suppression ofcannibalism in aquaculture is difficult to a杜ain, identi61ing and contr011ing the factors that might tdg号er cannibalism as early as possible is of greatimportance (Baras and Jobling 2002).1t was reported that cannibalistic fish may depend On cannibalism and di缶Cult to leam to accept artificial fbod (pa11er and LewiS 1987) Similarly obseNed in stdped basS入1'or011e saχαtilis (Braid and she11198D
14方
Chapter 3
Cannibalism was reported to initiate as soon as the feeding ability develops in dorada β1),C0元 1πoorei(Baras et al.2000) and catfish Pαπgasiαπod0π hypophthα1111US (Baras et al
ArifahRa1ⅡnahB血tisyed Muh町mlad : Bagtidcat丘Shuyst加 11e形UruS における仔魚負司育技術の改善
2010) at 21 and 3611AH respectivelv. R mav commence later in other sDecies such as African
Catfish C1αrias gariepjπUS (Mukai et al.2008), perch percajluviatilis (Baras et al.2003) and Pasific bluefin tuna 7hUππUS 0ガe11talis (sabate et al.2010) at 72 1Lヘ.11,10 釦d 14 dA11 respectively. cannibalism obseNed in bagrid catfish at 541Lヘ、H in chapter 2 Was considered to initiate early which was about 18 hours a丑er the first feeding. This could be related to the Unsuitable rearing condition f0110wing its noctumal behaviour of the laNae where they aggregate towardsthe darker area ofthe tank dudng the daytime, active swimming during the nigh杜ime, able to feed in dark condition and avoid light source. F0110W血g these, unsuitable rearing condition related to the photopedod and feeding schedule may tdgger cannibaⅡSm in bagrid catfish larvae and should be investigated
Light inauencesthe life cyde 6'om the embryonic stage (Downing and Litvak 2002) to the sexual maturation in teleosts (Nligaud et al.2010).1t is one of the main artiflcial 'zeit8ebers'(i.e. cue or synC11ronizer) that affects the diumal rhythm and behaviour of the 丘Sh. Receptivity and response of light in 丘Sh laNae is species speci丘C and changes with the developmental stages (Boeuf and Le Bai1 1999; puvanendran and Brown 2002). This includes changes in the retinal stNcture 負'om pure cones with high visual acuity to duplex retina (cones and rods) with high visual sensitivity (Boehlart 197& Kawamura et al.1984; Kawamura and lshida 1985)
Differe址 Photopedod (1ight phase) and scotopedod (dark phase) were reported to affect larval development, growth, survival, feed in仏ke, behaviour and metabolism of 負Sh differently (Hecht and pienaar 1993; Applebaum and Kamler 2000; Hossain et al.1998) Longer photopedod improves growth and suNivalin Atlantic cod Gadus "10rh記α (puvanendran and Brown 2002), snapper Pα81'us auratus (Fielder et al.2002), European sea
bass DiceπかαIchuS 1αhrιZX (vi11amizar et al.2009), catfish lPα11ago at記(Giri et al.2002), rainbow trout 0πCorh)πCh1ιS 1ア1yhss (sonmez et al.2009), red sea bream Pα8アUs hlql:or (Biswas et al.2006), Adantic halibut HTppoglosS1ιS hip,poglossus L.(simensen et al.2000) and tilapia oreochrohliS 元ilotiC1ιS (EI・sayed and Kawanna 2004). This is related to increasing feed intake as visual feeder at this stage. However, exposure t0 24 h light with 4096 more
近大水研報
