THE ROLE OF AVENALUMIN IN THE RESISTANCE
OF OATS TO CROWN RUST
Shigeyuki MAYAMA
CONTENTS I. GENERAL INTRODUCTION
11. THE PRODUCTION O F PHY
RESPONSE T O CROWN RUST, PUCCZNZA CORONATA F SP. AVENAE
. .
4 111 CORRELATION BETWEEN AVENALUMIN ACCUMULATION A N D THESPECIFIC RESISTANCE TO INCOMPATIBLE RACES O F T H ~ CROWN
RUST FUNGUS 3
IV EFFECTS O F ELEVATED TEMPERATURE AND a-AMINOOXYACE-
TATE ON AVENALUMIN ACCUMULATION AND HYPHAL GROWTH
IN OAT LEAVES 8
V. LOCALIZATION OF AVENALUMIN A T TIS
INFECTED WITH FUNGAL PATHOGENS 3
VI. MICROSPECTROPHOTOMETRIC ANALYSIS OF CELLULAR LOCATION
ABIOTIC ELICITORS
VIII. DETECTION OF ACCELERATED ACCUMULATION OF CINNAMIC ACID DERIVATIVES, THE PRECURSORS OF AVENALUMINS, A N D THE AMOUNT OF AVENALUMINS ACCUMULATED IN THE RUSTED
OAT LEAVES 45
IX. GENERAL DISCUSSION 50
X. SUMMARY A N D CONC 5 3
ACKNOWLEDGEMENT 55
CHAPTER I GENERAL INTRODUCTION
Plants in natural condition are exposed to many potentially parasitic microorganisms and insects, in addition to physical and chemical environmental stresses Each plant and parasite interaction is a struggle for survival between two organisms; plants resist to protect themselves from the parasite invasion and parasites try to attack plants for obtaining food materials. If plants are defeated by their attacks, we will often lose our food stuffs It is evident historically that food starvation occurred many times due t o the epidemic of plant diseases [ I , 361. The resistance and/or protection of plants against parasite infections are a n unavoidable a n d earnest desire of agricultural production.
The mechanisms for resistance of plants to parasite infection have been studied for a long time [22,23,47, 11 1, 137, 138, 1401. The resistance expression can be related either to the occurrence of physical structural barriers, or to the presence of antifungal compounds in plant cells. These mechanisms are also classified into constitutive or pre-infectional resistance and induced or post- infectional resistance, according to whether a resistance mechanism functions before or after infection. Plant defences against parasites appear to depend upon these various factors The major physical features for constitutive resistance have been indicated to be the thickness or hard- ness of the cuticle and cell wall, silicification of cell walls, stomata1 closure and the presence of schlerechyma [2], and for induced resistance, tissue lignification [3, 1321 and the formation of cork and papillae [64, 1321.
More attention in plant resistance has been given to the presence of antimicrobial compounds which are present pre-infectionally or induced post-infectionally in plant cells On the basis of their presence form in plants, Ingham [53] suggested that antifungal compounds should be classi- fied into four categories:
1. Prohibitins: pre-infectional materials which reduce or completely halt the in uiuo develop- ment of microorganisms
2. Inhibitins: pre-infectional materials which undergo a post-infectional increase in order to express fully their toxic potential
3. Post-inhibitins: post-infectional materials formed by the hydrolysis or oxidation of pre- existing substances.
4. Phytoalexins: post-infectional metabolites whose forn~ation involves either gene de-repres- sion or the activation of a latent enzyme system
Induced antifungal compounds like post-inhibitins or phytoalexins are considered to be important since their production in plant cells may relate to the specific interaction between both genotypes i n host plants and parasites, which determines that a given plant is susceptible or resistant t o different pathogen strains, and that a given pathogen strain is avirulent or virulent on different host cultivars. The genetical basis for the expression of the specific resistance in host cultivars a n d fungal race interactions has been known by the gene-for-gene concept postulated by Flor [34, 351, and explained in detail by Day [22] Flor [34] summarized the evidence from flux rust in the gene-for-gene hypothesis, postulating that during their evolution host plant and parasite develop complementary genetic systems, and for each conditioning rust reaction in the host plant there is a specific gene conditioning pathogenecity in the parasite. On the expression of the genes for resistance, there is a recognition mechanism involving the interfaces between the two organisms
that deter.mines whether disease reaction leads to induced resistance or induced susceptibility. It is now understood to be the key point for determination o f t h i s fundamental biological phenomenon, "specificity" [20, 21, 93, 1381
Up to the present time, many phytoalexins whose presence was originally hypothesized by Miiller and Borger [83] in 1941 have been formed in a t least 100 plant species representing 21 families [58]. Major phytoalexins in plants whose chemical structures have been identified are listed below (Table I)..
Table 1. Major phytoalexins in plants
-. Plant Pea Soybean French bean Broad bean Clover Sweet potato Potato Tomato Cotton Red pepper Grapevine Phytoalexin (trivial name) Pisatin Glyceollin Phaseollin Kievitone Wyer one acid Wyer one Medicar pin Ipomeamar one Rishitin Phytuberin Rishitin Hemigossypol Capsidiol a-Vinifer in Chemical class isoflavonoid isoflavonoid isoflavonoid isoflavonoid f uranoacetylene fur anoacet ylene isoflavonoid ter penoid ter penoid ter penoid ter penoid ter penoid ter penoid cyclic stilbene tr imer
Orchid Or chino1 dihydr ophenanthrene
Safflower Safynol polyacetylene
Carrot 6-Methoxymellein isocoumar in
- --
Relationships between phytoalexin production and the specific expression of resistance have been demonstrated especially in the interactions of potato-Phytophthora znfestans [ 1271, soybean-phy- tophthora megasperma f sp glycznea [56], and French bean-Colletotrlchum lzndemuthianum [6, 71. These findings provide a general understanding that phytoalexin production could be associated not only with the expression of disease resistance in plants but also with a mechanism through which the specific interactions of host plant and parasite could be mediated
Despite the widespread plant taxa known to produce phytoalexins and the many reports suggest- ing the production of phytoalexin in barley [90], corn [ 69, 701 and rice [89, 1301, the involvement of phytoalexins in antifungal resistance in the Gramineae has not been established [23, 58, 651. Although benzoquinone derivatives and momilactones were recently reported as phytoalexins of barley [32] and rice plants [13, 141, respectively, evaluation of these compounds in varietal resistance is still required Efforts for identification of phytoalexins from gramineous plants are important not only to generalize phytoalexins in plant defences, but also to determine real anti- microbial agents causing the specific resistance expression in major diseases of cereal plants such a s rust or powdery mildew of wheat and barley, where the genetical basis for disease resistance has been most extensively studied [23, 31, 481
the parasite have also been well analyzed [ 109,1101, and the mechanisms for the appearance of specificity in the interactions of oat cultivars and the rust races have been a subject to be explain- ed. A series of events in oat plants along with the incompatible reaction have been analyzed with special reference to cause or effect of resistance in oat plants [118, 120, 1431 N o real antifungal agents effective for the rust invasion have yet been found a s other rust or powdery mildew diseases of cereal plants. In any case, it is essentially important to identify antifungal agents associated with the specific resistance of oat to crown rust, as a prerequisite for understand- ing of the specificity of either resistance or susceptibility in the interactions of oat cultivars and the rust races
The present study was therefore conducted to examine whether the production of phytoalexin is involved in resistance of oat plants against crown rust invasion, as a step to detect the antifungal agents As a result, possible phytoalexins of oat plants, named avenalumins I, I1 and 111, were found to accumulate in the incompatible interactions In this paper, evidences for the detection and identification of avenalumins are described and the phytopathological significance of avena- lumin production is evaluated in relation to the expression of specific resistance in oat plants against crown rust fungi
CHAPTER 11
THE PRODUCTION OF PHYTOALEXINS, AVENALUMINS, BY OAT IN RESPONSE TO CROWN RUST, PUCCINIA CORONATA I?. SP. AVENAE
Using the disease system of oat cultivar Shokan 1 which responds resistingly to race 226 but susceptibly t o race 203, it was pursued whether the production of phytoalexin was involved in the incompatible interaction The methanol extracts from the incompatible a n d compatible interac- tions as well as the uninoculated leaves were carefully analyzed to detect the antifungal compounds specific to the incompatible reaction A remarkable accumulation of three distinct antifungal compounds was found in leaves which were responding to the infection with the incompatible race. Based on the various facts which are described in this paper, the compounds were regarded a s phytoalexins responsible for antifungal resistance in crown rust of oat plants, and given the trivial name a s avenalumins I, I1 and 111 The isolation a n d the chemical and biological properties of avenalumins are described in this chapter
MATERIALS AND METHODS
Growth and inoculation
01
plants. Two oat (Avena sativa Lo) cultivars, Shokan 1 a n d a linecontaining PC 38 gene for resistance to the crown rust, were grown on vermiculite a t 20-21°C in a growth chamber under continuous fluorescent illumination a t 8,000 lux.. Primary leaves of' 7-day- old seedlings were inoculated by a spray method with uredospores of race 226 or 203 of' Pucciniit
coronata Cda, f:, sp, avenue Fraser et Led. Usually 100 mg of' spores were mixed with about 2.5 g of
talc and dusted using a hand sprayer on the primary leaves of 10 to 15 pots (9 cm in diam.), each of which contained about 30 plants. The inoculated plants were then maintained under moist conditions for 15 hours and returned t o the growth chamber thereafter.. Race 226 was incompati- ble with Shokan 1 and compatible with PC 38.. Infection types of' the two cultivars with race 203 were the exact reverse of' that with race 226. The disease reactions were assessed by measuring the lengths of' all branched infection hyphae originating from substomatal vesicles, which were stained with alcoholic lactophenol containing aniline blue a n d visible under a n ordinary light microscope.
