博 士 学 位 論 文
Studies on development of the new method of control
using behavior regulators of Leptocorisa chinensis
(Dallas) (Hemiptera: Alydidae)
博 士 学 位 論 文
Studies on development of the new method of control
using behavior regulators of Leptocorisa chinensis
(Dallas) (Hemiptera: Alydidae)
(クモヘリカメムシの行動制御物質を使った新たな防除法の研究)
平成 29 年 5 月 17 日
Contents
Introduction 1
Chapter 1
Estimation of number of annual generations using effective heat unit of development for the rice bug, Leptocorisa chinensis (Dallas)
6
Chapter 2
Temperature and photoperiodic effects on induction and termination of diapause in female Leptocorisa chinensis
16
Chapter 3
A pecky rice-causing stink bug Leptocorisa chinensis escapes from volatiles emitted by excited conspecifics
31
Chapter 4
Evaluation of the behavior regulator of Leptocorisa chinensis to control this species in paddy fields
50
Summary 56
Acknowledgements 60
References 61
1
Introduction
While chemical pesticide use has increased agricultural production and
productivity, its use, overuse and misuse have caused negative externalities on human
health and the environment, as well as food safety (Kiritani 2000; Oida 2016). In
particular, the overuse of chemical pesticides has led to pest resistance, resurgence and
secondary outbreaks, which push farmers to use more new pesticides. To ease the
negative issues associated with pesticide use, integrated pest management (IPM)
technology, which aims to maximize farms’ economic profits, is introduced and
implemented in agricultural production worldwide. IPM refers to an ecologically-based
approach that makes the best use of all available technologies to sustainably manage
pest problems. The primary objective of IPM technology is to minimize chemical
pesticide use in relation to pest management, while maintaining or enhancing farms’ net
returns with minimal environmental degradation. Previous studies have shown that IPM
adoption significantly lowers pesticide use, saves production costs and maintains farm
productivity for adopters (Oida 2016). Thus, steps must be taken to further progress the
IPM program.
Peck (Fig. 1), primarily caused by stink bugs, is one of the key quality factors for
2
shriveled, and have poor milling quality. As the amount of pecky rice increases, the
quality and value of the crop are reduced. The adequate control of rice stink bugs during
heading can improve rice grade, quality and selling price, as kernels become discolored
when fed on by rice stink bugs in the milk or soft dough stages.
Fig. 1. Damage by stink bugs
The rice stink bug, Leptocorisa chinensis (Dallas) (Hemiptera: Alydidae) (Fig. 2),
is a major cosmetic pest. Leptocorisa chinensis is now recognized as one of the main
pests to cause pecky rice (Suzuki 2001). In Japan, this species occurs in mainland areas
(Takeuchi 2007), and is bivoltine throughout most of its distribution. During spring,
wild Gramineae species host L. chinensis, and the first generation of offspring
3
invades paddy fields and feeds on grains in the milk or soft dough stages, causing pecky
rice (Takeuchi et al. 2004b; Takeuchi 2007). While chemical insecticides are available,
the effective and environmentally benign control of this species in paddy fields is also
sought.
Fig. 2. Leptocorisa chinensis (Dallas) in life, on the top of Italian ryegrass, Lolium multiflorum Lam.
4
Pheromones (with Greek roots meaning ‘carrier of excitation’) are the chemicals
that an animal secretes or excretes that release a specific reaction, for example, a
definite behavior or developmental process’ in a member of the same species. Wilson
and Bossert (1963) divided pheromones into two groups: releasers, which induce an
immediate behavioral change including alarm, and primers, which initiate changes in
development, such as sexual maturation. The former does not result in immediate
behavioral changes, but means that the animal is predisposed to such changes (note here
that a releaser pheromone is not necessarily the same as a ‘releaser’ in the sense used by
ethologists such as Lorenz and Tinbergen to describe a trigger of instinctive behavior).
Knowledge that non-toxic and species-specific pheromones do not harm beneficial
species can be used to establish efficient and sustainable insect management strategies.
Disturbed stink bugs emit pungent volatile compounds that could have several
ecological functions including defense against predators or behavior regulation (Blum
1985; Aldrich 1995). A behavior regulator function might be used for the behavioral
control of stink bugs in agroecosystems. For example, in Sri Lanka, farmers collect and
squash stink bugs, placing them in bags around the field to reduce damage (Yamashita
personal communication). The volatiles from smashed bugs probably repel stink bugs in
5
used to repel conspecifics in rice fields and reduce the number of pecky rice grains. Leal
et al. (1996) identified the volatiles emitted from L. chinensis that were anaesthetized
with CO2 to minimize the release of defensive secretion, and found that a 5:1 mixture of
(E)-2-octenyl acetate and octanol, major components of the emissions of undisturbed L.
chinensis, was an attractant pheromone. However, the chemistry and ecological
functions of the volatiles from disturbed L. chinensis individuals have not yet been
studied.
The aim of the present study was to develop a new method of control using the
behavior regulator in L. chinensis. First, I examined the life history of L. chinensis to
predict and control L. chinensis invasion during the rice heading stage (Chapters 1 and
2). Second, I examined whether L. chinensis escaped from disturbed conspecifics in an
observation arena under laboratory conditions. Then, I analyzed the volatiles emitted by
both disturbed and undisturbed L. chinensis and observed the responses of undisturbed
L. chinensis to the components in the volatiles from disturbed conspecifics (Chapter 3).
Finally, I tried this control method using the behavior regulator of L. chinensis in paddy
fields and argued the significance of a new control method using the behavior regulator
6
Chapter 1
Estimation of the number of annual generations using effective heat unit of
development for the rice bug, Leptocorisa chinensis (Dallas)
Introduction
Understanding the life history of L. chinensis is an important prerequisite to
understanding the population dynamics of the pest in the field when using the behavior
regulator. With a detailed knowledge of the development of L. chinensis, it is possible to
predict the population fluctuations of this pest and then construct an effective control
program (Yao 2002). Leptocorisa chinensis invades paddy fields during heading, and
the end result is pecky rice is caused by L. chinensis (Takeuchi et al. 2004a). Thus, it is
important to predict and control L. chinensis invasions during the rice heading stage. In
particular, the thermal requirements (day-degrees) for development are often used to
estimate developmental periods because temperature has a major influence on the rate at
which insects develop (Howe 1967; Zaslavski 1988; Gordon 1998). However, little is
known about L. chinensis development.
The objectives of this study are to estimate the developmental threshold
7
history stages of L. chinensis. Such information could provide a means to construct a
practical model of the development of L. chinensis to establish integrated management.
Materials and Methods
Insects
Adults of L. chinensis were collected from a paddy field in Hyogo, in September
2002. The insects were kept at 25°C under a 16L–8D photoperiod in a plastic cage (9
cm dia.×5 cm height) with defrosted rice and distilled water (Yamashita 2010).
