博士論文
ナマズを中心とした水田利用魚類の繁殖生態 および保全に関する研究
Reproductive and conservation ecology of freshwater fishes using paddy fields,
with special reference to Far Eastern catfish
滋賀県⽴⼤学⼤学院 環境科学研究科 環境動態学専攻 舟尾 俊範
FUNAO Toshinori
2015 年
i
ナマズを中心とした水田利用魚類の繁殖生態および保全に関する研究
目次
第一章 序論 p. 1
第二章 標識再捕獲調査とベイズモデルによるナマズの繁殖個体群推定
2.1 はじめに p. 6
2.2 材料と方法
2.2.1 対象生物について p. 7
2.2.2 調査地 p. 8
2.2.3 水路における採集調査 p. 8
2.2.4 水田と水路における密度および成長調査 p. 9
2.2.5 成長解析 p. 9
2.3 結果
2.3.1 水路における個体数変動および場所間の体長組成の違い p. 10
2.3.2 水田と水路におけるナマズの密度変化および成長 p. 11
2.3.3 水路の各地点おける物理的環境 p. 12
2.4 考察
2.4.1 水田利用魚類にとっての水路環境 p. 12
2.4.2 水田と水路におけるナマズの密度および成長比較 p. 14
2.4.3 水路環境は水田利用魚類の繁殖に適するか p. 15
引用文献 p. 16
図表 p. 18
第三章 改修済み水路における水田利用魚類の繁殖実態と問題点
3.1 はじめに p. 27
3.2 材料と方法
3.2.1 調査地 p. 28
3.2.2 勾配測量 p. 29
3.2.3 定期採集調査 p. 31
3.2.4 補足採集調査およびモンドリ調査 p. 31
3.3 結果
3.3.1 環境要因の季節変化 p. 32
3.3.2 勾配と剪断応力 p. 33
3.3.3 採集生物について p. 33
ii
3.3.4 ナマズおよびコイ・フナ類について p. 33
2.4. 考察 p. 34
引用文献 p. 37
図表 p. 39
第四章 幹線水路に遡上するナマズの繁殖集団規模の推定
4.1 はじめに p. 46
4.2 材料と方法
4.2.1 調査地 p. 47
4.2.2 目視調査 p. 48
4.2.3 モンドリ調査 p. 49
4.2.4 網による捕獲調査 p. 50
4.2.5 ナマズ繁殖集団動態のモデリング p. 51
4.3 結果
4.3.1 目視および捕獲調査 p. 54
4.3.2 解析結果 p. 56
4.4 考察 p. 57
引用文献 p. 61
図表 p. 64
第五章 総合考察
5.1 水路で繁殖する水田利用魚類 p. 75 5.2 幹線水路が抱える問題点 p. 76 5.3 幹線水路に遡上してくる繁殖集団の規模 p. 77
5.4 今後の課題 p. 78
引用文献 p. 79
摘要 p. 80
謝辞 p. 83
1 第一章 序論
陸域と水域の境界には,冠水と干出を繰り返す不安定な環境が存在しており,この ような場所は水辺移行帯と呼ばれる(前畑, 2001a).常に水がある川や湖などの水域を恒 久的水域と呼ぶのに対して,恒久的水域の水位変動に伴って水辺移行帯に出現する水域 を一時的水域と呼ぶ(斉藤, 1997; 前畑, 2004など).一時的水域は降雨量に依存して形成 されるため,天候任せの非常に不安定な水域であるが,日本のように四季が存在し,ま とまった量の雨が降る季節が決まっている地域では,雨期に冠水し,乾季に干出すると いう一定のサイクルを年単位で繰り返している.
しかし,このような一時的水域はほとんどの場合浅い止水域であり,水温が激しく 変動し,いつ干上がるかわからないなど,水生生物にとっては過酷で不安定な環境であ る.一方で,一時的水域には豊富な栄養塩が流入するため,浅い水深による高水温条件 も相まって,プランクトンが大量発生する.さらに大型の捕食者が少ないこともあり,
小型水生生物にとっては好適な環境にもなり得る.このような特性から,一時的水域は 繁殖場所や採餌場所など短期的な生息環境として,多くの生物に利用されてきたことが 知られている(嘉田・遊麿, 2000; 根本, 2010など).
ところが,現在では自然の水辺移行帯はほとんど残されていない.広い一時的水域 が形成されるような平坦な水辺移行帯は,肥沃な土壌が堆積しやすいため,稲作の普及 とともに農地(主に水田)へと開拓されてきたためである(山崎, 1996).また,多くの 場所では,水害防止のため河川改修や護岸整備がすすめられ,現在では水域と陸域とが はっきりと分けられている.これらの人為的な環境改変により,大幅な増水がない限り,
大規模な一時的水域が現れることは現在ではほとんどない.
その少なくなった自然の一時的水域に替わって,水生生物に利用されてきた環境が 水田である.水田では,田植え時期には湛水し,それ以外の時期には陸域になるという
2
冠水と干出のサイクルが繰り返される.水田は,春先から夏にかけて毎年形成される人 工的な一時的水域として,稲作の普及以来,維持されてきた(嘉田・遊麿, 2000; 根本, 2010 など).水田では湛水直後には捕食者が少ないものの,しばらくしてプランクトンが大 量発生する(倉沢, 1955; 山元, 1987)など,自然の一時的水域にきわめてよく似た環境が 形成されている.自然の一時的水域が非常に少なくなっているにも関わらず,一時的水 域を利用する生物の中には現在も日本の平地に残っている生物種も存在しており,それ らは,水田を一時的水域の代替場所として利用してきたと考えられている(日鷹, 1998;
前畑, 2004など).また,かつては水田と恒久的水域との間は素掘りの水路で連続してい
たため,恒久的水域と一時的水域の両方を利用するタイプの水生生物も水田を利用でき た(斉藤, 1977; 嘉田・遊麿, 2000).
ところが,この水田の代替的な一時的水域としての機能も,日本では1960年代以降 に各地で行われてきた圃場整備により大きく損なわれたと考えられている(日鷹, 1998;
重松, 2010など).圃場整備されたほとんどの水田では,排水効率を上げるために水田と
小排水路の間,小排水路と幹線水路の間などに大きな落差が設けられた(片野, 1998). また,水路はコンクリートの2面張りや3面張りとなり,流速が速くなった.これらの 障壁を越えて,圃場整備が行われた水田に魚類が侵入するのは非常に困難であるため,
魚類が繁殖できる水田は非常に少なくなっていると考えられる.繁殖場所の減少は,個 体群規模の減少に直結すると考えられるため,水田を繁殖場所として利用する魚類は減 少 傾 向 に あ る と い わ れ て お り , 各 地 域 の レ ッ ド リ ス ト に 記 載 さ れ る 種 も 多 い(皆 川, 2009).
水田で繁殖する代表的な種類はナマズやコイ,フナ類,ドジョウなどである(内山,
2005; 重松, 2010など).本研究では,水田に依存した生活史を持つこれらの魚類を総称
して水田利用魚類と呼ぶこととする.このうち,ナマズは肉食性の大型魚類であり,日 本の淡水魚類の中では高次捕食者である.ナマズは繁殖および初期成長の時にのみ水田
3
を利用するため,水田地帯に生息するのは,繁殖のために一時的に侵入した場合を除き,
卵から幼魚までの時期に限られる(前畑, 2001b).水田にはもともと大型の水生生物が少 ないため,ナマズは幼魚であっても水田生物群集の中では高次の捕食者に属する.した がって,ナマズ個体群が維持されるためには,一時的水域と恒久的水域の二つの環境が 連続しており,なおかついずれの環境においてもエサとして十分な量の下位生物相が存 在していることが必要であると考えられる.このことから,水田利用魚類の繁殖場所と しての水田環境を評価する上で,ナマズは指標種として考えることができる.
