KIAA1199 遺伝子が司る新規ヒアルロン酸分解機構の解析
CBB:Coomassie Brilliant Blue CHC:clathrin heavy chain CPC:cetylpyridinium chloride CS:chondroitin sulfate
DMEM:Dulbecco's modified Eagle's medium DS:dermatan sulfate
DTT:dithiothreitol
EDTA:ethylenediaminetetraacetic acid EGF:epidermal growth factor
Endo H:endo-β-N-acetylglucosaminidase H FA:fluoresceinamine
FBS:fetal bovine serum GAG:glycosaminoglycan
GAPDH:glyceraldehyde 3-phosphate dehydrogenase GlcNAc:N-acetylglucosaimne
GM130:golgi matrix protein of 130 kDa HA:hyaluronan
HARE:hyaluronan receptor for endocytosis HAS:hyaluronan synthase
HEK:human embryonic kidney Hep:heparin
HGF:hepatocyte growth factor
HPLC:high performance liquid chromatography HRP:horseradish peroxidase
HS:heparan sulfate HYAL:hyaluronidase IFN:interferon IL:interleukin
LDS:lithium dodecyl sulfate MEM:minimum essential medium OA:osteoarthritis
PDI:protein disulfide isomerase
PVDF:polyvinylidene difluoride membranes RA:rheumatoid arthritis
RHAMM:receptor for hyaluronan-mediated motility RT:reverse transcription
第1章 正常ヒト皮膚線維芽細胞のHA 分解機構を担う KIAA1199 遺伝子の発見 第1節:序論
組織におけるHA の代謝は非常に速く,皮膚での代謝半減期は 1-1.5 日であり2),強力な HA 分解機構が存在することが予想される.これまでに,中村らは培養ヒト皮膚線維芽細胞 が外的に添加したHA を低分子化することを報告した17).また,申請者らも正常ヒト皮膚 線維芽細胞Detroit 551 株が培養系で旺盛に HA を分解することを見出した(Fig. 1-1A). しかしながら,その活性は繊細であり,細胞溶出液では消失することが大きな障壁となり, これまで皮膚のHA 分解研究の進展を阻んでいた.一方,組織の HA 分解仮説モデルとし て,CD44(HA 受容体)と 2 種類の HA 分解酵素(HYAL1 および HYAL2)が共同的に働 くという説が有力であった12).しかし,本モデルが組織のHA 分解に関わることを示す直 接的なエビデンスは乏しく,未だHA 分解機構については不明な点が多いのが現状である. そこで本研究では,生理的な皮膚の HA 分解機構を明らかにすることを目的とし,まずは 正常皮膚線維芽細胞を用いて HA 分解仮説モデルの検証を行った.その結果,皮膚線維芽 細胞にはCD44,HYAL1 および HYAL2 が関与しない,新規な HA 分解機構が存在すると 考えられたため,HA 分解に関わる未知の分子の同定を目的とし,線維芽細胞の培養系 HA 分解活性を指標に,その活性と発現の挙動が一致する遺伝子を包括的に探索した. 第2節:実験材料および実験方法 1-2-1 細胞培養
胎児由来正常ヒト皮膚線維芽細胞Detroit 551 株(American Type Culture Collection 社 製),HS27 株(American Type Culture Collection 社製)および NHDF-Ad 株(Takara Bio 社製)は10%(v/v)FBS(JRH Biosciences 社製)を含有する MEM 培地を用いて培養し た.HEK293 細胞(DS Pharma Biomedical 社製)および COS-7(DS Pharma Biomedical 社製)は10%(v/v)非働化 FBS,100 units/ml ペニシリンおよび 100 mg/ml ストレプト マイシンを含有するDMEM 培地を用いて培養した.
1-2-2 [3H]標識高分子 HA の調製
殿を行った.最終的に得られた沈殿画分を,Sepharose CL-2B ゲル濾過カラム (1 x 60 cm) に供与し,得られたVo 画分(> 1,000 kDa)を精製高分子[3H]HA とした.[3H]HA の放射 比活性は1.3-2.7 x 105 dpm/mg(2.2-4.5 kBq/mg)であり,Streptomyces hyaluronidase (Seikagaku Corporation 社製)にて消化されることを確認した(結果省略). 1-2-3 培養系 [3H]HA 分解活性の評価法 コンフルエントの細胞に40,000 dpm/ml の[3H]HA を添加し,一定期間培養後に得られ た培養上清をSepharose CL-2B ゲル濾過カラム(1 x 60 cm)に供与した.流速は 0.65 ml/min とし,2.5 ml/フラクションで分取後,シンチレーションカウンター(Aloka LSC-6100)にて各フラクションの放射活性を測定し,[3H]HA 分解物の分子量を評価した. カラムのキャリブレーションにはフルオレセインアミン蛍光標識(FA-)HA(FA-HA H1 (1,760 kDa),M1(907 kDa), L1(197 kDa), S1(56 kDa),T1(28 kDa),U1(9.8 kDa))(PG Research 社製)を用い,検出は励起波長:490 nm,蛍光波長:525 nm によ り行った.
