育すう初期の成長抑制がメスウズラの卵巣発育と体成分に及ぼす影響
山本義弘,橋口峰雄Effect of body growth during the early growing period on ovary development and body
composition of female Japanese quail
Yoshihiro YMAMOTO and Mineo HASHIGUCHI
Abstract
The body weights of female Japanese quail were regulated for 2 weeks from 7 days of age so that there became less weight 0, 10, 20 and 30% than a normal female quail, and thereafter quail were fed a diet with free access until sexual maturity. At sexual maturity, body growth, ovary growth and body composition were investigated. Age at sexual maturity was later for 20% and 30% groups than for 0% and 10% groups, but ovary weight and the size of ovarian follicle were not changed due to depressed body weight in early growing period. There was no difference in body fat deposition between groups, but body weight and body protein deposition were less for 20% and 30% groups than for 0% and 10% groups.
Key Words: Follicle growth, Sexual maturity, Body composition, Ovary,
緒 言 家禽における産卵機能は卵黄(卵子)と卵形成を経て 現れる.ニワトリにおいて産卵能力を発揮させるために 育成中の成長や体成分蓄積の調節を行うことがなされて いる.肉用鶏(1,2)と卵用鶏(3)において育成期の体重を 抑制すると,性成熟の発現は遅くなることが示されてい る.また育成期において体重の軽い産卵鶏は性成熟が遅 れるが(4,5),体重の重い鶏は性成熟が早くなる(6).こ のように育成期の体成長の調節は性成熟の発現に影響す ると考えられる.しかしながら,育成期の体成長抑制が 性成熟時の卵胞発育に対して影響するのかは良く知られ ていない.一方,家禽における性成熟の発現には体成分 がある一定以上になることが必要であるとの示唆がなさ れているが(7),体タンパク質と脂肪の蓄積と卵胞発育 との関係については明らかにされていない.肉用鶏にお いて育成期の成長抑制は性成熟時の体タンパク質を変化 させないまたは減少させると(8)報告されている.また, 育成初期の成長を抑制した場合,性成熟時の体脂肪蓄積 に関しても一致した見解は得られていない(2).本実験 ではメスウズラにおいて育成初期の成長抑制が卵巣の卵 胞発育に対して影響するかどうかを体成分の蓄積と関連 して検討した. 材料および方法
本実験では,68羽の日本ウズラ(Coturnix cotornix