そこで,ナマズを中心に水田利用魚類の生態について調査を行い,その水田地帯に おける生態を解明し,その知見に基づいて保全のための現実的な手法を検討することを 本研究の大きな目的とした.現在では,水田利用魚類が繁殖できる自然の一時的水域や その代替地として利用されてきた圃場整備以前の水田は非常に少なくなっている.一部 の個体は水田周辺の小水路や幹線水路で繁殖している可能性があるが,一時的水域では ない小水路や幹線水路が代替繁殖場所として機能しうるのかどうかについては,これま でにほとんど検証されたことがない.これらの水路の繁殖場所としての機能を検討する ため,いくつかの野外調査を行った.第1章では,水田地帯を流れる,小規模で流れが 穏やかな小排水路において,水田利用魚類の季節消長および成長を調べ,小排水路の繁 殖場所としての機能を検討した.第2章では,小排水路よりも恒久的水域により近い環 境である幹線水路で調査を行い,ナマズ,コイ,フナ類が繁殖する上での問題点につい て検討した.第3章では,幹線排水路において標識再捕法を用いたナマズの繁殖集団規 模の推定を行い,水田およびその周辺環境を利用しうる個体群が現在でも残存している のかどうかについて検討した.第4章では,これらの調査結果を総合し,現在の水田地 帯に存在する大小の水路が水田利用魚類の繁殖場所として果たしている役割,およびそ の保全の可能性について議論を行った.
4 引用文献
日鷹一雅. 1998. 水田における生物多様性とその修復.「水辺環境の保全‐生物群集の視
点から‐」江崎保男・田中哲夫(編), pp. 125−151. 朝倉書店, 東京.
嘉田由紀子・遊磨正秀. 2000. 水辺遊びの生態学 琵琶湖地域の三世代の語りから. 農山 漁村文化協会, 東京. pp. 210.
片野 修. 1998. 水田・農業水路の魚類群集. 「水辺環境の保全‐生物群集の視点から‐」
江崎保男・田中哲夫(編), pp. 67−79. 朝倉書店, 東京.
倉沢秀雄. 1955. 水田におけるPlanktonの消長. 日本生物地理学会会報, 16 : 428−432.
前畑政善. 2001a. 琵琶湖の魚と水辺エコトーン. 「知ってますかこの湖を びわ湖を語る
50章」琵琶湖百科編集委員会(編), pp. 73−78. サンライズ出版, 彦根.
前畑政善. 2001b. 魚類. 「滋賀の田園の生き物」滋賀自然環境研究会(編), pp. 116−128. 滋 賀県農政水産部,大津.
前畑政善. 2004. 琵琶湖の水辺移行帯—魚にとっての水田の役割—. 水, 46: 26−36.
皆川明子. 2009. 田んぼが育む淡水魚とその保全. 「春の小川の淡水魚」水谷正一・森淳
(編), pp. 9−29. 学報社, 東京.
根本正之. 2010. 身近な自然の保全生態学 生物の多様性を知る. 培風館, 東京. pp.213.
斉藤憲治. 1997. 淡水魚の繁殖場所としての一時的水域. 「日本の希少淡水魚の現状と系
統保存—よみがえれ日本産淡水魚—」長田芳和・細谷和海(編), pp. 194−204. 緑 書房, 東京.
斉藤憲治・片野修・小泉顕雄. 1988. 淡水魚の水田周辺における一時的水域への侵入と 産卵.日本生態学雑誌 38: 35−47.
重松敏則. 2010. 里山・里地の変貌.「よみがえれ 里山・里地・里海」重松敏則・JCVN(編), pp. 19−30. 築地書館, 東京.
5
内山りゅう. 2005. 田んぼの生き物図鑑. 山と渓谷社, 東京. pp. 320.
山元憲一. 1987. 水田におけるプランクトンの消長. 水産増殖. 34 : 261−268.
山崎不二夫. 1996. 水田ものがたり‐縄文時代から現代まで‐. 農山漁村文化協会, 東 京. pp. 188.
6
第二章 水田地帯の水路における魚類の成長と移動
2.1 はじめに
水田利用魚類にとっての本来の繁殖場所は,河川の増水などによって形成される自 然の一時的水域であったと考えられる.その環境が農地開発などにより減少し,さらに その代替的機能を担っていた水田も圃場整備によって恒久的水域との連続性が絶たれ たことにより,水田利用魚類の繁殖場所は慢性的に不足しているのが現状である(日鷹, 1998; 前畑, 2001; 水谷・森, 2009など).
その一方で,水田利用魚類の一部は水田周辺の排水路で産卵していることが知られ
ている(斉藤, 1997; 片野, 1998など).水田周辺の排水路が代替的な繁殖場所として利用
されている可能性はあるものの,それを定量的に調査した例は多くない.繁殖場所とな る水田や自然の一時的水域が限られている現状では,水路などでの繁殖状況を調べ,そ の場所の活用あるいは改善の可能性を探ることは,水田利用魚類の現実的な保全策を計 画する上で必要であると考えられる.
ただし,水田地帯に存在する水路的環境は,ごく小規模で流量も少ないものから,
小河川に匹敵するほどの規模を持つ幹線水路まで存在する.実際に一部の幹線水路は,
もともとは小河川であったものを改修し,水路として利用しているものもある.そのた め,水路の環境条件はきわめて多様であり,水田利用魚類による利用実態および生息場 所・繁殖場所としての価値も大きく異なっていることが予想される.全てを水田地帯の 水路として一概に論じることは,いたずらに混乱を招きかねない.水田利用魚類の保全 を目指す上では,これらの異なる水路環境の違いを適切に識別・分類し,ある程度個別 に議論することが必要になると考えられる.
本章では,水路環境の中でも特に水田に近い場所として,その水量の大半を水田か らの排水に依存する,小規模で流れの緩やかな水路(以降,小排水路とする)について,
7
水田利用魚類の繁殖場所としての価値について研究を行った.経時的に小排水路の上流 から下流にかけて水田利用魚類の仔稚魚の採集調査を行い,体サイズ分布の経時的変化 から成長速度を推定し,それら仔稚魚の生息環境としての小排水路の価値を検討した.
特にナマズSilurus asotusについては,この小排水路と周辺の水田で成長速度と密度を調 査・比較することにより,小排水路の生息環境としての質をより定量的に評価すること を試みた.また,ナマズやフナ類Carassius spp.などの稚魚はある程度まで成長すると恒 久的水域へと流下するといわれている(前畑, 2001)が,その移動の過程について詳しく は知られておらず,時間をかけて移動するのか,ごく短時間の間に恒久的水域へと移動 するのかは不明である.そこで,水路の上流部と下流部で個体数と採集個体の体長の変 動を比較して,水路の利用のされ方についても考察した.それらの結果に基づき,小排 水路が水田利用魚類の繁殖場所として活用できるかどうかを検討した.
2.2 材料と方法
2.2.1 対象生物について対象生物について対象生物について対象生物について
ナマズはナマズ目ナマズ科に属する肉食性の淡水魚である.産卵時には,一時的水 域に侵入し,繁殖する(川那部ほか, 2005; 内山, 2005など).フナ類はコイ目コイ科に属 する淡水魚で,ナマズと同様に,増水時に一時的水域に侵入し,産卵する(川那部ほか,
2005; 内山, 2005など).産卵後,親魚は速やかに流下し,一時的水域にはほとんど残ら
ない.この2種類はある程度の大きさまで成長すると流下し,恒久的水域へと生息場所 をかえるといわれている(前畑, 2001).
ドジョウMisgurnus anguillicaudatus(コイ目ドジョウ科)およびメダカOryzias latipes
latipes(ダツ目メダカ科)は水田や周辺の水路,小河川などを主な生息場所としている
(川那部ほか, 2005; 内山, 2005など).親魚は繁殖時以外も水田周辺の水域に生息してい るため,成長後もナマズやフナ類のように大規模な移動はしない.