feed consumption did not improve growth ofbarramundi ιαtes calcarif'er. This is associated With increased swimming and feed intake activity, thus increased energy consumption
1 4号
(Barl0工入l et al.199S). Longer scotopedod seems to improve the growth of noctumal species With re員.1ge seeking behaviour in light condition such as A丘'ican catfish (Applebaum and Kamler 2000; Hossain et al.1998), sutchi catflshPαπgasiα110doh h)pophthα1刀11ιS (Mukai et al 2010; Mukai 20ID and vuodu cat丘Sh Hetのみ,απCh"S I0π即y11is (Baras et al.1998). L喰ht Condition increased stress to photophobic species, thus reducing swimming activity and feed intake as we11 as displaying increased cannibalistic and terdtodal behaviour σ'1echt and Pienaar 1993). N入lhen designing an arti6Cia11ighting system for a given 6Sh species in culture, its padicular ec010gy and developmental sta今e which 工N'iⅡ affect its sensitivity should be Considered (vi11amizar et al.20ID
In accordance 工和ith the sensitivity ofHght, fish can be genera11y classified into diurnal
( 20 1 4 )
Species that is active and feeds dudng the day and noctumal species that is active and feeds during the night. There is also crepuscular species that is active and feeds during the twilight 丘'om dawn to dusk. Most nodumal species can be categorized into the crepuscular group (Blyth et al.1999). Di任erences in feeding period for diumal, noctumal and crepuscular Species may differ especia11y in the sensory ability where higher sensitivity was found 血 noctumal and crepuscular species due to insU錨Cient i11Uminance.1mproved feeding e伍Ciency based on their sensory ability permits better ingestion, digestion and assimilation (Biswas and Takeuchi 2003; Biswas et al.2010) because flsh can controlthe 企ed intake
ArifahRahmahBintisyedMuhan血ad : Bagddcatfishuyst弘S11e叛αruS におけるイ子魚飼育技術の改善
based on their energy demand and feeding rhythm (Hossain et al.1999; 20ob. Feeding time Was reported to affect feed intake and growth performances oflndian cat6Sh HeteroP元etιSies
五)ssilis (sundararai et al.1982) and channel catfish lctalur1ιS PU11Ctatus (Noeske et al.1985)
Catfish aeter0みrαπCh1ιS I0元宮lylhs fed during the nigh杜ime showed improved growth rate Compared to daytime with the same feedin8 rate 4くerdchuen and Legendre 199り. Nthough Asian seabasS ιαtes calcarιf'er were able to feed both dudng the daytime and nighttime, growth rate were insigniflcant with only daytime or 11ighttime feeding q王arpaz et al.2005) Nthough bagrid catfish is a noctumal fヒeder, they are usua11y fed during the daytime InsU伍Cient orinaccurate feeding time may lead to reduced feed intake and growth,increased Stawation that induce cannibalism and terdto"ality, thus reducing suNival (Hecht and Pienaar 1993; Hossain et al.20ob
Therefore, this study aims to investigate the manipulation ofphotopedod and feeding Schedule on the survival, cannibalisnL growth performances and swimming speed of laNal bagrid catfish to develop optima11aNiculture condition
3.2 MaterialsandMethods 3.2.1 PreparationoflaNae
Eggs were artiacia11y obtained in september 2011 at the center of c011aborative Research in Aquaculture, universiti Malaysia sabah・Kinki university in Nlalaysia using 2-3 year old broodfish of about 60o g each. Female received two intraperitoneal hormonal
