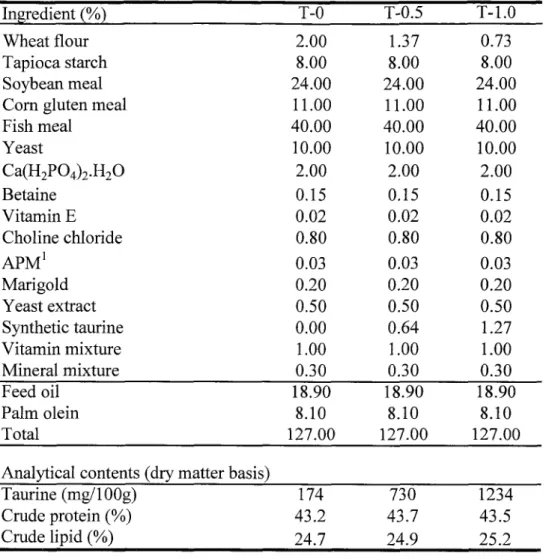
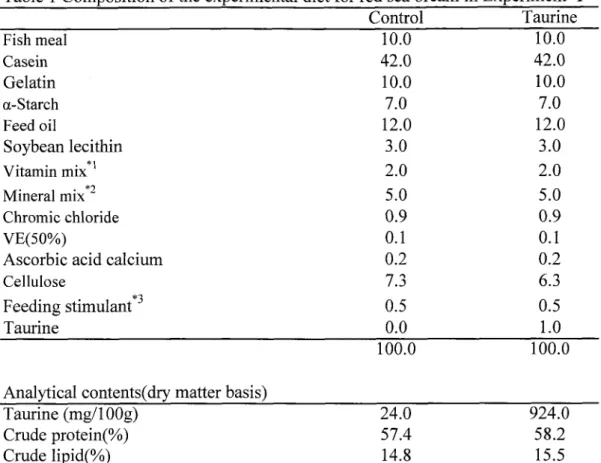
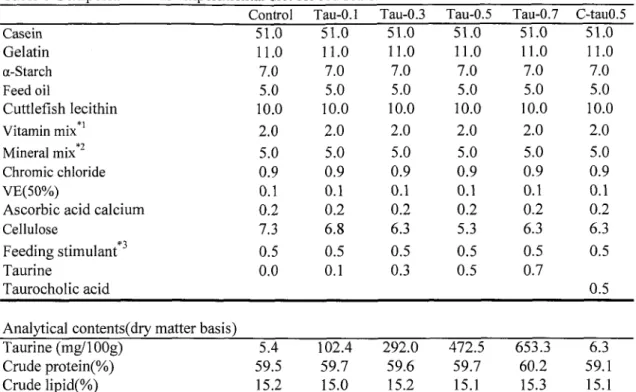
TUMSAT-OACIS Repository - Tokyo University of Marine Science and Technology (東京海洋大学)
海産魚類のタウリン要求とその役割
著者
松成 宏之
学位授与機関
東京水産大学
学位授与年度
2006
URL
http://id.nii.ac.jp/1342/00000708/
博士学位論文
海産魚類のタウリン要求とその役割
じ
サ ’籟平成18年度
(2007年3月)
東京海洋大学大学院
水産学研究科
資源育成学専攻
松成宏之
〔課程博士〕
博士論文内容の要旨
(2,000字程度)報告番号
課博第 号
氏 名松成 宏之
「海産魚類のタウリン要求とその役割」
タウリンはタンパク質を構成する成分ではなく、生体内に遊離状態で存在するアミノ酸
である。これまで魚類はタウリンを生合成できるため、餌・飼料中にタウリンは必要ない
とされてきた。しかしながら近年、海産魚類ではタウリンの合成能が微弱あるいは存在し
ないため、餌・飼料中にタウリンを要求する可能性が示唆された。本研究は、ブリやマダ
イなどの産業上重要な海産養殖魚類のタウリンの要求量とその役割を明らかにすることを
目的として、タウリンを強化したワムシおよびタウリンを添加した配合飼料を海産魚に給
与し、成長や体成分に及ぼす影響について検討した。また、魚粉を一切含まない海産魚用
の精製飼料の開発を試みた。第1章では、海産仔稚魚のふ化後の経過目数に伴うタウリン含量の変化を明らかにする
とともに、海産仔魚に対するタウリン強化ワムシの効果について検討した。人工採卵・ふ
化により得られたふ化仔魚から全長18cmまで飼育したブリ仔稚魚や、天然で採取した稚魚
とともに、人工種苗生産魚および天然魚が摂餌している餌・飼料のタウリン含量を測定し
た。その結果、天然魚は人工種苗生産魚の2∼4倍のタウリンを含有すること、ふ化直後に
タウリン含量が急激に減少すること、さらに天然魚の餌料(コペポーダ)と比較して人工
生産魚に給与する餌料(ワムシ)のタウリン含量は著しく低いことが明らかになった。こ
れらのことから、タウリンはブリ仔魚の成長に重要な役割を担うとともに、種苗生産で使
用する餌・飼料ヘタウリンを強化することにより、健全な種苗が得られる可能性が示唆さ
れた(第1節)。次に、ワムシヘのタウリン強化方法を検討したところ、ワムシ培養水槽へ
のタウリン添加量および強化時間に比例して、ワムシ中にタウリンが蓄積された。これら
のタウリン強化ワムシをふ化直後のマダラ仔魚に給与したところ、仔魚の成長が促進され
た。以上のことから、タウリンはふ化仔魚の発育に重要な栄養素であることが示唆された
(第2節)。るだけでなく、魚体中のタウリン含量は飼料中のタウリン含量の影響を大きく受けること
、が明らかになった。さらに遊離アミノ酸組成からタウリンを生合成する酵素活性が低いこ
とが示唆された(第1節)。親魚においては、飼料ヘタウリンを添加することにより、成熟
が促進されるとともに採卵量が増加し、さらに卵の受精率やふ化率が向上することが明ら
かとなった(第2節)。第3章では、マダイ稚魚に対する精製飼料中のタウリンの効果を検討した。一般的な養
魚用配合飼料には、主なタンパク質源として魚粉が5割以上含まれるため、タウリンの要
求性に関する研究には適さなかった。そこで、タンパク質源としてタウリンを含まないカ
ゼインを用いた飼料を新たに開発し、タウリンの添加量を調整した飼料をマダイ稚魚へ給
与した。これまで、海産魚はカゼイン飼料の摂餌が劣ると考えられていたが、開発した精
製飼料はマダイ稚魚の摂餌が極めて良好であった。低タウリン飼料をマダイ稚魚に給餌す
ると体色の暗色化や成長停滞などの異常がみられたが、タウリンを添加することにより、
これらの症状が改善された。さらに、飼料中のタウリンおよび含硫アミノ酸の1っである
シスチンがマダイ稚魚の摂餌行動に及ぼす影響について検討した結果、タウリン添加区で
は対照区と比較して、飼料を視認後、餌を飲み込んだ回数(摂餌回数)が有意に高くなる
こと、一方、シスチン添加区では飼料を視認後に餌を飲み込むまでに、1回以上吐き出す回
数が有意に高くなることが明らかになった。また、シスチン添加区では魚体中のタウリン
含量が増加しないことから、マダイは体内でシスチンをタウリンヘ十分量代謝できないこ
とが明らかになった(第1節)。次に、マダイ稚魚におけるタウリン要求量および脂質の消
化吸収におけるタウリンの役割について検討した結果、飼料中に約0.5%のタウリンを添加
することにより成長が促進されることから、マダイ稚魚は体内で十分量のタウリンを合成
・できず、マダイ稚魚は飼料中に約0.5%のタウリンを要求量することが明らかになった。ま
た、飼料へのタウリン添加により抱合胆汁酸濃度が上昇するとともに、肝臓中の粗脂肪含
量が有意に高い値を示したことから、飼料中のタウリンが脂質の消化吸収に影響を及ぼす
可能性が示唆された(第2節)。
以上の結果から、タウリンは数種の海産魚類において重要な栄養素であることが明らか
になった。今後、仔魚から稚魚期までの餌・飼料の改善を図ることが可能となるばかりで
なく、魚粉に代わるタンパク質原料の積極的な利用と、環境負荷低減飼料の開発に大きく
貢献するものと考えられる。さらに、海産魚用精製飼料が開発されたことから、海産魚に
海産魚類のタウリン要求とその役割
目 次緒 言
頁 ・1第1章 海産仔稚魚のタウリン含量とタウリン強化ワムシによる給与効果
第1節 人工種苗生産ブリ仔稚魚におけるタウリン含量の変化および天然稚魚と
の比較 (目本水産学会誌2003169,757−762,)・・・・・・・・・・・… 16
第2節 マダラ仔魚の成長および体組成に及ぼすタウリン強化ワムシの効果
(∠9伽ou1甜丁θ501θ刀oθ2005;53,297−304。)・・・・・・・・・・・・… 35
第2章 ブリ用配合飼料へのタウリン添加効果
第L節 飼料中のタウリンがブリ稚魚へ及ぼす影響
(乃訪θガθs501θ刀oθ2005;71,1131−1135.)・… ・63第2節 ブリ親魚の産卵に及ぼすタウリン添加飼料の影響
(躍shθガθ550∫θ刀oθ2006172,955−960.)・・・・・… ・82第3章 マダイ稚魚に対するタウリン添加精製飼料の影響
第1節 飼料中のタウリンおよびシスチンが
マダイ稚魚の摂餌行動に及ぼす影響・・・・・・・・・・・・… 103
第2節マダイ稚魚のタウリン要求量および胆汁酸への影響・・・・・・… 123
総 括
・140
謝 辞
・147
緒言
我が国の漁業生産量は1988年を境に急激に減少している。特に遠洋漁業や沖合漁
業では、国際的な200海里体制の定着及び公海漁業への規制強化や資源状態の悪化に
より、漁獲量はピーク時の半分以下となっている。このような状況の中で、つくり育
てる漁業などの適正な資源管理および増養殖手法により、わが国およびその周辺の水
域において持続的に資源を利用する必要性は極めて大きいと考えられる。つくり育て
る漁業は、栽培漁業と養殖業に大別される。栽培漁業は、水産動物の減耗が最も激し
い卵から仔稚魚の時期を人間の管理下において種苗を生産し、これを天然水域へ放流i
した上で適切な管理を行い、対象とする水産資源の持続的な利用を図ろうとするもの
である。一方、養殖業は、人工あるいは天然から入手した種苗を収穫まで人間が管理
する方法である。栽培漁業は、1963年に重要な水産資源を増やすために、魚介類の種苗生産・放流を
中心に瀬戸内海をモデル地域として始められた。その後、約40年の間に、文橡魚種の
拡大や生産尾数の増加などの著しい進歩があり、2000年には種苗放流対象魚種は約80
種、マダイ、ヒラメをはじめとする9種においては年間1,000万尾を超える種苗が放
流されている(水産庁・目本栽培漁業協会,2002)。現在では、このような生産尾数な
どの「量」から、種苗の形態や行動などの「質」の向上を図ることも求められている。種苗の質については、「健苗性」や「種苗性」の言葉で表わされる(塚本 1990)。健
苗性とは、形態的、生理的、および生化学的に健全であることを意味する。一方、種
苗性とは、種の特徴的な行動生態とそのための機能が十分に発達した種苗の質と定義
ヒラメ稚魚の放流効果を向上させるためには、人工種苗の種苗性について検討する必
要あることが報告された(古田,1998)。天然のヒラメ稚魚は、着底直後から全長100㎜ 前後に至るまで主にアミ類を摂餌することが知られおり(Subiyant et al.,1993)、朴ら(1997)はヒラメ稚魚の健全性の向上を目的に、天然ヒラメ稚魚の主な餌料生物で