8 2.2.2 調査地調査地調査地調査地
滋賀県彦根市松原の水田地帯を調査地とした.この水田地帯では幹線排水路が一本 しかなく,一帯の排水はこの幹線排水路へと流入する.この幹線排水路の流路延長は約
840 mであった.幹線水路は,西側では琵琶湖と接続し,南側では矢倉川と接続してい
る(図2.1).いずれの接続地点においても,平水時には水は水路から川へ向かって,あ るいは湖へ向かって流れていた.増水時には河川の増水の影響を受け,流れがよどんで いた.河川や琵琶湖から水路へ水が激しく流れこむような状況は観察されなかった.
この幹線水路は両岸がコンクリートで護岸されていた.底部には砂泥が堆積してい た.琵琶湖側下流部の一部は3面コンクリート張りになっていた.この水路の一部には 堆積して水面にでた土から陸生植物が生えていた.また,一部には抽水植物が生えてい る場所もみられた.しかし,ほとんどの場所では水生植物は見当たらなかった.
水路の琵琶湖側下流部,河川側下流部および中間点付近に調査地点を設け,それぞ
れSt. L,St. RおよびSt. Pとした(図2.1).なお,流れがよどみ,琵琶湖側と河川側ど
ちらに水が流れるか判断がつかない場所を中間点として定めたため,St. Pから両河口ま での距離は等しくない.この水路の幅はSt. Pで約110 cm,St. Lで約250 cm,St. Rで
約115 cmであった.また,St. Pに隣接する水田2枚(水田AおよびB)でも調査を行
った.この2枚の水田は予備調査を行った2011年5月27日の時点で,標準体長で10−20 mmのナマズの稚魚が多数確認された場所である.なお,魚類の標準体長とは,吻端か ら脊柱の末端までの長さを指し,一般的に用いられる体サイズの指標である.
2.2.3 水路における採集調査水路における採集調査水路における採集調査水路における採集調査
2011年6月13日から7月31日までの期間に3日に一回の頻度で調査を行った.水 路に設けたSt. P,St. LおよびSt. Rの3地点で仔稚魚を採集し,種類および個体数を記 録した.なお,フナ類およびヨシノボリ類の仔稚魚は同定が困難であったため,それぞ れフナ類およびヨシノボリ類とした.また,同時に採集された生物については可能な範
9
囲で同定し,種類のみ記録した.採集にはタモ網(D型ネット,幅36 cm,深さ35 cm, 目合い3 mm)を使用した.1地点での調査努力量を採集者1人で10分間とし,定量的 に評価できるように努めた.
調査開始前に,各地点で水温,水深および流速を測定した.流速を測定する際には,
植物片を流し,それが1 m流れる時間から表面流速を算出した.なお,St. LとRの結 果をまとめて扱う場合はSt. LRと表記した.
2.2.4 水田と水路における水田と水路における水田と水路における水田と水路における密度および成長調査密度および成長調査密度および成長調査密度および成長調査
水田AおよびB,およびSt. Pで,ナマズの密度および成長速度を調べた.調査期間
は2011年5月29日から,中干しにより調査不可能となる6月28日までである.ナマ ズが夜行性であること,ナマズ仔稚魚の餌となる動物プランクトンが日中には集中分布 する可能性があることを考慮し,20-22時の間に調査を行うこととした.
魚類は成長に伴って遊泳力が高くなって採集されにくくなるため,生残率を過小評 価する可能性がある.そこで,採集効率の低下を防ぐために,方形枠(30×30 cm,高さ
25 cm)を設置し,その中に入った生物を目合い1 mmの小型の手網で掬い取ることと
した.この方形枠を泥に食い込むように速やかに設置し,逃避する生物が極力少なくな るよう努めた.採集個体については,標準体長を測定したのち,採集地点に放流した.
各調査地点につき,任意の8ヶ所で採集した.なお,水田では,2辺で8地点(角の3 地点を含む)を掬い取ることとした.また,採集個体数が5個体に満たない場合は,随 時採集を行い,できる限り個体数を確保できるよう努めた.
2.2.5 成長解析成長解析成長解析成長解析
各地点で採集されたナマズ仔稚魚の体長の季節変化を,最尤法によって山数 1~3 の 混合正規分布に近似した.ただし,1個体からなるコホートがある場合にはパラメータ ー推定に支障をきたすため,各コホートの全長の標準偏差の最小値を0.5とする制約を つけた.続いてAIC(赤池情報量基準; Akaike, 1974)を基準として,AICが最小となる
10
最適な山数のモデルを選び出した.そして,それぞれの調査日に採集された各個体の体 長が,上記で得られた混合正規分布のどの山に属するかを,それぞれの山の重み付き確 率密度関数に基づいて判別した.
縦軸に体長,横軸に湛水後日数をとった際に,概ねなめらかな増加曲線に乗るグル ープを同一のコホートと見なした.そして,4回以上の調査日で個体が採集されたコホ ートについては,Richardsの成長曲線(Richards, 1959)への近似を行なった.ただし当該 コホートに属すると推定される個体が1個体だけの場合には重み(標準誤差平方の逆数)
が算出されないため,前後の調査日における全長の分散を荷重平均して重みとした.
2.3 結果
2.3.1 水路水路水路水路における個体数変動および場所間の体長組成の違いにおける個体数変動および場所間の体長組成の違いにおける個体数変動および場所間の体長組成の違いにおける個体数変動および場所間の体長組成の違い
水路での採集調査で,6種のべ699個体の魚類を採集した(表2.1).また,その他の 水生生物も10種類採集された.水田地帯で繁殖を行うといわれる魚種としては,フナ 類,ドジョウ,ナマズおよびメダカが採集された.
St. PおよびSt. LRのいずれの地点においても,ナマズおよびフナ類の採集個体数は
調査期間前半に多く,徐々に減少した(図2.2a,b).ナマズが採集されなくなる時期は,
St. Pの方がSt. LRよりも早く,7月上旬には採集されなくなった(図2.2a).St. Pでは,
60 mm以上のナマズは採集されず,40 mmまでの個体が多かった(図2.3a).一方,St. LR
では,やや成長し50 mmを超えた個体が多く採集された.St. PとSt. LRで採集された 個体の体長には有意な差が認められた(Mann-WhitneyのU 検定,P<0.01).7 月には,
St. Pではされなかった60 mmを超える個体が多く採集された(図2.4a).
フナ類については,St. PとSt. LRで採集されなくなる時期に大きな違いはみられな かったが,St. Pでは7月中旬以降採集されない日もあったのに対し,St. LRでは31日 の調査を除いて毎回採集された(図2.2b).また,St. Pに比べてSt. LRで大型の個体が
11
多く採集された(Mann-WhitneyのU検定,P<0.01)(図2.3b).St. Pでは期間を通して
30 mmを超える個体はほとんど採集されなかったが,St. LRでは6月下旬以降に30 mm
を超える個体が多く採集された(図2.4b).
ドジョウの採集個体数は調査期間初期には少なかったが,徐々に増加した(図2.2c). その増加傾向はSt. Pで特に顕著であった. St. Pでは,St. LRに比べて小型個体が多く 採集された(Mann-WhitneyのU検定,P<0.01)(図2.3c).小型個体が頻繁に採集され た時期は6月下旬以降であった(図2.4c).St. LRでは,体長60 mm以上の個体がしば しば採集された.また,St. Pと同様に6月下旬以降には20 mm前後の小型個体が頻繁 に採集されたが(図2.4c),その個体数はSt. Pに比べて少なかった(図2.3c).
メダカは 6 月下旬に採集され始めた.その採集個体数は安定せず,変動しながら推 移した(図2.2d).St. PとSt. LRで採集された個体の体長組成に大きな違いは認められ なかった(Mann-WhitneyのU検定,P = 0.49).