ja-ponica)のメスを使用した.1日齢から28日齢まで保温 器付きのバターリ育すう器で飼育し,その後性成熟に達 するまで産卵用の単飼ケージで飼育した.育すう温度は 慣行法に従い,調節した.照明は1日齢では連続照明と し,以後は,1日16時間照明(4:00∼20:00)8時間暗 黒とした.なお,本実験は香川大学農学部実験動物の取 扱いに関するガイドラインに従い行われた. メスウズラを7日齢に17羽ずつの4群に分け,21日齢 時の体重を対照区(0%区)より10%,20%および30% 抑制するように設定した10%区,20%区および30%区を 設けた.それぞれの区のウズラに育成飼料を給与し,7 日齢から21日齢までの飼料給与量は21日齢時の設定体重 になるように体重の推移をみながら調整した.また,21 日から性成熟までは飼料を自由に摂取させた.供試育成 飼料は粗蛋白質を24%,代謝エネルギーを2900kcal/kgを 含むように調整し,用いた飼料原料の配合割合はトウモ ロコシ57.72%,大豆粕32.67%,魚粉7.00%,コーンオ イル0.61%,DL-メチオニン0.09%,食塩0.21%,第三燐 酸カルシウム0.15%,炭酸カルシウム1.27%およびビタ ミン・ミネラルプレミックス0.28%であった.なお,飼 香川大学農学部学術報告 第60号 87∼90,2008
OLIVE 香川大学学術情報リポジトリ
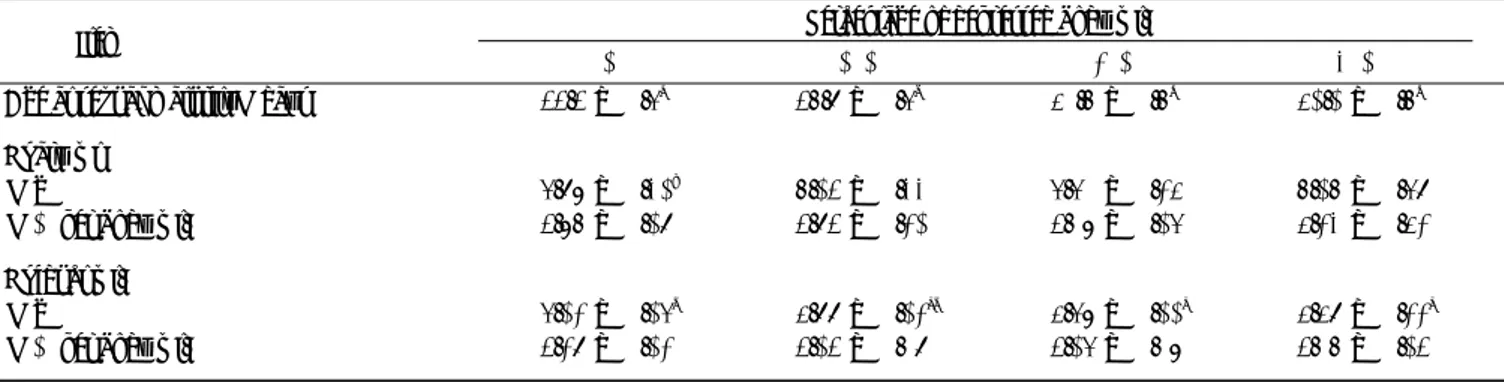
Tech. Bull. Fac. Agr. Kagawa Univ., Vol. 60, 2008 料中のビタミン,ミネラルおよび必須アミノ酸の量は NRC(9)の養分要求量を満たすように配合設計した. 性成熟時(ウズラが最初の卵を産卵した日)に体重を 測定した後,エーテル麻酔し断頭によりと殺した.脱羽 後直ちに開腹し,卵巣において閉鎖卵胞がないことと腹 腔内に卵ついしていないことを確認した.その後腹腔内 脂肪,卵巣,卵管およびと体を採取し,それらの重量を 測定後直ちに冷凍保存(−29℃)した.なお卵巣につい ては最大卵胞から第6卵胞まで採取し重量の測定を行 い,卵胞の発達を調べた.冷凍保存したと体は解凍し, 胸筋と骨格成長の指標である脛骨を採取し,重量と長さ をそれぞれ調べた. と体のタンパク質と脂肪量の測定は各区の性成熟時体 重が平均値に近い6羽を選び行った.と体(肝臓,卵巣 および卵管を除く)は解凍後ミンチにし,さらにハイパ ワーホモゲナイザー(増田理化製)でホモゲナイズした. と体の蛋白質と脂肪の量はホモゲナイズした試料から2 サンプルをそれぞれ採取しケルダール法(10)とF OLCH et al.の方法(11)に従い測定した. 得られた結果は一元配置の分散分析を行い,また 平均値の差の検定はFISHERのPLSD法によって行った (P<0.05). 結果および考察 育成初期に成長を抑制したメスウズラの性成熟日齢を 調べたところ(表1)性成熟日齢は21日齢時体重を20% と30%抑制した区が0%区(対照)および10%区より有 意に遅くなり,また10%区は0%区より遅延することを 示し,性成熟の発現は育成期の成長抑制率が高くなるほ ど遅延することが明らかになった.家禽において性成熟 と育成期の成長との関係はいくつかの報告がなされてお り,育成期の体重抑制は性成熟を遅延させることが肉用 鶏(12)と産卵鶏で(3)報告されている.性成熟は卵巣にお ける卵胞の発育によって支配されているので,育成初期 の成長抑制による性成熟の遅延は卵巣における卵胞発育 の遅延を示している.卵管重量は育成初期の体重抑制に より減少し,30%区が0%区より有意に減少したが,性 成熟時の卵巣重量は育成初期の体重抑制により変化しな かった(表1).育成期の体重と卵巣の発育について, 卵巣は育成期の体重が重いほうが発育していることが産 卵鶏(4)とウズラ(13)において報告されている.一方,肉 用鶏において卵巣重量は育成期の体重によって変化しな いことが示され(12),この結果は本実験の結果と一致し ていた.また,卵巣の卵胞の発育と育成期の体重制御と の関係については報告がなされていないので,本実験で は卵胞のうち最大卵胞から第6番目の卵胞の発育を調べ たところ(図1),それぞれの卵胞の発育は育成初期の 体重抑制によって違いは認められなかった.よって,ウ F1 F2 F3 F4 F5 F6
