Title 苔類ゼニゴケにおけるフィトクロムシグナル伝達機構の 解析( Dissertation̲全文 )
Author(s) 井上, 佳祐
Citation 京都大学
Issue Date 2016-09-23
URL https://doi.org/10.14989/doctor.k20035
Right
Type Thesis or Dissertation
Textversion ETD
Kyoto University
苔類ゼニゴケにおけるフィトクロム シグナル伝達機構の解析
井上 佳祐
1
目次
要旨 2
略語表 3
序論 5
結果 13
考察 35
材料と方法 39
文献 45
謝辞 54
2
要旨
フィトクロムは赤色光および遠赤色光を受容する光受容体であり、植物の成長や発生の 多くの面で重要な役割をもつ。シロイヌナズナを用いた解析から、フィトクロムは主に核内 で機能し、赤色光シグナルの抑制因子として働くPHYTOCHROME INTERACTING FACTOR
(PIF) というbHLH型転写因子の機能を阻害することで遺伝子発現を制御することが知られ
ている。これまでに、赤色光/遠赤色光可逆的な応答やフィトクロム遺伝子の存在は多くの 植物種で報告されているが、シロイヌナズナ以外の植物種におけるフィトクロムシグナル 伝達の分子機構に関してはほとんど知見がない。そこで本研究では基部陸上植物である苔 類ゼニゴケを用いて、フィトクロム自身の性質の解析とともに、そのシグナル伝達機構につ いて解析を行った。
まず、ゼニゴケにおけるフィトクロム遺伝子の探索を行い、1 分子種のフィトクロム
(MpPHY) をもつことを明らかにした。Mpphyの光に対する安定性および細胞内局在の解析
から、安定性は被子植物におけるⅡ型フィトクロムに、細胞内局在は被子植物におけるⅠ型 フィトクロムに類似した性質をもつことを明らかにした。野生型やMpPHY過剰発現株では 無性芽の発芽や、LHCBおよびPORといった光誘導性遺伝子の発現が赤色光/遠赤色光可逆 的に制御されたのに対し、恒常的に活性型になると予想された点変異を導入したMpphyY241H を発現する形質転換体ではそれらの応答の光依存性が喪失した。これらの結果から、Mpphy はシロイヌナズナのフィトクロムと同様に赤色光依存的に遺伝子発現を制御することで生 理応答を制御することが示唆された。
次に、Mpphyを介した遺伝子発現制御に関与する候補遺伝子として PIF 相同遺伝子の探 索を行い、ゼニゴケが1分子種のPIF (MpPIF) をもつことを明らかにした。相同組み換え によるMpPIFノックアウト株 (MppifKO) を用いた解析から、MppifKOではMpphyY241H発現 株と同様に無性芽の発芽や光誘導性遺伝子の発現の光依存性が喪失することを見出し、
MpPIF が暗黒下で無性芽の発芽や光誘導性遺伝子の発現を抑制するフィトクロムシグナル
伝達の抑制因子であることを明らかにした。また生化学的な解析から、MpPIF が暗黒下で 蓄積し、赤色光依存的に分解されること、MpphyY241H発現株では暗黒下でもMpPIFが蓄積 しないこと、活性型MpphyとAPAモチーフを介して光依存的に相互作用することを明らか にした。これらの結果から、MpPIFは赤色光依存的に活性型Mpphyと相互作用することで 分解されることが示唆された。以上の結果から、基部陸上植物であるゼニゴケがフィトクロ ムとPIF 転写因子との光依存的な相互作用および PIF 転写因子の分解による遺伝子発現制 御というシロイヌナズナと共通したフィトクロムシグナル伝達機構をもつことを明らかに した。本研究の結果、PIFを介した転写制御による赤色光シグナル伝達機構は陸上植物進化 の基部に位置する苔類の時点で既に獲得された形質であることが示唆され、陸上植物の共 通祖先において獲得されていた可能性が考えられた。
3
略語表
PHY PHYTOCHROME
PIF PHYTOCHROME INTERACTING FACTOR
LHCB LIGHT-HARVESTING CHLOROPHYLL A/B-BINDING
POR PROTOCHLOROPHYLLIDE OXIDOREDUCTASE
FHY1 FAR-RED ELONGATED HYPOCOTYL1
FHL FHY1-LIKE
TOC1 TIMING OF CAB EXPRESSION 1
PIL PIF3-LIKE
COP1 CONSTITUTIVE PHOTOMORPHOGENIC 1
SPA SUPPRESSOR OF PHYA-105
HY5 ELONGATED HYPOCOTYL5
EF1 ELONGATION FACTOR 1
LRL1 LJRHL1-LIKE 1
APT ADENINE PHOSPHORIBOSYL TRANSFERASE
SPT SPATULA
ALC ALCATRAZ
UGPase UDP-glucose pyrophosphorylase
SCF Suppressor of kinetochore protein1, Cullin, Ring-Box1 CRISPR clustered regularly interspaced short palindromic repeats
CAS9 CRISPR associated proteins
R red light
FR far-red light
Pr red light absorbing
Pfr far-red light absorbing
DNA deoxyribonucleic acid
cDNA complementary DNA
RACE rapid amplification of cDNA ends
RNA ribonucleic acid
mRNA messenger RNA
PCR polymerase chain reaction
qRT-PCR quantitative reverse transcription PCR
APA active phytochrome A-binding
APB active phytochrome B-binding
PB phytochromobilin
4
VLFR very low fluence response
LFR low fluence response
HIR high irradiance response
bHLH basic helix-loop-helix
CaMV cauliflower mosaic virus
CBD chitin binding domain
EST expressed-sequence tag
EDTA ethylenediaminetetraacetic acid
DMSO dimethyl sulfoxide
BSA bovine serum albumin
PBS phosphate buffered saline
DTT dithiothreitol
IPTG isopropyl β-D-thiogalactopyranoside
PMSF phenylmethylsulfonyl fluoride
SDS sodium dodecyl sulfate
PAGE polyacrylamide gel electrophoresis
PVDF polyvinylidene difluoride
MUSCLE Multiple Sequence Comparison by Log-Expectation
JTT Jones-Thornton-Taylor
JGI Joint Genome Institute
BLAST Basic Local Alignment Search Tool
LED light emitting diode
5
序論
植物の環境応答
移動能をもたない植物にとって、周囲の環境に自身を適応させることは生存する上で必 要不可欠である。そのため、植物は光や温度、重力などを受容し、それらに従って最適な形 態形成や運動応答を行えるように進化を遂げてきた。様々な環境情報の中でも、「光」は光 合成のエネルギー源であると同時に、日長や周囲の植物体との位置関係を知るための環境 情報としても重要な因子であり、生活環を通して植物の生長、発生に多大な影響を与える因 子の1つである。植物は光の強度、波長、方向、照射時間などを正確に感知し、自らの生体 情報に変換することで、種子の発芽、胚軸伸長阻害や子葉の展開といった光形態形成、成長 相転換といった組織レベルの形態形成から、光屈性や葉緑体定位運動といった細胞レベル の運動応答まで、様々な生理応答を示すことが知られている。