2.3.2 水田と水路におけるナマズの水田と水路におけるナマズの水田と水路におけるナマズの水田と水路におけるナマズの密度密度密度変化および成長密度変化および成長変化および成長変化および成長
水田では,方形枠を用いた調査でナマズを採集できた期間は 5 月29 日から 6 月13 日までであった.この方形枠で採集されたナマズの個体数は水田A,水田BおよびSt. P でそれぞれ18個体,24個体および97個体であった.
ナマズの初期密度はSt. Pで最も高く,水田の2倍以上の密度であった(図2.5).そ の後の密度はいずれの地点でも低下した.特に水田では,調査1回目から2回目にかけ ての急激に減少し,その残存率は30 %であった.一方,水路における残存率の低下は 水田に比べ緩やかであった.
水田AおよびBで方形枠調査および補足調査によりナマズを採集できた期間は5月 29日から6月24日までであった.水田AおよびBそれぞれで,のべ67個体および54 個体のナマズが採集された.また,同期間にSt. Pで採集されたナマズはのべ131個体 であった.
12
いずれの地点においても,採集個体の体長は徐々に増加する傾向にあった.各調査 日に採集されたナマズの体長を山数 1~3 の混合正規分布に近似し,得られたモデルの AICを求めたところ,5月29日のSt. Pの結果を除き,1~2山のモデルのAICが最小と なった(表2.2).孵化日を同一と仮定し,近似した成長曲線では,調査期間中の成長量 において著しい違いはみられなかった(図2.6).期間の前半では一日あたり1−1.5 mm 程度の成長がみられた.水田BおよびSt. Pでは,6月中旬以降,成長が鈍化する傾向 がみられたが,水田A では他の 2 地点に比べて成長率の低下は少なかった.期間の終 わりにおける予測値はいずれの地点の値も40 mm前後であった.
2.3.3 水路の各地点における物理的環境水路の各地点における物理的環境水路の各地点における物理的環境水路の各地点における物理的環境
各地点で測定した環境要因の季節変化を図2.7に示した.地点間の水温の変動に大 きな違いはみられなかった.各地点の水深には差があったが,その変動は同調していた.
流速に関しては,St. Pでは10 cm/sec以下の日が多かったのに対し,下流側の2地点で
は10 cm/sec以上の日がほとんどであった.
2.4 考察
2.4.1 水田水田水田利用水田利用利用利用 魚類魚類魚類にとって魚類にとってのにとってにとってののの水路水路水路環境水路環境環境環境
St.Pでは,7月にナマズはほとんど採集されなかった.また,St. Pでは60 mm以上 の個体は採集されなかった.一方,St. LRでは7月下旬までナマが採集され,7月に採 集された個体の多くは体長60−90 mmであった.ナマズの稚魚は水田で成長し,降雨な どで増水した際に水田から恒久的水域へと下るといわれており,その大きさは全長で
40−70 mmといわれる(前畑, 2001).St. Pで採集されなくなった大きさはこれに一致して
おり,成長したナマズは下流側へ移動したと考えられる.また,St. LRでは90 mm程 度まで成長した個体が採集されたことから,水路の環境条件によって,この程度の大き さまで成長できることがわかる.
13
以上のことから,ナマズにとって水路上流部は繁殖と初期成長の場所であり,水路 下流部は,移動経路および恒久的水域に移る前の成長の場所として機能していると考え られる.
St. Pでは体長30 mm以上のフナ類はほとんど採集されなかった.水田におけるニゴ
ロブナの成長率は徐々に鈍化することが知られており,水田が好適な生息環境となるの
は全長25 mmに達するまでといわれている(金尾ほか, 2009).7月中旬以降にSt. Pで採
集されたフナ類の体長は20−30 mmであり,それ以前と比べて成長した個体が採集され ることはなかった.このことから,St. Pでのフナ類の成長は,水田における成長と同様
に,20−30 mm程度で横ばいになると考えられる.また,この時期には採集される個体
数が少なくなったことから,流下を開始する大きさも20−30 mm程度であると考えられ る.
一方,St. LRでは,7月に採集された個体の多くは体長30 mm以上であった.金尾
ほか(2009)は水田でのフナ類の成長が鈍化する要因の一つとして,餌生物がプランクト
ンから底生生物に変化することを挙げている.水路の下流部では体長40 mm前後のフ ナ類の餌となる生物が多く生息しているのかもしれない.これらのことから,フナ類も ナマズと同様に,水路上流部を繁殖および初期成長の場所として利用し,成長しながら 流下していくと考えられた.ただし,フナ類はナマズに比べると調査前半で採集された 個体数が多かったにも拘わらず,後半に採集された個体数は少なかった.このことから,
水路に残って成長した個体は取り残されただけで,大半は速やかに流下したのかもしれ ない.
ドジョウについては,St. Pでの採集個体数は時期が下るにつれて増加する傾向にあ った.また,St. Pで採集されたドジョウの大半は体長60 mm以下の個体であった.ド ジョウのオスは体重2 gで,メスは体重3.5 gで繁殖可能であるといわれている(鈴木, 1983).この体重に相当する体長はおよそ60 mmと78 mmである(久保田, 1961).St. P
14
では,調査初期には50 mm以上の個体が採集され,6月中旬以降に小型の個体が多く採 集された.初期に採集された大型の個体は一歳魚以上の成魚であり,6月中旬以降に採 集された個体の大半は当歳魚であると考えられる.調査期間中,採集個体数が増加傾向 にあり,小型個体が多く採集されたことから,ドジョウは,水路あるいは周辺の水田で 随時繁殖していたと考えられる.また,各調査日に採集された最大個体の体長が増加傾 向にあったことから,St. Pの周辺でドジョウは成長できていたと考えられる.
St. LRにおいても,St. Pと同様に,ドジョウの個体数は増加する傾向にあった.た
だし,その増加の仕方はSt. Pに比べ緩やかであった.また,St. LRでは体長60 mmを 超える個体が調査期間を通して頻繁に採集された.St. LRはドジョウ仔稚魚の成長場所 であるとともに,成魚の生息場所にもなっていたと考えられる.水路の上流部で親魚が ほとんど確認されなかったことから,ドジョウもナマズやフナ類ほど極端ではないもの の,親魚と仔稚魚の成長する場所が異なる可能性がある.水路上流部は仔稚魚の成長場 所として,下流部は成長した個体の生息場所として機能していると考えられる.
メダカはSt. Pで6月下旬から採集され始め,St. LRにおいても7月上旬から採集さ
れ始めた.その採集個体数はフナ類やドジョウに比べると少なく,概ね横ばいで推移し
た.また,St. PとSt. LRの間で体長組成に違いはみられなかった.いずれの地点でも,
7月中旬に10−15 mmの小型の個体が採集されたことから,繁殖している可能性は高く,
水路で成長していると考えられるが,本調査ではそのことを裏付ける十分な結果は得ら れなかった.
2.4.2 水田と水路水田と水路水田と水路水田と水路におけるにおけるにおけるにおけるナマズの密度および成長比較ナマズの密度および成長比較ナマズの密度および成長比較ナマズの密度および成長比較
水田と水路でナマズの成長速度に大きな違いはみられなかった.初期密度は水路の 方が高く,その後の減少率も水路の方が小さかった.また,水田に比べて水路の方が成 長速度が遅いという傾向はみられなかった.これらのことから,本調査地では,水田だ けではなく,水路もナマズの繁殖場所として機能していたことが示唆される.
15
ナマズ稚魚の初期のエサは水田に豊富に発生する動物プランクトンである.本調査 地では,降雨時や水田への注水時には排水が水路へと流れ込んでおり,水田で発生した プランクトンが排水とともに水路へと供給されていたと考えられる.また,ナマズは同 種・他種を問わず,口に入る大きさの稚魚も捕食する.水路は水田に比べ,生物の密度 が高くなるため,ある程度成長した後は水田よりもエサに巡り合いやすい場所となるか もしれない.