Detectron of antijungal compounds assocrated wzth resrstance reactron Forty-eight hours after inocu- lation, the primary leaves ( 5 g fresh wt) inoculated with either race 226 or race 203 a n d the uninoculated leaves were dissected into segments after eliminating the top 1 cm of the leaves and dipped without maceration in 200 m l of hot methanol kept in a water bath (80°C) for 10 minutes The extract was filtered through a filter paper and the f i l t ~ a t e was then evaporated to dryness rn
vacuo at below 40°C The residue was dissolved in methanol and fractionated into 90 fractions
through a Sephadex LH 20 column (3
x
45 cm) with methanol as eluent. Each fraction (6 ml) was concentrated to 0 5 ml A 60~1-portion of each concentrated fraction was spotted onto silica gel plates (Merck GF,,,60; 0 25 mm) for thin layer chromatography (t 1 c ). Elution profiles of the fractions were obtained by measuring the u v -absorption a t 340nm of each spot on the t 1 c plate The t. 1 c plates were then developed with a solvent system of chloroform: methanol :water (65 : 35 : 10, v/v) The spots were detected by scanning with u v light in the range of 250 to 350 nm by a Hitachi fluorescence spectrophotorneter or by spraying with 20% sulfuric acid and heating a t 110°C for lominutes T o detect antifungal compounds specific to the incompatible interaction, antifungal activity of all the fractions developed on the t 1 c plates was screened by using the t 1 c plate bioassay method as described below
Isolation and spectral analyszs o j avenalumtns A large number of primary leaves of Shokan 1 were
inoculated with uredospores of incompatible race 226 About lO0g (fresh wt) of leaves were harvested 5 to 6 days after inoculation The methanol extract of the leaves was applied to a Sephadex LH 20 column (3 x 35 cm) and a crude fraction containing avenalumins was isolated This fraction was concentrated, and then applied t o a longen column (3 x 120 cm) of Sephadex LH 20 Fractions containing avenalumins I and 11, I and 111, II and 111 were collected by slowly eluting with methanol The individual avenalumins were then isolated on 0 35 mm t 1 c plates. The isolated avenalumins were further purified on the t 1 c and dissolved in methanol The u v. and fluorescence spectra were analyzed by a Hitachi u v spectrophotometer and a Hitachi fluorescent spectrophotometer MPF-4, respectively
Determination of chemlcal structures of avenalumrns I , II and III The chemical structures of purified
avenalumins I , I1 and III were elucidated[80]. Physicochemical analyses based on ultra violet, nuclear magnetic resonance (NMR) and mass spectra (MS) were conducted and the structures of avenalumins were proposed The proposed structures of avenalumins I , I1 and 111 were synthesized for confirmation of the structures Physicochemical data as well as antifungal activities of synthe- sized avenalumins 11, II and III were compared with those of isolated avenalumins I , I1 and 111
Bioassay for anttfungal actrvlty The antifungal compounds were detected on t 1 c plates using
uredospores of P coronata f sp avenae race 226 After the developed t I c plates were well mois- tened by spraying with water, uredospores were dispersed heavily on the plates a n d brushed to make a uniformly dense layer The plates were again sprayed with a mist of water and incubated in a dark chamber a t 20°C for 16 hours The presence of compounds active against uredospore germination as well a s germ-tube growth was recognized as orange-yellow spots contrasting to the background where the uredospores germinated and their original color became fainter
The antifungal activity of isolated and synthesized avenalumins and some phenylpropanoid compounds in aqueous solution was also assayed by the microscope-slide technique Uredospores of P coronata f sp avenae races 226 and 203 and Puccinla gramrnts f. sp trztrci Erikss et Henn. race 56 were floated on a droplet and allowed to germinate a t 20°C for 10 hours. The rate of germination and the length of germ-tube were measured
Quantrtative assay of avenalurnrn Leaf pieces of 5 cm length (1 to 6 cm portion from the leaf tip)
Sephadex LH 20 column (1
x
25 cm). The fraction containing avenalumins was diluted [with ap- propriate amount of methanol, depending on the concentration of avenalumins, to give complete separation of their spots each other on the t 1 c. The fractions were then chromatographed on the silica gel plates (0 25 mm) with the same solvent system a s above and scanned under 340 nm as exitation wavelength by the Hitachi fluorescence spectrophotometer Avenalumins were detected a s dark spots against fluorescence from the silica gel layer The amount of avenalumins separated on the t 1. c plate was estimated by u v -absorption and calculated by comparing with that of synthesized standard compound whose absorbance a t 340 nm responded linearly within the range of 0 5 to 3 0 pg The amount was expressed as pg per g fresh weight of leavesRESULTS
Detect~on o j antijiungal compounds in incompat~ble znteraction A comparison was made among the
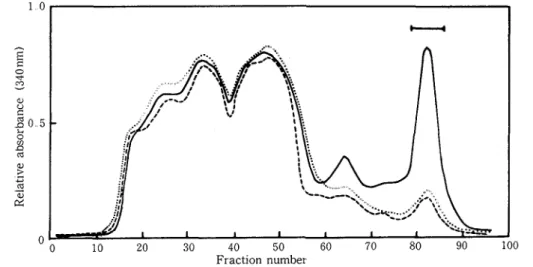
extracts of healthy leaves of Shokan 1 and the leaves inoculated with incompatible race 226 and compatible race 203 Elution profiles from the Sephadex columns bearing each of the extracts were examined under the u v wavelengths from 250 to 350nm The results indicated that u. v -absorption a t 340nm in fractions 75 to 85 was characteristic of the incompatible interaction (Fig 1) The portion corresponding to this peak was visualized on the column as a yellow-green fluorescence zone by irradiation with long wavelength u v -light and was referred to a s Fr. G
All of the fractions eluted from the column were further compared by t 1 c Three major and two minor compounds were detected in Fr G from the incompatible sample Of particular inter- est were the major spots a t Rf 0 30, 0 34 and 0 36, designated as avenalumins I, I1 and 111, respectively (Fig. 2) There was another blue-fluorescing spot detected in fractions 42 to 48 of the incompatible sample This spot was also absent in the fractions from susceptible and healthy leaves
Screening for antifungal compounds was conducted by the t 1 c. plate bioassay method on all the fractions shown in Fig. 1 from the incompatible and compatible interactions and the uninoculat- ed. Although the number of antifungal compounds detected by this method depended on the quantity of the fraction applied to the t 1. c. plates, it was clear that the major antifungal com-
Fractlon number
Fig 1 Elution profiles from Sephadex LH 20 column of methanol extracts of oat leaves (cv Shokan 1) 48 h after inoculation with incompatible race 226 (-) or compatible race 203 ( ) of Puccznza coronata f sp avenue and uninoculated control leaves (-.---) The bar indicates the fractions that contain avenalumins and is referred to as the FI G fraction