光受容体フィトクロム
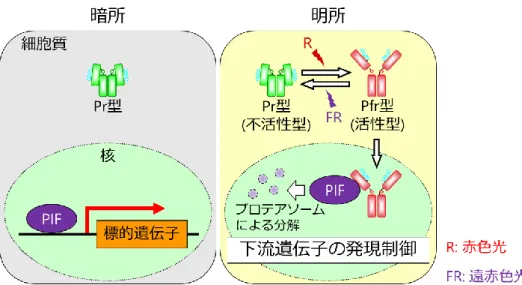
植物は光の情報を正確に感知するため、光の波長ごとに異なる光受容体をもっており、フ ィトクロム、クリプトクロム、フォトトロピン、ZEITLUPE/FLAVIN-BINDING, KELCH REPEAT, F-BOX1/LOV KELCH PROTEIN2、UV RESISTANCE LOCUS8がこれまでに同定さ れている (Kami et al., 2010; Ito et al., 2012; Jenkins, 2014; Christie et al., 2015)。これらの光受容 体の中で、フィトクロムは赤色光および遠赤色光を受容する唯一の光受容体であり、その大 きな特徴として赤色光を受容することで生理学的に不活性型であるPr型から活性型である Pfr型へ、遠赤色光を受容することでPfr型からPr型へ光可逆的に構造変化する「光可逆性」
をもつ (図1)。この性質によって、フィトクロムは光シグナルのON/OFFを素早く切り替え るスイッチとして機能することや、赤色光と遠赤色光の比を感知することが可能となって いる (Mancinelli, 1994)。また、Pr型がPfr型よりも熱力学的に安定なため、暗黒下では徐々 にPfr型からPr型へと変換される「暗反転」がみられる (Mancinelli, 1994)。
図1 フィトクロムの光可逆的な構造変化の模式図
6
フィトクロムの分子構造は大きく N 末端領域と C 末端領域に分けることができる (Nagatani, 2010)。N 末 端 領 域 に は 発 色 団 結 合 ド メ イ ン で あ る GAF (cGMP-specific phosphodiesterases, adenylyl cyclases, FhlA) ドメインや、Pfr型の安定性に関与すると考えら れるPHY (phytochrome-specific GAF-related ) ドメインなどが存在し、C末端領域にはPAS (PER, ARNT, SIM) ドメインやヒスチジンキナーゼ様ドメイン (histidine-kinase-related
domain; HKRD) が存在する。PASドメインやHKRDが存在することから、C末端領域がシ
グナル伝達に重要であると長年考えられてきたが、N末端領域にダイマー形成能と核移行 シグナルを付与したN末端phytochrome B (phyB) がphyB変異体を相補し、同様にN末端
phytochrome A (phyA) がphyA変異体を部分的に相補したことから、N末端領域がシグナル
伝達において重要であることが示されている (Matsushita et al., 2003; Oka et al., 2004; Mateos
et al., 2006)。このことから、C末端領域には二量体化に必要なドメインや核移行シグナル
配列が存在すると考えられている (Muller et al., 2009)。また、N末端領域とC末端領域の 結合が赤色光依存的に弱まることから、不活性型ではN末端領域がC末端領域に結合して 核移行能を抑制しており、活性型になって構造が変化することで抑制が解除されて核内に 移行する可能性が考えられている (Chen et al., 2005)。
フィトクロム発色団と発色団生合成酵素遺伝子
フィトクロムアポタンパク質が光を吸収し得る機能的なタンパク質となるためには、発 色団と結合することが不可欠である。フィトクロムの発色団は開環状テトラピロール化合 物であり、陸上植物ではフィトクロモビリン (PB) を発色団として持っていることが知ら れている。植物のフィトクロムホロタンパク質は GAFドメイン内に存在するシステイン残 基に発色団がチオエーテル結合によって共有結合している (Lagarias and Lagarias, 1989)。ま た、発色団が結合するシステイン残基を欠失させても僅かに光受容能がみられることから、
非共有結合的に発色団が結合できる可能性も示唆されている (Oka et al., 2011)。PBの生合 成酵素遺伝子はシロイヌナズナ (Arabidopsis thaliana) において白色光下で胚軸が徒長する hy1, hy2変異体から単離同定された (Muramoto et al., 1999; Kohchi et al., 2001)。
フィトクロムの機能による分類
被子植物のフィトクロムは暗所で高レベルに蓄積し、明所ですみやかに分解される光に 対して不安定なⅠ型フィトクロムと、明暗に大きく左右されずに存在する光に対して安定 なⅡ型フィトクロムに大別される。多くの植物種では、フィトクロムは遺伝子ファミリーを 形成しており、シロイヌナズナにおいてはphyA~Eの5分子種が同定されている (Clack et
al., 1994)。5分子種のフィトクロムのうち、phyAはⅠ型に、phyB~EはⅡ型フィトクロムに
分類される。この光に対する安定性に起因して、暗所ではphyAが、明所ではphyBが最も 多く蓄積することが示されている (Sharrock and Clack, 2002)。そのため、暗所における微弱 な光の受容には主にphyAが、明所における光の受容には主にphyBが機能していると考え
7
られている。イネ (Oryza sativa) やトウモロコシ (Zea mays) といった単子葉植物において も光に対して不安定なphyAと光に対して安定なphyBの存在が報告されており、分解の速 度は植物種間でやや異なっているものの I 型フィトクロムは赤色光照射後 2~4 時間以内に 分解がみられる (Reddy and Sharma, 1998; Xie et al., 2014)。
シロイヌナズナのフィトクロムは光依存的に核内に移行して機能することが知られてい る (Kircher et al., 1999; Yamaguchi et al., 1999; Huq et al., 2003; Klose et al., 2015)。この核移行 に関する分子機構もⅠ型とⅡ型のフィトクロムで異なっていることが明らかとなっている (Nagatani, 2004; Fankhauser and Chen, 2008)。phyAの核移行は赤色光または遠赤色光のいず れの照射によっても観察され、その核移行には FAR-RED ELONGATED HYPOCOTYL1 (FHY1)、FHY1-LIKE (FHL) といった因子を必要とすることが報告されている (Hiltbrunner et al., 2006; Genoud et al., 2008; Rausenberger et al., 2011)。一方で、phyBは赤色光下でのみ効 率的に核内に移行することが報告されており (Yamaguchi et al., 1999; Oka et al., 2012)、核移 行にFHY1やFHLを必要とせず、自身の核移行シグナルまたはフィトクロムシグナルに関 与する転写因子との結合を介して核内に移行しているのではないかと考えられている (Chen et al., 2005; Pfeiffer et al., 2012)。核に移行する速度もⅠ型とⅡ型のフィトクロムで異な っていることが知られており、phyA-GFP が光照射後 15分以内と短時間で核移行が観察さ れるのに対し (Kim et al., 2000)、phyB-GFPは赤色光照射後2時間程度で核内に観察され始 めることが報告されている (Gil et al., 2000)。
フィトクロムに制御される生理応答
フィトクロムによって制御される生理応答は、その光量依存性や光可逆性、相反則の有無 によって低光量反応 (low fluence response; LFR)、超低光量反応 (very low fluence response;
VLFR)、高照射反応 (high irradiance response; HIR) の3種類に分類される (Mancinelli, 1994;
Casal et al., 1998)。これらの応答の中で、LFRが最も広く知られており、典型的な赤色光/遠
赤色光可逆性や相反則を示す応答である。VLFRは波長の依存性がなく、非常に低い光量 (<
1 nmol m-2) のあらゆる波長の可視光で誘導され、光可逆性を示さないことから非常に少数
の Pfr 型のフィトクロムが応答を引き起こすのに十分である可能性が考えられている