2.4.3 水路環境は水田利用魚類の繁殖に適するか水路環境は水田利用魚類の繁殖に適するか水路環境は水田利用魚類の繁殖に適するか水路環境は水田利用魚類の繁殖に適するか
St. PとSt. LRでは,メダカを除いた3種について,体長に有意な差が認められた(図
2.3).いずれの種類の体長組成も,St. Pの方が小型個体が多い傾向にあった.これは,
St. Pが水田魚類の繁殖場所としての性質を備えていることを示している.すなわち,水
田地帯の水路は一時的水域に近い条件であるといえる.
St. LRに比べてSt. Pで採集されたナマズおよびフナ類が小型で,しかも調査期後半
にかけて採集されにくくなった.これは上流部(St. P)は初期成長の場所であり,成長 したのち移動したためであると考えられる.また,St. Pではドジョウの親魚はほとんど 採集されなかった.ドジョウもナマズやフナ類と同様に,水路上流部を主に繁殖場所と して利用しているためと考えられる.これらのことは,St. Pの方が一時的水域に近い環 境条件であり,St. LRが一時的水域と恒久的水域との中間的な環境であったことを示唆 している.
一時的水域と恒久的水域をつなぐ水路の中でも,水田周辺の小排水路は水田が冠水 している時期にのみ通水する場所も多く,一時的水域に準じる環境条件となっていると 考えられる.水田周辺の排水路では,一般的に水深は浅く,水田からの排水が流量の大 半を占める.そのため,一時的水域のメリットである,高水温・捕食者の不在などの環 境条件となる場所が多い.本研究では初期の餌となるプランクトンについて調査してい ないが,水田-水路間の成長速度が同程度であったことから,水路の餌条件も悪くなか
16
ったと考えられた.これらの環境条件が整う水路は水田地帯では少なくないと考えらえ る.水路は水田利用魚類にとって繁殖に十分な環境とはいえないまでも,繁殖可能な場 所であると考えられる.
引用文献
Akaike H. 1974. A new look at the statistical model identification, IEEE Transactions Automatic Control 19: 716−723.
日鷹一雅. 1998. 水田における生物多様性とその修復.「水辺環境の保全‐生物群集の視
点から‐」江崎保男・田中哲夫(編), pp. 125−151. 朝倉書店, 東京.
金尾滋史・大塚泰介・前畑政善・鈴木規慈・沢田裕一. 2009. ニゴロブナCarassius auratus grandoculis の 初 期 成 長 の 場 と し て の 水 田 の 有 効 性. 日 本 水 産 学 会 誌 75:
191−197.
片野 修. 1998. 水田整備事業の魚類への影響. 「水辺環境の保全—生物群集の視点から
—」江崎保男・田中哲夫(編), pp. 67–79. 朝倉書店, 東京.
川那部浩哉・水野信彦・細谷和海(編). 1989. 日本の淡水魚. 山と渓谷社, 東京. pp. 719.
久保田善二郎. 1961. ドジョウの生態に関する研究-Ⅳ 成長度および肥満度について. 農水講研報 11: 213−234.
前畑政善. 2001. 魚類. 「滋賀の田園の生き物」滋賀県自然環境研究会(編), pp. 116−128, 滋賀県農政水産部, 大津.
前畑政善. 2004. 琵琶湖の水辺移行帯—魚にとっての水田の役割—. 水 46: 26–36.
Richards, F. J. 1959. A Flexible Growth Function for Empirical Use. Journal of Experimental Botany 10: 290−300.
水谷正一・森 淳(編). 2009. 春の小川の淡水魚. 学報社, 東京. pp. 190.
斉藤憲治. 1997. 淡水魚の繁殖場所としての一時的水域. 「日本の希少淡水魚の現状と系
17
統保存—よみがえれ日本産淡水魚—」長田芳和・細谷和海(編), pp. 194−204. 緑 書房, 東京.
鈴木 亮. 1983. ドジョウ養殖の最新技術. 泰文館, 東京. pp. 56−57.
友田淑郎. 1978. 琵琶湖とナマズ 日本の野生動物10. 汐文社, 東京. pp. 326.
内山りゅう. 2005. 田んぼの生き物図鑑. 山と渓谷社, 東京. pp. 320.
18
表 2.1 採集された魚類およびその他の生物.各魚種の数字は個体数を示す.なお,アメン ボ類に関してはアメンボ属として,まとめて扱った.
Japanese name Scientific name
Fishes フナ類 Carassius sp. 146
ドジョウ Misgurnus anguillicaudatus 345
ナマズ Silurus asotus 71
メダカ Oryzias latipes 99
ヨシノボリ類 Rhinogobius sp. 36
ウキゴリ Gymnogobius urotaenia 2
Others アマガエル Hyla japonica -
トノサマガエル Rana nigromaculata - ヒメタニシ Sinotaia quadrata histrica - アメリカザリガニ Procambarus clarkii - シオカラトンボ類 Orthetrum sp. - コオイムシ Appasus japonicus - マツモムシ Notonecta triguttata - ハイイロゲンゴロウ Eretes sticticus - アメンボ類 Gerridae spp. - コミズムシ類 Sigara sp. -
19
表2.2 各調査日に採集されたナマズの体長を山数1~3の混合正規分布に近似して得られた 各モデルのAIC.網掛けされた値は最小値である.水田Bでは6月16日には十分な個体数 が採集されなかった.
No. of peak St. P Paddy A Paddy B
5/29 1 235.3 87.0 33.6
2 234.3 86.3 36.7
3 232.6 87.7 43.5
6/1 1 155.5 46.0 25.4
2 160.1 45.2 24.9
3 164.0 47.0 30.4
6/4 1 39.8 29.5 25.2
2 38.7 34.7 29.6
3 44.7 31.7 35.8
6/7 1 48.7 45.1 41.8
2 51.3 43.4 38.9
3 53.7 45.1 43.9
6/10 1 76.5 40.2 29.5
2 75.7 43.5 31.9
3 78.0 47.0 33.5
6/13 1 73.2 45.9 43.1
2 63.0 45.6 46.2
3 63.1 50.1 46.7
6/16 1 18.1 22.6
2 23.0 25.5
3 26.1 26.1
6/19 1 27.0 22.5 21.9
2 31.8 24.3 22.2
3 36.1 23.9 27.7
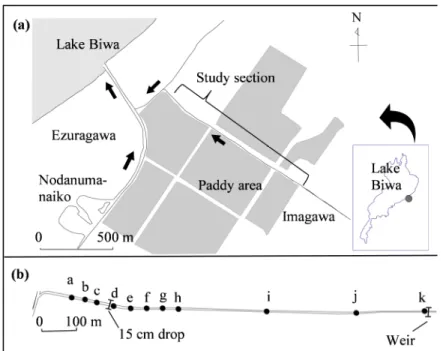
20 図2.1 調査地概要
濃灰色部分は水田地帯を表す.矢印は流れの方向を示す.○は 各調査地点St.L,PおよびRを示す.また,St.Pに隣接する□
は調査水田AおよびBを示す.
21
図2.2 St.PおよびSt.LRにおける各種の採集個体数の変動.点線と実線はそれぞれ
St.PおよびSt.LRの個体数の変動を示す.なお,St.LRの個体数はSt.LおよびSt.R の平均値である.(a)ナマズ,(b)フナ類,(c)ドジョウ,(d)メダカ.
No. of individuals
22
図2.3 St.PおよびSt.LRにおける各種の採集個体の体長組成.いずれの種についても,
横軸の一目盛りは5 mmである.(a)ナマズ,(b)フナ類,(c)ドジョウ,(d)メダカ.