Position in follicle seguence 3 2 1 Follicle wt. ( g )
Fig. 1 Follicle weight in each position of follicular se-quence in ovary of female quail depressed body growth during the early growing period (Mean ± SE. n = 17). F1 ∼ F6 = The largest follicle ∼ the sixth largest follicle. % = percentage of depressed body weight.
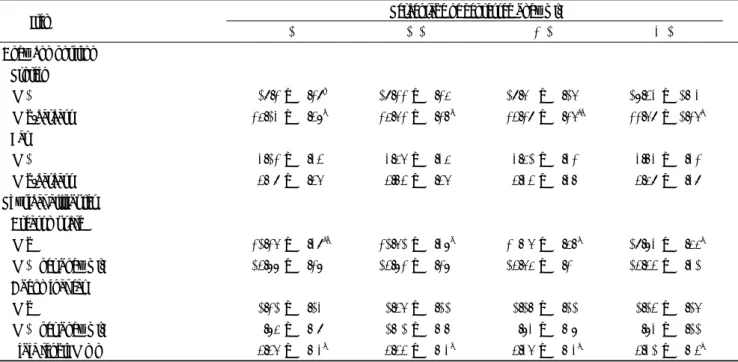
Table 1 Age and reproductive organ weight at sexual maturity of female quail depressed body growth during the early growing period
Item Percentage of depressed body wt.
0% 10% 20% 30%
Age at sexual maturity(days) 44.4 ± 0.6a 47.9 ± 0.6b 50.7 ± 0.7c 51.1 ± 0.7c
Ovary wt (g) 6.98 ± 0.31* 7.14 ± 0.33 6.60 ± 0.25 7.17 ± 0.69 (% per body wt.) 4.87 ± 0.19 4.94 ± 0.21 4.08 ± 0.16 5.23 ± 0.52 Oviduct wt. (g) 6.12 ± 0.16b 5.99 ± 0.12ab 5.68 ± 0.11a 5.59 ± 0.22a (% per body wt.) 4.29 ± 0.12 4.14 ± 0.09 4.16 ± 0.08 4.07 ± 0.14 *Mean±SE(n=17)
Means followed by different letter are statistically significant at 5% level. 88
山本義弘,橋口峰雄:体成長抑制が卵巣発育と体成分に及ぼす影響
Table 2 Body weight at sexual maturity of female quail depressed body growth during the early growing period
Item Percentage of depressed body wt.
0% 10% 20% 30%
Body wt. at sexual maturity(g) 143.0 ± 1.6bc 145.5 ± 1.9c 137.0 ± 2.1a 137.3 ± 2.3ab
Body wt. at stat(g) 62.7 ± 0.6 56.7 ± 0.4 49.6 ± 0.3 41.2 ± 0.7
Body gain(g) 80.3 88.8 87.4 96.1
*Mean±SE(n=17)
Means followed by different letter are statistically significant at 5% level.
Table 3 Body composition and physical attributes at sezual maturity of jemale quail depressed body growth during the early growing period.
Item Percentage of depressed body wt.