Avenalumin
Avenal.umin
Avenalumin
Time a f t e r inoculation ( h )
Fig 2 Thin-layer chromatogram of Fr G from oat leaves (cv Shokan 1) inoculated with compatible race 203 (left) or incompatible race 226 (right) of Pucclnla coronata f sp avenae Avenalumins were detected as dark spots under u v light; only the lower half of the chromatogram was photographed To demonstrate the presence of avenalumins I and I1 at 28 h after inoculation, a large amount of Fr G was applied to the t 1. c. plate; thus, avenalumins I1 and 111 are overlapped in this experiment
pounds which accumulated in the interaction were the avenalumins I , I1 and 111 The antifungal spot was also found in the fractions 42 to 48 and this antifungal spot, B1, was not found in either uninoculated or compatible race-inoculated leaves The spot fluoresced blue under eradiation with u v -light. However, antifungal activity seemed to be inferior to those of the avenalumins N o antifungal spot was detected a t Rf 0 4 5 and 0 5 5 from fractions 15 to 38 of the incompatible interaction on the t 1 c plates where the preformed antifungal substances of oat leaves, 26-desglu- coavenacosides A and B (26-DGAs) would be expected to be located
Chemical properties and structures of avenalumlns The pure avenalumins are white powders a n d
heat stable but, unlike other phytoalexins, they are highly water soluble.. The methanol solutions of the avenalumins have their maximum u. v -absorbance near 336 nm while the maximum fluores- cence emission spectra of avenalumins I , I1 and 111 are 490, 510 and 560 nm, respectively (Fig 3). The possibility that the avenalumins are present as glycosides in the leaves and they are released by enzymatic action was ruled out by the fact that homogenization of leaf' tissues did not result in the rapid release of' avenalumins. Avenalumins are highly aromatized compounds containing nitrogen and phenolic hydroxy groups.. Molecular fbrmulae were I : C16Hl,N0,, 11: CI7Hl3NO5, and 111: ClsH,,N04. Methanolysis of avenalumins showed that they are composed of' the f'ollow- ing structural moieties : 5-hydroxy-anthranilate in I, I1 and 111, p-coumarate in I , f'er ulate in I1 a n d p-hydroxyphenyl-pentadienate in 111. Acetyl derivatives of' avenalumins I , I1 and 111 were formed a n d used for spectrophotometric investigation with NMR and MS. The chemical structures of'
230 270 310 350 390
Wavelength (nm) Wavelength (nm)
Fig 3 U. v absorption (a) and fluorescence emission (b) spectra of avenalumins I (-), I I ( ) and I11 (---.--) in methanol. Excitation light for fluorescence emission, 410 nm.
0
Fig 4 The structures of avenalumins I, I1 and 111.
avenalumins were characterized as shown in Fig 4. The proposed structures were confirmed by total synthesis [80]. Avenalumin I was 2-[2-(4-hydroxyphenyl)ethenyl]-6-hydroxy-4H-3, 1-
benzoxazin-Cone Avenalumin I1 was 2-[2-(3-methoxyphenyl)ethenyl]-4H-3, l-benzoxazin-4- one Avenalumin I11 was 2-[4-(4-hydroxypheny1)-1, 3-butadienyll-4H-3, 1-benzoxazin-4-one.
Antcjungal actzvzty of avenalumzns The antifungal activity of the Fr G fractions was found to be
present only in those leaves inoculated with the incompatible race (Table 2) The reduction of germ-tube growth increased as the time after inoculation increased. Since avenalumins were the major compounds in Fr. G, inhibitory activity of isolated avenalumins was examined using two rust fungi (Table 3) Inhibition of germination and germ-tube growth was observed t o a similar extent on droplets of avenalumins I, I1 and 111 a t concentrations of 200pg/ml ar greater. About 50% inhibition of germ-tube growth was obtained a t 200-250 pglml There was no obvious differ- ence in sensitivity among P coronata f sp avenae races 226 and 203 and P gramznzs f. sp trzticl.
Table 2. Germ-tube growth of'Pucciizih coronata f sp avenae race 226 uredospores on solutions of Fr G which was extracted from oat leaves (cv Shokan 1) uninoculated and inoculated with incompatible race 226 or compati- ble race 203 of' Puccinia coronata f', sp avenae
Germ-tube length (,urn) of race 226 -
Time after
inoculation Uninoculated Inoculated
(h) Race 226 Race 203
-- (incompatible) (compatible)
28 515 354 49 7
Fr G from 1 5 g fresh weight leaves was dissolved in 1 ml of water
Uredospores were allowed to germinate for 16 h at 20°C Data are means of 100 uredospores observed in each of three replicates
Table 3 Germination and germ-tube growth of Puccin~a coronata f sp avenae and
Pucciniagramlnls f sp t r ~ t ~ c ~ uredospores on different concentrations of avenalumins
-- -
Germination (94) Germ-tube growth (pm) Concentration
(vglml) P coronata P graminis P coronata P graminis
Race 226 Race 203 Race 226 Race 203
Water control Avenalumin I 200 300 500 Avenalumin I1 200 300 500 Avenalumin I11 200 300 500
Uredospores were allowed to germinate for 10 h at 20°C Data are means of 100 uredospores observed in each of three replicates
It was confirmed that synthesized avenalumins I, I1 and 111 were also antifiingal a t equal level to those of natural avenalumins (Table 4).. The ED,, values of avenalumins I, I1 and I11 for the germ-tube growth were about 200pg/ml, respectively.. The antifungal activities of' avenalumins were then compared with those of' phenylpropanoid compounds which are possible precursors of avenalumins. The result showed that avenalumins were antifungal a t about equal level to those of' phenylpropanoids though the latter compounds showed higher activity by about 20-30% than those of avenalumins..
In the t. 1.. c. plate bioassay with the uredospores of race 226, avenalumins I, I1 and I11 gave clear spot a t the sites of corresponding Rf' values (Table 5). In this method, the minimum amount detecting the antifungal activity was 5 pg per spot for both avenalumins and the phenyl- propanoids, The acetylated derivatives of avenalumins had no antifungal activity, however.
Table 4 Antifungal activity of synthesized avenalumins I, I1 and I11 and phenolic acids against the germination and germ-tube growth of Puccinra coronata f sp avenae in the micro- scope-slide technique
- .- -- -- - Compounds Concentration Germination Germ-tube Inhibition rate
(pglml) (%) growth (pm) (%) Water contr 01 Avenalumin I Avenalumin I1 Avenalumin I11 Cinnamic acid p-Coumar ic acid Caff eic acid Ferulic acid
Table 5 Antifungal activity of synthesized avenalumins I, 11 and 111, their acetylated deriva- tives and phenolic acids in the t 1 c plate bioassay with Pucclnia coronata f sp avenae
-- pglspot Compounds -- 1 2 5 5 10 15 20 25 Avenalumin I - Avenalumin I1 - Avenalumin I11 - Acetylated avenalumin I - Acetylated avenalumin I1 -.