あるアミの栄養価について検討し、アミ類の重要な栄養成分として遊離アミノ酸の有
効性を明らかにした。さらに結晶アミノ酸を用いて遊離アミノ酸中のタウリンがヒラ
メの成長促進の効果があることを明らかにしている(朴ら,2001)。
タウリンは体構成タンパク質ではなく、生体内に遊離状態で存在するアミノ酸であ
り、浸透圧調整や神経伝達調整作用などの機能が知られている(Huxtable,1992)。タ
ウリンは、必須アミノ酸である含硫アミノ酸のメチオニンやシスチンから生合成され
ることが多くのほ乳類動物で明らかにされている。タウリンの生合成経路として、1)一
システイン→システインスルフィン酸→ヒポタウリン→タウリン,2)システイン→
システアミン→ヒポタウリン→タウリン,3)活性硫酸→システイン酸→タウリン
の3経路が推定されている。1)の経路でシステインからタウリンヘ代謝される過程の
中間代謝物であるシステインスルフィン酸は、システインスルフィン酸脱炭酸酵素
(CSD)によってタウリンの前駆物質であるヒポタウリンに脱炭酸される。仔ネコでは
このCSDの活性が弱いため、飼料からのタウリン供給が不足すると視覚異常が誘発さ
れること(Knopf et aL,1978)および生殖能力が著しく低下することや生まれた仔
ネコに異常がみられることなど、タウリンは必須の栄養素となっている。(Sturmanet
al.,1985;1986)。一方、魚類では、ニジマスを用いた飼育試験において飼料中にシ
スチンを添加すると魚体中のタウリン含量が増加すること(Walton et aL,1982)や、ら、魚類においてタウリンは必要ないとされてきた。このため、魚類におけるタウリ
ンに関する研究は、エキス成分としての研究が多く(Ozawa et a1.,1984;坂口ら,1988)、増養殖分野への応用をめざした研究はほとんど行われなかった。しかし最近の
研究により、タウリン合成に関与する酵素活性は魚種により大きく異なり、肝臓中の
CSD活性はニジマス、ティラピアなどの淡水魚では高く、マダイ、ブリ、ヒラメなど
の海水魚では低いことが明らかにされた(Yokoyamaet al.,20011Gotoeta1.,2001a, b,2003)。種苗生産を行うためには、ふ化率やふ化後の仔魚の生残率が高い良質の受精卵を安
定的に確保することともに、ふ化仔魚を効率的に飼育することが極めて重要となる。
天然海域での漁獲量の変動などに左右されずに良質の受精卵を安定的に確保するため
には、飼育環境下での採卵用の親魚の養成が必要となる。しかしながら、通常の飼育
環境下では成熟・産卵が進まない魚種が存在する。このような魚種では、水温や光周
期など環境の調節や、各種ホルモンの投与などの人為的な制御などの適切な親魚の養
成が必要になる。親魚の養成の中で卵に大きな影響を及ぼすものとして飼育環境の他
に、親魚の栄養状態がある(Izquierdoet aL,2001)。従来の親魚養成では、マアジ、サバ類などを主体とした冷凍生餌を給餌する場合が多いが、生餌は漁獲される時期や
場所により栄養組成が異なり不安定であることや、病原微生物侵入の懸念などの問題
がある。一方、配合飼料は栄養組成の調整が可能であり、さらに生餌に起因する病原
微生物の経口感染を未然に防除することが可能となる(虫明ら,2003)。海産魚類の親
魚の栄養要求については、マダイを用いて必須脂肪酸の欠乏により卵質が低下するこ
と(Watanabe et aL,1984a)、さらに飼料原料により卵質が影響を受けることを明ら和脂肪酸(n−3HUFA),アラキドン酸の要求量が明らかになっている(Furuita et al.,
200012003)。ブリでは配合飼料での親魚養成が可能であることが明らかにされ(虫明
ら,1995)、さらに配合飼料中に30㎎/kg前後のアスタキサンチンを添加することによ
り、採卵成績が向上することが明らかにされている(Verakmpiriya et aL,1997)。
このように魚種により異なるものの親魚の栄養要求についても、知見が蓄積されつつ
ある。しかしながら、親魚の栄養要求に関する研究には、親魚の飼育には大型の施設
が必要なことや、試験期問が長期にわたることなどから、試験を実施するに当たり困
難な条件が多くあることから、まだ十分とはいえない。
ふ化仔魚を効率的に飼育するためには、適切な餌料が必要になる。マダイ、ヒラメ、
ブリなどの海産魚類は、消化系諸器官がほとんど未分化な形でふ化する(渡辺,1985)。このため、これらの魚種では、ふ化後約10日ほどは、胃腺の働きなしで、餌・飼料を
消化吸収しなければならない。多くの人工飼料の主原料となる魚粉などのタンパク質
を十分に消化することが困難なため、実験規模では微粒子飼料の開発が行われている
ものの(Wang et a1,,2004)、完全に生物餌料と置き換わるまでには至っていない。現在でも、多くの魚種で大量培養が可能なワムシが初期仔魚の餌料として用いられて
いる。ワムシを用いた仔魚の飼育は、1960年に汽水産のシオミズツボワムシを普通海
水に馴致し、6乃1a郷伽o卿5幼.,6班αθ!勿の.などを餌料として飼育することによ
り、海水中で容易に増殖することを発見し、さらにこれを餌料として海産仔稚魚の飼
育を提唱したこと (伊藤1960)に始まる。1965年には日本栽培漁業協会の伯方島事
業場においてマダイふ化仔魚の飼育に使用された。海産クロレラを餌料としてワムシ
の培養を行ったが、ワムシの大量培養が必要になるとそれに見合う海産クロレラの供
年頃には、パン酵母のみで培養したワムシを仔魚に連続して給与すると摂餌開始HO
目後に仔魚の食欲減退、遊泳緩「曼、横転、腹部月畝茜などの症状が起こり、2−3日以内に全滅する現状が発生した。渡邉ら(1978)は、これらの原因は海産魚類の必須脂肪酸
であるn−3HUFAの欠乏によるものであることを明らかにした。さらに、海産クロレラ
によるワムシの二次培養(北島ら,1979),イカ肝油を資化させた酵母で生物餌料を
培養し強化する間接法(今田ら,1979),イカ肝油などのn−3㎜FAを多く含む油脂を
卵黄で乳化させ、直接ワムシやアルテミアの培養槽に入れ栄養強化する直接法
(Watanabe et al.,1982)を開発した。このようにEFA要求、特にEPAやDHAなどの
n−3㎜FAの要求量について検討されている(Takeuchi,1997)。しかしながら、ビタミ
ン類などについての検討があるものの、その他の栄養素についてはほとんど検討され
ていない。 一
わが国の養殖業は、戦後、順調に成長を続け日本の水産業における重要な地位を占
めるに至った。平成15年度の養殖生産量は125万トン、生産額は4,476億円に達し、
沿岸漁業に対する養殖の占める割合はそれぞれ約44%、47%となっている(水産年鑑
2006)。また養殖業はブリ、カンパチなどの高級魚を一般消費者に比較的身近な存在と
するなど、わが国における増養殖業は水産物の安定供給はもとより、高級品生産によ
る経済性追求あるいは伝統的食文化の保全に役立ってきた。しかしながら、近年生産
量の増加を目的とした過密養殖や過剰な餌の投与により、過度の有機物負荷が見られ
るなど、全国的に養殖魚場が悪化してきている。このような状況に適切に対応するた
めに、漁業協同組合等による養殖漁場の改善を促進するための措置及び特定の養殖水
産動植物の伝染性疾病のまん延の防止のための措置を講ずることにより、持続的な養
とするr持続的養殖生産確保法」案が提出され、平成11年5月14日可決・成立する
など、生産技術の開発と共に、環境の収容力などにも見合った適正な養殖事業とする
ため、生物特性と自然特性を考慮した環境保全型養殖業の展開が期待されている。主
な海面養殖は、現在では生餌ではなく主要なタンパク質源としてイワシ類を原料とし
た魚粉を大量に配合した飼料が利用されており、それらの魚粉の大半は南米等からの
輸入に依存している。しかし中国における養魚生産量の急増や魚、粉生産国における原
料となる魚の資源量変動等により、魚粉生産量は不安定かつ供給がひっ迫している状
況にある。このため、魚粉に依存しない配合飼料の開発が急務である。なかでも植物
性原料は、リンが少なく環境へのリン負荷軽減のためにも養魚飼料への積極的な利用
が望まれている。しかし、植物1生原料の配合に際しては、アミノ酸バランス、消化性
および生理阻害物質の存在などの問題があり(Liener,1989)、それらを改善するため
の処置を施さなければならない。これまでに主要な植物性代替タンパク質ヘメチオニ
ンやリジンなどのアミノ酸を添加することにより、その利用性が大幅に改善されるこ
とが明らかになっている(Takagi et aL,2001)。マダイやブリでは魚粉削減飼料を
給餌すると、成長の停滞や、肝臓が緑色に変色する「緑肝症」の魚が多発することが