Effect of temperature on development of eggs, nymphs and pre-oviposition
Developmental periods of L. chinensis eggs, nymphs and pre-oviposition were
studied at four constant temperatures (±0.5°C): 18, 22, 25 and 30°C (16L–8D). Newly
laid eggs (<24 h old) were collected from the adults of the stock culture mentioned
above. Newly hatched nymphs (<24 h old) and newly emerged and mated female adults
(<24 h old) were placed individually in the plastic cage as mentioned above. The bugs
were provided with unhulled rice frozen at the milky stages every 2 d. All eggs and
nymphs were checked at 24 h intervals for survival and the presence of exuviae, which
8
day were confined in the plastic cage and maintained under the same conditions until
the female laid the first egg.
Estimation of pre-oviposition period after overwintering
I attempted to estimate the preoviposition period after overwintering under two
sets of conditions for adults of L. chinensis. Thirty-six adult males and 32 adult females
were collected from fields just before they were placed in a cage (9 cm dia.×5 cm
height) at a site in a paddy field in Kasai, Hyogo Prefecture from 5 November 2003 to
12 February 2004. The 19 pairs of adults that survived in the field were moved to cages
(6 cm dia.×4 cm height) with one pair in the incubator at 25°C (16L–8D) in the
laboratory, and whether females laid eggs was recorded every day.
On 9 November 2001, 27 adults that survived in the field were placed in cages (6
cm dia.×4 cm height), with one pair of adults in the incubator at 25°C (16L–8D) in the
9
Results
Table 1-1 shows the developmental period for eggs, nymphs and the
pre-oviposition period of L. chinensis. The period for each stage decreased as the
temperature increased from 18 to 30°C.
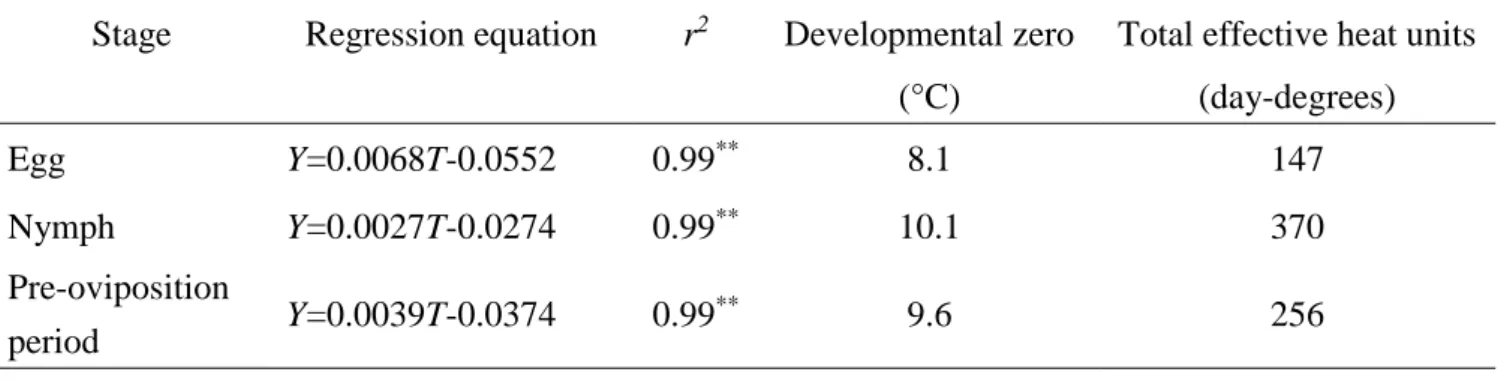
Table 1-2 shows the relationship between rearing temperature (T) and the rate of
development of L. chinensis. Developmental rates for eggs, nymphs and pre-oviposition
period increased linearly as the rearing temperature rose from 18 to 30°C. The rate of
development of the different life history stages in relation to temperature is expressed Table 1-1. Effects of temperature on developmental periods of eggs, nymphs and
pre-oviposition of adults in Leptocorisa chinensis
Temp. Eggs
Nymphs
Pre-ovispoition
(℃) d (mean±SD)
n
ad (mean±SD)
n
ad (mean±SD) n
a18 14.9 ± 2.6
89
47.1 ± 1.9
81
29.5 ± 3.6
14
22 10.4 ± 1.234
31.5 ± 1.7
29
21.1 ± 2.3
11
25 8.7 ± 0.7250
25.2 ± 2.5
142
17.3 ± 5.3
14
30 6.7 ± 0.7197 18.7 ± 2.1
172 12.4 ± 3.2
20
a Number of individuals tested.10
by the linear regression equation (Y=a×bT), where Y is the reciprocal of the number of
days (= development rate) and T is temperature (°C) (Patel and Schuster, 1983). There
was a significant linear relationship between temperature and development rates of eggs,
nymphs and pre-oviposition. The developmental zeros in eggs and nymphs were at 8.1
and 10.1°C, respectively, and the effective heat units were 147 day-degrees and 370
day-degrees, respectively. The developmental zero and effective heat unit for the
pre-oviposition period of nondiapause females were 9.6°C and 256 day-degrees,
respectively.
Table 1-2. Regression equations of developmental rate (Y) to rearing temperature (T) in Leptocorisa chinensis, developmental zero and total effective heat units calculated from the regression equation
Stage Regression equation r2 Developmental zero Total effective heat units
(°C) (day-degrees) Egg Y=0.0068T-0.0552 0.99** 8.1 147 Nymph Y=0.0027T-0.0274 0.99** 10.1 370 Pre-oviposition period Y=0.0039T-0.0374 0.99 ** 9.6 256 **p<0.01
11
The cumulative percentage of ovipositing females collected on 9 November 2001 and
12 February 2004 gradually increased (Fig.1-1). All females had laid their eggs by 41 d
after incubation. There was no significant difference in the days when egg laying started
after incubation between the populations collected on 9 November 2001 (28.6 d±5.7
SD) and 12 February 2004 (33.3 d ±4.9 SD) (p>0.05, t-test). From these results, an
effective heat unit of 469.7 day-degrees above 9.6°C, which is the threshold for females
of non-reproductive diapause, was assumed to be equal to that of reproductive diapause,
and was estimated to be required for the pre-oviposition period of overwintering
females. 0 20 40 60 80 100 1 6 11 16 21 26 31 36 41 46 C um ul at iv e % of ov iposi ti on
Day after incubation (25ºC,16L-8D)
Fig. 1-1. Pre-oviposition period of female, Leptocorisa chinensis transferred from the field to laboratory conditions. Solid and dashed lines indicate females collected on 9 November 2001 and 12 February 2004, respectively.
12
Discussion
In L. chinensis, Hasegawa et al. (1976) reported that this bug overwinters as adults.