(Shinomura et al., 1996)。HIRは長時間の連続した光照射が応答に必要であり、基本的には光
可逆性や相反則がみられないが、限られた厳密な実験条件下では相反則や光可逆性を示す ことが報告されている (Shinomura et al., 2000)。シロイヌナズナではⅠ型フィトクロムであ るphyAがVLFRと遠赤色光高照射反応 (FR-HIR) を制御し (Nagatani et al., 1993; Parks and Quail, 1993; Whitelam et al., 1993; Shinomura et al., 1996)、Ⅱ型フィトクロムであるphyBが LFRと赤色光高照射反応 (R-HIR) を制御している (Mancinelli, 1994; Shinomura et al., 1996)。
フィトクロム相互作用因子とシグナル伝達機構
フィトクロムが受容した光情報を下流因子に伝達するには何らかのタンパク質との相互
8
作用が必要であることが考えられたため、これまでに多くの相互作用因子の探索が行われ ている (Bae and Choi, 2008)。それらの相互作用因子の中には、phyAおよびphyBの両方に 結合する因子だけでなく、上述したFHY1やFHLのように一方のみに結合する因子も存在 する。さらに、それらの中でも赤色光依存的に Pfr 型のフィトクロムに結合する因子と、
Pfr/Pr型の区別なく結合する因子が存在する。このようなフィトクロム相互作用因子の中で、
代表的な因子として PHYTOCHROME INTERACTING FACTOR (PIF) ファミリーが挙げら れる (Leivar and Monte, 2014)。PIFファミリーはbHLH subfamily VII(a+b) に属するbHLH型 の転写因子であり (Pires and Dolan, 2010)、phyBのC末端領域を用いたYeast two-hybrid法 によって最初にPIF3が単離された (Ni et al., 1998)。その後、phyBのN末端領域や全長を用 いてPIF3 との相互作用が見直され、PIF3が赤色光依存的にPfr 型のphyBに結合すること や、その相互作用が光可逆的であることが明らかとなった (Ni et al., 1999)。さらに、赤色光 下で胚軸が短くなるsrl2 (short under red-light 2) 変異体の原因遺伝子としてPIF4が単離さ れ (Huq and Quail, 2002)、TIMING OF CAB EXPRESSION 1 (TOC1) と相互作用する因子の 中でPIF3と相同性の高い遺伝子としてPIF3-LIKE 1 (PIL1)、PIL2、PIL5、PIL6が単離され た (Yamashino et al., 2003)。その後、PIL遺伝子を含むbHLHサブファミリーとphyBの相互 作用解析が行われ、相互作用が確認された因子がそれぞれPIF1/PIL5、PIF5/PIL6、PIF6/PIL2 と名付けられた (Huq et al., 2004; Khanna et al., 2004)。また、その後bHLH72がPIF7として (Leivar et al., 2008)、bHLH16がPIF8として新たに同定されている (Leivar and Quail, 2011)。
これらのPIFファミリーはN末端側にAPB (Active phytochrome B binding) モチーフと呼ば れる領域が存在しており、その領域に点変異を入れるとphyBとの相互作用が阻害されるこ とから、活性型フィトクロムとの相互作用部位であると考えられている (Khanna et al., 2004)。
さらに、イネにおけるPIFファミリーにもN 末端側に保存性の高い領域が存在するが、シ ロイヌナズナのPIFファミリーのもつAPBモチーフとの相同性は低く、生物種間のAPBモ チーフの保存性は低いと考えられる (Nakamura et al., 2007)。また、PIF ファミリーの中で PIF1、PIF3の 2分子種は Pfr 型の phyA とも相互作用することが示されており (Zhu et al., 2000; Shimizu-Sato et al., 2002)、APA (Active phytochrome A binding) モチーフが存在すると考 えられているが、相互作用に必要な領域は未だ明確ではない (Al-Sady et al., 2006; Shen et al.,
2008)。PIF ファミリーは暗黒下や赤色光/遠赤色光比が低い環境下において多数の下流遺伝
子の発現制御を行うことで、フィトクロムシグナル伝達の負の制御因子として機能してい る。光を受容して活性型となったフィトクロムはPIFと相互作用し、PIFのリン酸化とそれ に続くユビキチン・プロテアソーム系による分解を引き起こす (図2)。この光依存的な分解 によるPIF蓄積量の変化が、種子の発芽、芽生えの脱黄化、胚軸伸長阻害、避陰応答といっ た生理応答を引き起こすことが明らかとなっている (Leivar and Monte, 2014)。
9
図2 PIFを介したフィトクロムシグナル伝達機構 (Castillon et al., 2007を参考に作製)
PIF以外の主要なフィトクロムシグナル伝達因子として、可視光のシグナル伝達において 中心的な抑制因子として機能するCONSTITUTIVE PHOTOMORPHOGENIC 1 (COP1) およ びSUPPRESSOR OF PHYA-105 (SPA) ファミリーを介した経路が知られている (Huang et al., 2014)。暗黒下においてCOP1/SPA複合体はELONGATED HYPOCOTYL5 (HY5) のような光 形態形成の促進因子を分解することで生育に好ましくない環境下での光形態形成を抑制し ている。近年の解析から、光を受容して活性型となったフィトクロムが SPA ファミリーと 光依存的に相互作用することで COP1/SPA 複合体の解離を促進することが報告されている (Lu et al., 2015; Sheerin et al., 2015)。
上記のようにシロイヌナズナにおける知見は多く存在しているが、フィトクロム依存的 な遺伝子発現制御は様々な被子植物や (Sawers et al., 2002; Takano et al., 2005; Kebrom et al., 2006; Takano et al., 2009; Kebrom et al., 2010)、裸子植物 (Alosi and Neale, 1992; Christensen et
al., 2002) においても報告されているのに対し、シロイヌナズナ以外の植物種におけるフィ
トクロムシグナルの分子機構に関する知見はほとんど存在しない。
基部植物のフィトクロムシグナル伝達機構
シダ植物やコケ植物蘚類といった基部植物においてもフィトクロムに関する知見が報告 されている (Suetsugu and Wada, 2007; Rosler et al., 2010)。シダ植物であるホウライシダ
(Adiantum capillus-veneris) では原糸体光屈性、葉緑体光定位運動など、被子植物では主にフ
ォトトロピンによって制御される応答が赤色光によって制御されることが報告されている (Rosler et al., 2010; Jaedicke et al., 2012)。コケ植物では、蘚類に属するヒメツリガネゴケ (Physcomitrella patens) やヤノウエノアカゴケ (Ceratodon purpureus) においてフィトクロム に関する知見が報告されている。ヒメツリガネゴケにはPpPHY1~4および PpPHY5a~5cの 7分子種が、ヤノウエノアカゴケにはCpPHY1~4の4分子種が存在し、原糸体光屈性や分岐
10
の形成、葉緑体光定位運動を制御していることが報告されている (Pasentsis et al., 1998;
Mittmann et al., 2004; Uenaka et al., 2005; Mittmann et al., 2009)。これらのフィトクロムの中で、
Cpphy1はC末端にHKRDの代わりにセリン/スレオニンキナーゼと高い相同性を示す配列