報告された(Maita et al.,1997;Takagi et aL,1999;Watanabe et a1.,1998)。当初、本原因は胆管に粘液胞子虫が詰まり、肝臓に胆汁が欝積し緑肝を呈するといわ
れていたが(Maita et al,,1998)、そのような飼料にタウリンを添加することで症状が軽減することから、飼料中のタウリン濃度が緑肝症に影響を及ぼしていることが示
唆された(Goto et al,,2001c)。魚粉の代替原料として有望視されている大豆油粕などにはタウリンがほとんど含まれていないため、飼料中のタウリンが不足する可能性
以上のことから、本研究では、海産魚類のタウリン要求と役割を明らかにするため
に、第1章では、海産仔稚魚のふ化後の経過日数に伴うタウリン含量の変化を明らか
にするとともに、海産仔魚に対するタウリン強化ワムシの効果について検討した。人
工採卵・ふ化により得られたふ化仔魚から全長18cmまで飼育したブリ仔稚魚や、天然
で採取した稚魚とともに、人工種苗生産魚および天然魚が摂餌している餌・飼料のタ
ウリン含量を測定した(第1節)。次に、ワムシヘのタウリン強化方法を検討するとと
もに、異なる濃度でタウリン強化したワムシをふ化直後のマダラ仔魚に給与すること
により、タウリンがふ化仔魚に及ぼす影響について検討した(第2節)。
第2章では、ブリ稚魚および親魚に及ぼす配合飼料中のタウリンの影響について検
討した。第1節では、0,0.5,L O,1.5,2.0%のタウリンを市販飼料に添加しブリ稚 魚に6週問給与し、成長をマダイ稚魚の成長や全魚、体の遊離アミノ酸組成等を調べた。第2節では、魚粉削減飼料に0,0.5,L O%のタウリンを添加した飼料を産卵前に約昼
ヶ月間、ブリ親魚へ給与し、親魚の成熟度、採卵量、受精率などを調べた。
第3章では、マダイ稚魚に対する精製飼料中のタウリンの効果を検討した。‘般的
な養魚用配合飼料には、主なタンパク質源として魚粉が5割以上含まれるため、タウ
リンの要求性に関する研究には適さなかった。そこで、タンパク質源としてタウリン
を含まないカゼインを用いた飼料を新たに開発し、1.0%のタウリンを添加した飼料を
マダイ稚魚へ給与した。さらに、飼料中のタウリンおよび含硫アミノ酸の1つである
シスチンがマダイ稚魚の摂餌行動およびアミノ酸代謝に及ぼす影響について検討する
ため、タウリンを0,0.5,1.0,2,0%,シスチンを1.0,2。0%添加した精製飼料を6週間給与した。マダイ稚魚の成長や全魚体の遊離アミノ酸組成等を調べるとともに、
るタウリン要求量および脂質の消化吸収におけるタウリンの役割を明らかにするため
に、タウリンを0,0.1,0.3,0.5,0.7%,タウロコール酸を0.5%添加した精製飼料を
本論文の一部は下記に報告済みである。
(1)学術雑誌等に発表した論文 1.松成宏之,竹内俊郎,村田裕子,高橋 誠,石橋矩久,中田 久,荒川敏久人工種苗生 産ブリ仔稚魚におけるタウリン含量の変化および天然稚魚1との比較 目本水産学会誌,69(5), 757−762, 2003. 2.Hiroyuki Matsunari,Daisuke Arai,Masahiko Koiso,Hiroshi Kuwada,Takayuki Takahashi, Toshio Takeuchi.Effect of fee(1ing rotifers enriche(i with taurine Gn grQwth performance and body composition of pac i f i c cod 6旨漉zs zzz80■roo鋤∂」∼どs larvae. ノ1σ4∂o〃!オ㎝θ5ヒ1θ刀oG 53, 297−304, 2005, 3. Hiroyuki Matsunari,Toshio Takeuchi,Makoto Takahashi, Kei ichi Mushiake. Effect of dietary taurine supPlementation on growth performance of ye1].owtai1 51θzブ01∂ 9乙!f/7σ乙!θ畑07∂6∂ juveni les. 乃’s々θヱゴθ55bゴθ刀065 71, 1129−1133, 2005. 4. Hiro ki Matsunari,Kazuhisa Hamada,Keiichi Mushiake, Toshio Takeuchi. Effects of taurine level s in broodstock〔ii et on the reproductive performance of yel lowtai l跡101∂ 9び1η卯θ塑認κa乃『shθガθ55ヒf㎝o畠72,955−960,2006. (2)学術雑誌等又は商業誌における解説、総説 1.竹内俊郎,松成宏之.タウリンの有効性養殖43(10),72−75,2006. (3)国内学会・シンポジウム等における発表 1.o竹内俊郎・松成宏之・石崎靖朗・金 信権・秋元淳志・高橋 誠・石橋矩久 ブリおよび マダイ稚魚に対するタウリンの添加効果平成13年度目本水産学会大会,p.102. 2.○秋元淳志・松成宏之・石崎靖朗・竹内俊郎・高橋 誠・宿輪 仁・石橋矩久 ブリ当歳魚 に対するタウリンの添加効果平成13年度目本水産学会大会,p。102. 3.○松成宏之・竹内俊郎・村田裕子・高橋 誠・浜田和久・虫明敬一 ブリ仔稚魚および幼魚 におけるタウリン含量の変動平成!4年度目本水産学会大会,p。125. 4.○浜田和久・松成宏之・虫明敬一・宿輪仁・竹内俊郎ブリ親魚の産卵に及ぽす親魚用配 合飼料へのタウリン添加効果一L採卵結果平成15年度目本水産学会大会,p.152. 5.○松成宏之・竹内俊郎・村田裕子・秋元淳志・浜田和久・虫明敬一 ブリ親魚の産卵に及ぼ す親魚用配合飼料へのタウリン添加効果一2.分析結果.平成15年度日本水産学会大会, p.152. 6.○松成宏之・竹内俊郎・古板博文・山本剛史・金信権・横山雅仁低タウリン飼料がマダ参考文献
Furuita,H.,Tanaka,H.,Yiamamoto,T,S㎞f飢shi,M.,Takeuchi,T,2000.Ef臣cts ofnβHUFA levels血broo戯ock(iiet on廿1e reproductive perR)nnance an(i egg an(i larval quaH取of血e Japanese且omde蔦Pαzα1∫6h吻solivα6ε欝。吻襯cぬ肥187,387−398. Fun口t亀H.,Yi mamoto,T,Sh㎞嬬N。,Suzuki,N.,丁盛euchi,T,,2003.Ef琵cts of arachidonic acid Ievels hl broodstock diet on larval an(l egg quality of廿1e Japanese floundeちPα7α1’6h吻s ol’vα‘θ鰐.匂μαo〃1孟耀220,725−735,古田普平 1998。ヒラメ人工種苗と天然稚魚の摂餌行動の比較 日水誌 64,393−
397. Goto,T,Ma鵬moto,T,Murakami,S,,肱kagi,S.,Hasumi,E,2003.Conve総ion ofcysteate into tau血e hl Hver offish.ノ㌃曽h、So孟69,216−218。 Goto,T,Maおumoto,T,Takagi,S.,2001b.Distribu廿onofthehepaticcysteaminedioxygenase ac丘vi丘es hl fish.乃曽h。5ヒ云67,1187−1189. Goto,T,mkagi,S。,Ich盟,T,Sakai,Tl,Endo,M.,Ybshida,T,,Ukawa,M.,MurataH.,2001c. Studies on the green Hver㎞cultured r鉗sea bream f6d low level and non岨sh meal diets: Relationship between hepa亘c協u血e and b丑iverdhl levels。E∫sh Sb孟67,58−63. Goto,T,Tiba,K,Sa㎞rad亀Y,%kagi,S.,2001a。Deもem血laHon ofhepa廿c cysteinesulf㎞ate d㏄arboxylase activity in fish by means of OPArprelabe㎞g and reverseヤhase hi⑳一perR)㎜ance liqui(l chromatographic separaUon.FZsh。Sヒ云67,553−555.今田克,影山百合明,渡辺武,北島力,藤田矢郎,米康夫,1979。魚介類種苗生
産用酵母(油脂酵母)の開発。日水誌 45,955−959。伊藤隆1960.輪虫の海水培養と保存について.三重大水研報3,708−740.