In the present study, we recognized that all the adult females collected in winter
gradually began to lay eggs after incubation (Fig. 1-1). An effective heat unit of 469.7
d-degrees was required for the pre-oviposition period of overwintering females,
although the effective heat unit for the pre-oviposition period of undiapaused females
was 256 day-degrees. We have never observed the level of egg maturation in
overwintering females, but Hasegawa et al. (1976) observed that the females have no
mature eggs in the winter. The same genus, Leptocorisa oratorius Fabricius is known to
have reproductive diapause (Ito et al. 1993). It is considered that adult L. chinensis
would undergo reproductive diapause in the winter. There was no significant difference
in the timing of egg laying after incubation between the populations collected on 9
November 2001 and 12 February 2004. It is thought that the level of ovary maturity
between both of the populations is the same. Since there was not much difference, the
effective heat unit for the pre-oviposition period of L. chinensis females in the
meteorological data for November to February from 2001 to 2004 was recorded with
the AMeDAS system in Kasai City, Hyogo Prefecture. A more intensive experiment
13
Here, the number of annual generations of L. chinensis was estimated based on
these data and the meteorological data from 2001 to 2003 recorded by the AMeDAS
system in Kasai City, Hyogo Prefecture. The starting point for the calculation was 1
January. The results indicated that adults of the first generation and second generation
emerged in mid-July and late August, respectively (Fig. 1-2). The possibility also
showed that adults of the third generation would appear depending on temperature. In
0
250
500
750
1000
Ef fect iv e cu m u lat iv e te m p e ratu re (da y -d eg re e)J F M A M J J A S O N
D
Preoviposition period
Nymph
Egg
OW
G1
G2
G3
0
250
500
750
1000
Ef fect iv e cu m u lat iv e te m p e ratu re (da y -d eg re e)J F M A M J J A S O N
D
Preoviposition period
Nymph
Egg
OW
G1
G2
G3
Fig. 1-2. Schematic seasonal advancement of Leptocorisa chinensis generations from 2001 to 2003 in Hyogo Prefecture, estimated from the developmental zero and effective heat units of eggs, nymphs, and the pre-oviposition period. OW: Overwintering generation, G1: First generation, G2: Second generation, G3: Third generation. 2001: solid lines, 2002: broken lines, 2003: single dotted lines.
14
the earlier study, two peaks were observed in July and September in Shiga Prefecture
(Hasegawa et al., 1976), and also in Hyogo Prefecture (Yamashita, unpublished) in the
Kinki region. The present results are consistent with these reports, in that the number of
annual generations of L. chinensis was two.
Earlier studies investigated the effects of constant temperature on the rate of
development of L. chinensis reared on unhulled rice in milky stages every 7 d (Ishizaki
et al., 2002). Developmental thresholds of 13.5, 12.0 and 18.0°C, and thermal constants
of 92.4, 316.2 and 247.7 d-degrees were estimated for the eggs, nymphs and
pre-oviposition period, respectively. Developmental rates obtained from those results
did not provide a good estimate of the developmental times of populations in the field.
A direct extrapolation of the results to the field did not fit the field fluctuation of L.
chinensis. The development of L. chinensis reported by Ishizaki et al. (2002) was longer
than that in the present results. The estimated lower developmental threshold
temperatures recorded in the present study differed slightly from those estimated using
the results of Ishizaki et al. (2002). These differences may be due to differences in the
quality and quantity of food materials the tested insects consumed or/and differences in
developmental thresholds between the geographic populations of L. chinensis from
15
These results would be useful to estimate the occurrence of each stage of L.
chinensis. However, to apply the developmental simulation to L. chinensis, further
studies are needed for the measurement of another developmental parameter as well as
16
Chapter 2
Temperature and photoperiodic effects on induction and termination of diapause
in female Leptocorisa chinensis
Introduction
In Chapter 1, the developmental threshold temperature (9.6°C) and the heat unit
requirement (469.7 day-degrees) during the preoviposition period of female L. chinensis
were estimated. It was also estimated that there are two or three annual generations.
However, the field population phenology of L. chinensis was predicted from these
results without considering the consequences of diapause. Both the induction and the
termination of diapause in insects can influence the seasonal timing of growth and
reproduction in the generation immediately after diapause and in subsequent generations
(Tauber et al. 1986). Therefore, it is important to understand diapause induction and
termination in L. chinensis to determine the field phenology. In a recent study,
Tachibana and Watanabe (2007) reported that adult L. chinensis collected from Tsukuba
(36.02°N, 140.10°E), Ibaragi Prefecture, Japan, undergo a winter reproductive diapause,
and the critical day length for diapause induction was 13.75 h/d. However, they did not
17
one of the sensitive stages of insects that undergo reproductive diapause (de Wilde 1954,
Tauber and Tauber 1976, Numata 2004, Danks 2007). In Japan, nymphal and adult L.
chinensis occur in the fields in autumn when the day length is shorter than 13.75 h
(Takeuchi et al. 2005, Yamashita et al. 2005, Tachibana and Watanabe 2007).
This chapter investigated the influence of photoperiod and temperature on the
induction and termination of reproductive diapause in female nymphal and adult L.
chinensis. These results contribute to understanding the life cycle of L. chinensis in the
field and to the establishment of integrated management strategies to counteract this
major pest to rice.
Materials and Methods
Temperature and photoperiodic effects during nymphal and adult stages on
female L. chinensis survival and oviposition
A culture of L. chinensis was established from adults collected in a paddy field
and neighborhood sites in Kasai (34.55°N, 134.59°E), Hyogo Prefecture, Japan, during
October 2005. Before experimentation, the insect culture was maintained for two
generations at 25 ± 0.5°C under a photoperiod of 16:8 (L:D) h in a plastic cage (9 cm
18
distilled water for drinking (Yamashita, 2010). Randomly selected, newly emerged
nymphs (<1 d old) from the third filial generation were maintained under either a
constant long-day (16:8 [L:D] h) or short-day (12:12 [L:D] h) photoperiod at 25±0.5°C.
Adults that emerged from each photoperiod group (<1 d old) were then randomly
allocated to one of the two photoperiods at 25±0.5°C, 20±0.5°C, or 15±0.50.5°C, for a
total of 12 experimental adult groups. The initial number of females per group was 11-
24. All adults were reared as male-female pairs in plastic cups (6 cm dia.×4 cm height)
sustained on frozen, then defrosted, milk-ripe stage rice, along with distilled water.
Mortality and the number of females that oviposited were recorded daily for 100 d.
Fisher exact probability test was used to determine the significance of the differences in
the survival rate among the different conditions.
Photoperiodic effects on survival and postdiapause reproductive recovery of
female L. chinensis
L. chinensis- cultures were established from adults collected from a wild grass
habitat near a paddy field in Kasai, Hyogo Prefecture, Japan, on 8 December 2005.
Until experiments began, they were maintained in an outdoor cage (9 cm dia.×5 cm
19
overwintering adults were transferred to the laboratory for experiments on 1 February
and 29 March 2006, where they were randomly allocated into male-female pairs and
maintained under either a constant long-day (16:8 [L:D] h) or short-day (12:12 [L:D] h)
photoperiod at 25±0.5°C and sustained on defrosted frozen rice in the milk-ripe stage
and distilled water for the duration of the experiment. The initial number of females was
10-12 per cohort. For comparison, field-collected adults from Kasai were transferred to
the laboratory on 10 June 2005 and maintained under either a constant long-day (16:8
[L:D] h) or short-day (12:12 [L:D] h) photoperiod at 25±0.5°C. Mortality and the
number of females that oviposited were recorded daily for the duration of the
20
Results
Temperature and photoperiodic effects during nymphal and adult stages on female
L. chinensis survival and oviposition
The effects of temperature and photoperiod on survival and oviposition of adults
reared as nymphs under a long-day photoperiod (16:8[L:D] h) are shown in Fig. 2-1. L.
chinensis females started ovipositing on day 11 after adult emergence at 25°C under a
long-day photoperiod, and >60% of the females oviposited during the experimental
period (Fig. 2-1a). At 20°C under a long-day photoperiod, adults started ovipositing on
day 21 after adult emergence, and the number of ovipositing females increased
gradually (Fig. 2-1c). Females under a long-day photoperiod at 15°C laid no eggs,
although they survived for the duration the experimental period (100 d; Fig. 2-1e).