1-2-4 マイクロアレイ遺伝子発現解析
ヒスタミン(10 mM)あるいは transforming growth factor (TGF)-β1(10 ng/ml)で刺 激したDetroit 551 線維芽細胞から,8 時間および 24 時間後に total RNA を抽出し,Agilent 44K4 chips(Agilent Technologies 社製)を用いて遺伝子発現を調べた.結果は,各遺伝子 の無刺激区と刺激区との発現比(倍)にて示した.
1-2-5 培養系HA 分解活性の反応速度の算出
[3H]HA と混合した 100,300,600 および 1,250 mg/ml の非標識ヒト臍帯高分子 HA(分 子量800~1,200 kDa)(Seikagaku Corporation 社製)を Detroit 551 線維芽細胞に添加後, 10 mM ヒスタミン存在下あるいは非存在下で 72 時間培養した.得られた培養上清を Sepharose CL-2B ゲル濾過カラムに供し,高分子 HA 画分の放射活性を測定した.[3H]HA 分解活性は高分子HA の減少量とし,次の式にて算出した.培養系[3H]HA 分解活性(mg/105 cells/72 h)=細胞に添加した HA 量(mg)x(培養前の高分子 HA 画分の放射活性―72 時 間培養後の高分子HA 画分の放射活性)/細胞に添加した全放射活性/細胞数(x 105 cells). 1-2-6 抗体
原とし,同ペプチドを用いてアフィニティー精製した.抗CD44 および抗 glyceraldehyde 3-phosphate dehydrogenase (GAPDH)抗体は Santa Cruz Biotechnology 社から購入した. コントロールラットIgG2aκは SouthernBiotech 社より購入した. 1-2-7 培養系HA 分解系の基質特異性 Detroit 551 線維芽細胞を 10 mg/ml FA-GAG(FA-HAs(H1,M1,L1,S1,T1 および U1),FA-コンドロイチン硫酸 A,C,D(FA-CSA,FA-CSC,FA-CSD),FA-デルマタン 硫酸(FA-DS),FA-ヘパリン(FA-Hep),FA-ヘパラン硫酸(FA-HS))(PG Research 社 製)を含む培地で72 時間培養後,得られた培養上清を Sepharose CL-6B ゲル濾過カラム (1 x 35 cm)に供与した.流速は 0.4 ml/min とし,1.6 ml/フラクションで分取後,各フ ラクションの蛍光強度(励起波長:490 nm,蛍光波長:525 nm)を測定して GAG の分解 を評価した. 1-2-8 ウェスタンブロット解析法
細胞溶出液をNuPAGE 4-12% Bis-Tris gels(Invitrogen 社製)に供与し,電気泳動後, 分離したタンパク質をPVDF 膜に転写した.転写後の PVDF 膜は抗 KIAA1199,抗 CD44, 抗HYAL2 あるいは抗 GAPDH 抗体と反応させた.抗 KIAA1199 抗体の検出には HRP 標 識抗ラットIgG 抗体(Jackson ImmunoResearch 社製),抗 CD44,抗 HYAL2 および抗 GAPDH 抗体の検出には HRP 標識抗ラビット IgG 抗体(DAKO 社製)を用いた.バンド の検出には SuperSignal West Pico Chemiluminescent Substrate system (Thermo Scientific 社製)を用いた.
1-2-9 RT-PCR 法およびリアルタイム PCR 法
Total RNA の抽出は RNeasy(Qiagen 社製)を用い,逆転写反応には High Capacity cDNA Archive Kit(Applied Biosystems 社製)を用いた.HYAL1(GenBank accession No, U_96078),HYAL2(U_09577),CD44(NM_000610)および GAPDH(NM_002046) (内部標準遺伝子)mRNA の検出には以下の PCR プライマーを用いた.