injections ofovaprim (0.50 ml kg'1, synde1 1ntemationa11nc., canada) at 6 h inteNal while
male received one hormonalinjection (0.25 ml kg、1) atthe same time as the second injection
for female. At ovulation (8 hours a丑er the second injection at 28゜C), eggs and milt were gently stripped and mixed to initiate fe此iliZ飢ion. Fertilized eggs were spread onto a net 工刃ith the 磁esh opening of l m識 in a l,ooo lincubation tank until hatching. T11roughout the
experiment, the tank was kept under natura11ight condition with 12 h light and 12 h dark a2L:12D) at 1472 and B lx respectively ai8ht meter, EXπC 401025) while water temperature, dissolved oxygen and pH were measured twice daily at 28.3士1.9゜C,5.5士0.7 mg
1、1 and 7.0士0.2 respectively. Larvae hatched approximately 24 h a丘er fe此ilization were
transfened and acclimatized in an 80 ltransparent recirculated aquarium with a stocMngdensity of 30 laNae l、1 Prior to experiment. The recirculation system was only initiated at 18
hA1王 When the laNae were able to maintain a horizontalposition and aggregated atthe lower Corner ofthe tank (obseNed in chapter 2)近大水研報 14号 ( 2014 )
Experimentaldesi晉n 3.2.2
A two、factorial experiment was conducted to investigate the effects of t11ree Photopedods a11d t11ree feeding schedules on the suNival, cannibalisnL growth performances and swimming activity from 2 t0 14 dA11 1arvae (13 days). At 24 11A1王,1aNae were transfened int027 tanks of71 aquarium 丘Ⅱed with 5 1 dechlorinated 丘eshwater at a stocking
density of20 larvae l、1. At the same time, the mean initial weight (0.003士0.oo g) assumed to
be equalwas measured using pooled sample of loo laNae 負'om the acclimatization tank. This Was due to the difficulties in measuring individual weight of2411A111aNae since they were too sma11. A total of lo larvae were anesthetized with V20 diluted Transmore containing a・ methyl quinoline as active ingredient (Nika Trading, Malaysia) and measured for total body IengthThe experiment tanks represents t11ree different photopedods; D 24 h light (24ι),1D 12 h 1地ht:12 h dark a2L12D) and 1Ⅱ) 24 h d釘k (24D) condition and t血ee diff釘ent 企eding Schedules; D day and oight feeding (DNF),Ⅱ) day feeding (DF) and night feeding (NF). The tanks were 迅Uminated with auorescent light (40 \刃) positioned 20 cm above the tank. The Iight provided constant Hght intensiw of 3,Bo lx for the light phase and o lx fbr 仙e dark
ArifahRahlnahB血tisyedMuha1血ad ; Bagtidcat負Shuyst加11e形UruS における仔魚飼育技術の改善
Pbase (1i今ht meter, EXπC 401025). For 12L12D treatment,1ights were manua11y switched On and 0任at 07:oo and 19:oo respectively. Nlthe experimenttanks were covered with black doth respective to their photopedodsto maintain constantlightintensity. The black doth was Slightly uncovered just enough to conduct feeding and siphoning offaeces and uneaten food Where the tanks were exposed to externa11ightintensity of30 IX
First feeding for the laNae was provided with rotifer (Brachi0π1ιS phCαガ1is sp
Complex) at 3011AH (according to chapter 2) at 20 individuals ml、1. At 2 dA11, experiment
Was initiated by feeding the larvae with lrte1ア1iajl'a11CisCαπα nauplius (20 individual ml"1)