Females under a short-day (12:12 [L:D] h) photoperiod at 25°C started ovipositing on
day 10 after adult emergence. They ceased ovipositing within a few days but survived
for the duration of the experimental period (Fig. 2-1b). Females under a short-day
21
Fig. 2-1. Effects of temperature and photoperiod on survival and oviposition of adult female L. chinensis reared as nymphs under a long-day photoperiod of 16:8 (L:D) h under 25°C (a and b) , 20°C (c and d), and15°C (e and f). Solid and broken lines represent the percentage of adult female surviving and ovipositing, respectively, The initial number of newly emerged females (<1 d old) was 11-24 (n).
Days after transfer
e 15°C n=12 f 15°C n=12 d 20°C n=11 c 20°C n=18 b 25°C n=24 16L- 8D 12L-12D a 25°C n=18
22
The effects of temperature and photoperiod on survival and oviposition of adults
that were reared as nymphs under a short-day photoperiod (12:12 [L:D] h) are shown in
Fig. 2-2. Females under a long-day photoperiod (16:8 [L:D] h) at 25°C started
ovipositing on day 40 after the start of the experiment and the number of ovipositing
females increased gradually (Fig. 2-2a). Under a long-day photoperiod at 20°C, a small
number of females oviposited (Fig. 2-2c). However, no females oviposited under any
other conditions (Fig. 2-2e and f).
There were no significant differences in the survival rate at day 100 between the
females in each photoperiodic condition in the same temperature in Figs. 2-1 and 2 (P>
23
Fig. 2-2. Effects of temperature and photoperiod on survival and oviposition of adult female L. chinensis reared as nymphs under a short-day photoperiod of 12:12 (L:D) h under 25°C (a and b), 20°C (c and d), and 15°C (e and f). Solid and broken lines represent the percentage of adult female surviving and ovipositing, respectively. The initial number of newly emerged females (< 1 d old) was 11-24 (n).
Days after transfer
e 15°C n=12 f 15°C n=11 d 20°C n=11 c 20°C n=12 b 25°C n=12 16L- 8D 12L-12D a 25°C n=12
24
Photoperiodic effects on survival and postdiapause reproductive recovery of
female L. chinensis
Adult females transferred from the overwintering cage in the field to the long-day
photoperiod at 25°C in the laboratory on both 1 February and 29 March commenced
ovipositing day ≈40 after transfer (Fig. 2-3a and c). However, no adult females that
were transferred on either date to the short-day photoperiod at 25°C oviposited for the
duration of the experiment (150 d; Fig. 2-3b and d). Field-collected adult females that
were transferred to a long-day photoperiod at 25°C in the laboratory on 10 June
commenced oviposition almost immediately and continued to lay eggs for the duration
of the experiment (Fig. 2-3e). Similarly, collected adult females transferred to a
short-day photoperiod at 25°C in the laboratory on the same day commenced
ovipositing immediately. However, after 14 d the number of ovipositing females
25
Fig. 2-3. Survival and oviposition of adult female L. chinensis transferred from field conditions to the laboratory and maintained at 25°C and either a 16:8 (L:D) h (left) or 12:12 (L:D) h (right) photoperiod. Solid and broken lines represent the percentage of adult female surviving and ovipositing, respectively. The initial number females was 10-12 (n). Survivorship and oviposition data reported in a-d are for adults collected on 8 December 2005 and in e and f for adults collected on 10 June 2005. Dates when adults were transferred to the laboratory conditions are shown.
Days after transfer
b 1 Feb. n=11 a 1 Feb. n=10 e 10 Jun. n=12 f 10 Jun. n=11 d 29 Mar. n=11 c 29 Mar. n=10
26
Discussion
Nymphal and adult stages are the most sensitive to photoperiod and temperature
in many insects that undergo reproductive diapause (de Wilde 1954, Tauber and Tauber
1976, Numata 2004, Danks 2007). In L. chinensis, oviposition curves differed between
similarly maintained adults that had been reared as nymphs under differing photoperiod
conditions (Figs. 2-1 and 2). Likewise, oviposition was influenced by differing
photoperiod conditions during the adult stage of nymphs that were reared under the
same conditions (Figs. 2-1 and 2). More specifically, adult females that had been reared
as nymphs under the long-day photoperiod oviposited for only a short time under
short-day conditions at 25°C (Fig. 2-1b). Females did not oviposit under a short-day
photoperiod at 20°C, even when they were reared as nymphs under the long-day
photoperiod (Fig. 2-1d). These findings suggest that short-day photoperiod conditions
during the adult stage induce diapause in L. chinensis females. This is consistent with
the findings of Tachibana and Watanabe (2007).
There were also differences in oviposition patterns between the adult stages of L.
chinensis nymphs reared under the long-day and short-day photoperiods (Fig. 2-1a and
c , Fig. 2-2a and c). Adults began ovipositing after ≈40 d under a long-day photoperiod,
27
Females required constant long-day photoperiod conditions to initiate oviposition,
indicating that in L. chinensis, not only adult but also nymphal stages are sensitive in the
process of reproductive diapause induction. Thus, both adults and nymphs can undergo
a reproductive diapause. This is further supported by the fact that in Japan, both nymphs
and adults can be found in the field when the daylength is shorter than the critical
photoperiod of 13.75 h/d. (Takeuchi et al. 2005, Yamashita et al. 2005, Tachibana and
Watanabe 2007). Thus, it was concluded that L. chinensis females exhibit a facultative
adult diapause and that photoperiod controls its’ induction and termination, confirming
the results of Tachibana and Watanabe (2007).
There were also differences in oviposition patterns between the adult stages of L.
chinensis nymphs reared under the long-day and short-day photoperiods (Figs.2-1a and
c and 2a and c). Adults began ovipositing after ≈40 d under a long-day photoperiod,
when they had been reared as nymphs under a short-day photoperiod (Fig. 2-2a and c).
Females required constant long-day photoperiod conditions to initiate oviposition,
indicating that in L. chinensis, not only adult but also nymphal stages are sensitive in the
process of reproductive diapause induction. Thus, both adults and nymphs can undergo
a reproductive diapause. This is further supported by the fact that in Japan, both
28
critical photoperiod of 13.75 h/d. (Takeuchi et al. 2005, Yamashita et al. 2005,
Tachibana and Watanabe 2007). Thus, it was concluded that L. chinensis females
exhibit a facultative adult diapause and that photoperiod controls its’ induction and
termination, confirming the results of Tachibana and Watanabe (2007).
The results of the transfer experiments also indicate L. chinensis females exhibit a
facultative adult diapause. For example, when transferred to 25°C in the laboratory,
field overwintering adult females did not commence oviposition when maintained under
short-day conditions but commenced ovipositing after ≈40 d under long-day
photoperiod conditions (Fig. 2-3). Moreover, field-collected, post overwintering (June)
female adults maintained under short-day conditions did not oviposit as readily or as
consistently as those maintained under the long-day photoperiod at 25°C (Fig. 2-3).