をもった特殊な分子である (Thummler et al., 1992)。Cpphy1以外は典型的な陸上植物フィト クロムと共通したドメイン構造ではあるが、それらのエキソン・イントロン構造は被子植物 のフィトクロムと異なっていることが知られている (Mittmann et al., 2009)。また、Ppphy1~4 はプロトプラストを用いた一過的発現系において細胞質に局在することが示されており、
核移行シグナルを付加したPpphy1 およびPpphy2 が葉緑体光定位運動に機能的でないこと から、上記の応答はPpphyの細胞質における機能であることが示唆されている (Uenaka and Kadota, 2007)。このように基部植物のフィトクロムに関する知見は細胞質における機能が多 く存在するが、近年になって核における機能がいくつか報告されている。ホウライシダでは、
マイクロビームによって核内のフィトクロムを選択的に活性化することによって胞子発芽 を誘導できることが報告されている (Tsuboi et al., 2012)。ヒメツリガネゴケでは、連続遠赤 色光下においてPpPHY1およびPpPHY3が細胞質から核内へと移行し、FR-HIR様の応答を 制御することが報告されている (Possart and Hiltbrunner, 2013)。しかしながら、基部植物に おけるフィトクロムシグナル伝達の分子機構や遺伝子発現制御の有無は未だ直接的に示さ れていない。近年の系統解析の結果から、被子植物や蘚類を始めとする多くの植物種では種 特異的な遺伝子重複によってフィトクロムの分子種が増加していることが示唆されている (Li et al., 2015)。一方で、苔類やツノゴケ類では遺伝子重複がみられず、これらの種におけ る 1 分子種のフィトクロムがもつ祖先的な性質を明らかにすることが、フィトクロム遺伝 子の分子進化を考えるうえで重要であると期待されている (Li et al., 2015)。
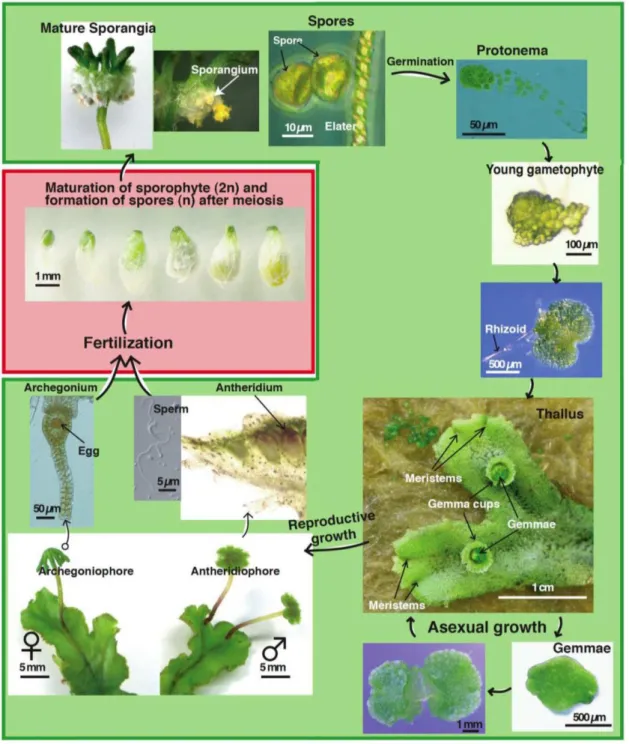
苔類ゼニゴケ (Marchantia polymorpha)
ゼニゴケは苔類に属するコケ植物で、その生活環の大部分を半数体である配偶体で過ご すことが知られている (図3)。葉状体 (Thallus) の背側には葉緑体に富んだ同化糸が存在す る気室が存在し、無性生殖器官である杯状体 (Gemma cup) や無性芽 (Gemma) が形成され る。葉状体の腹側には地表に密着するための仮根 (Rhizoid) や腹鱗片が形成される。ゼニゴ ケは雌雄異株であり、造精器 (Antheridium) または造卵器 (Archegonium) をそれぞれの株が 形成することで有性生殖を行うことが知られている。ゼニゴケは陸上植物進化の基部に位 置すると考えられている苔類に属することから (Qiu et al., 2006)、植物の進化を考える上で も非常に重要なモデルであり、被子植物に比べて体制が単純かつ遺伝子の冗長性が低いた め、陸上植物の祖先的なシグナル伝達機構を備えている可能性が考えられる。従って、ゼニ ゴケを用いた研究により、陸上植物に普遍的なシグナル伝達機構の根幹となる分子機構を 理解できると期待される。また、ゼニゴケは生活環の大半が半数体であるため、変異体・形 質転換体の表現型が当代で観察できる、無性生殖によってクローンを簡単に大量に増殖で きるといった生物学的な利点を備えているだけでなく、Joint Genome Institute (JGI) による
11
ゲノム解読によりシグナル伝達に関与する遺伝子の冗長性が低いことが明らかとねってい ること (未発表データ)、高効率の形質転換法や (Ishizaki et al., 2008)、相同組み換えによる ジーンターゲティング(Ishizaki et al., 2013)、CRISPR/CAS9を用いたゲノム編集といった様々 な遺伝学的ツールが整備されており (Sugano et al., 2014)、基部陸上植物の新しいモデル植物 の1つとなっている。
図3 ゼニゴケの生活環
12
過去に、ゼニゴケの赤色光応答がいくつか報告されている。明暗周期で培養しているゼニ ゴケの明期終了時に遠赤色光をパルス照射すると、葉状体の垂直方向への成長やクロロフ ィル量の減少が観察され、これらの応答は遠赤色光の後に赤色光を照射することで打ち消 されたことからフィトクロムによる制御が示唆されている (Fredericq and De Greef, 1966;
Hartmann and Jenkins, 1984)。また、赤色光および遠赤色光のパルス照射を用いた解析から、
葉状体の老化や無性芽の発芽が赤色光/遠赤色光可逆性を示すことからフィトクロムによっ て制御されることが示唆されている (Ninnemann and Halbsguth, 1965; De Greef et al., 1971;
Otto and Halbsguth, 1976)。さらに、ゼニゴケのフィトクロムが葉状体の切断片からの再生過 程において、細胞周期への再進入や再生芽の形態形成を制御することが近年報告されてい る (Nishihama et al., 2015)。
本研究では、苔類ゼニゴケをモデルとしてフィトクロムの祖先的な性質を解析するとと もに、そのシグナル伝達機構について解析することで、陸上植物のフィトクロムシグナル伝 達機構の進化を解明することを目的に研究を行い、ゼニゴケがそれぞれ 1 分子種のフィト クロムとPIF転写因子をもつことを明らかにした。また、生化学的、細胞生物学的な解析か ら、1分子種のフィトクロムが被子植物におけるI型およびⅡ型フィトクロムの特徴を併せ もつことを明らかにした。さらに、陸上植物進化の基部に位置する苔類ゼニゴケが、フィト クロムと PIF 転写因子の光依存的な相互作用および PIF の分解による遺伝子発現制御機構 をもつことを明らかとした。
13
結果
フィトクロム遺伝子の単離
ゼニゴケにおける赤色光シグナル伝達機構を明らかにするため、cDNAライブラリーおよ びゲノムライブラリーを用いて赤色光受容体であるフィトクロム遺伝子の探索を行った。
その結果、ゼニゴケはゲノム上にフィトクロム相同遺伝子 (MpPHY) を 1 分子種のみもつ ことが明らかとなった。得られたMpPHYのゲノム配列を調べたところ、陸上植物の典型的 なフィトクロム遺伝子とイントロン挿入位置が完全に保存されていた (図4)。次に、アミノ 酸配列を比較したところ、陸上植物の典型的なフィトクロムと同じドメイン構造をもつこ とが明らかとなった (図4)。そこで、Mpphyの分光学的性質を明らかにするため、発色団と してPBの付加したMpphyのN 末端領域を大腸菌で発現、精製し、その吸収スペクトル を測定した。その結果、組み換えMpphyタンパク質は赤色光/遠赤色光可逆的な分光学的性 質を示した (図 5)。これらの結果から、Mpphy は陸上植物の典型的なフィトクロムと共通 した分光学的性質をもつ機能的なフィトクロムであることが示唆された。
14
図4 ゼニゴケおよびシロイヌナズナのフィトクロムのマルチプルアライメント
赤い矢じりはイントロン挿入位置を示す。赤いアスタリスクは発色団結合残基を、青いアス タリスクは光可逆的な構造変化に重要な残基を、マゼンタのアスタリスクはphyAにおける ユビキチン結合残基を示す。赤、青、マゼンタ、黄、緑、紫の下線はそれぞれ N-terminal extension,、PAS domain、GAF domain、PHY domain、PAS-related domain、histidine kinase-related domain (HKRD) を示す。
15 図5 組み換えMpphyタンパク質の分光学的性質
発色団としてPBを付加したN 末端Mpphyを大腸菌で発現・精製し、吸収スペクトルを 測定した。(A) Mpphy(N612) の吸収スペクトル。遠赤色光照射後の吸収スペクトル (青線) および赤色光照射後の吸収スペクトル (赤線) を示す。(B) 遠赤色光照射後の吸収スペクト ルから赤色光照射後の吸収スペクトルを引いた差スペクトルを示す。
Mpphyの光に対する安定性の解析
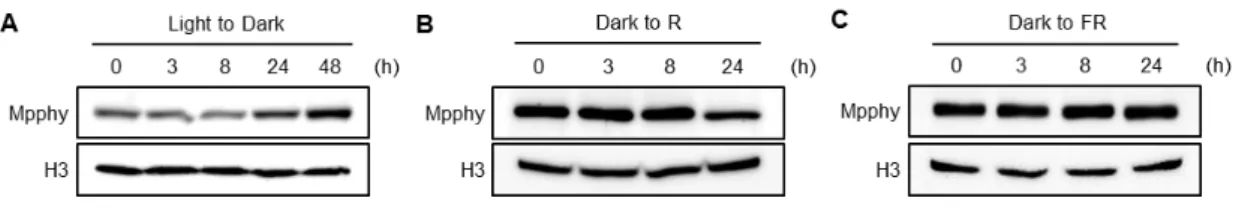
1分子種しかないMpphyが被子植物におけるI型フィトクロムまたはII型フィトクロム のどちらに近い性質をもつか明らかにするため、Mpphy の光に対する安定性を解析した。
連続白色光下で10日間培養した植物体を暗黒下に移し、Mpphyタンパク質の蓄積量の変化 を継時的に解析したところ、Mpphy タンパク質の蓄積量は暗処理後わずかに増加した (図