Unti1 6 dA11 While artificial feed (otohime B2 Containing approximately 50% protein and 10% 1ipid) was fed 負'om 2 t0 14 dA11to satiation' T11ree feeding schedules of4 h intervals Were as f0110WS: D DNF at 08:00,12:00,16:00,20:00,00:oo and 04:0011) DF at 08:00,12:00, 16:oo and ⅡD NF at 20:00,00:oo and 04:00. Totalweight ofan途Cial feed provided fbr each tank was measured atthe end ofthe experiment. Each combination between the photopedods and feeding schedules were done in tdplicates
During the experiment, dead laNae 6'om each tank were c0Ⅱected before each feeding time and obseNed immediately under the light microscope (Model EC1ゆSe E600, Nikon, Japan) at 4X magnification. A11y larvae with bitten marks on its body or missing dudng each Sampling time were characterized as cannibalism. At 6,10 a11d 14 dA11, ten randomly Sampled laNae were then anaesthetized with ν20 diluted Transmore to measure the individualtotalbody length. Atthe same time, SUNiving laNae 丘om each tank were counted
to measure the suNival. The anesthetized laNae were a110wed to revive and retumed to their
respective tanks
At the end of the experiment, a11 SUNiving laNae 丑'om each tank were pooled and Weighed. SUNival(S) and cannibalism (C) were calculated as: S (96)=価nal number of IaNae/initial number of larvae)*10o and c (9'、0)=(number of injured and disappeared
IaNae/initial number of larvae)*10o respectively. Growth performances; feed intake (FD, apparent feed conversion ratio (AFCR), specific growth rate (SGR) and coe伍Cient of Variation for total body length (CV) were determined using the formula: FI (3)= final feed
Weight (g)- initial feed weight(g); AFCR = feed 即Ve"(g)*weight 部in (g)"; SGR (%d')
(1n final weight - 1n initial weighり*days、1*100) and cv =(standard deviation*mean total
body length'1)*10o respectively. Apparent feed conversion ratio (AFCR) was used instead of
FCR in this study because uneaten feed were not measured due to the sma11 quantity and easily dissolved in the water. A11 resultS 圦lere presented for 14 dAH except fbr survival, Cannibalism and totalbody length which were presented for 6,10 and 14 dAHBehaviour observation was conducted using a high de丘nition inftared night vision Video camera (Yashica, ADV・5181'1D, Japan) positioned above the experimentaltank. TWO Sets of additional LED infrared light (56 LED per set) were assembled towards the sides of the video camera and a reaection tape was placed under the tank to improve the visibility of IaNal movement in dark condition. The peak radiation ofthe l"ED in丑'ared light waS 940111n Which was outside the range of absorption ofthe visual pigmentS 血 Catfishes (Lythgoe and Padddge 1989). video recordings were made fbr lo min at 08:00,12:00,16:00,20:00,00:00
and 04:oo at 14 dAH fbr a11treatments. swimming speed (cm sec、1) of lo sec was randomly
analyzed 丘om the video recordings which represents the swimming activity of the larvae (sabate et al.2010). video images were repeatedly played back with a personal computer Where their temporal changes in position were manua11y recorded on a transparent plastic Sheet placed onthe monitor近大水研報 14 万' ( 20 1 4 )