HYAL1 sense, 5’-CAGGCGTGAGCTGGATGGAGA-3’; HYAL1 anti-sense, 5’-GTATGTGCAACACCGTGTGGC-3’; HYAL2 sense, 5’-GAGTTCGCAGCACAGCAGTTC-3’; HYAL2 anti-sense, 5’-CACCCCAGAGGATGACACCAG-3’; CD44 sense, 5’-TCCCAGTATGACACATATTGC-3’; CD44 anti-sense, 5’-CACCTTCTTCGACTGTTGAC-3’; GAPDH sense, 5’-ACCACAGTCCATGCCATCAC-3’; GAPDH anti-sense, 5’-TCCACCACCCTGTTGCTGTA-3’.
FGD4-2, 5’-AUGUGAUGCUGCAAAGUUAAGCUCC-3’; TMTC1-1, 5’-UAAUACAGUGCUCCCAAAGGUGACA-3’; TMTC1-2, 5’-UUUGGUUUCAGCUGGAGAGCCUUGU-3’; RASL11A-1, 5’-UAGACCAGCCGUGAAUACAGCUUGC-3’; RASL11A-2, 5’-AUUUGGACAGGGAAUCGACGACCUG-3’; C6orf32-1, 5’-AUACCAGGUGAUUUCCAGGUUCAGU-3’; C6orf32-2, 5’-UUUCAAGGCCCUGUAGACUUCUUCC-3’; AK024243-1, 5’-UACUCUUGGUCCCUGAUCAAAGUCC-3’; AK024243-2, 5’-AUCAGUCUGAGUUAAAGGCCAGCUC-3’; FLJ35700-1, 5’-UUUCCAAGGGCAGAGCCAGCAUUUG-3’; FLJ35700-2, 5’-UGAGAAGUCUGUUCUCGUCACUCUG-3’; FLJ36644-1, 5’-UUAACUGGGACCACAUCGCCAUUUG-3’; FLJ36644-2, 5’-UUUAUUAUGACCUUCCUCCCGCUGC-3’; KLHL23-1, 5’-AAACAUGUAACACCAUAGCUCUCCC-3’; KLHL23-2, 5’-UAUAGAUCCACACUGUGUCAAGAGC-3’; BX350256-1, 5’-UAGGCCUGCAUAAUACAGUCCUAUG-3’; BX350256-2, 5’-UAAUGUGGGAUCCUUAUCAGCAUGC-3’; ENST00000372493-1, 5’-AAUAUAGGCUACGCAGUUGUGGAGG-3’; ENST00000372493-2, 5’-UGAUCCUCCCAGUUUCUUCUUGCUC-3’; THC2404909-1, 5’-UCAUCUUUCACUGGAGCCCGGAAGC-3’; THC2404909-2, 5’-UUUCCAAGGAGUCACGUGGUCGCUC-3’; A_32_P171043-1, 5’-AGAUGUCGGGCAUUAACUCAGAAGG-3’; A_32_P171043-2, 5’-UUUACUUGUUGGGCUAUGAUACUCC-3’. HYAL2およびCD44に対する2 種類の siRNA(Invitrogen 社製)は以下を用いた. HYAL2-1, 5’-AAUCUUUGAGGUACUGGCAGGUCUC-3’; HYAL2-2, 5’-AUAUUGUGCCUGUUUGACUAUGCGG-3’; CD44-1, 5’-AUUCAAAUCGAUCUGCGCCAGGCUC-3’; CD44-2, 5’-AAAUGCACCAUUUCCUGAGACUUGC-3’;
これらのsiRNA は Lipofectamine RNAiMAX(Invitrogen 社製)を用いて細胞に導入した. 1-2-11 遺伝子発現プラスミドの構築と細胞への導入法
後48 時間の細胞を用いた.組換え DNA実験は㈱カネボウ化粧品の倫理審査で承認された. 1-2-12 ヒト皮膚試料の調製
正常ヒト皮膚試料は,Stephens & Associates, Inc.(Dallas, Colorado, USA)にて,3 名 のボランティア(女性:61-65 歳)の顔面目尻より,バイオプシーパンチにより採取した. 皮膚の採取はStephens & Associates, Inc.および㈱カネボウ化粧品の倫理審査で承認され, インフォームドコンセントは手術前にボランティアより得た.