Lquierdo,M.S。,Femandez−Palacios,H,Tacon AGJ。,E焼ct of broodstock nutddon on repro(1uc廿ve perR)nnance offish。∠〈1(1zイα6π1魏形200119,25−42。北島力,藤田矢郎,大和史人,米康夫,渡辺武,1979.クロレラで2次培養したパン
酵母ワムシの餌料効果 日水誌 45,469−471.㎞。P凪S伽1副へ㎞s柱。n&瓢H&ye亀Kg1978。願血e:舳esse翻n面e曲r
dle cat.Jlノ〉初〃:108,773−778. LienerI。E.,1989.Antinut且tional factors inlegume5eeds:stateofthea並.Ih:J。Huisman,TEB. van der Poel and I.E Liener(E(iitors),RecentAdvances ofResearch inAnt加uthtional Factors血k騨eSe。ds・恥d・c陶e血騨←14・
Mai砥M。,Aoki,H,Yamagata,Y,SatohラS.,0㎞oto,N,Wa旋mabe,Tl,1998.Plasma. biochemisUyand.dieaseresistanceinyenow副琵danon。fishmealdiet乃shPα孟ho乙33,59−63.虫明敬一,本藤 靖,崎山一孝,浜田和久,堀田卓朗,吉田一範2003.日本栽培漁業
協会における親魚養成技術開発の現状と今後の課題(総説).栽培技研 30,79−100. 虫明敬一,河野一利,Visu面驚ralunp㎡y灸渡邉武,1995.市販ソフトドライペレットをOzawa,A,Aoki,S.,Suzuki,K,Sugimoto,M,F唾鳳丁,Ts両i,K,1984.丁泡u血econtent圭nfish εmd shens.Nippon SuisanGakkaishi。37,561−567。
朴 光植,竹内俊郎,青海忠久,中添純一,1997.ヒラメ稚魚の生物餌料としてのア
ミの栄養価に関する研究 水産増殖 45,371−378.
朴光植竹内俊郎青海畝横山雅仁2・・1.飼料中のタウリン漉ラメ稚魚の成
長および魚体内のタウリン濃度に及ぼす影響.日水誌67,238−243.
Park,Gs。,Takeuchi,T,Ybkoyama,M.,seikai,T、,2002。QPt㎞al die町tau血e level fbr gro嚇hofjuvenUeJapanese且otmderP醒α1’oh吻sollvαoε濾乃sんSo云68,824−829。坂口守彦,村田道代,1998 タウリン,魚介類のエキス成分 恒星社厚生閣,東京,
56−65. St㎜an,JA,Moret乙,RM.,French,JH.,Wisniewski,HM.,1985.1加血e d韻ciency hl the develophlg cat:persistence ofthe cerebellar extemal granule cemayer Jコ糖矧osα1∼θ&13, 405−416. S厩m砥JA.,G肛gano,AD.,Messing,JM.,㎞面,H,1986.Fe㎞em鴫m認tal血ede昼ciency: ef驚ctonmotherandof魯p血g。」八勉疏116,655−667. Subiyanto,L,H批繊L,Sen砥丁,1993.L餌al se賃lementoftheJapanesenounderomsandy beaches of廿1e YatsushhD sea,Japan。ハ似2pon Sμなαn Gα蜘なh’59,1121−1128。水産年鑑編集委員会(2006): 水産年鑑 水産祉 東京
水産庁・目本栽培漁業協会(2002)平成12年度栽培漁業種苗生産,入手・放流実績(全
国).105 mkagi,S.,Hoso㎞w嬬H.,Sh㎞eno,S。,M田ta,M.,Ukawa,M.,Ueno,S.,1999.U出iza廿on of soy prote血concen柱ate血a diet貴》r red sea bream,P㎎ア鰯〃吻りκ翻Zsαnzoz器hoんz∫47,77−87, 艶kagi,S.,Sh㎞eno,S.,Hosokawa,H。,Ukaw灸M.,2001.Ef陀ct of Iysine and me血onine supplementadonto asoyprotein concentratediet fbrred seabream−Pα解s〃吻加瓦5h翫孟67, 1088−1096. %keuchi,T,1997.Essential f激y acid requirements ofaqua丘c animals with emphasis on fish Iarvaeand行ngerlings。RθvF肋So云5,1−25。 艶keuchi,T,Park,GS。,Seikai,Tl,Ybkoyama,M.,200L肱u血e contentinJapanese且ounder Pα7α1’oh孟伽ol’vαoθz器丁&S.and red sea bream P卿鰐n吻り7T,&S.du血9血e peho(10f see(i production.!望(1襯oz41孟ノ∼θ£32,244−248. Takeuchi,T,,Sh丘眠 Y,Vね㎞be,T,199L Suitable protehl an(i Hpid levels hl diet R)r 行uger㎞gs ofred seabreamPα野麗s溺φoπム勉ρonS麗sαn Gα纐’sh正57,293−299。塚本勝巳 1990.種苗性のはなし さいばい 55,20−24.
V¢ra㎞np墳y灸V,Mushiake,K.,Kawano,K.,Watanabe,T,,1997。Supplemental ef驚ct of astaxan仕血1血broo(聴tock diets on dle quali取ofyellov副eggs.況曽h,Soよ63,816−823. W飢ton,MJ.,CoweヱCB.,Adron,JW.,1982.Methioninemetabolisminrξ血bow廿out聞diets ofdif艶血gme踊o血eandcystineootent.」轍疏112,1525−1535.wiang,Q。りmkeuchi,T,Hirota,T,Ishid灸s。,Miyakawa,H,,Hayasawa,H,2004。apPlica丘onof ㎡croparticle diets R)r Japanese且ounder P醒α110hりzs ollvααヲzイs larvae。乃sh,S砿70,611−619. Watanabe,T,Aoki,H.,Sh㎞amoto,K,Hadzuma M.,Mai砥M.,Yiamagata,Y,K辻on,V, Satoh,S。,1998。Athaltocultureyellovvtailwithnon一五shmea1(1iets.瓦sh.S6孟64,505−512。
渡辺武,荒川敏久,北島力,福所那彦,藤田矢郎1978。脂肪酸組成からみた仔稚魚
用生物餌料の栄養伍 日水誌44,1223−1227. Wa荘1nabe,T,Itoh,A,Murakami,A.,聴ukash㎞a,Y,Ki吻㎞a,S.,F両ita,S。,1984b.Ef驚ctof nuhi哲onal quaHty ofcliets given to broodstock on廿1e verge of spav血g on repro(1uc廿on ofre(1 sea brean1.1984b。ノ〉な2pon Sz〃sαn Gα蜘∫5h云50,1023−1028. Watξmabe,T,Ohta,C.,Kit勾㎞a,C.,F噸ita,S.,1982.㎞provement ofdietaly value ofb血e sh血lp Alte㎡a sa㎞a fbr Iarvae by fヒedingthem onω3highly unsa缶rated伽y acids。ハ吻po〃 Sz泥sαnσα蜘Zsh孟48,1775−1782. Watanabe,T,Ohhash,S。,Ito瓦A。,Kit勾㎞a,C.,F両ita,S.,1984a Effヒct of nutritional compositon ofdiets on che㎡cal components ofre(1seabream broodstock and eggs produ㏄d。 吻pon5協1sαnGα㎞護sh云50,503−515. Watanabe,T,驚ra㎞nphiya,V,Watanebe,K,Viswana甑K.,Satoh,S.,1998。Feedhlg of 面hbowroutwithnon一五shmealdiets.π曽h。5厩69,242−248。渡辺良朗 1985。仔魚の消化吸収機構,水産学シリーズ54 養魚飼料 恒星社厚生閣,
東京,89−98.Ybkoyama,M,Nakazoe,」。,1992.Accumulation and excretion of tau血e in面hbow廿out
(Onoo吻noh欝n呈吻ss),偽d(1iets supplemented with methionine,cystille,and ta面ne.Co〃ψ.