0% 10% 20% 30% Body composition Protein (%) 19.2 ± 0.29* 19.22 ± 0.25 19.60 ± 0.16 18.43 ± 1.03 (g/carcass) 25.13 ± 0.48b 25.62 ± 0.27b 24.29 ± 0.26ab 22.69 ± 1.26a Fat (%) 3.12 ± 0.34 3.56 ± 0.35 3.51 ± 0.32 3.73 ± 0.32 (g/carcass) 4.09 ± 0.46 4.74 ± 0.46 4.34 ± 0.37 4.59 ± 0.39 Physical attributes Breast muscle (g) 21.26 ± 0.39ab 21.61 ± 0.38b 20.06 ± 0.57a 19.83 ± 0.55a (% per body wt.) 14.88 ± 0.28 14.82 ± 0.28 14.65 ± 0.20 14.44 ± 0.31 Abdominal fat (g) 1.21 ± 0.13 1.46 ± 0.11 1.17 ± 0.11 1.14 ± 0.16 (% per body wt.) 0.85 ± 0.09 1.01 ± 0.07 0.83 ± 0.08 0.83 ± 0.11 Tibia length(mm) 4.46 ± 0.03b 4.44 ± 0.03b 4.36 ± 0.03a 4.31 ± 0.04a
*Mean±SE(n=6 in body composition, and n=17 in physical attributes) Means followed by different letter are statistically significant at 5% level.
ズラにおける育成初期の体重抑制は性成熟に達するまで の卵胞の発育を遅延させるが,性成熟に達した卵巣の卵 胞発育には影響しないものと思われる. 表2に示すように,性成熟時の体重は0%区と10%区 の間に差がなかったが,20%区と30%区は0%区と10% 区に比べ低い傾向を示した.また,21日齢から性成熟ま での増体量は体重抑制を行った区が0%区より多くなっ た.性成熟時の体重が軽かった20%区と30%区は性成熟 の遅延が認められ(表1),性成熟の発現に性成熟時の 体重が関係していることが考えられる.このことについ て,BRODY et al.(14)が主張しているように,性成熟の開 始にはある一定以上の体重に達することが必要であると すれば,本実験条件では性成熟に必要な体重は約137g となる.しかしながら,性成熟日齢の早かったウズラの 体重は143gと145gであり性成熟に達する前に137gに 到達していたことになる.このように性成熟に必要な体 重に達した時点で性成熟が発現しなかったのは,BRODY et al.(7)が主張する体重以外の要因も性成熟に関係して いることが推察される. 本実験において体タンパク質量は20%区と30%区が 0%区と10区より有意に低いかまたは低い傾向を示し (表3),育成初期の成長抑制は体タンパク質の蓄積を低 くすることが明らかになった.また,頸骨の長さも体タ ンパク質と同様な関係がみられ,育成初期の成長抑制は 骨格成長を抑制することが認められた.本実験において 性成熟は20%区と30%区が0%区と10区に比べ遅くなる ことを示したが(表1),この性成熟日齢の遅延は育成 初期の成長抑制による体たんぱく質の蓄積または骨格成 長の抑制が関係しているかもしれないが,この点につい て更なる検討が必要である.一方,体脂肪量と腹腔内脂 89
OLIVE 香川大学学術情報リポジトリ
Tech. Bull. Fac. Agr. Kagawa Univ., Vol. 60, 2008 肪量は育成初期の成長抑制によって違いが認められな かった(表3).このように,育成初期に成長を抑制し ても性成熟時の体脂肪蓄積に変化がみられないことは Brody et al.(14)が述べているように性成熟の発現には体 脂肪のある一定量の蓄積が必要であることを示唆してい るのかもしれない. 摘 要 メスウズラにおける育成初期の成長抑制が卵巣の卵胞 引 用 文 献
⑴ ROBBINS, K.R., MCGHEE G.C., SEI, P.O. and BEAUCHENE
R.E.: Effect of restriction on growth, body weight, and egg production of broiler females through 68 weeks age. Poultry Science, 65, 2226-2231 (1986).