1-2-13 免疫組織染色法
免疫組織染色はホルマリン固定したヒト皮膚組織のパラフィン切片を用いて実施した. 抗原賦活には,切片試料を再水和後,1 mM EDTA(pH 8.0)において,95°C で 15 分処理 した.内因性のペルオキシダーゼ活性は,3%(v/v)H2O2を含むPBS で 15 分処理して失 活させた.ブロッキングには,10%(v/v)donkey serum を含む BSA/PBS-T(1%(w/v) BSA を含む PBS-T)を用い,4 mg/ml の抗 KIAA1199 抗体もしくはコントロールラット IgG2aκと 4°C で 18 時間反応させた.PBS で洗浄後,BSA/PBS-T で 500 倍希釈した HRP 標識抗ラットIgG 抗体 (Jackson ImmunoResearch 社製) と室温で 1 時間反応させ,発色 はVector® VIP kit(紫色; Vector Laboratories 社製)で行った.
1-2-14 in situハイブリダイゼーション法
Digoxigenin 標識した KIAA1199 ア ンチセンスおよ びセンスプローブ(GenBank accession No. NM_018689,nucleotides 5729-6352)を合成し,in situハイブリダイゼー ションはホルマリン固定した正常ヒト皮膚組織のパラフィン切片を用いて実施した19). 1-2-15 統計処理
統計解析はSAS システムにより実施した.Dunnett’s multiple comparison test でコン トロールとの有意性を解析した.P値0.05 未満を統計的に有意とした.
第3節:実験結果
1-3-1 培養皮膚線維芽細胞を用いたCD44,HYAL1 および HYAL2 依存的な HA 分 解仮説モデルの検証
551 線維芽細胞おけるこれらの分子の発現を RT-PCR で調べたところ,CD44 および HYAL2は発現していたが,HYAL1 の発現は認められなかった(Fig. 1-1B).次に,発現
Fig. 1-1. HA degradation by cultured normal human skin fibroblasts independent of CD44, HYAL1 and HYAL2. (A) Detroit 551 skin fibroblasts were cultured with [3H]HA for 48 h and HA fragments in the culture medium were examined by size exclusion chromatography. (B) Expression of CD44, HYAL2 and HYAL1 by RT-PCR. GAPDH, a loading control. (C and D) CD44 and HYAL2 were knocked down by treating cells with siRNAs to CD44 or HYAL2. For controls, the cells were transfected with control non-silencing siRNA (Control siRNA). Cells with siRNA were cultured with [3H]HA for 48 h, and degraded HA was examined by size exclusion chromatography. Knockdown efficiency for CD44 and HYAL2 was evaluated by immunoblotting (Insets). GAPDH, a loading control. Representative data for two siRNAs are shown.
Fig. 1-2. Endogenous HA-specific depolymerization by Detroit 551 fibroblasts in culture. The cells were cultured with FA-GAGs: FA-HA H1, FA-CSA, FA-CSC, FA-CSD,
が認められたCD44およびHYAL2についてsiRNA を用いて発現抑制した結果,いずれの 発現抑制においてもHA 分解には全く影響が認められなかった(Fig. 1-1C, D).これらの ことから,皮膚線維芽細胞にはCD44,HYAL1 および HYAL2 が関与しない,新規な HA 分解機構が存在すると考えられた. 1-3-2 培養皮膚線維芽細胞のHA 分解に関わるKIAA1199遺伝子の発見 線維芽細胞のHA 分解を担う新規な分子の同定を目的とし,皮膚線維芽細胞の培養系 HA 分解活性と発現の挙動が一致する遺伝子の探索を試みた.まずはinterleukin (IL)-1α,IL-1β, TGF-α,TGF-β1,epidermal growth factor (EGF),hepatocyte growth factor (HGF), interferon (IFN)-γ,tumor necrosis factor (TNF)-αおよびヒスタミンで Detroit 551 線維芽 細胞を刺激したところ,培養系HA 分解活性はヒスタミンにより顕著に亢進し,TGF-β1 に
Fig. 1-3. HA degradation mediated by KIAA1199 and regulation of KIAA1199 expression
by histamine and TGF-β1. (A) Effect of histamine and TGF-β1 on HA depolymerization.