The termination of diapause in L. chinensis might also be infuenced by
temperature. When females were reared as nymphs under a short-day photoperiod, and
maintained as adults under a long-day photoperiod, oviposition began after ≈40 d, but
the percentage that oviposited over the experimental period of 100 d was lower at 20°C
than at 25°C (Fig. 2-2a and c). Females did not lay any eggs at 15°C regardless of the
photoperiod conditions. Tachibana and Watanabe (2007) assumed that reproductive
29
longer than the critical photoperiod and when temperatures exceeded a given high,
although females never began oviposition during the 30-d experiments. In our study,
field overwintering adult females transferred to 25°C in the laboratory did not
commence oviposition when maintained under short-day conditions but did so under
long-day conditions after ≈40 d (Fig. 2-3a and c). These results suggest that, to
commence oviposition after winter, female L. chinensis require a long-day photoperiod
and ≈40 d at temperatures >20°C.
Oviposition patterns for the 1 February cohort were basically the same as those for
the 29 March cohort when they were transferred to both short-and long-day photoperiod
conditions (Fig. 2-3a and d). These results indicate that changes in photoperiod or
temperature did not terminate diapause in L. chinensis females by 29 March. However,
by 10 June, adult female L. chinensis were able to oviposit readily when transferred to
long-day photoperiod conditions and had thus recovered their reproductive ability after
diapause. The females also oviposited for a short period when they were transferred to
short-day photoperiod conditions, but the number of ovipositing females gradually
decreased (Fig. 2-3f). This pattern was similar to that of adult females reared as nymphs
under a long-day photoperiod and then held under short-day photoperiod at 25°C (Fig.
30 even after winter.
Additional studies are needed to evaluate the developmental parameters for field
populations of L. chinensis in various climatic regions. Furthermore, in the current study,
the diapause of L. chinensis was evaluated using oviposition and survival data without
examining the condition of the ovaries and fat body. To get more information about
31
Chapter 3
A pecky rice-causing stink bug Leptocorisa chinensis escapes from volatiles emitted
by excited conspecifics
Introduction
Disturbed stink bugs emit pungent volatile compounds that could have several
ecological functions including behavior regulation and defense against predators (Blum
1985; Aldrich 1995). The behavior regulator function might be used for the behavioral
control of stink bugs in agroecosystems. The volatiles from smashed bugs likely repel
other stink bugs in the area. We hypothesized that the volatiles emitted from disturbed L.
chinensis could be used to repel conspecifics in rice fields and reduce the number of
pecky rice grains.
Leal et al. (1996) identified the volatiles emitted from L. chinensis anaesthetized
with CO2to minimize the release of defensive secretions, and found that a 5:1 mixture
of (E)-2-octenyl acetate and octanol, major components of the emissions of undisturbed
L. chinensis was an attractant pheromone. However, the chemistry and ecological
functions of the volatiles from disturbed L. chinensis individuals have not yet been
32
The objective of this study was to clarify whether the volatiles from disturbed L.
chinensis elicit excitement and escape behavior in conspecifics as a first step to test the
above hypothesis. We first investigated whether L. chinensis escaped from disturbed
conspecifics in an observation arena under laboratory conditions. We then analyzed the
volatiles emitted by both disturbed and undisturbed L. chinensis and observed the
responses of undisturbed L. chinensis to the components in the volatiles from disturbed
conspecifics.
Materials and methods
Insects
A colony of L. chinensis was established from adults collected from paddy fields
in Kasai, Hyogo Prefecture, Japan, in October 2005, and maintained for two generations
prior to the experiments. Males and females were kept in a plastic cage (9 cm dia.×5 cm
height) in a climate controlled room at 25 ± 0.5°C under a photoperiod of 16L:8D, with
distilled water in a small Petri dish and rice panicles (ca. 50 mm) in the milk-ripe stage
as food. After mating occurred in the cage, I used them for the experiments. For
33
Fig. 3-1. Bioassay set-up to evaluate the responses of Leptocorisa chinensis to alarm odours . An undiaturbed adult was introduced into an observation arena made from two plastic cups (85mm diameter 40mm height), and then either (A) an L.chinensis or (B) a piece of filter paper imoregnated with a synthetic chemical was introduced through the hole (18mm diameter).
Observation arena An excited L. chinensis
Rice grains for food A piece of filter paper with sample A plastic cup
Wet cotton towel for water supply Hole (5mm diam.) A calm L.chinensis (ca.1.5cm long) Hole (18mm diam.) 8.5cm 4 cm water ( A ) ( B )
34
Bioassays
The bioassay set-up was constructed to evaluate the excitement and escape
behaviour of undisturbed L. chinensis to cues from excited conspecifics (Fig. 3-1).
Water (60 cc) was provided in a lidded plastic cup (85 mm diameter, 40 mm height).
The lid had a 5-mm-diameter opening, into which a piece of wet cotton towel was
inserted. I placed 10 rice seeds at the milk-ripe stage on top of part of the lid as food. I
covered the lid with an inverted plastic cup of the same size with one escape hole (18
mm diameter) at the side. I called the inside of the inverted plastic cup the observation
arena.
To introduce an undisturbed individual into the arena, we carefully transferred one
L. chinensis from a rearing cage to a test tube (18 mm diameter, 120 mm length), and
the opening of the test tube was connected to the opening in the inverted plastic cup to
allow the bug (initial occupant) to enter the arena. When the bug stopped moving, it was
considered undisturbed. For the control experiment, we introduced an undisturbed bug
into the arena in the same way. For experimental treatments, we introduced an
individual of the same gender using forceps with gentle nipping to disturb it and cause it
to emit the pungent odour. The two individuals were distinguishable because the second,
35
initial occupant was observed for 3 min. When the occupant raised its antennae to scan
the air and started walking actively, it was judged to be excited. When it left the arena
through the hole, we judged that it had escaped. We measured the duration of time until
it showed excitement and escape behaviour. The introduced disturbed individual was
actively walking in a circle on the ceiling of the arena. During the observation, the two
individuals did not interact in the arena.
To further test the ecological functions of the pungent volatiles from L. chinensis,
I conducted the same bioassays using the dominant chemical components. A pure
compound (1 or 10 μg) was applied to a piece of filter paper (10×10 mm2) and
introduced into the observation arena containing an undisturbed L. chinensis through the
opening (Fig. 1-1). The amounts of pure compound [(E)-2- octenal] were determined
based on chemical analyses: the amounts released by one disturbed female per minute
were ca. 4.5–14μg . We measured the duration of time until the bug showed excitement
and escape behaviour. The bioassays were conducted in a climate-controlled room (25 ±
3°C, 50–60% relative humidity; during 10:00–16:00). I repeated each experiment, 5–10
times per day on 2–4 experimental days.