0 5 10
1 2 3 4 5 6 7 8 9 10 11 12 13 14 0
5 10
1 2 3 4 5 6 7 8 9 10 11 12 13 14
0 10 20 30
1 2 3 4 5 6 7 8 9 10
0 10 20 30
1 2 3 4 5 6 7 8 9 10
S. asotus Carassius spp.
St. P
St. LR
St. P
St. LR
0 20 40
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 0
20 40
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
0 10 20 30
1 2 3 4 5 6 7 8
0 10 20 30
1 2 3 4 5 6 7 8
M. anguillicaudatus O. latipes
St. P
St. LR St. P
St. LR
30 40 50 60 70 80 90 10 20 30 40 50
10 20 30 40 50 60 70 80 90 0 10 20 30 40
(a)
(c)
(b)
(d)
No. of individuals
Standard length (mm)
23 0
20 40 60 80 100
6/11 6/21 7/1 7/11 7/21 7/31
0 20 40 60 80 100
6/11 6/21 7/1 7/11 7/21 7/31
0 10 20 30 40 50
6/11 6/21 7/1 7/11 7/21 7/31
0 10 20 30 40 50
6/11 6/21 7/1 7/11 7/21 7/31
0 20 40 60 80 100
6/11 6/21 7/1 7/11 7/21 7/31
0 20 40 60 80 100
6/11 6/21 7/1 7/11 7/21 7/31
0 5 10 15 20 25 30 35
6/1 6/21 7/11 7/31
0 5 10 15 20 25 30 35
6/11 6/21 7/1 7/11 7/21 7/31
図2.4 St.PおよびSt.LRで採集された各種の体長の季節変化.1プロットが採集され
た1個体の体長を表す.(a)ナマズ,(b)フナ類,(c)ドジョウ,(d)メダカ.
Standard length (mm)
(a) (b)
(c) (d)
S. asotus St. P
Carassius spp. St. P
M. anguillicaudatus St. P
O. latipes St. P
St. LR St. LR
St. LR St. LR
24
図2.5 水田A,Bおよび水路St.Pにおけるナマズの密度(個 体数 / m2)(上段)および残存率の推移(下段).残存率は5
月29日の個体数を100として算出した.
0 10 20 30 40 50 60 70
5/29 6/1 6/4 6/7 6/10 6/13
Paddy A Paddy B St. P
0 50 100
5/28 5/31 6/3 6/6 6/9 6/12
Paddy A Paddy B St. P
5/29 6/1 6/4 6/7 6/10 6/13
Survival rate (%) Density (No. of individuals / m2 )
25
0 10 20 30 40 50
5/27 6/1 6/6 6/11 6/16 6/21
St. P St. P Paddy A Paddy A Paddy B Paddy B
S ta nda rd l eng th (m m )
Days
図2.6 各地点で採集されたナマズの体長の平均値およびモデル予測値.点が 各調査日に採集されたナマズの体長の平均値を示し,線が各地点のモデル予 測値を示す.
26
図2.7 水路の各地点(St. P,LおよびR)にお ける物理要因の季節変化.上段:水温,中段:
水深,下段:流速.
0 10 20 30 40
St.P St.L St.R
0 5 10 15 20
0 10 20 30
6/16 6/23 6/30 7/7 7/14 7/21 7/28
Water temperature (℃) Flow velocity (cm/sec) Water depth (cm)
27
第三章 幹線水路における水田利用魚類の繁殖実態と問題点
3.1 はじめに
第二章では水田地帯の水路のうち特に規模の小さい排水路について,水田利用魚類の 成長および移動について調べ,ナマズSilurus asotus・フナ類Carassius spp.をはじめとす る水田利用魚類の繁殖場所として小排水路が有効に機能している場合があることを述 べた.その理由として,小排水路は水田に比較的近い環境であるために,水田利用魚類 の繁殖場所として好適な条件を備えていることが考えられた.しかしその一方で,この ような小排水路は規模が小さいために全体の面積も限られており,水田利用魚類の個体 群を維持する上では限定的な貢献しか期待できないかもしれない.また,恒久的水域と 水田をつなぐ水系のなかで,小排水路は水路のもっとも上流部にあたり,水田利用魚類 の親世代個体群が生息している恒久的水域と離れている.その間に,落差などにより恒 久的水域との連続性が保たれていない場所も多く存在する.そのため,全ての小排水路 が生息場所として機能できるわけでもないだろう.
そこで本章では,より下流側に位置し,小排水路が多数流入する幹線水路を研究の対 象とした.多くの幹線水路や小河川では常に水が流れており,恒久的水域に相当すると 考えられる.そのため,一時的水域を産卵場所とする魚類の産卵場所として適していな いと一般的には考えられる.加えて,幹線水路は両岸や底をコンクリートで護岸されて いる場所がほとんどである.一方で,田植え時期の水田地帯では,幹線水路の流量の大 半を水田からの排水が占めることもあり,水質などの条件については小排水路と似た条 件である可能性がある.水田利用魚類が幹線水路で産卵している例は少なくないと考え られるが,本来は一時的水域を利用する水田利用魚類がどの程度幹線水路を含む恒久的 水域で繁殖しているかについては,ほとんど知見がないのが現状である.
幹線水路における水田利用魚類の繁殖実態を報告した数少ない例として,滋賀県彦根
28
市を流れる江面川水系において採集された仔稚魚の種を報告したSuzuki et al. (2008)が 挙げられる.Suzuki et al. (2008)は採集した仔稚魚について,その形態とRAPD (Random
Amplified Polymorphic DNA)法に基づいて種の同定を行い,9種の魚類を報告している.
この報告はコイ Cyprinus carpio およびフナ類を主な対象とした定性的な調査に基づい ており,それ以外の魚種について,あるいは経時的・定量的な評価は行っていない.ま
た,Suzuki et al. (2008)以外には,水田利用魚類の繁殖場所としての幹線水路の価値を評
価した例は見当たらない.
本研究では,幹線水路における水田利用魚類の繁殖実態を明らかにするため,Suzuki
et al. (2008)でコイおよびフナ類の成長が確認された江面川水系の今川を調査地とし,コ
イ・フナ類に加えてナマズの卵と仔稚魚を対象として経時的・定量的な調査を行った.
さらに,調査地における卵の流されやすさを評価するため,水路の測量およびその結果 に基づいた物理的な環境条件の分析を行った.これらの結果を総合し,現在の幹線水路 の産卵場所としての機能および改善が必要な点について検討した.
3.2 材料と方法
3.2.1 調査地調査地調査地調査地
滋賀県彦根市八坂町を流れる江面川支流の今川で調査を行った(図 3.1).今川は流
路延長1431 mの河川である.その主な水源は,開出今町集落内に存在する複数の湧水
である.今川は開出今集落を流れ出た後,水田地帯を流れる.そして水田地帯の末端で 南西方向へとほぼ直角に屈曲し,江面川に流入する(以下,この合流点のことを今川河 口と称する).江面川は今川と合流した地点から約360 m下流で琵琶湖に流入している.
また江面川本流は上流側で野田沼とつながっている.
今川は人工的に直線化された河川であり,その両岸はコンクリート張りである.今 川河口とその約300 m上流の間の区間では,直径5−20 cm程度の礫が河床を覆っている.
29
それより上流ではコンクリート河床が連続しているが,部分的に砂泥で覆われている区 間も存在する.
今川河口から約460 m上流の地点には落差約15 cmの段差がある(この段差の上流 側の地点は採集調査のSt. dに相当する).この段差より下流側の河床はコンクリート張 りだが,その上に直径20−50 cm程度の礫が配されており,砂泥が堆積している場所も 多い.また,今川河口の上流約270 mから410 mまでの範囲にはヨシPhragmites australis
やマコモZizania latifoliaなどの抽水植物が多く繁茂しており,流れが緩やかになってい
た.
水田地帯区間の今川は農業用の排水路として利用されており,この区間では複数の 小排水路が今川に流入している.水田からの排水のほとんどはパイプを介して小排水路 に流入している.また,この地域のほとんどの水田では,琵琶湖や地下水からポンプに より取水している.したがって今川周辺の水田には魚類が侵入することは不可能である.