Cells were treated with or without histamine (10mM) or TGF-β1 (10 ng/ml) and cultured with [3H]HA for 48 h. The HA degrading activity was analyzed by chromatography. Control, untreated cells. (B) Abrogation of HA degrading activity by knockdown of KIAA1199 with two different siRNAs specific for KIAA1199. The cells were cultured with [3H]HA for 48 h, and HA degradation was determined. Inset, immunoblotting for KIAA1199 and GAPDH (a loading control). (C) Kinetic study of HA degradation by cells treated with or without histamine for 72 h. Control and Histamine, cells treated with vehicle alone and histamine, respectively. (D and
E) The expression levels of KIAA1199 mRNA and protein in cells treated with histamine,
1-3-3 KIAA1199 cDNA を導入した細胞の HA 分解能 HA 分解における KIAA1199 の機能を調べるため,培養系で HA を分解しない HEK293 細胞とCOS-7 細胞にKIAA1199の 全長cDNA を導入した.その結果, KIAA1199 cDNA を一過的に導入 した細胞は,[3H]HA を 10-100 kDa の中間サイズの HA 断片に分解し たのに対し,コントロールのプラス ミドを導入した細胞は HA 分解活 性を示さなかった.これらの結果か ら,KIAA1199遺伝子を導入した細 胞は,皮膚線維芽細胞と同様のHA 分解能を新たに獲得したことが分 かった(Fig. 1-4A, B).以上より,
Table 1-1. Genes up-regulated by histamine and down-regulated by TGF-β1 in Detroit 551
skin fibroblasts.
Detroit 551 cells were cultured in the presence and absence of 10 mM histamine or 10 ng/ml TGF-β1. At 8 h and 24 h after the treatment, total RNAs were isolated and subjected to microarray gene expression analysis. Values (fold induction) represent the ratio of expression levels in the stimulated cells compared with the un-treated control cells.
Fig. 1-4. HA depolymerization by KIAA1199
transfectants. (A and B) HEK293 and COS-7 cells were
先述の皮膚線維芽細胞における KIAA1199 発現抑制実験の結果(Fig. 1-3B)と合わせ, KIAA1199 は HA 分解に必須な因子と考えられた.
1-3-4 正常ヒト皮膚の線維芽細胞におけるKIAA1199 の発現
in situハイブリダイゼーション法(Fig. 1-5A)および免疫組織染色法(Fig. 1-5B)によ り,KIAA1199 は主に正常ヒト皮膚真皮の線維芽細胞で発現していることを見出した.一 方,表皮細胞において,KIAA1199 mRNA の発現は認められたが(Fig. 1-5A),KIAA1199 タンパク質の発現は認められず(Fig. 1-5B),KIAA1199 は表皮細胞において機能していな い可能性が考えられた. 第4 節:考察 これまで,CD44,HYAL1 および HYAL2 が HA 分解における主要な役割を担う因子と 考えられてきた.しかし,今回の研究により HYALs や CD44 が関与しない,KIAA1199 依存的な新規HA 分解機構が存在することが明らかとなった.全身の HA のターンオーバ ーにおいて,末梢の組織で合成された分子量1,000 kDa 以上の高分子 HA は,まずは組織 において10-100 kDa の中間サイズの HA に分解され,その大部分がリンパ管に入り,リン パ節にて HYAL1 により更なる分解を受ける 2).リンパ管で分解されなかった HA(~10 kDa)は血管に運ばれ,最終的には HYAL1 や HYAL2 が高発現している肝臓,腎臓および 脾臓において9, 13),HARE を介したクラスリン経路で細胞内に取り込まれ,リソソームに て完全に消化されると考えられている2).これまでに,HYAL1 欠損によって引き起こされ るIX 型ムコ多糖症患者や,HYAL2 ノックアウトマウスの血清においては HA レベルが上 昇することが報告されている13, 16).さらに,IX 型ムコ多糖症患者のマクロファージや線維 芽細胞13),またHYAL1 ノックアウトマウスの軟骨細胞のリソソームにおいては HA が蓄 積することが報告されている15).これらのことから,HYAL1 や HYAL2 はリンパ節,肝臓, 腎臓および脾臓などの臓器や,マクロファージ,線維芽細胞および軟骨細胞などにおいて,