6A)。次に、2日間の暗処理によってMpphyを蓄積させた植物体を連続赤色光または連続遠
赤色光下に移し、Mpphyタンパク質蓄積量の継時的な変化を調べた。その結果、Mpphyは 赤色光照射後24時間でわずかに減少するものの、赤色光照射後8時間以内または遠赤色光 下では蓄積量の変化が全くみられなかった (図6B, C)。被子植物のI型フィトクロムは赤色 光照射後 2~4 時間以内に著しい蓄積量の減少を示すことから (Reddy and Sharma, 1998;
Sharrock and Clack, 2002; Xie et al., 2014)、MpphyはII型フィトクロムと類似した光安定性を 示すことが明らかとなった。
16
図6 Mpphyタンパク質の光に対する安定性の評価
抗 Mpphy 抗体を用いて野生型における内性 Mpphy タンパク質蓄積量の光に対する挙動を
解析した。連続白色光下で10日間培養した植物体を次に示す光条件に移して解析を行った。
(A) 暗黒下、(B) 48時間の暗処理+連続赤色光 (30 µmol photons m-2 s-1)、(C) 48時間の暗処理 +連続遠赤色光 (30 µmol photons m-2 s-1)。抗Histone H3抗体の結果をコントロールとして示 した。
Mpphyの細胞内局在解析
シロイヌナズナにおいて、I型フィトクロムであるphyAは赤色光および遠赤色光のいず れの照射においても細胞質から核内への移行がみられるのに対し、II型フィトクロムである phyB は赤色光照射下でのみ効率的な核移行がみられることが知られている (Kircher et al., 1999; Yamaguchi et al., 1999)。そこで、Mpphyの細胞内局在パターンがI型およびII型のど ちらに近いのか明らかにするため、MpphyのC末端に蛍光タンパク質Citrineを付加した融 合タンパク質を構成的発現プロモーターである CaMV 35S プロモーター (pro35S) の制御下 で発現するコンストラクトを導入した形質転換体 (pro35S:MpPHY-Citrine) を作出した。
pro35S:MpPHY-Citrine株の無性芽を連続白色光下で1日間培養後、6日間の暗処理を施し、
暗処理直後または暗処理後に赤色光または遠赤色光を 10 分間照射して蛍光観察を行った。
その結果、Mpphy-Citrine の蛍光は暗黒下では細胞の周縁部でのみ観察されたのに対し (図
7)、赤色光または遠赤色光照射後には主に核での蛍光が観察された (図7)。
17 図7 Mpphy-Citrineの細胞内局在解析
pro35S:MpPHY-Citrine株を連続白色光下で1日間培養後、暗黒下に6日間移したサンプル (上 段) および暗処理後に連続赤色光 (30 µmol photons m-2 s-1; 中段) または連続遠赤色光 (30 µmol photons m-2 s-1; 下段) を10分間照射したサンプルの蛍光観察。スケールバーは20 µm。
また、光可逆的な構造変化に重要なチロシン残基 (Tyr241) をヒスチジン残基に置換し、
恒常的に活性型として機能することが予想される1アミノ酸置換体 (MpphyY241H) を用いて 同様の解析を行ったところ、MpphyY241H-Citrine は暗黒下においても核での蛍光が観察され た (図8)。組み換えMpphyY241Hタンパク質はphyBY276Hと同様に光可逆的な吸光スペクトル の変化が完全に喪失したことから (図 9)、phyBY276Hと同様に生体内で活性型として機能す る可能性が考えられた (Su and Lagarias, 2007)。これらの結果から、Mpphyは光照射によっ て活性型になると核内へと移行することが示唆された。
18 図8 MpphyY241H-Citrineの細胞内局在解析
pro35S:MpPHYY241H-Citrine株を連続白色光下で1日間培養後、暗黒下に6日間移したサンプ ル (上段) および暗処理後に連続赤色光 (30 µmol photons m-2 s-1; 中段) または連続遠赤色光 (30 µmol photons m-2 s-1; 下段) を10分間照射したサンプルの蛍光観察。スケールバーは20 µm。
図9 組み換えMpphyY241Hタンパク質の分光学的性質
発色団としてPBを付加したN 末端MpphyY241Hを大腸菌で発現・精製し、吸収スペクト ルを測定した。MpphyY241H(N612) の吸収スペクトル。遠赤色光照射後の吸収スペクトル (青 線) および赤色光照射後の吸収スペクトル (赤線) を示す。(B) 遠赤色光照射後の吸収スペ クトルから赤色光照射後の吸収スペクトルを引いた差スペクトルを示す。
19
光依存的なMpphyの細胞内局在変化をさらに確認するため、細胞分画による生化学的手 法によって内性Mpphyタンパク質の細胞内局在を解析した。連続白色光下で10日間培養し た植物体に対して2日間の暗処理を施し、赤色光または遠赤色光を照射して10分および60 分後のサンプルについて細胞分画を行い、ウエスタンブロットによって細胞内局在を解析 した。その結果、蛍光観察の結果と一致して赤色光または遠赤色光照射のいずれにおいても 10分後に核内での Mpphy 蓄積量が増加した (図 10)。しかし、光照射に対する迅速な核移 行は蛍光観察と一致していたが、細胞分画の結果から大部分のMpphyタンパク質は光照射 後も細胞質に残っていることが明らかとなった (図10)。さらに、遠赤色光照射サンプルの 核画分において移動度が変化したバンドが確認されたことから (図 10B)、遠赤色光照射下 での核内のフィトクロムは何らかの翻訳後修飾を受けることが明らかとなった。以上の結 果から、Mpphy は赤色光および遠赤色光のいずれの照射でも核内に移行するという被子植 物のI型フィトクロムと同様の細胞内局在パターンを示すことが明らかとなった。
図10 細胞分画による内性Mpphyタンパク質の細胞内局在解析
細胞分画およびウエスタンブロットによる細胞内局在解析。連続白色光下で10日間培養し た植物体に48時間の暗処理を施した後、連続赤色光 (30 µmol photons m-2 s-1; A) または連 続遠赤色光 (30 µmol photons m-2 s-1; B) 照射下に移し、図に示した時間でサンプリングを行 い、細胞質画分 (Nucleus-depleted) および核画分 (Nucleus-enriched) を用いてウエスタンブ ロットを行った。抗UGPase 抗体を細胞質マーカーとして、抗 Histone H3抗体を核マーカ ーとして用いた。
20
Mpphyが制御する生理応答の解析
過去に、ゼニゴケの無性芽が赤色光依存的に発芽し、遠赤色光によって発芽が抑制される ことから、無性芽の発芽がフィトクロムによって制御されることが示唆されている (Otto
and Halbsguth, 1976)。そこで、野生型の MpPHY 遺伝子を構成的発現プロモーターである
ELONGATION FACTOR 1 (MpEF1) プロモーターの制御下で発現するコンストラクトを導
入した MpPHY 過剰発現株 (proMpEF1:MpPHY) および恒常的に活性型として機能すること
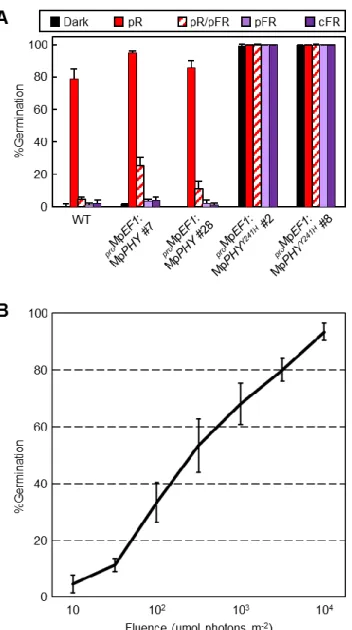
が予想される点変異をもつMpPHYY241HをMpEF1プロモーターの制御下で発現するコンス トラクトを導入した恒常活性型MpPHY発現株 (proMpEF1:MpPHYY241H) を用いて、Mpphyが 無性芽の発芽に関与するかどうかを解析した。緑色安全光下で無性芽を寒天培地に播き、暗 黒下で1日間吸水させた後、様々な光処理を施して発芽した個体数をカウントした。その結 果、野生型の無性芽では過去の報告と一致して赤色光のパルス照射によって発芽が誘導さ れ、赤色光に続く遠赤色光によって発芽が阻害される赤色光/遠赤色光可逆性が見られた
(図11A)。また、赤色光によって発芽が誘導された個体数は光量依存的に増加した (図11B)。
さらに、遠赤色光のパルス照射や連続遠赤色光は無性芽の発芽に有効でないことが明らか となった (図11A)。これらの結果から、ゼニゴケの無性芽の発芽はVLFRやFR-HIRではな く、典型的な光可逆性を示すLFR に分類されると考えられた。proMpEF1:MpPHY では赤色 光によって発芽誘導される個体数が野生型と比較して僅かに増加したが、遠赤色光による 発芽の阻害は野生型と同程度にみられた (図 11A)。対照的に、proMpEF1:MpPHYY241Hでは、