Then the responses of males and females of L. chinensis were tested to
36
to excite the bugs. In this study, we needed (E)-2-octenal to volatilise slowly to measure
the air concentrations, so we dissolved (E)-2-octenal in methanol (10% v/v) and then
diluted with distilled water to make 0.1, 0.01, and 0.001% solutions. For each bioassay,
we used 1 mL solution impregnated into a piece of moist cotton wool (2 cm×2 cm×0.5
cm) on the same size of aluminium foil. We conducted experiments in a mesh cage (30
×30 × 50 cm3: 2 mm mesh) with an electric fan (flow rate 50 cm/s; SY124010L, 40 mm
× 30 mm × 10 mm thickness; Size Corporation, Tokyo, Japan) in the centre of the cage.
We carefully introduced 4–5 undisturbed L. chinensis (either males or females) into the
mesh cage as described above. When the bugs stopped moving, they were considered
undisturbed. We then placed a piece of impregnated cotton wool on the fan. We
observed the flight behavior for 5 min. In our preliminary behavioural observations of
both sexes in the mesh cage, we confirmed that flying individuals did not elicit any
behavioural responses in undisturbed individuals. For both males and females, we
conducted the experiments 4 times (0.1% solution and 0.01% solution) and 3 times
(0.001% solution) on 1–3 experimental days in a climate-controlled room (25 ± 3°C,
37
Volatile collection and chemical analysis
Either a male or a female L. chinensis was used for volatile sampling. For
collection from a disturbed individual, it was nipped with forceps. Immediately
afterwards, it was put in a 200-mL glass bottle with an air inlet and outlet (55 mm
diameter, 110 mm height). For the collection of volatiles from an undisturbed individual,
it was allowed to walk into the volatile collection vial from the rearing cage. An
undisturbed individual was motionless, walking, or flying in the bottle. Volatile
collections were done on four individuals per gender and treatment during daytime
(25±3°C, 50–60 % relative humidity; during 10:00–15:00).
The headspace volatiles in the glass bottles were collected for 1 min (disturbed
adult) or 10 min (undisturbed) at flow rate of 100 mL/min using Tenax adsorbent in a
glass tube (Tenax TA 20/35 100 mg; 3-mm inner diameter (ID), 160 mm long; GL
Science, Tokyo, Japan). The volatile collection time was determined based on
preliminary chemical analyses. The trapped compounds from disturbed adults were
eluted with 2 mL n-hexane (Wako Pure Chemical Industries Ltd.) containing n-eicosane
(0.5 μg; Wako Pure Chemical Industries Ltd.) as internal standard for the recovery rate.
The eluate was concentrated by nitrogen gas flow to 10 μl. One microliter of
38
chromatograph–mass spectrometer (GC–MS; GC: Agilent 6890 with HP-5MS capillary
column: 30 m long, 0.25 mm i.d. and 0.25 μm film thickness; MS: Agilent 5973 mass
selective detector, 70 eV with He as carrier gas; Agilent, Santa Clara, CA, USA). The
GC oven temperature was programmed to rise from 40°C (5 min hold) to 280°C at
15°C/min. The compounds were identified by comparing their mass spectra and
retention times with those of authentic compounds and quantified using a calibration
curve of the respective compound.
Air in the flight cage with volatilised (E)-2-octenal was collected for 3 min at flow
rate of 100 mL/min using Tenax adsorbent in a glass tube [Tenax TA 20/35 100 mg;
3-mm inner diameter (ID), 160 mm long]. The collections were repeated four times for
each of the four concentrations. The volatile collection time was determined based on
preliminary chemical analyses. The collected volatile compounds were analysed by
GC–MS as described above, except for the injection method. The GC–MS was
equipped with a thermal desorption cold trap injector (TCT; CP4010, Chrompack, The
Netherlands). Headspace volatiles collected on Tenax-TA were released in the TCT
thermal desorption unit at 220°C for 8 min in He flow. The desorbed compounds were
collected in the TCT cold trap unit (SIL5CB-coated fused silica capillary) at -130°C.
39 capillary column of the GC.
Chemicals
Hexyl acetate, octyl acetate, octanal, (E)-2-octenal and octanol were purchased
from Wako Pure Chemical Industries Ltd., Osaka, Japan. (E)-2-Octenyl acetate and
(Z)-2-octenyl acetate were synthesised by Sumika Technoservice Corporation (Hyogo,
Japan).
Statistical analyses
The amounts of each volatile compound emitted from males and females were
compared by t test. The time durations needed for excitation and escape were tested by
the Kaplan–Meier time-to-event model using a log-rank test statistic. All statistical tests,
except multiple-comparison tests, had a significance of 0.05. For multiple comparisons
of duration, we adjusted the significance according to Holm’s sequentially rejective
Bonferroni test to reduce type I errors. The multiple comparisons involved three
log-rank tests, therefore the lowest of the three P values was compared with a = 0.0167
(0.05/3), the second lowest with α = 0.025 (0.05/2), and the third lowest with α = 0.05
40
(E)-2-octenal were analysed using two-way analysis of variance (ANOVA) with factors
concentration and sex, and their interaction. The data of the flight proportion were
arcsine square root transformed before two-way ANOVA, and were weighted in the
analysis by the number of individuals released into the mesh cage. All statistical
analyses were conducted using the JMP software package (version 9.0.2; SAS Institute,
Cary, NC, USA).
Results
Response of L. chinensis to volatiles emitted from conspecifics
Following the introduction of a disturbed same-sex conspecific, all undisturbed
females (n = 19) and males (n = 16) exhibited excitement within 40 s (Fig. 3-2a) and
escaped the arena within 3 min (Fig. 3-2b), with no differences between the sexes (P =
0.14 and P = 0.72 log-rank test, respectively). When an excited individual touched the
hole (18 mm diameter) with antennae, the individual immediately walked out of the
arena through the hole and flew to the fluorescent lights on the ceiling. In contrast,
addition of an undisturbed same-sex conspecific elicited no excitement or escape
41
Chemical analyses of volatiles emitted from L. chinensis
females with a disturbed female
Undisturbed males with a disturbed male
Undisturbed females with an undisturbed female males with an undisturbed male
Fig. 3-2. Proportions of excitement and escape of undisturbed Leptocorisa chinensis females (n = 19) and males (n = 16) when exposed to undisturbed or disturbed conspecifics of the same gender. Lines with the same lower-case letter are not significantly different (Holm’s sequentially rejective Bonferromi test after Kaplan-Meier time-to-event model using a log-rank test statistic, P<0.05). Females (n = 19) and males (n = 16) exposed to an undisturbed individual showed no excitement/escape behavior.
42
Chemical analyses of volatiles emitted from L. chinensis
Volatiles from undisturbed males and females were below detectable levels (data
not shown). When disturbed, both male and female L. chinensis adults (n = 4) emitted
hexyl acetate, octyl acetate, (Z)-3-octenyl acetate, (E)-2-octenyl acetate, octanal,
(E)-2-octenal and octanol (Table 3-1). Females emitted significantly higher amounts
of hexyl acetate, octyl acetate, (E)-2-octenyl acetate, (E)-2-octenal and octanol than
males (t test) (Table 3-1).