⑵ FATTORI,T.R., WILSON H.R., HARMS, R.H., MATHER F.B.,
MILES, R.D. and BUTCHER G.D.: Response of broiler
breeder females to feed restriction below recommended levels. 3.Characterizing the onset of sexual maturity. Poultry Science, 72,2044-2051 (1993).
⑶ HURWITZ, S. and PLAVNIK, I.: Severe feed restriction
pul-lets during the early growing period: Performance and relationships among age, body weight, and egg weight at the onset of production. Poultry Science, 68,914-924 (1989).
⑷ RENDEN, J.A., BENOFF, F.H., WILLIAMS, J.C. and B
USH-ONG, R.D.: Examination of the physical characteristics in
diverse group of dwarf white leghorn pullets before and after first oviposition. Poultry Science, 69,16-26. (1990)
⑸ BENOFF, F.H. and RENDEN, J.A.: Divergent selection for
mature body weight in dwarf white leghorns. 1. Growth and reproductive responses to selection. Poultry Science, 62,1931-1937 (1983).
⑹ CUNNINGHAM, D.C. and MORRISON, W.D. : Dietary
en-ergy and fat content as factors in the nutrition developing egg strain pullets and young hens. 1. Effect on several parameters and body composition at sexual maturity. Poultry Science, 55, 85-97 (1976).
⑺ BRODY ,T.B., SEIGEL, P.B. and CHERRY, J.A.: Age, body
wight and body composition requirements for the onset sexual maturity of dwarf and normal chickens. British Poultry Science 25, 245-252 (1984).
⑻ RENEMA, R.A., ROBINSON, F.E., NEWCOMBE, M. and
MCKEY, R.I.: Effect of body weight and feed
alloca-tion during sexual maturaalloca-tion in broiler breeder hens. 1. Growth and carcass characteristics. Poultry Science, 78, 619-628 (1999).
⑼ NRC: National Research Council. Nutrient requirements of poultry. National Academy of Science, Washington DC. (1994).
⑽ AOAC: Official Methods of Analysis. 13th edition.
Wash-ington, DC, Association of Official Analytical chemists (1980).
⑾ FOLCH, J., LEES, M. and SLOANE-STANLEY, G.H.: A
simple method for the isolation and purification of total lipids from animal tissue Journal Biological Chemistry, 226, 497-509 (1957).
⑿ FATTORI, T.R., WILSON, H.R. HARMS, R.H., MATHER, F.B.,
MILES, R.D. and BUTCHER, G.D.: Response of broiler
breeder females to feed restriction below recommended levels. 3. Characterizing the onset of sexual maturity. Poultry Science, 72, 2044-2051 (1993).
⒀ OGUZ, I., ALTAN, O., KIRKPINAR, F. and SETTER, P.: Body
weight, carcass characteristics, organ weights, abdominal fat, and lipid content of liver and carcass in two lines of Japanese quail (Coturnix coturnix japonica), unselected and selected for four week body weight. British Poultry Science, 37, 579-588 (1996).
⒁ BRODY, T., EITAN, Y., SOLLER, M., NIR, I. and NITSAN,
Z.: Compensatory growth and sexual maturity in broiler females reared under severe food restriction from day of hatching. British Poultry Science, 21,437-446 (1980).
(2007年10月31日受理) 発育および体成分に及ぼす影響を調べるために,21日齢 時の体重を0%,10%,20%および30%に抑制した4つ の群を設け,性成熟まで飼料を自由に摂取させた.性 成熟日齢は20%と30%区が0%と10%区より遅くなった が,卵巣重量および卵胞の大きさは体重抑制によって変 化しなかった.また性成熟時体重は20%と30%区が0% と10%区より軽くなった.また,体脂肪蓄積は育成初期 の成長抑制により影響されなかったが,体タンパク質蓄 積と脛骨の長さは20%と30%区が他の区より少なくなっ た. 90