Fig. 1-5. Expression of KIAA1199 by dermal fibroblasts in normal h u m a n s k i n . ( A ) T h e m R N A
第2章 KIAA1199 が司る HA 分解機構の特性解析 第1節:序論
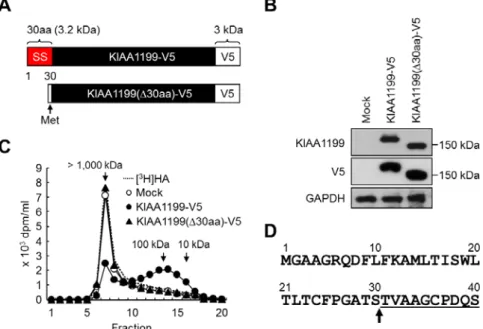
KIAA1199遺伝子は,1,361 のアミノ酸からなる分子量 153 kDa のタンパク質をコード する4,083 bp の open reading frame(ORF)を含んでいる24).KIAA1199 のアミノ酸配 列はHYAL を含む他のタンパク質と相同性を示さず9, 12),また複数のHA 結合タンパク質 で共通に認められるHA-リンクモジュール25)やB(X7)B HA-結合モチーフ26)も認められな い.一方,KIAA1199 には,細胞外リガンドとの結合が推測されている G8 ドメイン27), 高分子多糖の加水分解への関与が示唆されているPbH1 ドメイン28),および機能が不明の GG ドメイン29)が存在することが報告されている(Fig. 2-1).また,KIAA1199 は 7 つの 推定N型糖鎖修飾部位を有し,N 末端 30 アミノ酸は疎水性の高いアミノ酸で構成されてい ることから,シグナル配列と予想されている28).しかし,これまでにKIAA1199 分子の機 能については明らかにされていない. そこで本研究では,新たに樹立した KIAA1199 安定発現 HEK293 細胞を用 い,KIAA1199 の機能および培養系で のHA 分解の様式,さらに KIAA1199 タンパク質の成熟化と細胞内局在制御 について詳細に検討した. また,マウスにはヒトKIAA1199と DNA 配列で 86.8%,アミノ酸配列で 91%の相同性を示すホモログ遺伝子 (mKiaa1199)が存在する 24).ヒト KIAA1199 と 同 様 , こ れ ま で mKiaa1199の機能は明らかにされてい ないため,本研究では,mKiaa1199 の 機能についても検討した. 第2節:実験材料および実験方法 2-2-1 細胞培養 HEK293 細胞は第1章で記載した方法に従い培養した. 2-2-2 KIAA1199-V5およびKIAA1199(∆30aa)-V5遺伝子発現プラスミドの構築 C 末端に V5 タグを有する KIAA1199(KIAA1199-V5)の発現プラスミドは,KIAA1199 の全長cDNA を pcDNA3.1/V5-His プラスミド(Invitrogen 社製)に挿入して構築した.N
Fig. 2-1. The schematic representation of the structure of full-length KIAA1199. Functional
末端30 アミノ酸を欠失した KIAA1199-V5(KIAA1199(∆30aa)-V5)の遺伝子発現プラス ミドは,1番目のメチオニン残基から30 番目のセリン残基を ATG 開始コドンで置換して 作製した.組換えDNA 実験は㈱カネボウ化粧品の倫理審査で承認された.
2-2-3 マウスKiaa1199 ホモログ遺伝子(mKiaa1199)発現プラスミドの構築 mKiaa1199 cDNA(GenBank accession No. AB_103331)は,マウス脳由来 cDNA (Takara Bio 社製)を鋳型として RT-PCR により増幅し,塩基配列は Applied Biosystems 3730xl DNA Analyzer(Life Technologies 社製)を用いて確認した.mKiaa1199発現プラ スミドは,mKiaa1199 cDNA を pcDNA3.1(-)プラスミド(Invitrogen 社製)に挿入して構 築した.組換えDNA 実験は㈱カネボウ化粧品の倫理審査で承認された.
2 - 2 - 4 KIAA1199 遺 伝 子 安 定 発 現 細 胞 (KIAA1199/HEK293 細 胞 ) お よ び mKiaa1199遺伝子安定発現細胞(mKiaa1199/HEK293 細胞)の樹立
KIAA1199あるいはmKiaa1199発現プラスミドをHEK293 細胞に導入し,800 mg/ml の G418(Sigma 社製)を含有する培地にて,プラスミドが導入されたクローンを選抜した. さらにその中から,ウェスタンブロット法により KIAA1199(mKiaa1199)タンパク質を 発現し,かつ培養系HA 分解活性を有する細胞を取得した.