3.2.3 Statisticalanalysis
Nl da仇 Were statistica11y analyzed using spss version 11.0. The effects of Photopedod and feeding schedule and their interaction on suNival and growth pedormances
Were tested using two-way analysis ofvariance (ANOVA). Data with signi丘Cant differences Were 員lrther compared among the means using post HOC Tukey'S HSD test u)<0.05). The Swimming speed was analyzed usin旦 one・way analysis ofvariance (ANOVA). The normality Ofvariance was analyzed using shapiro・叉Vilk test
ArifahRa1血ahBintisyedMuha血lad : Ba即idcat負Shuyst加πe"1UruS における仔魚貧司育技術の改善
3.3
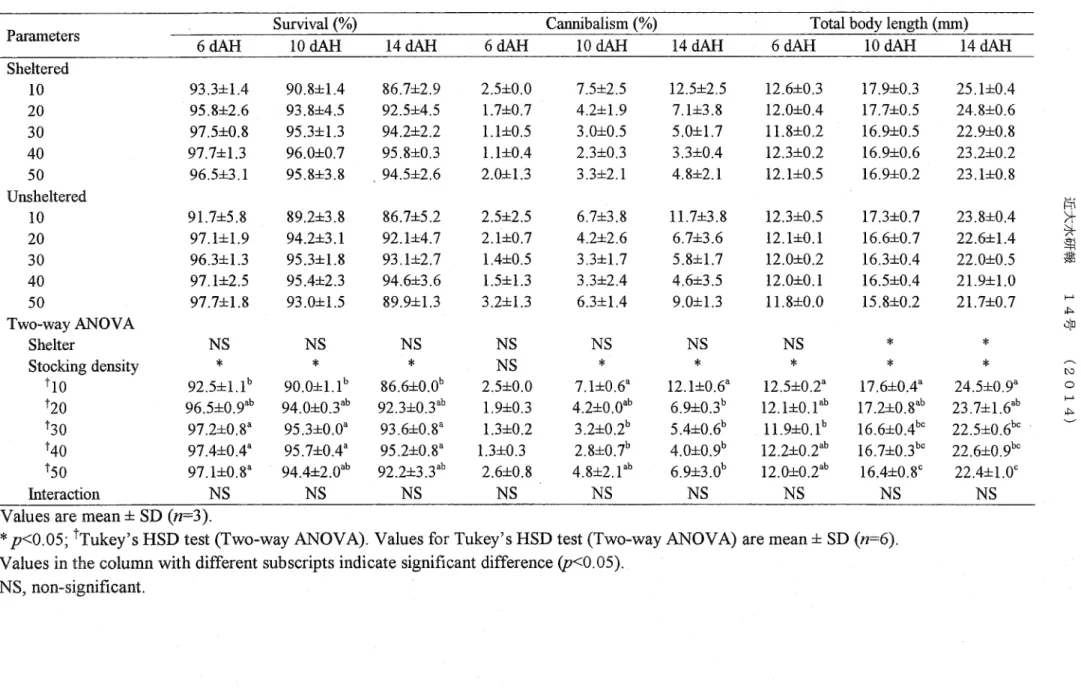
TWO・way ANOVA analysis indicated that photoperiod did not show any significant effects on a11 Parameters u)>0.05). Feeding schedule showed significant effects on the SUNival and total body length at 6,10 and 14 dA11 U)<0.049 and pく0.009; Pく0.033 and Pく0.000; Pく0.013 and pく0.ooo respectively). However, feeding schedule only showed
Significant effects on the cannibalism at lo u)<0.043) a11d 14 dAH U)<0.013). post Hoc test for two・way ANOVA indicated that for survival, day and night feeding (DNF) was Significandy 11igher than day feeding (DF) at 6,10 and 14 dAH. For cannibalism, DNF was Significantly lower than NF at lo and 14 dA11. For total body length, DNF was signiflcantly higherthan NF at 6 dA王l and DNF was signi丘Cantly higherthan both DF and NF for lo and
14 dAH (Table 3.D
TWO・way ANOVA analysis also indicated that feeding schedule showed significant effects on the feed intake u)<0.000), final weight Uフ<0.039) and coe缶Cient of variation q)<0.024) but showed no significant effects on the sGR and AFCR U)>0.05). post Hoc test for two・way ANOVA showed that for feed intake and final weight, DNF was significantly higher than DF and NF. For coe伍Cient of variation, NF was significantly higher than DF (Table 3.2). No signiflcant interaction u)>0.05) was obseNed between photopedod and feeding schedule in a11 Paramete犯(TableS 3.1 and 3.2)
Results
Iable 3.1 SUNival, cannibalism and totalbody length ofdifferent feeding schedules under different photoperiods at 6,10 and 14 dA11 Parameters 241ight DNF 121ight:12 dark DNF 6dAH 24 dark DNF 97.0士1.フ 93.3士2.9 96.0士1.フ Suwival(%) 10dAH 95.0士0.0 89.7士2.5 89.7士32 TWO・wayANOVA Photoperiod Feeding schedule TDNF ↑DF tNF Interaction 98.3士1.5 89.7士72 85.7士7.8 14dAH 93.7士0.6 86.0士3.0 86.7士2.5 94.7士4.0 86.3士7.8 83.3士8.5 93.7士7.1 86,7士8.1 92.7士3.2 6 dA11
Values are mean 士 SD (π=3) *Pく0.05
tpostHoctestfortwo-way ANOVA (1Ukey'S HSD)
Values in the column with different subscripts indicate signiticant diaerence u?<0.05) NS, non・significant; DNF, day and night feedin矧 DF, day feeding; NF, night feedin号Cannibalism (96) 1.0士1'0 4.3士1.5 2.3士1.5 94.0士3.0 83.7士7.0 81.3士8.3 93.0士7.2 85.0士7.8 90.7士4.9 NS 10dAH 96.3士2.4゜ 89.9士3.3b 91.4士5北 NS * 4.0士1.0 6.0士1.フ 8.0士2.6 1.0士1.0 8.0士6.1 8.3士4.0 883士5.1 82.7士7.2 88.7士4.9 NS 14 dA11 942士1.1゜ 87.0士2.4 87.9士4.0北 NS * 5.0士1.0 12.0士2.0 H.0士1.フ 2.3士1.5 10.7士7.6 B'7士8.1 4.7士5.0 63士5.9 4.3士1.5 NS 6dAH T0仏lbody length (mm) 92.0士3.2゜ 84.1士3.ob 85.6士3 8北 NS * 10.4士0.8 10.4士0.8 10.2士0.6 4.3士2.3 B.0士7.5 14.3士7.6 5.0士4.6 8.7士6.