Table 3-1. Amounts of volatile compounds recorded in headspace of disturbed male and female Leptocorisa chinensis (n = 4)
Compound ng (relative amounts %) P value( t test)
Male Female Hexyl acetate 27±14 (0.5) 193±40 (1.2) 0.007 Octyl acetate 273±86 (5.5) 1170±347 (7.1) 0.046 (Z)-3-octenyl acetate 23±12 (0.4) 82±27 (0.5) 0.09 (E)-2-octenyl acetate 41±31 (0.8) 590±198 (3.6) 0.03 Octanal 22± 6 (0.4) 29± 3 (0.2) 0.31 (E)-2-octenal 4536±1191 (92.0) 14341±2208 (87.0) 0.008 Octanol 10±10 (0.2) 72±16(0.4) 0.02
43
Response of L. chinensis to synthetic compounds of volatiles emitted from a
disturbed individual
I studied the responses of L. chinensis females to three volatile compounds, i.e.
octyl acetate, (E)-2-octenyl acetate and (E)-2-octenal, which were predominant in
volatiles emitted from a disturbed male and/or female (Table 3-1). The numbers of
individuals tested for each compound is shown in Fig. 3-3. Octyl acetate was the least
active in eliciting excitement/escape behaviour at two doses (Holm’s sequentially
rejective Bonferroni test after logrank test) (Fig. 3-3a, b). At 1-μg dose, no individuals
escaped from the arena when offered octyl acetate, while 60–90% of individuals
escaped when offered (E)-2-octenyl acetate or (E)-2-octenal with no significant
differences between the compounds (P = 0.29, log-rank test) (Fig. 3-3c). At the 10-μg
dose, the proportion of escape for octyl acetate was significantly lower than for the
other two compounds, while there were no significant differences between
(E)-2-octenyl acetate and (E)-2-octenal (Holm’s sequentially rejective Bonferroni test
44
Undisturbed Undisturbed
Fig. 3-3. Proportions of excitement (a, b) and escape (c, d) of undisturbed Leptocorisa chinensis female when exposed to synthetic chemicals at different concentrations of (a, c) 1 μg and (b, d) 10 μg. Lines with the same lower-case letter are not significantly different (Holm’s sequentially rejective Bonferroni test after Kaplan–Meier time-to-event model using a log-rank test statistic, P<0.05). n = 19 for (E)-2-octenal, 17 for (E)-2-octenyl acetate, 7 for octyl acetate
45
Concentration of (E)-2-octenal in air needed to induce escape behaviour in L.
chinensis
Males started showing flight behaviour when exposed to 0.001% solution of
(E)-2-octenal, while females started showing flight behaviour when exposed to 0.01%
solution (Fig. 3-4). The solution concentration significantly affected the flight behaviour
(F2,16 = 10.19, P = 0.001). However, effects of sex and the interaction (concentration 9
sex) were not significant (sex: F1,16 = 10.13, P = 0.72; interaction: F2,16 = 2.44, P = 0.30).
The headspace analyses of air in the cage showed that 0.01 and 0.1% solutions of (E)-
2-octenal resulted in 1.5 ppbV and 9.3 ppbV, respectively. The concentration of
46
Discussion
Undisturbed males and females of L. chinensis became excited and escaped from
the arena when exposed to previously excited individuals of the same gender. Their
behaviours suggested that they responded to volatiles emitted from disturbed
conspecifics. Alternatively, visual/physical cues, such as walking and sounds associated Fig. 3-4. Proportions of flight behavior of undisturbed Leptocorisa chinensis males and female in mesh cage when offered emulsified (E)-2-octenal at different concentrations
47
with movement, might have affected the excitation and escape behaviour. To clarify the
effects of the volatiles, we conducted chemical analyses and tested the effects of the
volatile components on the behaviour of L. chinensis.
Most of the volatiles found from the disrupted individuals were C8 aldehydes,
alcohol and acetates. The major compounds found in the headspace of disturbed males
and females were (E)-2-octenal and octyl acetate. (E)-2-Octenyl acetate was also one of
the major compounds in the headspace of disrupted females. Gunawardena and
Bandumathie (1993) reported that the chemical compositions of defensive secretions
produced by disturbed Leptocorisa oratorius males and females were similar; the two
major components in both were (E)-2-octenal (76% v/v) and octyl acetate (16%w/w). In
this study, in total, females emitted~3 times more volatiles than males. A similar trend
was reported in disturbed and undisturbed Lygus lineolaris (Wardle et al. 2003).
The relative amounts of (E)-2-octenal and (E)-2-octenyl acetate were 87–92% and
0.8–3.6%, respectively. Thus, we concluded that (E)-2-octenal was one of the major
factors eliciting the excitement and escape behaviour in L. chinensis females in the
arena when a disturbed individual of the same gender was introduced. No significant
differences in escape proportions were observed between males and females (Fig. 3-2b),
48
escape behaviour in L. chinensis males. This conclusion was supported by the bioassays
using emulsified (E)-2-octenal in the mesh cage.
The proportions of escape in response to a disturbed female and to synthetic
volatile compounds [(E)-2-octenal and (E)-2-octenyl acetate (1-μg dose)] were not
significantly different (P = 0.23, log-rank test). The low activity of hexyl acetate
suggested that the location of the double bond at the (E)-2 position was more important
than the presence of the aldehyde group. Further studies are needed to evaluate this idea.
Further, we did not test the effects of minor compounds (less than ca. 1% in the blends
from both genders: hexyl acetate, (Z)-3-octenyl acetate, octanal and octanol) on
excitement and escape behaviours. Studies on the ecological functions of such volatiles
are needed as well.
Leptocorisa chinensis males are strongly attracted to a 5:1 mixture of
(E)-2-octenyl acetate and octanol (Leal et al. 1996; Watanabe et al. 2009; Fukatsu et al.
2012). These compounds were also detected from males and females at ratios of
approximately 2:1 and 20:1, respectively, in our study. Leal et al. (1996) also reported
that a whole blend did not attract males and the attractiveness of a binary mixture
decreased with addition of (Z)-3-octenyl acetate. In this study, we also detected
49
(E)-2-octenyl acetate and octanol emitted from a disrupted adult, if any, would have
been hampered by the different ratios and/or by the presence of other compounds, such
as (Z)-3-octenyl acetate.
Leal et al. (1996) identified compounds found in this study as well as nonanal and
(E)-2-octenol in the headspace and hexane extracts of both male and female L. chinensis
that were anaesthetised with CO2 to minimize the release of defensive secretion.
CO2-induced anaesthesia might have resulted in production of small amounts of
volatile compounds. Our inability to detect volatiles from undisturbed males and
females was probably due to differences in collection methods.
As mentioned in the ‘‘Introduction,’’ in Sri Lanka, farmers protect rice plants by
putting smashed adult stink bugs, including Leptocorisa species, around their fields
(Yamashita, personal communication). Our data suggest that the volatiles from these
smashed bugs would have repelled stink bugs from the agricultural fields. In this study,
we clarified that (E)-2-octenal and (E)-2-octenyl acetate are involved in the
excitement/escape behavior of L. chinensis. Continual release of synthetic (E)-2- octenal
in paddy fields at concentration of ca. 2 ppbV or higher is expected to protect rice grains
from L. chinensis damage during critical stages of the growing season. I in chapter 4
50
Chapter 4
Evaluation of the behavior regulator of Leptocorisa chinensis to control this species
in paddy fields
Introduction
The previous chapter outlines the possible use of volatile compounds eliciting
excitement/escape behavior in L. chinensis to control this species in paddy fields
(Chapter 3). The life history of L. chinensis has also been estimated to predict and
control L. chinensis invasion during the rice heading stage (Chapters 1 and 2).