Acetylated avenalumin I11 -
Cinnamic acid
+
p-Coumar ic acid -
Caff eic acid
-
Ferulic acid -
Accumulation of avenalumins during the rnfection process The amount of avenalumins i n the prima-
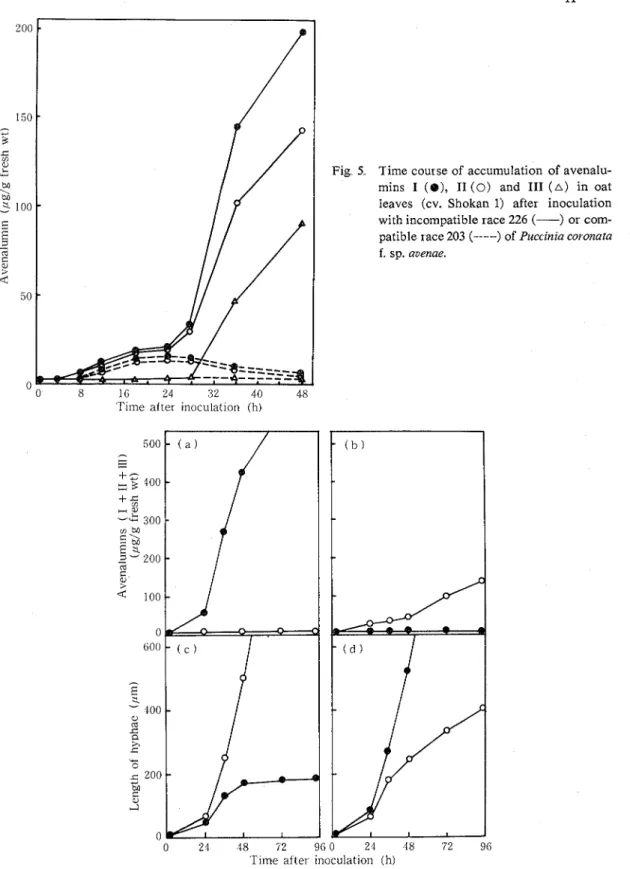
r y leaves of Shokan l following inoculation with incompatible race 226 or compatible race 203 was estimated during disease development (Fig 5) I n both incompatible a n d compatible interac- tions, avenalumins were detected a s early a s 10 t o 12 hours after inoculation, when substomatal vesicles were formed in the stomata1 cavities T h e amounts detected were very low a t this stage, a n d a slow increase was observed up t o 24 hours after inoculation for both incompatible a n d compatible interactions
Fig 5 Time course of accumulation of avenalu- mins I ( 8 ) , I1 (0) and 111 ( A ) in oat leaves (cv. Shokan 1) after inoculation with incompatible race 226 (-) or com- patible race 203 (- .--) of Puccznia coronata
f sp. avenae
'Time after inoculation ( h )
0 21 18 52 96 0 2 1 18 '72 96 'Time after inoculatior~ ( h )
Fig, 6,. A correlation between avenalumin accumulation and fungal growth in primary leaves of' oat infected with Puccinih coronata f' sp auenae xaces 226 [(a), (c)] and 203 [(b), (d)] respectively..
Race 226 is incompatible with cv. Shokan 1 ( @ ) and compatible with the PC 38 line ( 0 ) .
inoculation and avenalumin 111 after 36 hours At 48 hours after inoculation, when fungal growth ceased completely, about 400 pg of total avenalumins per gram fresh weight had accumulated. In contrast to the rapid accumulation of the avenalumins in the incompatible response, avenalumins I and I1 decreased after 28 hours in the compatible interaction and no avenalumins were detected from 2 to 6 days after inoculation. Avenalumin 111 was not detected in the compatible interaction throughout the experimental period.
The accumulation of avenalumins was also examined in the oat line possessing resistant gene PC 38 whose infection types with races 226 and 203 were the opposite of those of Shokan 1. The contents of total avenalumins increased in this oat line after inoculation with race 203 but not with race 226 The result, thus, indicated that the accumulation of avenalumins was associated with race-cultivar resistance (Fig 6) In both incompatible combinations, the time of rapid accu- mulation of avenalumins coincided with the time of retardation of growth of infection hyphae. The fact that the amount of avenalumins in leaves of the line possessing the PC gene was consid- erably lower than in Shokan 1 may reflect differences in the degree of resistance between the cultivars. In the PC 38 line, the cessation of growth of intercellular hyphae of the incompatible race occurred between 48 to 60 hours after inoculation, while in Shokan 1 it occurred between 28 and 36 hours after inoculation (Fig 6).
DISCUSSION
The data presented in this chapter demonstrate that the avenalumins are phytoalexins of oat plants. Avenalumins are extensively accumulated in the incompatible host-parasite combinations.. The time of' major accumulation of the compounds coincided with the time of cessation of fhngal growth within tissues. No major compounds other than avenalumins I, I1 and 111 seemed to participate in antifungal activity of Fr.. 6 , since the antifungal activity in Fr. G (Table 2) is rough- ly in accord with that of total avenalumins accumulated in vi'vo (Fig. 5), a s estimated by the in vitro data in Table 3 Avenalumins are unique phytoalexins in that they are highly hydrophilic and contain nitrogen Avenalumins I, I1 and I11 are the first chemically-identified phytoalexin of' cereal plants infected with rust fbngi.
It has been demonstrated that the cereals, rye, wheat and maize contain glucosides of dihydroxy- benzoxazolinone and glucosidases, which interact in homogenized tissues resulting in the rapid release of' antifungal aglucones [I351 In a wheat-stem rust system, Deverall [23] found a n antifhngal compound in wheat seedlings which had expressed to P..graminis f: sp.. tritici: The com- pound was identified as benzoxazolinone which was released fiom a glucosidic precursor in hypersensitively responding host cells.. However, formation of avenalumins is not the same as that of' the antifhngal substance described by Deverall [23], since the homogenized leaves of' uninoculated oat leaves did not produce avenalumins. Also, the slow accumulation of' avenalu- mins during the process of' infection may indicate that they are not released from immediate precursors but are synthesized in response to inkction..
In oat leaves, the preformed antif ungal compounds, 26-DGAs referred as post-inhibitin, have been shown to be activated from the immediate precursors, avenacosides A and B, by /3-glucosidases in injured leaf' tissues [55, 721 .. The possible significance of' these preformed substances has been discussed for the host-parasite interactions of' oat leaves [ 1061. In this study, however, no post-, infectional increase of 26-DGAs was detected in oat leaves which had responded to the incompati- ble race of the crown rust fungus. This suggests that the formation of' 26-DGAs from the immediate precursors is not involved in rust resistance, contrary to the case of' stem rust resistance in wheat.