フ 8.0士4.4 NS NS 22士6.2' 6.2士1.9゜ S.0士3.1^ NS 10 dA11 17.0士0.2 15.2士0.1 14.7士0.4 11.3士0.2 102士0.1 10.0士0.0 8.7士3.5 13.7士5.8 9.7士4.フ NS 14dAH 3.8士1,4b 8.4士2.4釦 9.9士3.3' NS * 22.8士0.4 19.9士0.4 19.9士0.4 17.1士0.1 14.8士0.2 15.0士0.3 10.9士0.4 10.9土0.6 10.0士0.0 NS 6.0土2.4b 12.9士0.9゜ H.7士2.4゜ NS * 22.6士0.3 20.0士02 20.4士0.5 17.2士0,5 15.4士0.1 14.9士0.3 NS 10.9士0.ダ 10.5士0.4北 10.0士0.1b NS * 22.5士2.1 20.3士0.フ 20.5士0.6 NS 17.1士0.1゜ 15.1士0.3b 14.9士0.2b NS * NS 22.6士0.2' 20.0士0.2 20.3士0.3b NS * >十ラ十劃鳶 一心d (舶0一ど 班N 斑N DF N , NO、ー,
Iable 3.2 Feed intake, apparent feed conversion ratio, speci6C growth rate, final wei号ht and coe伍Cient ofvariation of different feeding Schedules under different photoperiods at 14 dA11
Parameters 24 Hght DNF 121ight:12 dark DNF 24 dark DNF Feed intake (幻 0.038士0.006 0.019士0.010 0.012土0.007 TWO・wayANOVA Photoperiod Feeding schedule fDNF ↑DF TNF Interaction 0.041士0.008 0.015士0.008 0.022士0.007 AFCR
Values are mean 士 SD (π=3) *Pく0.05
tpost Hoc test fortwo、way ANOVA (1Ukey'S HSD). values in the column with differentsubscrゆtsindicate signi丘Cant di丘erence u)<0.05)
NS, non、S喰nificant; DNF, day and night feeding; DF, day feedin島 NF, night feeding; AFCR apparent feed conversion ratio; SGR specific
growth rate 0.4士0.1 0.3士0.2 0.2士02 0.047士0.007 0.020士0.012 0.024士0.017 0.4士0.2 0.2土0.0 0.3士0.1 SGR ψ0) NS * 0.042士0.005゜ 0.018士0.003 0.019士0.006 NS 27.9士1.9 24.8士1.9 23.9士3.0 0.4士0.1 0.3士02 0.3士0.3 Final weight (3) 27.2士3.3 23.9士2.フ 24.7士3.2 NS NS 0.4士0.0゜ 0.3士0.1゜ 0.3士0.1' NS 0.H5士0.029 0.077士0.018 0.071士0.029 27.9士1.9 25.3士3.3 26.1士2.5 Coe缶Cient ofvariation (%) 0.109士0.041 0.070土0.024 0.079士0.029 NS NS 27.7士0.4゜ 24.7士0.フ' 24.9士1.1' NS 0.115士0.029 0.085士0.033 0.093士0.033 3.3士1.1 4.3士1.1 3.7士04 4.4士1.9 3.7士0.6 6.5士0.5 NS * 0.113士0.003゜ 0.077士0.008b 0.081士0.011 NS 3.9士2.1 3.3士0.8 5.8土0.9 NS 3.9士0.6北 3.8士0.5b 5.3士1.5" NS * と一響匁N冒'昌W巨一如てnル旨冨亘且一WN四式0昌跨ぷ貫ミミ言令雙一堅券団鴫鴬識S昇嚇
器
DF N 班N ' NO、司,Swimming speed representing the swimming activity of three different photoperiods (24L,12L:12D and 24D) manipulated undert1Ⅱee diffeNm feedmg schedules(DNF, DF and トJF) were presented in Fig.3.1 respectively. The white and black bar represents the natural Photoperiod ofdaytime (07:00-19:0の and nigh伽me a9:00-070の. The figures with grey and White area representsthe time thatthe larvae were fed and unfed respectively. For DNF (Fig 3.1a), there were 2 Significantly u)<0.05) higher peaks of activity at 12:oo and 20:oo for a11 Photoperiods. A110ther peak of higher activity at 04:0o could be identi丘ed fbr 24L and
12L:12D in DNF treatmentifthe activity for 24 h cyde showed similar continuoustrend. For DF (F喰.3.1b), adivity auctuated with a decrease ftom 08:oo t0 18:oo for aⅡ Photoperiods The activity slightly increased 丘om 18:oo t0 20:oo but decreased again until 04:00. The range of activity for daytime and nigh札ime irrespective of photopedod were 220-4.06 and
2.03-3.62 Cm sec、1 respectively. For night feeding (Fig.3.1C), activity increased for 24L and
12L:12D but decreased for 24D 丘om 08:oo t0 18:00. The activity slightly increased 丘om 1&oo t020:oo for a11 Photopedods. However,the activity 員lrther increased for 24L, remained Constant for 12L:12D and decreased for 24D. The range ofactivity for daytime and nighttime