ほぼ全ての個体が暗黒下においても発芽し、遠赤色光による発芽の阻害が全くみられなか った (図11A)。qRT-PCRによってproMpEF1:MpPHYY241HおよびproMpEF1:MpPHYにおける
MpPHY発現量を調べたところ、各形質転換体におけるMpPHY発現量にほとんど差がみら
れなかったことから (図12)、proMpEF1:MpPHYY241Hの表現型は発現量の差ではなく、アミノ 酸置換の影響であることが示された。これらの結果から、過去の観察結果と一致して (Nishihama et al., 2015)、MpphyY241Hが生体内で恒常的に活性型として機能することが強く支 持された。
21
図11 無性芽の発芽の光質依存性と光量依存性
(A) 野生型 (WT)、proMpEF1:MpPHYおよびproMpEF1:MpPHYY241H株の様々な光環境下にお ける発芽率の測定。暗黒下で吸水させた無性芽を、そのまま暗黒下に維持したサンプル (Dark)、赤色光 (3000 µmol photons m-2) をパルス照射した後で暗黒下に戻したサンプル (pR)、
赤色光 (3000 µmol photons m-2) に続いて遠赤色光 (9000 µmol photons m-2) をパルス照射し た後で暗黒下に戻したサンプル (pR/pFR)、遠赤色光 (9000 µmol photons m-2) をパルス照射 したサンプル (pFR) および連続遠赤色光下 (30 µmol photons m-2 s-1) に移したサンプル
(cFR) の発芽した個体数の割合を示す。一度の解析に 50の無性芽を用いた。グラフは独立
した3回の実験における平均値を、エラーバーは標準偏差を示す。
(B) 野生型の無性芽を暗黒下で吸水させ、様々な強度の赤色光を5分間照射した後で暗黒下 に戻し、6日後に発芽率を測定した。一度の解析に50の無性芽を用いた。グラフは独立し た3回の実験における平均値を、エラーバーは標準偏差を示す。
22 図12 形質転換体におけるMpPHY発現量の比較
野生型 (WT)、proMpEF1:MpPHYおよびproMpEF1:MpPHYY241H株におけるMpPHY発現量の 比較。連続白色光下で10日間培養した植物体を解析に用いた。MpEF1を内部標準遺伝子と して用いた。グラフは独立した 3 回の実験における平均値を、エラーバーは標準誤差を示 す。
23
Mpphyは光依存的に下流遺伝子の発現を制御する
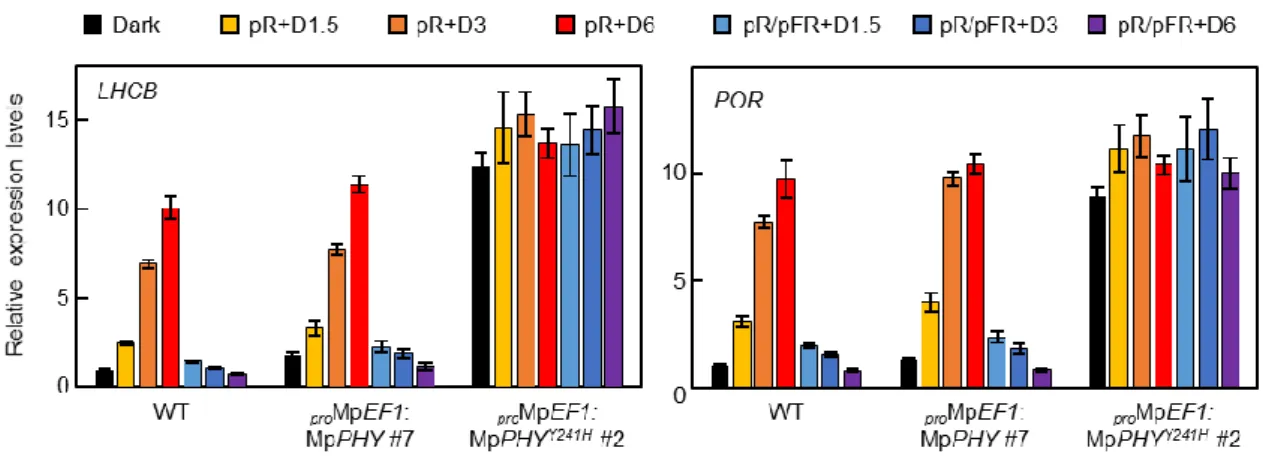
シロイヌナズナにおいて、フィトクロムは光に応答して多数の下流遺伝子の発現を制御 することが知られている (Quail, 2010)。そこで、Mpphyがシロイヌナズナのフィトクロムと 同様に光依存的な遺伝子発現を制御するかどうか調べた。連続白色光下で10日間培養した 植物体に対して48時間の暗処理を施した後、赤色光をパルス照射して暗黒下に戻したサン プルおよび赤色光パルスの直後に遠赤色光をパルス照射して暗黒下に戻したサンプルにつ い て LIGHT-HARVESTING CHLOROPHYLL A/B-BINDING PROTEIN (LHCB) お よ び PROTOCHLOROPHYLLIDE OXIDOREDUCTASE (POR) のmRNA蓄積量を解析した。野生型
およびproMpEF1:MpPHYではLHCBおよびPORのmRNA蓄積量は赤色光照射依存的に増
加し、赤色光照射に続く遠赤色光照射によってmRNA蓄積量の増加が阻害された (図13)。
対照的に、proMpEF1:MpPHYY241Hでは暗黒下においてもLHCBおよびPORのmRNA蓄積量 が増加しており、遠赤色光によるmRNA蓄積量の増加の阻害もみられなかった (図13)。こ れらの結果からMpphyは光誘導性遺伝子の発現を制御することが示唆された。
図13 光誘導性遺伝子の発現解析
野生型 (WT)、proMpEF1:MpPHYおよびproMpEF1:MpPHYY241H株におけるLHCBおよびPOR の発現量の比較。連続白色光下で7日間培養した植物体に48時間の暗処理を施し (Dark)、
赤色光 (3000 µmol photons m-2; pR) または赤色光 (3000 µmol photons m-2) に続いて遠赤色 光 (9000 µmol photons m-2; pR/pFR) をパルス照射して暗黒下に戻した後、図に示した時間で サンプリングを行った。MpEF1 および MpAPT を内部標準遺伝子として用いた。グラフは 独立した3回の実験における平均値を、エラーバーは標準誤差を示す。
24 ゼニゴケはPIF転写因子を1分子種のみもつ
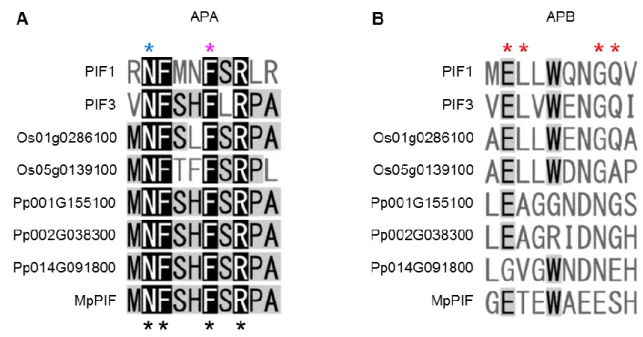
これまでの解析から、フィトクロム依存的な遺伝子発現制御がゼニゴケにおいてもみら れたため、次にMpphyシグナルに関与する転写因子を探索した。PIFファミリーはbHLH転 写因子のSubfamily VII(a+b) に属することが報告されている (Pires and Dolan, 2010)。ゼニゴ ケゲノムデータベースに対するBLASTおよび系統解析の結果から、ゼニゴケはシロイヌナ ズナやイネのPIF ファミリーと同じクレードに属する遺伝子を 1分子種のみもつことが明 らかとなった (図14)。この候補遺伝子 (MpPIF) はC末端領域にbHLHドメインが非常に 高度に保存されており、シロイヌナズナのPIF1およびPIF3に存在するphyAとの相互作用 に重要であると予想されるAPAモチーフをもつことが明らかとなった (図15)。しかしなが
ら、MpPIFはシロイヌナズナの全てのPIFファミリー転写因子に存在するphyBとの相互作
用に重要なAPBモチーフをもたないことが明らかとなった (図15)。また、ヒメツリガネゴ ケにおける PIF 相同遺伝子も APB モチーフをもたず、APA モチーフのみをもつことから
(図 16)、APB モチーフは陸上植物進化の過程で獲得された形質であることが示唆された。
また、BLAST解析の結果、MpPIFと最も類似性の高かったMapoly0502s0001は、根毛形成 に関与することが報告されているLRL1と同じクレード (Subfamily XI) に属した (Karas et
al., 2009)。Subfamily XIに属する遺伝子にはAPAモチーフやAPBモチーフがみつからなか
ったことから、ゼニゴケがPIF転写因子を1分子種のみもつことが強く支持された。
25 図14 ゼニゴケPIF相同遺伝子の探索
bHLHドメインのマルチプルアライメントを用いて最尤法により系統樹を作製した。外群と してクロレラおよびクラミドモナスの bHLH 配列を用いた。系統樹内の数字はブートスト ラップ確率を、バーはアミノ酸の置換率を示す。At, Arabidopsis thaliana; Os, Oryza sativa; Pp, Physcomitrella patens; Mp or Mapoly, Marchantia polymorpha; Cv, Chlorella vulgaris; Cr, Chlamydomonas reinhardtii。
26
図15 MpPIFとシロイヌナズナのPIFファミリーのマルチプルアライメント
赤とマゼンタの下線はそれぞれAPBモチーフとbHLHドメインを示す。青枠は推定APAモ チーフを示す。赤いアスタリスクはPIFファミリーとphyBの結合に重要な残基を示す。青 のアスタリスクはphyAとPIF1の結合に重要な残基を、マゼンタのアスタリスクはphyAと PIF3の結合に重要な残基を示す。
27
図16 APAモチーフおよびAPBモチーフのマルチプルアライメント
MpPIFとシロイヌナズナ、イネ、ヒメツリガネゴケのPIF相同遺伝子におけるAPAモチー
フ (A) およびAPBモチーフ (B) のアミノ酸配列の比較。青のアスタリスクはphyAとPIF1 の結合に重要な残基を、マゼンタのアスタリスクはphyA とPIF3の結合に重要な残基を示 す。黒のアスタリスクは高度に保存された残基を示す。赤のアスタリスクはPIFファミリー とphyBの結合に重要な残基を示す。
MpPIFはMpphyシグナル伝達の抑制因子である
MpPIFがMpphyシグナルに関与するかどうかを明らかにするため、相同組み換えによる