今川河口の上流約250 m,670 m,1000 m,1220 mおよび1430 mの各点で,ある程 度の流量を有する水路が合流している.これらのうち河口の上流約670 mと約1000 m の地点で流入する水路は,流量が特に多かった.
今川河口の上流約1.5 kmの地点には勾配が0.58(約30度)の傾斜部があり,その落
差は約85 cmである.魚類がこの傾斜部より上流へ遡上することは非常に困難である.
この傾斜部を調査範囲の最上流端とした.今川河口からこの傾斜部までの範囲には,魚 類の遡上を妨げる構造物は見当たらなかった.
今川では,ヨシやエビモPotamogeton perfoliatusなどの抽水・沈水植物が部分的に繁 茂していた.川幅を任意の数地点で測定したところ,最小値は219 cm,最大値は316 cm であった.川幅は下流ほど広くなる傾向にあった.
3.2.2 勾配測量勾配測量勾配測量勾配測量
今川河口の上流約330 mから670 mの範囲で河床高の水準測量を行い,これをもとに
30
河床勾配を求めた.この区間にはSt. a−hが含まれる.測量にはレーザーレベルと標尺
を用い,St. dを基準点として,20 m間隔で河床の比高を測った.また,St. dの下流40
mの地点より下流側では細かな起伏が多かったため,5 m間隔で測量した.測量した比 高と測定地点間の距離から,St. dの上流側および下流側の2区間について,それぞれの 勾配を求めた.
次に,St. dの上流側および下流側の2区間で,魚類の卵などの物体を流そうとする力
がどの程度違うかを比較するため,水流が河床に与える剪断応力を求めた.その算出に は,採集調査のSt. a,c,dおよびhにおける水深を用いた.St. aおよび cは河口から
460 mの地点にある段差の下流側区間の両端に位置する地点,St. dおよびhは段差の上
流側区間の両端に位置する地点である.なお,調査を行った最上流地点はSt. kである が,勾配を求めるための測量を行った地点の中ではSt. hが最上流部である.以下,St. a からcまでの区間を区間a−cと称し,St. dからhまでの区間を区間d−hと称する.各地 点での測定は2008年5月30日に行った.
剪断応力τは以下の式から求められる.
τ = (1)
ただし,ρは水の密度,gは重力加速度,Rは径深,Iは勾配である.また径深Rは 以下の式で求められる.
= (2)
ただし,Aは各区間の流積,Sは潤辺である.調査対象の水路は矩形断面を呈してい るので,Sは以下の式で求められる.
= 2 + (3)
ただし,Hは水深,Wは水路幅である.
剪断応力を求めるため,St. aおよびcの水深から平均水深を求め,それをこの区間の 平均水深とみなした.同様にSt. dおよびhの水深から求めた平均値を上流区間の平均
31
水深とした.また,ρ = 1000 kg m-3,g = 9.8 m s-2,W = 3.1 mとして計算した.
3.2.3 定期採集調査定期採集調査定期採集調査定期採集調査
タモ網を用いて卵および仔稚魚の採集調査を行った.使用したタモ網は幅40 cmのD 型ネットで,深さ30cm,網目1mm×1mmである.調査期間は2008年5月14日から7 月31日までとし,この79日間に調査を58回行った.調査を行う時間は基本的に15時 から17時の間とした.2008年5月14日から5月19日まではSt. aからhの8地点で調 査を行った.各地点の間隔は40から50 m間隔である.5月30日以降にはSt. iからk を追加した11地点で調査を行った(図4.1.b).St. iからkの3地点の間隔は200ないし
280 mである.卵および遊泳力の低い仔魚の採集を主な目的としたため,網の先端の平
らな部分を河床につけ,上流側に向かって河床をこするように掬い取った.河床の 40
cm×40 cmの範囲を掬い取るようにし,取りこぼしをなくすために同じ場所で3回掬い
取った.また,各地点につき両岸沿い2か所で掬い取った.採集した卵および仔稚魚を 計数し,仔稚魚については標準体長を測定したのち,同地点に放流した.10 mm以下の 小型の個体については全長を測定した.なお,ナマズおよびコイ・フナ類の卵はそれぞ れ黄緑色の粒径 3 mm 程度の卵と半透明の粒径 1.4−2.5 mm の卵である(川那部ほか,
1986)ことから,肉眼による観察で区別することができる.
St. dで水温,水深および流速を測定した.流速を測定するときは,植物片などを流し,
1 m流れる時間を計測し,流速を算出した.また,国土交通省近畿地方整備局琵琶湖河 川 事 務 所 の ウ ェ ブ サ イ ト(http://www.biwakokasen.go.jp/graph2/csvlist.html)か ら 琵 琶 湖 の 水位のデータを得た.
3.2.4 補足採集調査補足採集調査補足採集調査補足採集調査およびモンドリ調査およびモンドリ調査およびモンドリ調査およびモンドリ調査
定期採集調査では,卵の取りこぼしを少なくするために,河床をこするように掬い取 る方法をとった.この方法では,成長して遊泳力が高くなった稚魚は網から逃げてしま うため,採集効率が低くなる.コイ・フナ類とナマズそれぞれの稚魚がどの程度生き残
32
っているかを確認するために補足調査を行った.補足調査には定期採集調査で用いた網 と同じ網を用いた.主に壁沿いや植物が茂った場所などで,タモ網にむかってけりこむ ように掬い取りを行った.採集された魚類の個体数および標準体長を記録し,採集地点 に放流した.2008年6月2日,7月1日,8日,16日および28日,8月14日に計6回 の調査を行った.採集者は1人とし,調査時間は15分とした.
大型魚類を採集するため,モンドリと呼ばれるワナを設置した.このモンドリは幅
62 cm,長さ92 cm,高さ54 cmのドーム型で,その目合いは1 cm×1 cmである.モン
ドリを使ったこの調査は,調査地に存在する魚類の個体数や体サイズの変化を相対的に 評価することを目的としたため,モンドリ内部に餌などの誘引剤は入れなかった.モン ドリの設置期間は2008年5月21日から8月1日までの73日間である.採集物の確認 は1,2日に1回程度とし,計59回確認した.採集された魚類については,種数,個体 数および標準体長を記録したのち,採集地点に放流した.
3.3 結果
3.3.1 環境要因の季節変化環境要因の季節変化環境要因の季節変化環境要因の季節変化
本調査期間中の水温は緩やかに上昇する傾向にあった(図3.2a).調査期間中の最低 水温は19.0℃,最高水温は32.4℃であった.水深は6月中旬まで17 cm以上を保ってお り,それ以降は増水時を除いて5−10 cm前後で推移した(図3.2b).流速は6月中旬ま
で主に20−40 cm/secで推移したが,それ以降は速くなり、40−60 cm/secの日がほとんど
だった(図3.2c).
琵琶湖の水位は管理水位に基づいて操作されている.2008 年の琵琶湖の管理水位は
6月16日まで+30 cmに,それ以降は−20 cmに設定されていた.なお,琵琶湖の水位は
鳥居川水位観測所の零点高を水位0 mとして定められている.実際の水位は+10 cm程 度で推移して,6月上旬から緩やかに低下し,6月中旬に管理水位である−20 cm前後と
33 なった(図3.2d).
3.3.2 勾配と剪断応力勾配と剪断応力勾配と剪断応力勾配と剪断応力
平均勾配は区間a−cおよび区間d−hそれぞれで0.0005および0.0013であった(図3.3). これは区間a−cおよび区間d−hにおいて,100 mの間にそれぞれ5 cmおよび13 cmの 落差があることを示す.剪断応力は区間a−cおよび区間d−hそれぞれで1.75 Nm-2およ び2.26 Nm-2であった.
3.3.3 採集生物について採集生物について採集生物について採集生物について
定期採集調査,補足採集調査およびモンドリ調査で採集された生物を表3.1に示した.