Blo6hθ肱P勿sioよ102A,565−568。
Ybkoyama M,TakeuchiエPark GS,Nakazoe J・,200L Hepa怠c cysteinesulp㎞ate
第1章
海産仔稚魚のタウリン含量とタウリン強化ワムシによる給与効果
第1節
人工種苗生産ブリ仔稚魚におけるタウリン含量の変化および天然稚
人工種苗生産ブリ仔稚魚におけるタウリン含量の変化および天然稚魚との比較
短縮表題 ブリ仔稚魚のタウリン含量の変化
松成宏之,1竹内俊郎,1*村田裕子,2高橋 誠3石橋矩久,3・中田 久,4荒川敏久
4
1東京水産大学資源育成学科,2水産総合研究センター中央水産研究所,3日本栽培漁業協
会五島事業場,4長崎県総合水産試験場 Changes in the taurine content (iuring the early growth stages of artificial ly produce(i yellowtail and a comparison with their wild fish Hiroyuki MATSUNARI,1 Toshio TAKEUCHI, i※ Yuko MURATA,2 Makoto TAKAHASHI,3 Norihi sa ISHIBASHI,3a Hisashi CHUDA,4AND Toshihisa ARAKAWA4 1Department of Aquatic Biosciences,Tokyo Univers ity of Fi sheries,Minato,Tokyo, 108−84,77, 2National Research Institute of Fisheries Science, Fisheries Re$earch Agency,Yokohama,Kanagawa,236−8648,3Goto Station,Japan Sea−Farmirlg Associat ion, Tamanoura,Nagasaki853−0501,4Nagasaki Prefectural Institute of Fisheries,Taira, Nagasaki, 851−2213 *Tel Fax :81−3−5463−0545. Emai l :take@toky−o−u−fish.ac.jpa現所属:日本栽培漁業協会南伊豆事業場(Minamiizu Stati・n,Japan Sea−Faming
Association, Irozaki, Shizuoka, 415−0156,Japan)人工種苗生産ブリ仔稚魚におけるタウリン含量の変化および天然稚魚との比較
松成宏之,竹内俊郎(東水大),村田裕子(中央水研),
高橋 誠,石橋矩久(日栽協),
中田 久,荒川敏久(長崎水試)
異なる餌料系列を用いた種苗生産過程におけるブリ仔稚魚および飼育に用いたワム
シ、アルテミア、天然コペポーダ、冷凍天然コペポーダ、配合飼料のタウリン含量お
よび天然稚魚におけるタウリン含量との違いを調べた。その結果、人工種苗生産過程
におけるブリ仔稚魚のタウリン含量は、餌・飼料中のタウリン含量の影響を受けるこ
と、特に開口時までに多くの遊離アミノ酸が減少するのに対して、タウリンは開口後
のワムシ給餌期に大きく減少すること、また人工種苗生産稚魚は天然稚魚に比べて、
タウリン含量が著しく少ないことが明らかとなった。
キーワード
ブリ,仔稚魚人工種苗,天然稚魚ジタウリン,遊離アミノ酸
Changes in the taurine content (iuring the early stages of artificial ly reare(i yel lowtai l and a comparison with their wi ld fish Hiroyuki MATSUNARI, Toshio TAKEUCHI, Yuko MURATA, Makoto TAKAHASHI, Norihi sa ISHIBASI, Hi sashi CHUDA, and Toshihi sa ARAKAWA This experiment was con(iucte(i to investigate the changes in taurine content during the development of artificial ly produce(i yellowtai15θrゴ01∂(1乙1fη(1飢∂oゲ∂孟∂ larvae an(玉juveniles.Besi(les this, taurine content of wild caught and reare(i yel lowtai l was compared. Free amino aci(is content in larvae an(i juveni les pro(iuced at the Goto Station of Japan Sea−Farming Association an(i Nagasaki Prefecture Institute of Fisheries (TL, 4.2−186.8mm) and juveniles caught from the coastal area of Nagasaki Prefecture (TL, 29.7−179.O mm) were analyzed. The content of Most of free amino acids such as lysine, leucine, i soleucine, 、alanine decreased between the ferti l ized egg stage and open−mouth stage, with the exception of taurine in artificially reared fi sh.The taurine content in the whole body of the artificially reare(i larvae decrease(i rapi(ily during the rotifer feeding.The amount of taurine in the wi ld fish was higぬer compare(i to the culture(l fi sh,丁五ese results reveal that taurine has arl important role compared to other free amino aci(is, suggest ing l ive foo(i and(1iet use(i for yel lowtai l cu!ture would not satisfy taurine requirement of yel lowtai l.
ブリ 施ガ01∂σθ1超θθr雄∂6∂(Te㎜inck et Schlegel)は、日本の魚類養殖におい
て最も生産量が多く重要な魚種の1つである。ブリの養殖は体長数cmから10数cm
のモジャコと呼ばれる天然の稚魚を採捕したものを用いて行うため種苗の供給は、モ
ジャコ漁に左右され、不安定である。計画的かつ安定的な種苗供給の実現のためには、人工種苗生産技術の確立が必要不可欠であると考えられる。しかしながら、現在の人
工種苗生産では生残率が低く、また形態異常の発生など改善しなければならない点も
多い。ブリの栄養要求に関する研究は古くから行われており、稚魚の脂質およびタンパク
質の適正量、n−3高度不飽和脂肪酸要求、1
2)仔稚魚のドコサヘキサエン酸(DHA)要 求、3)稚魚および幼魚のアミノ酸要求4−6)などが明らかにされている。含硫アミノ酸の1つであるタウリンは水産動物の組織に広く分布し、しかも多量に
含まれている。その生理作用として、海産生物では浸透圧調節物質として知られてい
.』る程度で知見は乏しかったが、近年タウリンの魚類に及ぼす影響について研究が進められている。ヒラメ盈畑!10h訪” o万旧oθ認稚魚において飼料中のタウリンが稚
魚の健全な育成に極めて有効であることが明らかになるとともに、7〉ヒラメ仔稚魚で
は体内でのタウリン合成に関与するシステイン硫酸脱炭酸酵素(CSD)の活性が微弱で
あること、またブリではこの酵素活性がヒラメよりもさらに低いことが明らかにされ
ている。8)また、マダイおよびヒラメではふ化直後に魚体中のタウリン含量が減少す
ることも明らかにされた。9) ブリ稚魚については、これまで、一般組成、無機質含
量10)、脂質組成および脂肪酸組成11)について飼育魚と天然魚の比較を行ってきたが、タウリンのブリ仔稚魚に対する効果についての研究はこれまで行われていない。そこ
的に、人工種苗生産過程におけるブリ仔稚魚のタウリン含量の変動を調べるとともに、
天然稚魚との比較を行った。材料および方法
人工種苗は、日本栽培漁業協会五島事業場において生産された種苗(五島産と略記)、および長崎県総合水産試験場において生産された種苗(長崎産と略記)を用いた。五
島産は、60m3陸上コンクリート水槽を用い、飼育水温は22℃で飼育した。仔稚魚の給
餌に用いた飼料系列をFig,1に示す。給餌したワムシおよびアルテミアの2次強化に
は、アクアラン(BASFジャパン㈱製)を生物餌料水槽1kL当りそれぞれ100および
200g使用した。また、ワムシ給餌期間中は、飼育水にナンノクロロプシスを50万セ
ル/mLとなるように添加した。
長崎産は、100m3陸上コンクリート水槽を用い、飼育水温は22℃で飼育した。給餌