It is of special interest that avenalumins are highly hydrophilic; it is thus considered, a s Daly postulated [ 191, that these avenalumins may have more contact with invading organisms in infect- ed host tissues than other reported phytoalexins which are generally low in water solubility
There are two phases of avenalumin accumulation during the infection process The first phase is the prehaustorial stage of fungal development when no visible host responses such a s cellular collapse are observed. In this phase, a small amount of avenalumins was detected in both incom- patible and compatible combinations The second phase is the period of early haustorium formation a n d the initial period of host cell collapse a t infected sites Rapid accumulation of the avenalumins is observed in this stage but only in the incompatible combinations In the compatible combina- tion, the amount of avenalumins decreased as haustorial formation proceeded It appears therefore that avenalumin accumulation could be controlled in a t least two phases during infection; one is non-specific induction a s the stage of stomata1 penetration and the other is race-cultivar specific regulation during the process of fungal development within leaf tissues
CHAPTER 111
CORRELATION BETWEEN AVENALUMIN ACCUMULATION AND THE SPECIFIC RESISTANCE TO INCOMPATIBLE RACES OF THE CROWN RUST FUNGUS As described in the previous chapter, avenalumins, the phytoalexins of oat plants, were accumu- lated in the incompatible oat-rust interaction Chemical analysis of avenalumins showed that they are highly hydrophilic, luminescent nitrogen-containing phenolic compounds, and the chemical structures were elucidated One may ask if the production of avenalumins is really associated with the mechanisms specifying the varietal resistance in the crown rust of oat
Phytoalexins have been considered to have a n important role in disease resistance on the basis of their fungitoxicity a n d the site, time and concentration of their accumulation in infected tissues in relation to restriction of fungal development [6, 7,8, 56, 104, 105, 1451 The role of avenalumins in resistance of oats to crown rust should also be evaluated in regard to those properties of phytoalexin accumulation
In oats many major genes for resistance to crown rust have been identified a n d studied by Simons and coworkers [ 109, 1101 In this study, 21 oat PC lines each possessing a single resistance gene and two crown rust races were employed. The relationship between resistance gene expres- sion and the rapidity and amount of avenalumin accumulation was examined in detail in the 42 combinations between the PC lines and fungal races The role of avenalumins in expression of PC genes against crown rust is discussed in this chapter
MATERIALS AND METHODS
Plants and fungi': Twenty-one cultivars of' oat whose genes fbr resistance to crown rust had been
identified [ 1101, and cv. Shokan 1 were employed for the present study.. The identified resistance genes and reactions to races 203 and 226 of P. coronata f.. sp, avenae are listed in Table 6. In this study, these oat varieties are referred to as
PC
lines, and are designated by the gene number instead of the code for the varieties.. PC lines 2, 14, 51, 52, 53, 57, 58, 59 and 61 were supplied by Dr. M. D. Simons, USDA, Iowa State University and PC 54, 55 and 56 by Dr. D. E.. Harder, Agri- culture Canada Research Station, Winnipeg, Canada. PC 35, 38, 39, 40, 45, 46, 4'7, 48 and 50, originally from Dr. J. W. Martens, Agriculture Canada Research Station, Winnipeg, Canada, were donated by Dr. S , Sato, Tsukuba University, Japan. All the PC lines were grown in the universityfarm of Kagawa University and the seeds were yielded and supplied for the present study. Among the PC lines, PC 35, 38, 39, 40, 45, 46, 47, 48, 50, 54, 55 a n d 56 are all near-isogenic to each other, and PC 51 and 52 are also near-isogenic (Dr M D Simons, personal communication)
Growth and znoculation of plants Dehusked seeds were soaked in water for 2 hours and incubated
on wet filter papers in Petri dishes for 36 hours About 200 uniformly germinated seeds were selected and sown on vermiculite in a planter (15x 2 5 x 10cm) and grown in a growth chamber a t 20-21°C under illumination for 16 hours daily with 10,000 lux fluorescent and incandescent lamps.. Seven day-old seedlings of each PC line were inoculated as stated in the previous chapter except that 90mg spores mixed with 3 g of talc were dusted onto the primary seedling leaves in each planter About 15% of stomata were infected by inoculation with this mixture
Obsevvatlon o j fungal development Intercellular hyphae of crown rust in infected leaves were
observed using a modification of the Calcofluor technique employed by Rohringer et a1 [ l o o ] Preliminary tests inidicated that 5 minutes staining with a Calcofluor White (Kayaphor FB R08618, Nippon Kayaku C o , L'PD Japan) solution (0 1%) facilitated simultaneous observation of infec- tion hyphae and host cell collapse as shown in stem rust-infected wheat leaves [loo] ;however, 16-20 hours staining with the fluorescence developer was necessary for complete visualization of all branched hyphae developing from substomatal vesicles of crown rust fungi Leaves stained for 20 hours were observed under a Zeiss research microscope equipped with a epifluorescence condenser IIIRS and a mercury high-pressure lamp, HBO 100 W/2 using a Zeiss HBO 100/2 power supply The filters used were as follows: BP400-440 excitation filter with peak transmittance of 400-440
Table 6 Oat varieties carrying identified genes for crown rust resistance and their infection types with races 203 and 226
- --. - .- .- -- -- . . -.--p.---p----..--...-. Infection type Variety PC gene - . .- - Race 203 Race 226 Victor.ia Ascencao D-137 CW-491-4 F-366 F-83 F-169 F-290 C. I.. 8081A F-158 CW486 Iowa X434 Iowa X421 H441 CAV 1830 CAV 4963 CAV 1964 H555 TAM-0-301 TAM-0-312 Coker 234 Shokan 1 2 14 35 38 39 40 45 46 47 48 50 5 1 52 53 54 55 56 57 58 59 61 Unknown
nm, chromatic beam splitter FT460 a n d barrier filter LP470. The lengths of stained infection hyphae developing horizontally from a vesicle were measured; the average lengths from 100 infec- tion sites were obtained for all samples and these were statistically compared
The classification of infection type was done in this investigation, partly with some microscopical observation, as follows: Immune to highly resistant: (0)-almost no visible symptom, no infection- hyphal mat, some small necrotic fleck. Moderately resistant: (1)-no uredia, few small infection- hyphal mat, necrotic or chlorotic fleck present (2)-few small uredia, plentiful small infection- hyphal mat, some necrotic areas seldom without uredia (3)-plentiful small-size uredia; necrotic areas seldom without uredia Susceptible: (4)-abundant medium to large uredia without necrosis or chlorosis
Quantztatzve estzmatlon of avenalumln accumulatzon The quantitative assay for avenalumins was
carried out a s described in the previous chapter Segments 5 c m long cut from 1 cm behind the tip of thirty infected primary leaves were used for the quantitation The methanol-soluble fraction containing avenalumins was separated on a Sephadex LH 20 column, evaporated to dryness under vacuum and usually redissolved in 1 m l of methanol The methanol-soluble fraction was then chromatographed on a silica gel GFZs4 (Merck) thin-layer plate (0 35 mm) The amount of avena- lumins I and I1 on the plate was estimated by scanning under a Hitachi fluorescence spectrophoto- meter MPF-4 Although avenalumin I11 was detected as a trace in certain PC lines, the amount was negligible as compared to avenalumins I and I1
RESULTS
Correlation between avenalumin accumulation and resistance expression in various PC llnes. Analysis of the relationship between resistance expression and avenalumin accumulation in rust-infected leaves of'the PC lines was conducted by separately inoculating with both races 203 and 226.. The experiments were done in two series, one with PC 35 to 50 and the other with PC 2, 14 and 51-61. The oat cultivar Shokan 1 which was highly resistant to race 226 and susceptible to race 203 was used in both experiments as a reference line.. The results are summarized together in Tables '7 a n d 8 for race 203 and 226 respectively. The degree of resistance of PC lines toward races 203 and 226 o f ' P . coronata f: sp.. avenue was determined by the length of intercellular hyphae and final infection type. Although hyphal length was roughly correlated with the infection type, a more detailed order of the resistance of' PC lines within the same infection type was obtained by comparing the hyphal lengths. Up to 24 hours after inoculation, no significant variance in the growth of'intercel- lular hyphae was found among the race-PC line combinations However, retardation of' hyphal growth in leaves possessing the genes PC 14, 48, 50 and 59 for race 203 and PC 48, 50, 5 1, 52 and 53 fbr race 226 was observed by 36 hours after inoculation.. At 36 and 48 hours after inoculation, average lengths of'intercellular hyphae in the PC lines diff'ered significantly and it was possible to classify the reactions approximately into three groups: immune to highly resistant, moderately resistant and susceptible.. In Tables 7 and 8, the PC lines were grouped in the order of' resistance, based on the length of intercellular hyphae a t 48 hours after inoculation and final infection types. The orders of the PC lines assessed by both markers were parallel with each other except for a few lines.. The approximate time to detect retardation of growth of' inftction hyphae in each PC line was also indicated.. Restriction of' fhngal growth in highly resistant lines was observed a t 36 a n d 48 hours after inoculation, whereas retardation of' the growth in moderately resistant lines occurred 72 to 92 hours after inoculation.. The completely susceptible lines, infection type 4, were assumed to show no retardation of'hyphal growth during the experimental period up to 144 hours.