MpPIFノックアウト株 (MppifKO) を作出し (図17)、前述した無性芽の発芽の表現型につい
て解析した。その結果、MppifKOでは、proMpEF1:MpPHYY241Hと同様に光条件に関わらずほぼ 全ての無性芽が発芽した (図18A)。さらに、この発芽の光依存性の喪失は、MpPIF ゲノム 断片を導入した相補株 (proMpPIF:MpPIF/MppifKO) で野生型と同程度に回復したことから
(図18A)、MpPIFがMppifKOにおける発芽の光依存性の喪失の責任遺伝子であることが示さ
れた。
次に、Mpphy依存的な LHCBおよびPORの発現制御におけるMpPIFの機能について解 析した。その結果、MppifKOでは赤色光または遠赤色光パルス照射後に徐々に発現量が減少 するものの、暗黒下においてもLHCBおよびPORの発現量が野生型より増加していた (図
18B)。また、相補株では野生型と同程度までLHCBおよびPORの発現パターンが回復した
(図18B)。以上の結果から、MpPIFはMpphyシグナル伝達経路において無性芽の発芽およ
び光誘導性遺伝子の発現を負に制御することが示唆された。
28 図17 相同組み換えによるMppifKO株の作出
(A) 相同組み換えに用いたコンストラクトの模式図。矢印はジェノタイピングに用いたプラ イマー (a~c) を示す。(B) MppifKO株のジェノタイピング。(C) 野生型 (WT)、MppifKO、
proMpPIF:MpPIF/MppifKO株におけるMpPIF発現量の比較。MpEF1を内部標準遺伝子として 用いた。
29
図18 MppifKO株における無性芽の発芽の表現型と光誘導性遺伝子の発現量
(A) 野生型 (WT)、MppifKOおよびproMpPIF:MpPIF/MppifKO株の様々な光環境下における発 芽率の測定。光条件は図11Aと同様に行った。一度の解析に50の無性芽を用いた。グラフ は独立した3回の実験における平均値を、エラーバーは標準偏差を示す。(B) 野生型 (WT)、
MppifKOおよびproMpPIF:MpPIF/MppifKO株におけるLHCBおよびPORの発現量の比較。光 条件は図13と同様に行った。MpEF1およびMpAPTを内部標準遺伝子として用いた。グラ フは独立した3回の実験における平均値を、エラーバーは標準誤差を示す。
30
Mpphyは光依存的にMpPIFタンパク質蓄積量を制御する
MpPIFがMpphyシグナル伝達経路における遺伝子発現制御に関与することが示唆された
ため、光依存的な遺伝子発現制御の分子機構について解析した。シロイヌナズナにおいて、
活性型のフィトクロムが PIF ファミリー転写因子と相互作用することで、PIF のユビキチ ン・プロテアソーム系による分解を誘起することが知られている (Al-Sady et al., 2006; Shen et al., 2007; Shen et al., 2008)。そこで、まずMpPIFタンパク質蓄積量の光に対する挙動につ いて解析した。連続白色光下で10 日間培養した植物体に対して 48時間の暗処理を施した 後、連続赤色光あるいは連続遠赤色光下に移してMpPIFタンパク質蓄積量について解析し た。その結果、MpPIF タンパク質は連続白色光下の植物体ではほとんど検出されなかった のに対し、暗処理後24時間以降で顕著に蓄積量が増加した (図19A)。また、この蓄積量の 増加は、暗処理直前の遠赤色光照射によって促進された (図 19B)。これは遠赤色光によっ
て Mpphy が速やかに不活性型へと変化したことによるものと考えられ、活性型 Mpphy が
MpPIFタンパク質の蓄積を阻害する可能性が考えられた。暗黒下で蓄積したMpPIFタンパ
ク質は赤色光照射によって30分以内に速やかに分解されたのに対して (図19C)、遠赤色光 照射ではタンパク質蓄積量に変化がみられなかった (図 19D)。赤色光照射条件において
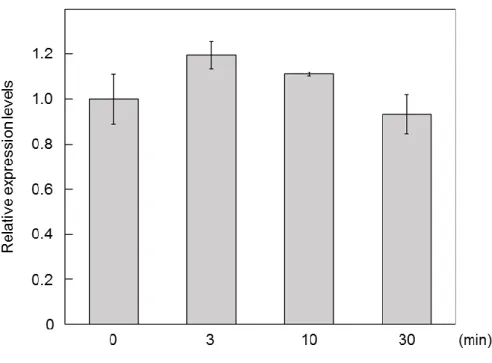
MpPIFのmRNA 発現量には変動がみられなかったことから (図20)、MpPIF タンパク質蓄
積量の速やかな減少はタンパク質レベルでの制御であることが示唆された。さらに、26Sプ ロテアソームの阻害剤である MG132 の処理によって、赤色光による MpPIFの蓄積量の減 少が阻害されたことから (図19E)、赤色光下でのMpPIF蓄積量の減少にユビキチン・プロ テアソーム系が関与することが示唆された。また、proMpEF1:MpPHYY241Hでは暗黒下におい
てもMpPIFタンパク質の蓄積がみられなかった (図21)。以上の結果から、赤色光下におけ
るMpPIFタンパク質の速やかな分解は活性型Mpphyによって引き起こされることが示唆さ
れた。
31 図19 MpPIFタンパク質の光に対する挙動の解析
抗MpPIF抗体を用いて野生型における内性MpPIFタンパク質蓄積量の光に対する挙動を解
析した。連続白色光下で10日間培養した植物体を次に示す光条件下に移してサンプリング を行った。(A) 暗黒下; (B) 遠赤色光照射後(9000 µmol photons m-2) に暗黒下 (+pFR) または 光照射せずに暗黒下 (-pFR); (C) および (D) 48 時間の暗処理後に連続赤色光 (30 µmol photons m-2 s-1; [C]) または連続遠赤色光下 (30 µmol photons m-2 s-1; [D]) (E) 連続白色光下で 10日間培養した植物体に48時間の暗処理を施し (D)、その後赤色光 (30 µmol photons m-2 s-
1) を30分間照射した (R)。赤色光照射の30分前に50 µM MG132またはDMSOを処理し
た。抗Histone H3抗体の結果をコントロールとして示した。
32 図20 赤色光に対するMpPIF発現量の継時的変化
連続白色光下で 10 日間培養した植物体に 48 時間の暗処理を施した後、連続赤色光下 (30 µmol photons m-2 s-1) に移して図に示した時間でサンプリングを行った。MpEF1を内部標準 遺伝子として用いた。グラフは独立した3回の実験における平均値を、エラーバーは標準誤 差を示す。
図21 形質転換体におけるMpPIFタンパク質蓄積量の比較
抗MpPIF抗体を用いて野生型、proMpEF1:MpPHY、proMpEF1:MpPHYY241H、 MppifKO株の暗 黒下における内性MpPIFタンパク質蓄積量を解析した。連続白色光下で10日間培養した植 物体を暗黒下に移して48時間後にサンプリングした。抗Histone H3抗体の結果をコントロ ールとして示した。
33
続いて、MpphyとMpPIFの相互作用および、その光依存性について解析した。大腸菌で 発現・精製したN末端Mpphy-PBと無細胞タンパク質発現系で調整したFLAG-MpPIFを
用いてin vitroプルダウン解析を行った。その結果、活性型のMpphyはFLAG-MpPIFと相
互作用を示したのに対し、不活性型のMpphyやMpPHYアポタンパク質はFLAG-MpPIFと 相互作用を示さなかった (図22A)。次に、活性型MpphyとMpPIFの相互作用にAPAモチ ーフが必要であるかどうか調べるため、APA モチーフ内の2 つのフェニルアラニン残基を アラニンに置換した MpPIFF297A, F300Aを作出し、Mpphy との相互作用を解析したところ、
MpPIFF297A, F300Aは活性型Mpphyと相互作用を示さなかった (図22B)。このことから、APA モチーフは Mpphy との相互作用に必要であることが示唆された。次に、生体内における
MpphyとMpPIFの光依存的な相互作用を明らかにするため、3FLAGタグを付加したMpPIF
を MppifKOに導入した proMpPIF:MpPIF-3FLAG/MppifKOを用いて共免疫沈降による相互作 用解析を行った。連続白色光下で10日間培養した植物体に対して48時間の暗処理を施し、
赤色光のみを照射した植物体および赤色光照射後に遠赤色光を照射した植物体を用いて、
FLAG抗体による共免疫沈降を行った。その結果、赤色光照射サンプルにおいてMpphyと
MpPIF の相互作用がみられ、その相互作用は遠赤色光照射によって阻害された (図 22C)。
これらの結果から、MpPIFは活性型のMpphyと優先的に相互作用することが示唆された。