したワムシは、午前の給餌にはプラスアクアラン(BASFジャパン㈱製)を生物餌料水
槽1kL当り60g、午後の給餌にはマリングロス(日清製油㈱製)を0.5Lとすじこ乳
化油(日清サイエンス㈱製)15gを併用して添加し、水温23−24℃で16−20時問の強
化を行った。アルテミアは午前の給餌ではDmセルコ(長瀬産業㈱/1㎜)を生物餌料
水槽1kL当り6009、午後の給餌にはマリングロスを1Lとすじこ乳化油309を併用
して添加し、水温23−24℃で16−20時間強化した後給餌した。飼育水には、ナンノク
ロロプシスを50万セル/mLとなるように添加した。仔稚魚および生物餌料のサンプ
ルは、水道水でよく水洗いした後水分をよく拭き取り、湿重量でそれぞれ約2gを採取
天然穐魚は、5月24日に長崎県五島列島沖で採集したもの(五島沖採捕)、および
5月16日から17目の間に長崎県総合水産試験場・調査船わかづる第2回航海で採集
(長崎調査船採捕)し、それぞれサイズ別に分別したものを用いた。仔稚魚は一20℃で、ワムシおよびアルテミアは一80℃で凍結保存したものを後目分
析に供した。分析方法
供試魚は包丁で細かくした後、遠心粉砕機(長谷川式スーパーファイブレーター ㈱
長谷川鉄工所製)を用いてすり潰した。ワムシおよびアルテミアは解凍後、遠心分離
機H−200型国産遠’亡機社製)を用いて10,000rpm(5590×g)で10分間遠心分離し
た。これらを、真空凍結乾燥機(共和真空技術RLE−206型)でそれぞれ乾燥させ、乳鉢
を用いて細かくすり潰し分析に供した。市販飼料は乳鉢ですり潰し均一にした後分析
に供した。試料は遠沈管に入れ、2%スルホサリチル酸を加えホモジナイザーを用いて
均一化した後、3000rpm(1509×g)で15分間遠心分離した。さらに2%スルホサリチ
ル酸を加え、上澄み液を50mLに定容したものをアミノ酸自動分析機(Model8500−A:
日立製作所製)を用いて分析した。なお分析はプールサンプルとした。
結果
餌・飼料の分析結果
離アミノ酸の総量は100g当り、ワムシ・アルテミア・天然コペポーダ・冷凍コペポ
ーダ・配合飼料では、それぞれ2855mg,3753㎎,7733mg,12119mgおよび2288mgであっ
た。その中でタウリン含量は100g当りそれぞれ84.7㎎、668mg、1182㎎、461㎎およ
び439mgであり、ワムシ中のタウリン含量が最も低い値を示していた。
人工種苗生産魚の分析結果
五島産と長崎産の人工種苗生産魚の遊離アミノ酸含量をそれぞれ、Tables2および
3に示す。
魚体中の遊離アミノ酸含量は、五島産では受精卵で12968mg/100gあったが、日齢3
の開口では2908mg/100gであった。その後は、約3000mg/100gとほぼ一定であった。
また長崎産でも同様に、受精卵で12270mg/100gあったものが、日齢3では3578㎎/100g
まで大きく減少し、両機関でほぼ同様の傾向がみられた。その中で、アルギニン,リジ
ン,ロイシン,イソロイシンなどが目齢1までに特に大きく減少していた。一方、メチ
オニン,トリプトファンは、ふ化に伴う変化は少なかった。この様な遊離アミノ酸の変
化の中で、タウリンのみがやや増加する傾向を示した、すなわち五島産では、受精卵
の410mg/100gから日齢3の626mg/100gへ、長崎産でも受精卵の680mg/100gから目
齢3の799mg/100gと増加した。
開口後になると、タウリンの含量が大きく変動した。すなわち、五島産、長崎産と
もに、目齢15∼19(全長:TL,7∼11㎜)までに大幅に減少した。その後目齢23∼42(TL9 ∼28㎜)で増加するが、TL40㎜を過ぎると減少し、一定となった(Fig。2)。天然稚魚の遊離アミノ酸分析結果をTable4に示す。
天然稚魚は、人工種苗生産魚と比較すると魚体中の遊離アミノ酸含量が多い傾向がみ
られた。五島沖採捕のものはサイズに関わらず約3700㎎/100gとほぼ一定であった。
一方、長崎調査船採捕のものではTL30㎜で約6000㎎/100gあったものが、TL120㎜で
は五島沖採捕のものとほぼ同様の約4000mg/100gまで減少した。
次に、魚体中のタウリン含量をみると、平均TL30∼42㎜で、2200∼2400㎎/100g含
まれていたものが成長に伴い減少する傾向を示した(Fig.2)。先の人工種苗と比較し、天然稚魚はタウリンを同じサイズで2∼4倍多く含んでいた。
考察
今回の分析結果から人工種苗生産過程におけるブリの遊離アミノ酸含量は、目齢3
の開口までに大幅に減少することが明らかになった。しかしその変動は、アミノ酸の
種類により異なった。すなわち受精卵中700㎎/100g以上と多く含まれたアルギニン,
リジン,ロイシン,イソロイシンなどは日齢1までに大きく減少した。またフェニルア
ラニン,チロシンは日齢3の開口までに緩やかに減少した。海産仔魚では遊離アミノ酸
が艀化前にはタンパク質合成に、ふ化後はエネルギー源に使用されることが明らかに
されている。12)これらのアミノ酸は、ブリでも同様に卵発生および艀化直後に重要な
働きをするのではないかと考えられる。このように各種のアミノ酸が開口までに減少
する中、タウリンのみが若干増加した。
を示したが、その後五島、長崎産ともに、約3000mg/100g前後で推移し大きな変動は
みられなかった。一方、タウリン含量は大きく変動した。五島産では日齢19で
39.4㎎/100g、長崎産では日齢15で203㎎/100gとなり、ワムシ給餌期間中は魚体中
のタウリン含量が大幅に減少した。その後アルテミアや、天然コペポーダを給餌する
ことにより、五島産では日齢32で738㎎/100mg、長崎産では目齢23で1253㎎/100g
と高くなっている。さらに生物餌料から配合飼料へと給餌した餌・飼料の変化に伴い、五島、長崎産ともに600mg/100g前後となった。ブリでは人工種苗生産過程における魚
体中のタウリン含量は、給餌した餌・飼料のタウリン含量に極めて近い値を示すこと
が明らかになった。つまりブリの仔稚魚は、餌・飼料の影響を大きく受けることから、
体内でのタウリンの合成能が低い可能性が考えられる。またブリの魚体中のタウリン
含量は、ふ化後20日までに大幅に減少することが明らかになった。海産仔稚魚では必
須の栄養素であるDHAが、ふ化後10目前後までに魚体内で著しく減少する13) こと
が明らかになっているが、タウリンでも同様の傾向が確認された。現在、ブリの人工
種苗生産過程では開口直後はワムシの単独給餌を行っている。しかし、本研究により
ワムシはその他の餌・飼料と比較するとタウリン含量が極めて少ないことが明らかに
なった。人工種苗生産過程では、ワムシ単用給餌時期に魚体中のタウリン含量が大き
く減少していることから、この時期は体内での消費量が供給量を上回り、給餌したワ
ムシのタウリン含量がブリ仔魚のタウリン要求量を満たしていない可能性が示唆され
る。ブリでは、必須の栄養素であるDHAをワムシに強化することにより高い生残率お
よび活力を示すことが明らかにされている。14)このようなことから、ワムシヘのタウ
リン強化について検討する必要があると考えられた。
リの食性については、TL約30mmから魚類の捕食を開始するが、約60mmまでは小
型椀脚類や枝角類を、60∼70mmを超えると大型のプランクトンを主食とする。以後
は魚類への依存度が高まり、TL約130mm以上になると完全に魚食陸に転換する。15)
このように、TL30mmから130mm前後にかけて食性が大きく変化することが知られ
ている。人工種苗生産過程では、給餌した餌・飼料のタウリン含量に極めて近い値を
示す傾向が確認されたことから、天然稚魚の魚体中のタウリン含量が成長するに伴い
減少しているのは、捕食生物の変化によることが一因であると考えられる。一方、ブ
リの人工種苗生産過程ではTL約30mm以降は配合飼料のみの給餌を行っている。こ
のため人工種苗のタウリン含量が成長に伴う変化はなく約700mg/100g前後で一定
であったのは、飼料中のタウリン含量に大きな変化がなかったためと推察される。天
然稚魚と人工種苗生産魚を比甦すると・TL30㎜前後の時期には2倍以上もの大きな差
が認められた。魚体中のタウリン含量の差がどのような影響を及ぼすのかについては
明らかではないが、ヒラメでは、タウリン含量の異なる配合飼料を給餌することによ
り、成長や飼料効率の改善、さらには摂餌行動が機敏になることが報告されている。
lG17)天然稚魚と人工種苗生産魚のタウリン含量の差が大きい仔稚魚期におけるブリの
タウリン要求量やタウリンがブリヘ及ぼす影響については今後詳細に検討する必要が
あろう。文 献
1) Takeuchi T, Shi ina Y, Watanabe T, Sekiya S, Imaizumi K. Suitable protein and l ipid level s in(iiet for fingerlings of yellowtai/.ハゲ夘o刀&!1s∂刀 佑ノ惚15方1 1992;58:1333−1339.2) TakeuchiT,ShiinaY,WatanabeT,Sekiy『aS,ImaizumiK.Suitablelevels
of n−3 highly『unsaturate(i fatty aci(1s in diet for fingerl ings of yel lowtai l.躍夘o刀5αfs8刀62漁1訪11992;58:1341−1346.