Table 7.. Resistance as measured by three parameters and avenalumin accumulation in primary leaves of PC lines infected with Puccznza coronata f sp avenue race 203
---- - -- -- - - -
-- --
Hyphal length" Time of Avenalumins I & I1 (,~g/g fresh wt)
& growth -- --
PC lines infection type retarda- Time after inoculation (h) (in parentheses) tion (hr) 24
- - - - -- - - - 36 48 72 96 144 (Highly resistant) 14 165 (0) 36 22 83 198 238 363 401 50 178 (0) 36 25 90 190 278 347 479 5 1 189 (0) 36 12 40 113 290 196 209 53 193 (0) 36 10 63 282 351 371 463 (Moderately resistant) 38 243 (1) 72 tr tr 21 113 156 86 (Susceptible) Shokan 1 426 (4) - tr tr tr tr tr tr
a The average length (pm) of intercellular hyphae developing from each substomatal vesicle 48 h after inoculation
b The remarkable retadation of growth of the intercellular hyphae was observed at indicated time after inoculation tr =trace
to the ordei of resistance to race 203 (Table 7) and race 226 (Table 8) There appears to be a relationship between avenalumin accumulation and degree of resistance associated with various genes In lines with resistance genes causing earlier restriction of rust development, the production of avenalumins was generally more rapid and abundant than in lines with resistance genes which retarded fungal growth a t a later stage The clear relationship was evident between the highly and moderately resistant groups; the rapid accumulation of avenalumins was observed a t the early infection period of 24 to 48 hours post-inoculation in the former group, whereas little accumulation of avenalumins was detected in the latter one. In moderately resistant group, the accumulation of avenalumin was observed after 72 hours post-inoculation though the amount of accumulated avenalumins seemed to be those of the highly resistant lines. The time of rapid accumulation of avenalumins corresponded to the time of retardation of growth of infection hyphae In a strict sense of the correlation between the amount of avenalumin and the hyphal length, there seemed to be some apparent exceptions such a s for
PC
40 and 48 to race 226 andPC
35 to race 203.Table 8 Resistance as measured by three parameters and avenalumin accumulation in primary leaves of PC lines infected with Puccznia coronata f sp avenae race 226
- -- .--- --
Hyphal length" Time of Avenalumins I & I1 (pglg fresh wt) & growth
PC lines infection type retarda- Time after inoculation (h)
(in parenthesis) tion (hr) 24 36 48 72 96 144
(Highly resistant) 51 150 (0) 50 160 (0) Shokan 1 168 (0) 53 171 (0) 14 172 (0) 52 181 (0) 59 186 (0) 58 205 (0) 61 224 (0) 48 314 (0-1) (Moderately resistant) 5 7 230 (1) 55 244 (1) 2 312 (1) 40 374 (1) 56 340 (2) 35 390 (2) 54 392 (2) 46 436 (3) (Susceptible) 4 7 452 (4) 39 460 (4) 38 528 (4) 45 560 (4)
a The average length (/*m) of intercellular hyphae developing from each substomatal vesicle 48 h after inoculation
b The remarkable retardation of growth of the intercellular hyphae was observed at indicated time after inoculation tr = trace
However, the retardation of growth of the intercellular hyphae seemed to be in accord with the time of avenalumin accumulation in these interactions It should be also pointed out that several lines such a s PC 2,38, 39 and Shokan 1 responded oppositely to races 203 and 226, and avenalu- mins markedly accumulated only in the incompatible combinations Very little accumulation of avenalumins was found in any of the compatible interactions in the present study.
DISCUSSION
It was demonstrated in this investigation that in oat leaves production of' avenalumins was directly proportional to the degree of resistance expression against crown rust; in other words, susceptibility of' oat cultivars to races of' the rust was inversely related to the time and rate of accumulation of' avenalumins.. These facts strongly suggest that avenalumins may be associated with the specific resistance expression in the interactions between oat cultivars and rust races. Resistance expression by the various genes, associated with the restriction of' the growth of the
intercellular hyphae, occurred a t different times after inoculation It appears, therefore, that in the expression of the various resistance genes a single mechanism associated with the production of avenalumins could be involved in restricting fungal development At present, there is no evidence
in relation to regulation of differential time and rate of avenalumin production in the incompatible rust-oat interactions
For flax leaves infected with Melampsora lznz,Keen and Littlefield [62] presented similar results: production of phenolic phytoalexins, conyfer yl aldehyde was highly correlated with the expression of multiple allelic genes for resistance They suggested that in flax a single mechanism for expres- sion of resistance to M llnz may exist, and that the distinguishing character of the various resistance alleles is differential recognition efficiency of the incompatible fungus by plant cells. The present data obtained for crown rust of oat with the various gene combinations also suggest the presence of gene-specific recognition mechanisms that regulate differential avenalumin production, and that the production of avenalumin is regulated as a result of resistance gene expression.
CHAPTER I V
EFFECTS O F ELEVATED TEMPERATURE AND a-AMINOOXYACETATE O N AVENALUMIN ACCUMULATION AND HYPHAL GROWTH IN OAT LEAVES Attempts were further made to ascertain the role of avenalumins in resistance by altering their zn vivo concentration by growing the plants a t elevated temperatures and by treatment of the plants with a-aminooxyacetate, a n inhibitor of phenylalanine ammonia-lyase
The effect of temperature on phytoalexin accumulation has been studied [18, 56, 60, 841 ; for instance, glyceollin was not detected a t 50°C or above 40°C [56, 841 and resistance in soybean to P megasperma f sp glycznea was lost a t temperatures above 45°C [ 15, 16, 561. When cv. Harosoy soybean leaves inoculated with Pseudomonas syr ingae pv glycznea race 1 were maintained a t '31°C, no glyceollin was produced and a fully compatible reaction occurred [60] In potato tubers either infected with P znfestans [ 181 or treated with the fungal wall-released elicitor, rishitin accumulation was inhibited by incubating a t 25 to 35OC and the tubers exhibited decreased resist- ance to Phytophthora capszcl and P znfestans [ 1241 In rust diseases of cereal plants, normal resistance reactions are diminished by incubation a t elevated temperatures [ 10, 77,82, 119, 1481. In cv Shokan 1 oat leaves inoculated with race 226, resistance and hypersensitive necrosis were reduced when inoculated seedlings were incubated a t high temperatures [ 1191.
a-Aminooxyacetate is a competitive inhibitor of phenylalanine ammonia-lyase [75]. Since this enzyme is associated with the production of avenalumins, it was determined whether a-aminooxy- acetate would block avenalumin production and promote fungus development within leaf tissues.
M A T E R I A L S A N D M E T H O D S
Planfs and pathogen culture and inoculation methods: The cultivar of' oat used in the present study was Shokan 1, which is highly resistant to P.. coronata f', sp avenue race 226.. The oats were grown on vermiculite in pots (diam.. 9 cm) in a growth chamber a t 20°C by illuminating for 16 hours daily with 10,000 lux fluorescent lamps. Inoculation of 7-day-old seedlings was achieved by dusting with a mixture of' talc and uredospores and incubating in a moist chamber for 16 hours a s de- scribed in Chapter 11..