2-2-5 抗体
ター(Roche Diagnostics 社製)を含む 50 mM リン酸バッファー(pH 6.0)中で破砕し, 細胞溶出液を調製した.得られた細胞溶出液を,非標識 GAG(CSA,CSC,CSD,DS, Hep,HS),あるいは非標識 HA(HA-H2 (1,452 kDa),HA-M2(1,039 kDa),HA-L2 (219 kDa),HA-S2(52 kDa),HA-T2(28 kDa))(PG Research 社製)と 37°C で 1 時 間インキュベートし,GAG と結合タンパク質との複合体を形成させた.ネガティブコント ロールには超純水を用いた.GAG-タンパク質複合体を沈殿させるために最終濃度が 1% (w/v)になるよう CPC を加え,更に 37°C で 1 時間インキュベートした.遠心して回収し た沈殿画分を,1%(w/v)CPC を含む 30 mM NaCl 溶液で 3 回洗浄後,NuPAGE LDS サ ンプルバッファー(Invitrogen 社製)に溶解した.得られた試料を NuPAGE 4-12% Bis-Tris gels(Invitrogen 社製)に供し,沈殿画分の KIAA1199(mKiaa1199)を,抗 KIAA1199 抗体を用いたウェスタンブロット法により検出した.
2-2-9 KIAA1199 の GAG 結合活性評価-2
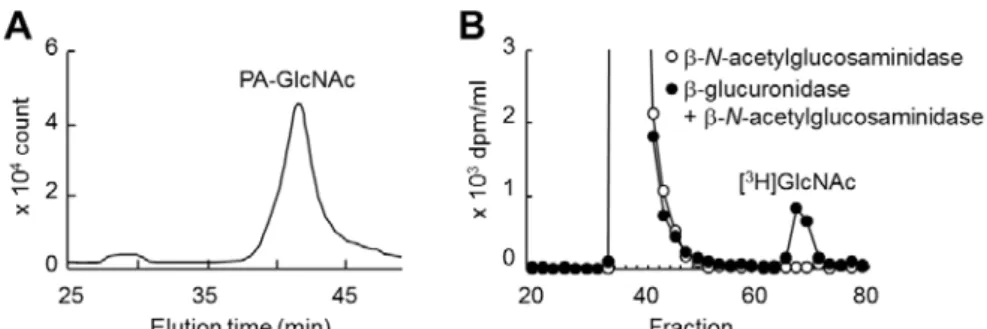
G-25 ゲル濾過カラム(1 x 107 cm)(GE Healthcare 社製)に供して評価した.GlcNAc の溶出位置は既存の方法31)に従い決定した. 還元末端糖の同定には,KIAA1199/HEK293 細胞を 100 mg/ml の非標識ヒト臍帯 HA を 含む培地で72 時間培養し,得られた培養上清に含まれる HA 分解産物を,Sepharose CL-2B ゲル濾過カラムを用いて回収した.HA 分解産物の還元末端糖は,2-アミノピリミジンによ りピリジルアミノ化(PA)し32),得られたPA-HA は 4 N HCl により単糖にまで加水分解 した.その際,N-アセチル基の脱離により生じたグルコサミンは,再度 5%(v/v)無水酢 酸にてN-アセチル化した33).得られたPA-GlcNAc は,TSK-gel Sugar AXI (内径 4.6 mm x 15 cm; Tosoh Corporation 社製)を用いた逆相 HPLC にて分離し,励起波長:310 nm,蛍 光波長:380 nm で検出した.
2-2-11 RNA 干渉法
2-2-13 免疫沈降法
KIAA1199/HEK293 細胞を,1%(v/v)Nonidet P-40 およびプロテアーゼインヒビター (Roche Diagnostics 社製)を含む 50 mM Tris-HCl バッファー(pH 8.0)中で破砕し,細 胞溶出液を調製した.得られた細胞溶出液中のKIAA1199 は,抗 KIAA1199 抗体を結合さ せたDynal magnetic beads protein G(Invitrogen 社製)を用いて免疫沈降した.ネガテ ィブコントロールにはラットIgG2aκを用いた.ビーズに結合したタンパク質は,NuPAGE LDS サンプルバッファー(Invitrogen 社製)で回収し,NuPAGE 4-12% Bis-Tris gels (Invitrogen 社製)で電気泳動後,抗 KIAA1199 および抗 CHC 抗体を用いたウェスタン 法による解析にて評価した.