3) 竹内俊郎,石崎靖朗,渡邉 武,今泉圭之輔清水 健D皿含量が異なるワムシ
を摂餌したブリ仔稚魚のアルテミア摂餌期におけるDHA要求.日水誌1998;64:270
−275. 4) Ruchimat T, Masumoto T, Hosokawa H, an(i Shimeno S. Quantitative methionine requirement of yel lowtai l (5θrfo1∂ (1乙!117(7〃θ塑oγ∂言∂). 、4(1α∂oz!!オ砿θ 19971 15Q: 113−122. 5) Ruchimat T,Masumoto T,Hosokawa H,Itoh Y,an(i Shimeno S. Quantitat ive lysine re(1uirement of yellowtai1 (5θ■fo1∂9乙!fη〔7乙!θz30γ∂6∂). 。4(7μ3α!1孟砿θ 1997; 158: 331−339. 6) Ruchimat T, Masumoto T, Itoh Y, and Shimeno S. Quantitative arginine requirement of juven i l e 飴rlo1∂ (7乙!fησz!θT30ゲ∂6a ノヴ5カ.5りf,1996; 64: 348−349.7) 朴 光植,竹内俊郎,青海忠久,横山雅仁飼料中のタウリンがヒラメ穐魚の成
長および魚体内のタウリン濃度に及ぼす影響 日水誌2001;67:238−243。8) YokoyamaM,TakeuchiT,ParkGS,NakazoeJ.Hepaticcysteinesulphinate
(lecarboxylase activity in fis姓. 、49α∂α!1ゑ 危sレ 2001; 32: 216−220. 9) Takeuchi T,Park GS,Seikai T, Yokoyama M.Taurine content in Japanese flounder搦r∂110力孟カア5『011レ∂oθαs T.&S.an(i re(i sea bream舶騨びs〃z∂』メor T.&S.(iuring the period of see〔i pro(iuction, ヨ(7μ∂o‘11乙 ノRθ5 2001; 32: 244−248.10) 荒川敏久,石崎靖朗,中田 久,清水 健,有元 操,竹内俊郎.飼育およ
び天然ブリ稚魚の一般成分および無機質含量の比較。日水誌2001;67:794−800.
11) 荒川敏久,石崎靖朗,中田 久,清水 健,有元 撮竹内俊郎.飼育およ
び天然ブリ稚魚の脂質組成および脂肪酸組成の比較 目水誌2002168:374−381.
12) Ronnestad I. Fyhn H. J。 Metabol ic aspects of free amino aci(is indevelopingmarinefisheggsandlarvae.忍θv1θ雁加冊畝5砿19931(3),239−259.
13) 竹内俊郎:魚類における 必須脂肪酸要求の多様性.化学と生物
1991;29:571−580. 14) Furuita H,Takeuchi T,Watanabe T,Fujimoto H, Sekiya S, Imizu血i K, Requirements of larval yel lowtai l for eicosapentaenoic aci(i, (iocosahexaenoic ac i(i,and n−3 hi ghly unsaturated fatty ac i(エ ノゼ5カ.5ヒし乙1996;62:372−379.15) 安楽正照,畔田正格.流れ藻に付随するブリ稚仔魚の食性.西水研報
1965;23:15−45.16) 朴 光植ヒラメ稚魚の健全性に対するタウリンの添加効果に関する研究博
士論文,東京水産大学,東京,1999.1 7) Park GS,Takeuchi T,Yokoyama M,
for growth of juveni le Japanese
ノウD5五.5ヒンーz’.2002;68:824−829.
Se ikai T. Opt imal di etary taurine level
' 1;
>
, O bO =1
O O Lh LO l 1: O Q: ID O O , l O Q' ,5 , LO ' ' ' I E 1: b5O
O
-¥
bO E ' = O ' :: O O1
O C:, O C: E 'S O e LL-
o JD c:'H
' o 1: CQ o Lo E E oO
C OC:
ONC: O'T: LLLO
O c: o 'Dc
:=,T5-
o o * l q) : L(D Lh.-4J o ac ._ , s c * ** , s : :: ) e E'NLocoocoo(ol¥ ico r
,occ;d(1;r ai cir(oicc;cocooLocoa)Loc¥1(0 '-
c¥lcOo,NOcocC,,Oc0 r[s)r¥
cOr;LC;c¥lC:;r( c icioc;o
co aDLr)-cOOco-Lr)aO
_cocNcO r(o rc¥lLo
r-Locf)o) hco o'N
rcoodcc; cl; c¥iadd
rc¥1 co
cd F
oo r rc J
c l_ c¥1 co_ _a)r __ r
,O-
riOc¥1,Nc¥lL:) -O
c¥1 cl;
c0 FcT;dr; ciL,:;O
co,oc¥ -OhLO_cocO
c¥1--h
c9Lo Lf)r¥cooci)
:c¥i r ci Foc;Lri
co Loo, ) cl; O,C,
hco _cocohLoh
_co_
-Loco
o = .- = O co oca o o oc:c= :: Q
=o
- - : 5 a) =- .::- .-. o
=0
E5:1 . >u)o0 '
t
._
ooa': ;!oQ
o',U
<J!:cLHJ 2 >HH
._ J S O ・ S * , S_ i : ) e e )i : O :or¥h(oo)co
c:;r r
0id
o
coc' r-coc¥lcl'cl'_,Nco0 rcoo'c:)
'r¥coc¥ic ': o;c¥lco
2::oeo rco(o
cv)'0 rLoC¥lOCO cO
CY).O(N 'd ・1-'
COCi)LO LO
'N{ CO LO;: r::CN-LC'
coh Lo")
co'o
a:; F c¥iad
Lc;d
coco ocl)
coco Loco
c:)cl,-h ¥cool¥col¥
cl;a:;c¥id_r
Lf)Oco-Lr)co
co__c¥t-:._ Ip
o '
o ,o (:f o o o ,D 0' ,DIE Q) !: s: c: - .- -- = o ' ' : c o o :: ,!' ':: c!: CUj a) co LH :OOCO<0_
CO CO CO ,N C a (:i C¥l co ci coh
h
O
CO LOh
CO CO F LO CO C¥l 'D , oH
8
C:l O 151
O L ,5 O L c5 o : LE ,5 ,: 1: o o o !: ; o ,o co cg J: L 'D t ,5 E 1:O
O
-¥
E C C O O l9 o O C E ,D a) L LL ,:f C¥l U:) cT: ) to a) ,Q coh
: o) r) (T, 17 CO Lr)h
t : C l CO c o) : QO cQ taO bO O ,D t: ,D LL = : o ' cT' Loo
r -+1 co cc; co o) I!i +1 Lo o) Go cc; +1 cl' h; (o co Ls) +1 Lo.o
r) co. co +1 c . c l r r ci +1o
cl; ao c i +1 ,:)d
c l c:).o
+1 o) c:;d
+1 (o. rd
+1 c ] t s ) s c) , : ; ts)oL,? !:c?r qlco's)c¥lco
c o)-co r r r dc co
c¥ico,r)- --co t
-c¥lcoo
o r
c¥1 OhOCO-
a:;cd(O :coLf)c,:;oc;,C,O
-O r(C,c-O,r) rc¥ c tLf)(o-h
c¥]co rO1- tr¥cor)coo
C; :c ia:;c'icl;c id ' ' ' -,Nc:) c',0 rc¥ ,NGoc lc ( rLoc¥l -Lr)
co roh_o)co rcococo
Lf)(c;o:;r (:icic¥ioir(r t-oco rLf)G・c lf)col¥- r
e)hci) rc¥1 rohL,Dc
-cl;tr)coh: : Ftsic;(,:; Fc ir heoG0 rc¥lcN Lo-
=,N
Lo co,oo t
COLO
':)o--ci cT; Fa a:;c i : :Lc;c:; rco r rcocl)LoLs) rcoo)_
-Lr).O)Lf)':) CO r
O)COO
L,DOi= ・ ・(o r¥
rr¥ Lf)
r(LC;r CC,Lr)O) Lr)1r- tCOcoh_ Lo r(:) cF)c¥lhrh- cOO cC!
hcOcNo)hl¥ c・cO
(:)[!)
cOOOr)Col¥ -O
hl¥ CO
CNCN =-
----_h rcoc l,!)ao rocoLr)
co cc; : :cicc;c ic ici,:;oc¥lcol¥ rcor¥coco
Lf)co__c ,
r rl-1-
l:tO,Nr¥coorh
c¥lcocol-tc,),NCOcv,,:¥lc lrhcv,,N
COO,N,:),Oco r
CO Lo r
r- l- * -1-co c:)1-co'N1-1-cot:)
c¥! occ'l r
oo cr)da:;u:;r
(sicicd-h
coo)_ uDc9ocF) r(:)
-co
co : _o) rcoco-
rc9(o,D c: -- c ,D
oc
ocQ
c
s o .: ; . s : rr¥Lo- ,,o o
o
d,:iLc;c¥io ci
cv)coLr)a:;_o-r
---CO.Lr)(Ol tCOGO(:) r toCOL:)LOcC;Cdr- COcO-
rO, tS)C¥lCNc C¥lL,D,NOo. rcD L':; ' ' (c・:;T: l-r¥CO cv)lF'I=*COCOCOcv) L:)C¥:C¥:C¥lCOCf)Cv,0_OO
ododod,:)Od ' '- t
OCOC¥lC Jhl rCO -Cv,': COo)coco,N= to)
oidc¥i-r( Fd
cotr)c Jcoco r,r)-co,N
Lo
co r
'N r- ol¥
05c¥iL,:; t:d,:;tsi
o rcoc J0 ico
-aoc¥1-l¥ t0 Fr¥Cv)C ,:)O r C¥ioiCY)- 'cl; COCF)Cv)L,DCv,(oLr)r O
r ,N
-tco,Nr¥.1:to,o oicNc¥icN-cicYi c )eoci) 'c¥ll ro)coc¥1
-c;)o)- cOui)Lr)_ LOl¥ rts)Lf)OcdcNOOlf),NO
,O-c¥:c¥1-- r
COC') rr- T-a)
. . CO
OOC:)COO C(i
' : LOCO(:)l¥G" C¥drh COf*
T-1-Cll-rG'coO(,) ,:)
,S)O"ot,:; Fr: od
C:) ,N
corh- L!)