However, to the best of our knowledge, the efficacy of the behavior regulator of L.
chinensis against the control of this species has not yet been investigated in paddy fields.
The aim of the present study was to establish a program for the pest control of L.
chinensis in paddy fields using a behavior regulator of L. chinensis. To achieve this goal,
we investigated the effectiveness of the behavior regulator of L. chinensis in paddy
fields.
Materials and Methods
We conducted behavior regulator release experiments in two open paddy fields at
51
August and September 2006. The experimental details are summarized in Fig.4-1, We
conducted two release experiments, trials 1 and 2. Each experiment involved two plots,
one in which behavior regulators were released and a control plot that received no
treatment (Fig. 4-1). Two paddy fields were transplanted with seedlings at intervals of
18 to 20 days from mid-May. Each field had an area of 350–600 m2 and adjoined the
other, and the rice variety was Koshihikari. No insecticides were used in any of the
fields throughout the study. A fixed plot within each field of about 288 m2 (12 m × 24
m) was used for a routine population census. The routine population census was
conducted from August to September. The number of L. chinensis and peck rice were
systematically counted in 49 hills per plot on each census date. The concentration of the
behavior regulator was also estimated using Twister (Gerstel, M¨ulheim and der Ruhr,
Germany; 1 mm film thickness × 10 mm length), which uses magnetic stir bars coated
with polydimethylsiloxane, in each plot on each census date. The Twister was set as
shown in Fig.4-1. After one day, the Twister was taken to the laboratory and analyzed
by gas chromatograph–mass spectrometer (GC-MS).
To test for the effects of the number of L. chiensis and pecky rice on the behavior
regulator, the data were analyzed by t-test and one-way analysis of variance (ANOVA)
52
Results and Discussion
The number of L. chinensis and the percentage of pecky rice in the plots with the
behavior regulator were lower than those in control plots (Figs. 4-2 and 4-3). In
particular, the number of pecky rice in each plot with the behavior regulator was
surprising less than 0.1, which is an excellent value for the investigation of rice grade
(Fig. 4-4). The concentrate of behavior regulator was recognized to be higher than the
threshold to release excitement/escape behavior in L. chinensis.
53
Fig. 4-2 Number of N. chinensis in alarm pheromones and control sites.
P<0.05
N
o
.
o
f
L.c
h
in
en
si
s
54
Based on these results for the open paddy fields, the behavior regulator would be
effective for the control of L. chinensis to reduce pecky rice. Thus, the utilization of the
behavior regulator can be considered a new technique for pest control. The behavior
regulator does not harm beneficial species and could become a key technique for
efficient and sustainable insect management strategies in the future.
1 2 0.0 0.2 0.4 0.6 0.8 1.0 1.2 V2 V1 Control Alarm pheromones % of p ec ky ri ce P<0.05
Fig. 4-3 Percentage of pecky rice in alarm pheromones and control sites. Vales translate to arcsince.
55 0 2000000 4000000 6000000 8000000 10000000 12000000 14000000 16000000 18000000 0 5 10 15 20 25 30 35 C once nt ra ti on of al ar m pher om one
Distance from alarm pheromones (m)
Fig. 4-4 Relationship between distance from alarm pheromones and concentration of them in a paddy field. Different letters indicate significant differences (P<0.05 Bonferroni corrections after ANOVA). Red line indicates the threshold.
56
Summary
The rice stink bug Leptocorisa chinensis (Dallas) (Hemiptera: Alydidae) is a
major cosmetic pest, and is one of the main causes of pecky rice (Suzuki, 2001). Thus,
the effective and environmentally benign control of this species in paddy fields is
essential, without the need for chemical insecticides. The aim of the present study was
to develop a new method of control using a behavior regulator found in L. chinensis.
In Capters 1 and 2, I examined the life history of L. chinensis to predict and
control L. chinensis invasion during the rice heading stage. First, I investigated the
effects of a range of constant temperatures (18°C–30°C) on the development of eggs,
nymphs and the pre-oviposition period of adult females in L. chinensis. The duration of
all stages decreased as the temperature increased from 18°C to 30°C. Developmental
thresholds of 8.1°C, 10.1°C and 9.6°C were estimated for the eggs, nymphs and
pre-oviposition period, respectively. Thermal constants of 147 and 370 degree-days
above the thresholds of 8.1 degree-days and 10.1°C were required for the development
of the eggs and nymphs, respectively. The pre-oviposition period required 256
degree-days above the developmental threshold of 9.6°C. After placing individuals
collected in November and February in an incubator in the laboratory, 469.7
57
to be equal to that of diapause female adults, were estimated to be required for the
pre-oviposition period of individuals that had overwintered. The present results were
useful to predict the field population phenology of L. chinensis in Japan.
Next, the effects of temperature and photoperiod on diapause induction and
termination in L. chinensis were studied under constant conditions or by using transfer
experimental protocols. Nymphs were reared either under a long-day (16:8 [L:D] h) or a
short-day (12:12 [L:D] h) photoperiod at 25°C in the laboratory. Females oviposited at
25°C and 20°C under the long-day photoperiod. However, females did not lay eggs
within 100 d at 15°C under the long-day photoperiod or at 20°C and 15°C under
short-day conditions. At 25°C, when nymphs were reared under a long-day photoperiod
and transferred to a short-day photoperiod on the day of adult emergence, females
started oviposition in 10 d but stopped shortly thereafter. When nymphs were reared
under a short-day photoperiod and transferred to a long-day photoperiod on the day of
adult emergence, females started oviposition in 40 d. Females that had been transferred
from the field on 1 February and 29 March to long-day laboratory conditions at 25°C
started oviposition in 40 d. However, females that had overwintered in the field were
transferred to a short-day photoperiod at 25°C on 1 February and 29 March did not start
58
considered to be sensitive to reproductive diapause induction and termination signals
both before and after overwintering. Female L. chinensis require not only a long-day
photoperiod and 40 d at a temperature of 20°C or higher but also threshold temperatures
for the start of oviposition even after the winter. From the results of Chapters 1 and 2, I
was able to predict L. chinensis invasions during the rice heading stage.
In Chapter 3, I examined whether L. chinensis escaped from disturbed
conspecifics in an observation arena under laboratory conditions. When an undisturbed
individual of the same gender was introduced into the arena, the initial occupying L.
chinensis did not show any behavioral responses. However, when a disturbed
conspecific of the same gender was introduced, the initial occupant was immediately
excited and escaped from the arena through a hole, suggesting that the pungent volatiles
from the disturbed conspecific caused excitement/escape behavior.
Next in Chapter 3, I analyzed the volatiles emitted by both disturbed and
undisturbed L. chinensis and observed the responses of the undisturbed L. chinensis to
the components in the volatiles from disturbed conspecifics. Chemical analyses using a
GC–MS showed that disturbed adults of both sexes emitted octanal, (E)-2-octenal,
octanol, hexyl acetate, (Z)-3-octenyl acetate, octyl acetate and (E)-2-octenyl acetate.