Effects ojgrowth temperature.:. 'To examine the effect of' growth temperature on avenalumin accu- mulation and hyphal growth, plants were inoculated and, after 16 hours in a dew chamber held
a t 20°C, were grown at temperatures of 15, 20, 25, 30 and 35°C The infected plants were also transferred to 30°C after the plants were incubated at 20°C for 36 or 72 hours after inoculation Treated leaves were collected at intervals during infection development and leaf segments 5 cnl long were excised I cm behind the tips and avenalumin accumulation and hyphal growth were determined a s described in earlier chapters
Treatment wzth a-amznooxyacetate ( A O A ) The infected leaves were cut a t the stems 16 hours after
inoculation and fed through their cut stems with Kasugai nutrient solution containing AOA a t the concentrations of 10-200jlM or with the nutrient culture solution alone a s control The pH of all the solutions was adjusted to 5 8 The duration of AOA feedings was 8 hours and it was carried out during the light period The leaves were then transferred to a fresh culture solution, pH 5 8 The quantification of avenalumins and observation of hyphal growth in the treated leaves were conducted a t intervals during the next 48 hours
Estzmatzon of avenalumzn content in treated leaves The quantitative estimation of avenalumin
accumulation in treated leaves was conducted as described in Chapter I1 Twenty primary leaves were extracted with hot methanol and this was fractionated by Sephadex LH 20column chromato- graphy The fraction containing avenalumins was then chromatographed on Silica gel (Merck GF,,,) t 1 c plates, where avenalumins were visualized as dark spots against background fluores- cence under a long u v -wavelength The a m w n t sf avenalumins was thus quantified by compar- ing negative fluorescence of the spots with that of a standard compound a t a n excitation wavelength of 340 nm with a Hitachi fluorescence spectrophotometer
Quantztatlon of hyphal growth and fluorescent, collapsed host cells The intercellular hyphae in
infected leaves were stained with the Calcofluor brightner for 16-20 hours The lengths of intercel- lular hyphae developing horizontally from a substomatal vesicle were measured under a Zeiss epifluorescence microscope as described in Chapter III For observation of fluorescent collapsed cells, the infected leaves were fixed with an alcoholic lactophenol solution and stained with Calcofluor for 10 minutes to acertain the penetration points of crown rust The number of col- lapsed host cells was observed by fluorescence observation using BP400-440 excitation filter (peak transmittance 400-440 nm), chromatic beam splitter FT460 and barrier filter LP470
RESUL TS
Effect of temperature on avenalumzn accumulation, hyphal growth and cellular collapse Avenalumin
accumulation was dependent on the temperature for plant growth (Fig 7a) The optimal temper- ature for avenalumin accumulation in oat plants was 20°C In oat leaves infected with race 226, avenalumin accumulation after 48 and 72 hours was largely reduced at elevated temperatures; a t 25°C the accumulation at 48 and 72 hours after inoculation was about half of that at 20°C and only traces of avenalumins were detected in plants kept a t 30 and 35°C Slightly increased hyphal growth was observed a t 25"C, but a considerable increase occurred in plants incubated a t 30°C (Fig 7b) A large mycelial colony was found in the latter tissues, but uredospores rarely formed a t 10 days after inoculation No promotion of fungus occurred a t 35°C
Avenalumin concentrations attained by incubation of race 226-inoculated leaves for 36 hours a t 20°C decreased when the plants were transferred to 30°C (Fig 8a) Only small amounts of avenalumins were detected a t 48 hours after transfer, and this reduction of avenalumin content coincided with the continuous growth of hyphae (Fig 8b) No subsequent growth of intercellular hyphae was observed when plants were transferred from 20°C to 30°C a t 72 hours after inoculation
At 20°C, hypersensitive collapsed cells were formed as early as 24 hours after inoculation and observed a t all infection sites a t 36 hours after inoculation The collapsed cells formed a t 36
Fig. '7.
Temperature
Effect of temperature on (a) the accumulation of avenalumins I (-) and I1 (----) at 48 ( 9 ) and 72 ( 0 ) h after inoculation and (b) fungus growth at the corresponding times in oat leaves (cv.. Shokan 1) infected with Puccinia coronata f. sp. avenae race 226.
0 2 1 $8 '72 96 120 l i t
Time after inoculation ( h ) 0 24 48 72 96 120 144 Fig 8. Effect of temperature shift on (a) the accumulation of avenalumins I (-) and I1 (----) and
(b) fungus growth in oat leaves (cv Shokan 1) infected with Puccinia coronataf sp avenae race 226 The infected plants were grown at 20°C ( 0 ) and transferred to 30°C ( 0 ) at 36 and 72
h post-inoculation
a n d 72 hours after inoculation a t 20°C remained after the plants were transferred t o 30°C. T h e average number of collapsed cells per infection site w a s 2.10 a t 20°C 36 hours after inoculation a n d it increased t o 2.77 a t 30°C 24 hours after the transfer..
Effect of a-amiizoox,yacetate ( A O A ) on the production of avenalumins and hyphal growth.: In vivo partial inhibition of' the production of avenalumins i n leaves inoculated with the "incompatible"
Time after inoculation (h)
Fig 9. Effect of a-aminooxyacetate on the accumulation of avenalumin I in oat leaves (cv Shokan 1) infected with Puccznia coronata f sp avenae race 226 The bar indicates the period of AOA feeding through the cut stems of infected plants. (0) AOA (200 pM), ( 8 ) control
Table 9. Effects of various concentrations of a-aminooxyacetate on inhibition of avenalumin accumulation after 48 h and fungus growth at 72 h after inoculation in oat leaves (cv Shokan 1) infected with Puccznia coronata f sp avenae race 226
Avenalumin (pg/g fresh wt) Hyphal length Treatment
I I1 & I11 Total % inhibition (pm)
Water contr 01 534 212 746
-
251AOA 10pM 135 56 191 74 -
25 pM 110 56 166 78 37 1
50 pM 97 tr 9 7 87 3 75
100pM 87 tr 87 88 693
Table 10 Effect of a-aminooxyacetate treatment on the formation of fluorescent collapsed cells (FCC) in oat leaves (cv Shokan 1) infected with Puccinia coronata f sp avenae race 226
Total number of' FCC
at 350 infection sites* % reduction in FCC Water control
AOA 10pM 25 pM 50 pM 100 pM
race was associated with AOA treatment (Fig 9) The rapid accumulation of avenalumins be- tween 36 and 48 hours after inoculation was almost completely prevented when AOA was supplied a t 16-24 hours after inoculation, after stomata1 penetration had occurred but before fungal pene- tration into mesophyll cells. At 72 hours after inoculation, the inhibition was reduced Inhibition was effective even when AOA was supplied a t low concentrations; about 80% inhibition was obtained a t 10-20pM and 90% was observed a t concentrations of 50-100 pM (Table 9) In AOA-treated infected leaves, the promotion of hyphal growth was observed 72 hours after inocula- tion (Table 9) Small mycelial colonies developed a t each infection site in the treated leaves after the initial period of infection; however, the growth was retarded thereafter when the inhibition of avenalumin accumulation was reduced (cf Fig 9) and no uredia developed Hypersensitive autofluorescent collapsed cells were first observed at infection sites a t 24 hours after inoculation. Treatment with AOA did not appreciably affect the formation of collapsed cells since only ca 20-40s inhibition was observed (Table 10)
DISCUSSION
The results in this chapter confirm the conclusions in the earlier chapters and further indicate that the production of avenalumins by oat leaves responding incompatibly to the rust race is directly involved in expression of resistance Increased susceptibility and suppression of avenalumin accumulation after temperature elevation and AOA application could result from either suppres- sion of phytoalexin production or promotion of phytoalexin degradation These treatments were applied after the determination of incompatibility as demonstrated in the incompatible race 226- Shokan 1 interaction [I201 Thus, increased susceptibility in this case implies that a metabolic control of accumulation of phytoalexins may affect disease reactions in normally incompatible host-parasite interactions where a recognition reaction towards incompatibility had already been triggered. Such a mechanism may also explain apparent cases of increased sllsceptibility in addi- tion to the type of suppression of incompatible recognition suggested elsewhere [29]. Similar experiments using a n inhibitor of synthesis of precursors of the phytoalexin glyceollin have been conducted in soybean systems The results showed that the herbicide glyphosate, a n inhibitor of the synthesis of chorismate, inhibited glyceollin synthesis in soybean leaves inoculated with P. syrcngae pv glycznea or hypocotyls inoculated with P megasperma f sp glycznea and greatly re- duced resistance expression in the soybean tissues to both pathogens [49]
In many reports with stem rust [77], leaf rust [ 111 and yellow rust [74] of wheat and crown rust of oats [96, 1201, it has been suggested that hypersensitive necrosis is not essential for inhibited rust growth The effect of elevated temperatures on the formation of collapsed cells in the leaves observed here also indicated that the promotion of fungal growth was initiated even though col- lapsed cells occurred in a similar way to that in leaves incubated a t 20°C. This observation is basically similar to those with the temperature senstive Sr 6 gene of wheat to stem rust [77], a n d implies that hypersensitively-collapsed host cells are not themselves inhibitory to the development of race 226 in Shokan 1 leaves However, the accumulation of avenalumins appears to be required for resistance expression to rust infection. The severe damage to race 226 in Shokan 1 leaves appears to occur before 72 hours after inoculation under normal conditions (cf. Fig, 8), and this may be due to prolonged exposure to the accumulated avenalumins
By transfering avenalumin-containing leaves from 20 to 30°C, it was found that the avenalmins were metabolically degraded in the infected tissues, as reported for other phytoalexins such a s rishitin [52, 541, lubimin [52], capsidiol [I131 and glyceollin [146]. It is therefore likely that the rate of avenalumin biosynthesis a t 20°C is higher than that for degradation; a t 30°C, either the
biosynthesis of avenalumin was prevented or the rate of biosynthesis was far less than that of biodegradation. The present data suggest that phenylalanine ammonia-lyase is involved in bio- synthesis of the avenalumins As shown in Fig 4, a part of the molecules of avenalumins are constructed by phenylpropanoid compounds; thus, the present results confirmed the probable involvement of phenylalanine ammonia-lyase in the biosythesis of the avenalumins.