:-
Oc¥lc C:) c l co l::__ l:
o
'-o <Q L0 coh
h
a) r co coo
coO
h
,N COh
O) C¥l q:; GO L,-C¥Ih
h
':¥iO
q:; r L:) co CO C¥l ,:) CO ,N CN cdO
O
,N LO : r Lr) r OC; a:) ,:) C lqJ :2 c ' u, c:I J( ,:,
Z
c9 ; o la; 1: O L C:, , (g o y: t: c5 =1
o o o=
; o Q, Q, , l Ja Lo , , ,1, E1
o
o
-¥
E dJ = = o o1
O (g O C ,U O cE io co ; t Lr) ; c¥a t ,Qh
Go a) c LO C:f CO c5 D O O o l a, :E O LL C O o) cc; +1 (o. o) o) co. q:) +1 co F r co : +1 oi c¥l o. cv' +1 Qo.o
r. +[ oi Ls)d
+1 co.h
c ld
+1 Lo :d
+i C , s o ooq:). rr¥r q:)col¥ rq:) r cddci :Lc;c¥i ' 'ddco
-o
c 4r r G0 rGo'Nc¥J r(,' roLoco,N Fl¥-
a)o)o-L,:;(';,:¥i ed F F Foc;c ic;
Ls)o
'N-aocoGoeo,Nc 'Na:)Lo
co r ro-
(oo)Lt)h ro
oc lcia5h:o;dLc;Qir(
o
coo,Na)a) tcoc¥JLor) r
c0 r,:?-OL!)u)cO(c,l¥o
LC;C:;LO(,ir c ic¥iciL,:;e:;ci-Ls)
eo__cl,qouDGDc¥lLo ra)
OGOCOCOcl,a:,hOc,,COO
0:;cd :cc; :c:;Er:;,,{ : F FhCO,NC¥ 1!)C- r
r-cooc¥lLr)o--1-
r-o)Lo-ocooocY,c¥J r
Lor¥(c,Lo rr),,ol:r rLs)-coLorhOcY) rcocd:)cOcOOLS)cO, ) r rCYi ・ F 0:;cc;
-LOCOLS)-CNOCr)
Lr) CO _cor¥
,N,N
-l-C0 r
C¥1 ・ ・1-COCv)a:)CO,OO)
CC;GO(O ' ' ' ' ' '
1-r¥CO1- 'C,O・:t CY)CY)O)OCO r ・-r,・,OT-a:)'1= tC
1-coO cl, r0 rc c:)
:cli q:),,:;dLc;r L,:; coo)co dcc;a:icoh Lr) r-
rLr)Ls)_coLs)r Lo-
Oco-
a)c¥lhl¥_
1 ) o , : r¥cocococ "No) r:cl;da; Fcc;(sio-o(o
rcoc¥1-- -(o
cooco r tl')o)
csia:;c¥ic¥ia,coa5<0h t to)eoa)
-,N,N
thcoco
od :c ic¥icOoh
(:)Lf)coc cdcicl;,N (oLr) r
_-,Nc¥]
co,NOh O
;OG6ci o)
OO
r d
rcOO co
GO_c¥lc¥1(D r_
h(:)O t
Co LO-
FciLti COCi
_ hCOCO
e rcO -C¥l c Lf) thc90(:, 'NL':; qo '; coOa)
tco dc,:,r-'N
c co
rooLo-(:ic,ic:;c¥ied rc¥io rcoLo,Nc h
-c¥1-q:Dc¥Ito- co r
O
dLs,,: r TJ ci
or Loc¥lc¥1(Oh O)--c
l-c¥lo-
rr¥_
todcor rr¥ ' '
f¥ cor o)t-Gr) I ; q:)o)'N- '!:' O) O; aDo
co c { C¥l ,:) LO CO cr)d
r c l QOO
,N ,r) CO C¥l CD c¥i a) o) od [ a)o
c r co o, "D c c Jc5 o I dJ = ,:I O
1
; .S1
o J2 o o=
th o u'c5,
Lo dJ, ' c:' E1
O
O
-¥
E dJ = o dJ = O o1
O o ' E a, ,D LL q: ,DS
H
,!!o L,:, ,5 co co coZ
'o a! o dJ oO
r . a:) +1 c¥l cri C c¥l +1o
c i o) t cri +1 Lo. co c l c¥l +1h
(:s c l r. +1 o. o) r t : +1 Go r Go : +1 c io
a:) : +1 o.h
Lo r +! Lr). : E EJ
h: S ) S c) . ¥* . s :; i : _L!2r coc¥Its)cocoL:?coo ' rur)c¥iC; ' ' ' ' ' -c lcorhts) c¥lcoco-cNcN rc¥1'Ncor d
-,:) trh(0 rr¥Ls) oc¥l
- rr
c:)ocoLr)0 FT- dco
o
C cYir dcN_
--r¥r r¥_(oLoco_
(oco- c lcohc)c'o)
:
(OC :(c;r(c;c i ;r¥-
,Nco'o-
r)oe)_r¥coc)_
oo)r tr coc l -o
o . .o
c: c¥i Fdai ' o:;co rLc;
-co
O'Na:) 'c 1--o tcocv)
c¥lc¥lc¥1--'N-o
,:)codrth'No)oc coo)
r(co_cococococoho
coco CNC¥l ,N o
- -coLo
COLr)(O r C
tO-
CpLt) t
Lf5i
i-CO t ,(OCOGOLr)LO-
c l,:):c :Ci ' '
-Lr) ,co0 toc dcoo-
c i Ls)oo)
c;d a:;c¥lc l Fcic l- '
coco c¥1': rcNc¥ll:rLoco
1-Ftc¥l0 :o)I¥Is) O
Ci(c;c i F ' ' ' ' ' '(Y) Lo -cF)c lc¥lC:)- a:;
Qr)r rc¥lcNcocNc¥lc,,Lo -cC,
C ・ ・,NC¥1 tCOO flr)CO
LC;,N-OC; :c,i Fec; :c,:;d
tO-cr)GOLOc,)C¥lL,)(C,--,N
o !: o G) o-o : (D c c:J: c: 'E ' o ,D ccc: E
- -5 o o o J
0 : :,D,:
h'55 t; G, :: 5 : i ' .- ': > oL>
<JICLHJg: >HH
._ J s c) . s ' * . s : : ) :e e s o : coor¥(o
Lo ol¥.r¥.
dc¥ih:o-
Goooco
co tc¥lco
oco-
coco tJF) cir(a:;r a:; (c; co cooa) foo o
c l -c¥1-r,OL,:,.,0 t(OCOOcoco :LO : ;
O
C¥1-C¥1-CO_
OC0 tC
C¥lh,:)CO tcOOO
:dd :,: L,:;
COLOOOco t r
GOcOC¥lco---C tTc !oc¥J_co0
r¥hcoc r
coco-t Fhc¥a r,Nr¥
hhCOCO
COdoi COOO
Ol¥ dC¥i,d
-LO
_LOCOL,Do
. toor r c¥l
-d '
-ur)F(Yid
co Lo tLr)o_ _ tc¥lco
r¥Ococ¥1(O r r
r a:; ' 'co O_ ,a:;
coc,) to)_ _ tcN,N
0 t,,OC¥lC¥lLO r O)LCiG ici,:) Lc;, i
_(OLr)COC
(OC0 r
)-- 1:
O '
O CD ,5 O O 'D O 0' a): OCCC(UC =
::E5 J' CQ:=
::!QLO
'T:> OC"L
:
5 5(/D<0_
GOh
CO (:) GOo
cs co CO Ci a) Lf)h
oi q:) ,:) Lo co c¥th
co coh
coo
co (Dh
co o_ Qq r coo
,:i ,:)h
co ,a ' OH
Goto
St.Rotifer enriched with AQUARAN
A rtemla enriched with AQUARAN
Commercial diet
Nagasaki
Pref .Rotifer enriched with different kinds of emulsified
Artem/ enriched with different kinds of emulsified
Wiid zooplankton
Frozen zooplankton Eggs of red sea bream Commercial diet
O 10 20
30 40
Days after hatching
50 60