irrespective ofphotopedod were l.32-3.42 and 3.14-4.65 Cm sec'1 respectively
ArifahRa1如ahBintisyedMuha1血ad : Bagridcat6Shuyst伽11e抗U地S における仔魚飼育技術の改善 a) 5 4 ^24L 3 2 →コー12L:12D Day and Night Feeding
b) 、 269 ・ 4 ^24D 3 b (24D) (24L) 2 ^ DayFeedi11g (24L) (12Lt12D) C) 4:00 Cd 4 b (12L桧D) bc (24D) 3 (24り 2 bc (24L) bc (24D) ー'ー'-1 (12L 12D)
Fi容.3.1 Swimming speed fbrt11ree photopedods; 24L,12L12D and 24D manipulated under three different feeding schedules a) day and night feeding (DNF), b) day feeding (DF) and c) night feeding (NF). Different subscripts within the same line indicate significant difference
ψく0'05, one、way ANOVA). white and black bar represents natural photopedod ofdaytime
and nighttime respectively. Grey area representsthe time thatthe laNae were fed0 Ni帥tFeed註lg &00 b(12L柁の (24り d (24の 12:00 12L:12D) d (24D) 1&00 20:00 Hours b 0:00 (一.oom 婁口)でご身⑳三ΞΞ冨め 0 5 0 5
3.4
Survivalwas significantly improved for DNF compared W北h DF but was insignificant With NF for 6,10 and 14 dA11. Lower survival for DF and NF (Table 3.D correlated with higher cannibalism at 14 dA11 due t010w feed intake similarly reported in African catfish (Hecht and pien肌r 1993; AI・Hafedh and Ali 2004), Japanese aounder paralichthys Ohvace1ιS (Dou et al.2000) and Atlantic cod Gadus "10rh解α(Folkvord 199D demonstrating that significantly higher feed intake in DNF in this study can reduce the cannibalism. The results proved that ba8rid catfish shows noctumal characteristics. However, feed should not Only be provided during the nighttime or only dudng the daytime as conducted in the Common aquaculture practice.1hisis because they showed higher activity orfヒeding demand both dudng the daytime and nighttime (Fig.3.D. From the results of swimmin号 Speed, although laNae were fed 6 times daily fbr DNF (Fig.3.1a), they stiⅡ Show increased activity at 12:00 (indicates daytime) and 20:00 (indicates nighttime).1n additiotL increased activity Was also obseNed during the daytime when the laNae were fed (grey area of Fig.3.1b), Where the range ofactivity was similarto the nighttime. This means that although the laNae Showed noctumal charactedsticS 工Vith increased activity at ni8ht (Fig.3.1C), provision offヒed during the daytime was not rejected by the laNae, hence increasing the activity. Feeding demand both during 血e daytime and nighttime was also reported by Alnomsakun et al a998の Where 入10加α intake in bagrid catfish lalvae was Mgh during the nighttime but was not signi丘Cant dudng the daytime. This could be related to the high metabolism and digestion Where previous study indicated that gastdc emptying time for 2・15 dA11ba8rid catfish larvae to be between 211r 30 min and 4 h lo min (Alnomsakun et al.1998b). Lalvae at 12 and 15 dA11 Showed less than twice faster gastdc emptying time compared to earlier life stages indicatin今 increased digestive ability (Atrlomsakun et al.1998b) therefore requiring higher
feed intake in shorter inteNals. This coincided weⅡ With increased cannibalism as the larvae
Discussion