34 図22 MpphyとMpPIFの相互作用解析
(A) および (B) MpphyとMpPIFのプルダウン解析。大腸菌で発現・精製したMpphyホロタ ンパク質およびアポタンパク質と無細胞タンパク質発現系で調整したFLAG-MpPIFおよび FLAG-MpPIFF297A,F300A (mAPA) を解析に用いた。FLAG-MpPIFとMpPHY-CBDアポタンパク
質、PfrおよびPr型のMpphy-CBDホロタンパク質によるプルダウン解析 (A)、およびFLAG-
MpPIFまたはFLAG-MpPIFF297A,F300AとPfrおよびPr型のMpphy-CBDホロタンパク質のプ ルダウン解析 (B)。キチンレジンに結合したタンパク質を抗Mpphy抗体および抗FLAG抗 体で検出した。(C) MpPIF-3FLAG 発現株を用いた共免疫沈降解析。連続白色光下で10日 間培養した植物体に48時間の暗処理を施した後、赤色光を照射 (9000 µmol photons m-2; R) または赤色光照射後に遠赤色光を照射 (10,800 µmol photons m-2; R/FR) してサンプリングを 行った。細胞分画によって得られた核画分のタンパク質を抗 FLAG 抗体で免疫沈降し、抗
Mpphy抗体または抗FLAG抗体で検出した。
35
考察
Mpphyの光に対する安定性および細胞内局在について
今回の解析から、Mpphy は被子植物のⅡ型フィトクロムと同様に光に対して安定である ことが明らかになった (図 6)。興味深いことに、Mpphy は光に対して安定であるにも関わ らず、被子植物のⅠ型フィトクロムにおける主要なユビキチン化部位が保存されていた (Rattanapisit et al., 2015)。これらから、ユビキチン・プロテアソーム系を介したⅠ型フィトク ロムの速やかな分解は、遺伝子重複によるフィトクロムの機能分化とともに陸上植物進化 の過程で獲得された可能性が考えられた。一方で、細胞内局在に関しては、Mpphyは赤色光 または遠赤色光のいずれの照射においても速やかに核内へと移行するⅠ型フィトクロム様 の性質をもつことが明らかとなった (図7、10)。シロイヌナズナのphyAも同様に赤色光ま たは遠赤色光のいずれの照射によっても核内へと移行するが、赤色光下では速やかに分解 されるため、遠赤色光下でのみ核内へと蓄積する (Oka et al., 2012)。これに対して、Mpphy は赤色光下でも比較的安定なため、赤色光下でも遠赤色光下と同程度に核内に蓄積できる 可能性が考えられた。シロイヌナズナにおいて、FHY1およびFHLがphyAの遠赤色光下で の核移行に重要であることが知られている (Hiltbrunner et al., 2006; Genoud et al., 2008;
Rausenberger et al., 2011)。近年、ヒメツリガネゴケにおけるFHY1ホモログがシロイヌナズ
ナの FHY1/FHL と同様に遠赤色光照射下におけるフィトクロムの核移行に重要であること
が示された (Possart and Hiltbrunner, 2013)。FHY1相同遺伝子はゼニゴケのゲノム上にも存在 するため、ゼニゴケのFHY1ホモログも遠赤色光下におけるMpphyの核移行に関与する可 能性が考えられた。以上の結果から、Mpphy はⅡ型フィトクロム様の光安定性をもちなが ら、Ⅰ型フィトクロム様の細胞内局在パターンを示すというⅠ型、Ⅱ型の両方の性質を併せ もつフィトクロムであることが示唆された。車軸藻Mesotaenium caldariorum のフィトクロ ムはⅠ型、Ⅱ型のどちらにも分類できない性質をもつことが報告されている (Morand et al., 1993)。今回の結果は、陸上植物進化の初期におけるフィトクロムはⅠ型・Ⅱ型のどちらに も分類できないという仮説を支持しており、両方の性質を併せもった祖先的な陸上植物フ ィトクロムから維管束植物の進化の過程でⅠ型・Ⅱ型へと機能分化した可能性が考えられ た。
Mpphyが制御する生理応答
Mpphy は無性芽の発芽や光誘導性遺伝子の発現を赤色光依存的に制御し、これらの応答
は赤色光に続く遠赤色光の照射によって阻害されたことから、Mpphyは被子植物のII型フ ィトクロムと同様に典型的な光可逆性を示すLFR を制御することが示された (図11、13)。
興味深いことに、Mpphyの核移行は遠赤色光下でも観察されたにも関わらず (図7、10)、遠 赤色光のパルス照射や連続遠赤色光は無性芽の発芽や光誘導性遺伝子の発現に有効ではな かった (図11)。そのため、これらの応答は単純にMpphyの細胞内局在だけでは説明できず、
36
核内のフィトクロムの活性状態が重要であることが示唆された。シロイヌナズナにおいて は、phyA は FR照射下で核内に局在し、VLFR や FR-HIRを制御することが知られている (Shinomura et al., 1996; Shinomura et al., 2000)。さらに、FR照射下でのフィトクロムの核局在 やFR-HIR様の応答はヒメツリガネゴケにおいても観察されている (Possart and Hiltbrunner,
2013)。今回の解析からは遠赤色光下で核に局在したMpphyの生理学的な意義は明らかにで
きなかったが、過去の知見からゼニゴケの成長相転換には遠赤色光照射が必要であること が報告されている (Chiyoda et al., 2008; Kubota et al., 2014)。そのため、遠赤色光下で核に局
在したMpphyは成長相転換の制御に関与する可能性が考えられるが、さらなる検証が必要
である。細胞分画による細胞内局在解析の結果、遠赤色光下でのみ核内のMpphyが何らか の翻訳後修飾を受けることが明らかとなった (図 10)。シロイヌナズナにおいて、核内の phyAが遠赤色光下でのみリン酸化されることが報告されており、このリン酸化はCOP1と の相互作用を促進することが示唆されている (Saijo et al., 2008)。このことから、phyAと同
様にMpphyも遠赤色光下において核内でリン酸化される可能性が考えられた。
PIFを介した転写制御による赤色光シグナル伝達
シロイヌナズナにおいて、PIF転写因子はフィトクロムによるシグナル伝達の主要な構成 因子であることが知られている (Leivar and Monte, 2014)。しかしながら、これまでにシロイ ヌナズナ以外の植物種によるPIFの機能はほとんど報告されていない。今回、ゼニゴケに1 分子種のみ存在するMpPIFがフィトクロムシグナル伝達に関与し、無性芽の発芽や光誘導 性遺伝子の発現を抑制することが示された (図18)。シロイヌナズナのPIFファミリーは暗 黒下で蓄積し、赤色光下で速やかに分解することが知られている (Leivar and Monte, 2014)。
これに対し、イネのOsPIL1はOsphyBと相互作用せず、光に対して安定であることが報告 されている (Todaka et al., 2012)。今回、MpPIFは暗黒下で蓄積し、おそらく活性型Mpphyと の相互作用を介して赤色光下でのみ選択的に分解されることが示唆された (図19、22)。こ のことから、MpPIF はシロイヌナズナの PIF ファミリーと共通した分子機構によって活性 型フィトクロムによって機能を阻害されることが示唆された。これらの結果から、陸上植物 進化の基部に位置する苔類が、フィトクロムと PIF 転写因子の相互作用を介した転写制御 機構を既に獲得していたことが示唆された。シロイヌナズナにおいて、PIF転写因子は分解 に先立ってリン酸化されることが報告されている (Al-Sady et al., 2006; Shen et al., 2007; Shen
et al., 2008)。刺激に応じたリン酸化とそれに続くSCF複合体による分解は多くの真核生物
において報告されている (Skaar et al., 2013)。そのため、MpPIFも分解に先立って赤色光依 存的にリン酸化される可能性が考えられるが、さらなる検証が必要である。
今回得られた知見から、ゼニゴケとシロイヌナズナにおける PIF を介したフィトクロム シグナル伝達機構の共通点と相違点が明らかとなった (図23)。ゼニゴケでは、赤色光下に
おいてMpphyは核内に移行し、MpPIFとAPAモチーフを介して相互作用することでユビキ
チン・プロテアソーム系によるMpPIFの分解を誘起する (図19、22)。シロイヌナズナでは、