本調査の対象種であるナマズおよびコイ・フナ類を含め,14種のべ 591 個体の魚類が 採集された.最も多く採集されたのはコイ・フナ類で,次いでミナミメダカOryzias latipes およびヨシノボリ類Rhinogobius spp.が多かった.
また,魚類以外にもアメリカザリガニProcambarus clarkii,エビ類,トンボ幼生,ウ
シガエルRana catesbeianaなどが採集された.
3.3.4 ナマズおよびコイ・フナ類についてナマズおよびコイ・フナ類についてナマズおよびコイ・フナ類についてナマズおよびコイ・フナ類について
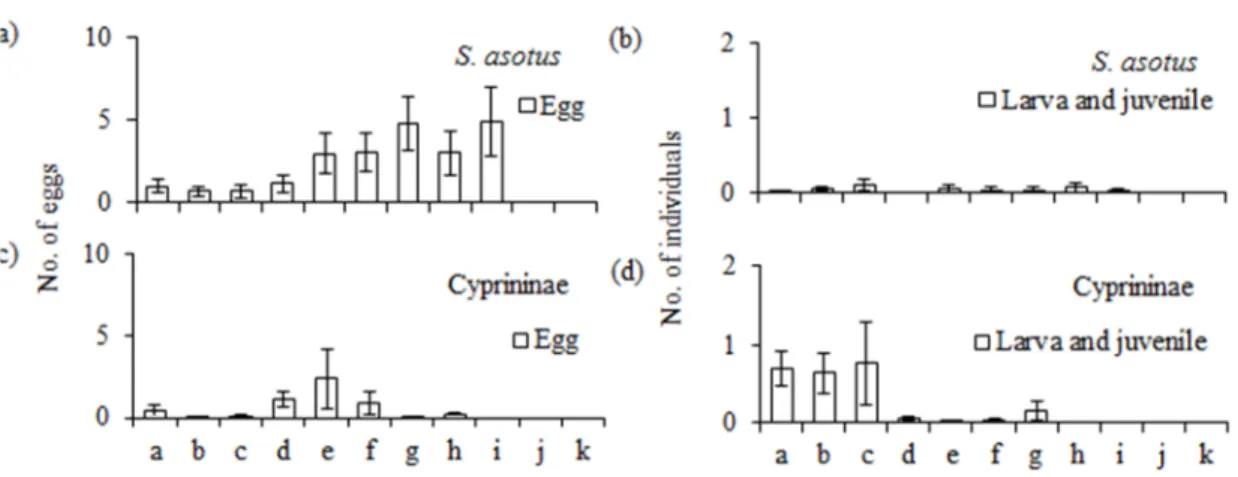
定期採集調査で,のべ1210個のナマズの卵を採集した.ナマズの卵は58回の調査の うち30回採集された.調査を開始した5月中旬から6月中旬にかけて頻繁に採集され た(図3.4).
またコイ・フナ類の卵はのべ 308 個採集できた.コイ・フナ類の卵が採集された回 数は58回の調査のうち11回であり,ナマズに比べ採集頻度は少なかった.多く採集で きた時期は,ナマズの卵と同様に5月中旬から6月中旬にかけてであった(図3.4).
定期採集調査で採集できたナマズの仔稚魚はのべ22個体であった.ナマズの仔稚魚 は58回の調査のうち8回の調査で採集された.採集されたナマズの仔稚魚の大きさは
3−21 mmで,その平均値±標準偏差は7.0 ± 6.3 mmであった.
これに対し,コイ・フナ類の仔稚魚はのべ 136 個体採集された.コイ・フナ類の仔
34
稚魚は58回の調査のうち23回の調査で採集された.採集されたコイ・フナ類の仔稚魚
は6−39 mmで,平均値±標準偏差は12.9 ± 5.5 mmであった.
ナマズおよびコイ・フナ類のいずれの種類についても,体長20 mm以下の個体が95 % 以上を占めた(図3.5).ナマズの方が仔魚の占める割合が高く,全長10 mm以下の個 体が7割以上であったのに対し,コイ・フナ類では4割程度であった.
ナマズの卵は調査範囲の中流部で多く採集される傾向にあった.これに対し,コイ・
フナ類の卵は比較的下流側の地点で多く採集された(図3.6).コイ・フナ類の仔稚魚は 下流側で多く採集された.ナマズの仔稚魚は中・下流部で採集された.やや成長した稚 魚が採集された地点は下流部のみであった.
3.4 考察
琵琶湖の水位が低下した6月中旬以降には本調査地の水深も浅くなった.本調査地の 水深は琵琶湖の水位の影響を受けていたと考えられる.比較的水深の深かった5月には,
ナマズおよびコイ・フナ類のいずれの卵も多く採集された.5月までは,流速も比較的 遅く,本調査地の水深や流速は琵琶湖の水位による影響を受けて,止水環境に近くなっ ていたと考えられる.
測量の結果,区間a−cの剪断応力は1.75 Nm-2であり,区間d−h(2.26 Nm-2)よりも 小さい値となった.河床にある物体が水流により運ばれるためには,ある一定値以上の 剪断応力が河床に加わる必要がある.すなわち,河床に加わる剪断応力が大きいほど,
河床にある物体は運ばれやすい環境となる.本研究では卵の粒径および密度を測定して いない.また,対象種の卵の密度についての既存の報告は見当たらず,卵が流され始め る剪断応力を求めることはできない.しかし,区間a−cの剪断応力はd−hのそれの8割 以下であることから,区間a−cの方が区間d−hに比べて卵が運搬されにくいといえる.
さらに,区間a−cには微起伏がある上,水生植物が部分的に繁茂していることから,こ
35
れらが水への抵抗を大きくしていると考えられる.また琵琶湖の水位の影響を受けて水 深が深く保たれていたことも考慮すると,区間a−cでは卵が流されにくく滞留しやすい 環境にあると考えられた.しかし,実際にはこの区間でコイ・フナ類の仔稚魚は多く観 察されたものの,ナマズの仔稚魚は少なかった.
定期採集調査でのべ136個体のコイ・フナ類の仔稚魚を採集した.Suzuki et al. (2008) は本研究の調査地である今川においてコイ・フナ類の仔稚魚を採集し,RAPD分析によ りコイ,ギンブナおよびニゴロブナの3種が含まれていたことを報告した.また,Suzuki
et al. (2008)は10回の調査を通して,コイ,ギンブナおよびニゴロブナをそれぞれ118
個体,316個体および150個体採集した.それらの体長分布の変化から,今川が産卵お よび初期成長場所として機能していることを示している.
一方,Suzuki et al. (2008)が採集した仔稚魚の中に含まれていたナマズは1個体のみで
あった.また,本調査においても,採集されたナマズの仔稚魚の個体数は 22個体であ り,コイ・フナ類の仔稚魚の個体数136個体に比べ少なかった.これに対し採集された 卵数では,コイ・フナ類よりもナマズの方が多く,約4倍であった.調査地点はナマズ が比較的多く産卵している場所であること,孵化に要する期間に違いがあることから,
卵数のみで単純な比較はできない.しかし,卵と仔稚魚の採集回数を比較しても,ナマ ズの卵と仔魚が採集された回数がそれぞれ30回と4回であったのに対し,コイ・フナ 類の卵と仔魚が採集された回数はいずれも11 回であった.このことからナマズの卵は ほとんど孵化できていない,あるいは多くが孵化直後に死滅していると考えられる.
本調査の主な対象種であるナマズおよびコイ・フナ類の卵はいずれも粘着性をもつこ と(Kobayakawa, 1985; 川那部ほか, 1989; 田崎・金澤, 2001)は知られているものの,その 粘着力の強さについて明確に比較した文献は見当たらない.一般的に,コイ・フナ類の 卵は水田などで産卵された場合,イネや雑草に粘着すると記載されることが多いのに対 し,ナマズの卵は周囲に泥をまとって目立たなくなっているなどと記載されることが多