CHAPTER V
LOCALIZATION OF AVENALUMIN AT TISSUE LEVEL IN OAT LEAVES INFECTED WITH FUNGAL PATHOGENS
It was shown in the previous chapters that the production of avenalumins could be closely asso- ciated with the expression of the genes for resistance to crown rust invasion However, the fact does not rule out the possibility that avenalumins might be accumulated in, the cells which locate far beyond the region of developing rust hyphae even though the apparent relationship was found. Thus, it is most important to know whether avenalumins occur a t infection sites where the phytoalexins might contact the infection hyphae and accumulate there into a concentration suffi- cient to inhibit the fungal development
Analysis of phytoalexin location in the micro-site of infected tissues has been carried out in detail
in C lmdemuthzanum-infected bean [ 6 , 71, P megasperma f sp. glysinea-infected soybean [56, 1451
and Po znfestans-infected potato [ 104, 1051 Sato et al [ 1041 emphasized that rishitin was synthesized in the neighboring live cells adjacent t o the infected cells and finally transferred to accumulate into the necrotic dead cells, as revealed by extractions of sliced sections of infected tubers In bean infected with avilurent races of the anthracnose fungus, phaseollin accumulation is restricted to small areas of necrotic tissues a t infection sites. If the phaseollin is in the necrotic cells alone, its concentration in the cells greatly exceeds 10 pglml which prevents germ-tube growth in vltro [7] In soybean hypocotyls, Yoshikawa et a l [ 1451 also showed by sliced sections of the infected tissues that glyceollin accumulated a t the localized infection sites of the resistant soybean hypocotyls to concentration to exceed the ED9, value for inhibition of the fungal growth
Primary leaves of oat are thin tissues and the sites of rust penetration are restricted in the sites of stomata; thus, it is impossible to estimate avenalumin accumulation in the stomatal micro-sites by slicing or excision of the infected tissues as done in those plant tissues In this chapter, there- fore, the localization and concentration of avenalumins in the stomatal sites infected with crown rust fungus were analyzed a t tissue level without making the sliced sections to demonstrate if avenalumins accumulated are effective for preventing the hyphal growth m vlvo. It was also analyed whether avenalumins might accumulate in the epidermal cells of oat leaves infected with other fungal pathogens that could penetrate through the cuticular layer.
MATERIALS A N D METHODS
Plants and fungi The incompatible combination between oat cv Shokan 1 and crown rust race 226 was used for the present study. Pyricularia or yzae and Pyricularla gr lsea, non-pathogens of oat plant, and Drechslera avenae were also employed a s pathogens which infect oat leaves through cuticular layer The conidia of Pyricularla fungi and D. avencze were obtained by culturing on oat-meal agar and V-8 juice agar media respectively.
Growth and lnoculat~on of plants The plants were grown a s mentioned before and used for inoculation 7 days after sowing T o obtain different rates of stomatal infection by the fungus,
inoculation was made by dusting various densities of spores diluted with talc powder The number of stomata penetrated by the fungus was counted for both abaxial and adaxial surfaces of leaves, and expressed as percentage of total stomata In order to assess the extent of avenalumin accu- mulation in infected and neighboring uninfected area, a partial inoculation was made A 1 cm- long region, 3 to 4 c m from the tip, of the highly resistant Shokan 1 leaves was inoculated by brushing uredospores of race 226 onto the abaxial surface The rate of stomatal infection in the inoculated surface was adjusted to about 10 and 30%
The conidia of the saprophytic fungal pathogens were suspended in water and used for inocula- tion of the abaxial surface of the leaves by brushing After inoculation for 14 hours in a moist chamber, the plants were placed and grown in the growth chamber
Quantztatzve estzmatzon of avenalumzn accumulatzon The estimation of avenalumins was carried out a s described previously In the case of the partial inoculation, the inoculated region and the next two 1 cm regions towards the tip were excised 36 hours after inoculation; fifty excised segments for each sample were weighed and used for the avenalumin estimation
For the leaves infected with the saprophytic fungal pathogens, the lower layers were peeled out from the infected leaves 24 hours after inoculation The epidermal strips and the remaining epidermis-stripped leaves were supplied separately for estimation of avenalumins The concentra- tions of avenalumins accumulated in the stomata1 area infected by the rust fungi and the epidermal cells infected by the saprophytic fungal pathogens were estimated by dividing the amout of avenalumins accumulated in the infected tissues by the volumes of each infection sites, a s described in the result section
Detectzon of avenalumzn zn zntercellular fluid The intercellular Auid from rust-infected primary
leaves of oat was collected by the method reported by Rathmell and Sequeira [98] Sixty infected leaves were collected a t intervals after inoculation and cut into 3 cm segments eliminating the tip 1 5 c m The cut ends were rinsed in water and the segments were wrapped in a layer of cheese cloth and then immersed in distilled water in a test tube Water was infiltrated zn vacuo for three periods of one minute each The leaf pieces were then removed from water and unwrapped and their surfaces were blotted dry with filter paper The leaf segments were again wrapped in cheese cloth, placed length-wise in a Spitz glass tube and centrifuged a t about 6 0 0 x g for 10 minutes. About 0 2 m l of intercellular fluid was obtained in the bottom of the tube Avenalumins contained in the fluid were separated by Sephadex LH 20column chromatography (1
x
25 cm) and quantified by t I c and fluorescence spectrophotometry a s described above. The enzymatic activity of glucose-6-phosphate dehydrogenase in the fluid was assayed as described by Rathmell and Sequeira [98] in order to determine leakage of cell contents into the intercellular spacesRESULTS
Dependency of avenalumin accumulation on znfectzon rate through stomata The experiments were
made with the Shokan 1-race 226 system AvenaIumins accumulated in the partly-infected por- tion of the primary leaves and none to little in the neighboring uninfected tissues (Fig 10) The results also indicated that the extent of avenalumin accumlation in the inoculated area was higher in leaves with higher infection rate In a subsequent experiment, amount of avenalumin was analyzed in relation to stomatal infection rates (Fig 11) The results clearly showed that avena- lumin accumulation in the infected leaves a t 36 hours after inoculation was directly related to the proportion of stomata infected up to 50% The correlation coefficient between infection rate and the amount of avenalumins was 0.98, and the regression line for the data of Figure 11 was Y =