2-2-14 各種阻害剤の培養系[3H]HA 分解への影響
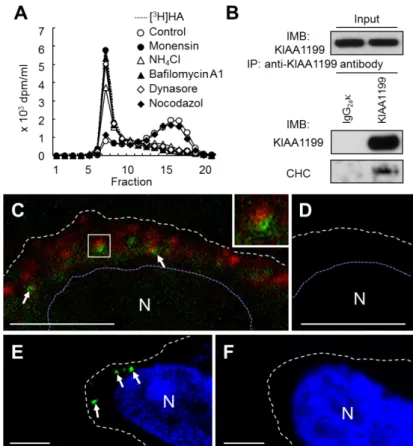
KIAA1199/HEK293 細胞を 5 mM モネンシン(MP Biomedicals 社製),10 mM NH4Cl (Wako 社製),1 nM バフィロマイシン A1(Calbiochem 社製),60 mM Dynasore(Merck 社製)あるいは0.3 mM ノコダゾール(Calbiochem 社製)を含む培地で 3 時間培養後, [3H]HA を添加し,さらに 6 時間培養した.得られた培養上清を Sepharose CL-2B ゲル濾 過カラム(1 x 60 cm)に供与し,HA 分解活性を評価した. 2-2-15 外因性HA の細胞内局在解析-1 マルチチャンバースライドで培養したKIAA1199/HEK293 細胞を,0.1 mg/ml のビオチ ン標識高分子HA(PG research 社製)を含む培地で 1 時間インキュベートした.コントロ ールとして,予めStreptomyces hyaluronidase(Seikagaku Corporation 社製)で消化し たビオチン標識高分子HA を用いた.細胞を 4%(w/v)パラホルムアルデヒドを含む PBS で固定後,ストレプトアビジン標識 Alexa-Fluor 488(Invitrogen 社製)でビオチン標識 HA を標識した.TO-PRO-3 にて核を染色後,試料を vectashield にて包埋し,Zeiss LSM 510 共焦点顕微鏡を用いて観察した.
2-2-16 外的に添加したHA の細胞内局在-2
2-2-17 N 末端アミノ酸配列解析
KIAA1199-V5 発現細胞を,1%(v/v)Nonidet P-40 およびプロテアーゼインヒビター (Roche Diagnostics 社製)を含む 50 mM Tris-HCl バッファー(pH 8.0)中で破砕し,細 胞溶出液を調製した.細胞溶出液に含まれるKIAA1199-V5 は,抗 KIAA1199 抗体を結合 させたDynal magnetic beads protein G(Invitrogen 社製)により,濃縮・精製した.ビ ーズに結合したKIAA1199-V5 は,NuPAGE LDS サンプルバッファー(Invitrogen 社製) で回収し,NuPAGE 4-12% Bis-Tris gels(Invitrogen 社製)で電気泳動後,PVDF 膜に転 写した.PVDF 膜を CBB 染色後,KIAA1199-V5 に相当するバンドを切出し,N 末端から 10 番目のアミノ酸までを,エドマン分解法により決定した. 2-2-18 脱糖鎖修飾 KIAA1199-V5 あるいは KIAA1199(∆30aa)-V5 発現細胞を,プロテアーゼインヒビター (Roche Diagnostics 社製)を含む 50 mM リン酸バッファー(pH 7.0)中で破砕し,細胞 溶出液を調製した.N-グリカナーゼによる脱糖鎖処理には,細胞溶出液を 0.1%(w/v)SDS および50 mM β-メルカプトエタノール存在下で 5 分間ボイル後,0.1 U/ml のN-グリカナ ーゼ(Glyco 社製)により,37°C で 20 時間インキュベートした.Endo H 処理による脱糖 鎖処理には,細胞溶出液を0.5%(w/v)SDS および 40 mM DTT 存在下で 10 分間ボイル 後,25,000 U/ml の Endo H(New England BioLabs 社製)により,37°C で 20 時間イン キュベートした.得られた試料をNuPAGE 6% Tris-glysine gels(Invitrogen 社製)に供 し,抗KIAA1199 抗体を用いたウェスタンブロット法により KIAA1199 タンパク質を検出 した.
2-2-19 mKiaa1199/HEK293 細胞および KIAA1199/HEK293 細胞による HA 分解 産物の分子量評価
mKiaa1199/HEK293 細胞および KIAA1199/HEK293 細胞を,FA-HA H1(10 mg/ml) を含む培地で72 時間培養後,得られた培養上清を,0.8 M NaNO3を含む0.05M CAPS バ ッ フ ァ ー (pH10.0)で平衡化した TSK-GEL G5000PW(7.5 mm x 30 cm; Tosoh Corporation 社製)に供与した.流速は 0.5 ml/min とし,蛍光強度にて HA を検出した. カラムのキャリブレーションにはFA-HA(FA-HA H1(1,760 kDa),M1(907 kDa), L1 (197 kDa),S1(56 kDa),T1(28 kDa),U1(9.8 kDa),3K(2.6 kDa))(PG Research 社製)を用いた.
第3節:実験結果
2-3-1 KIAA1199 分子の機能解析
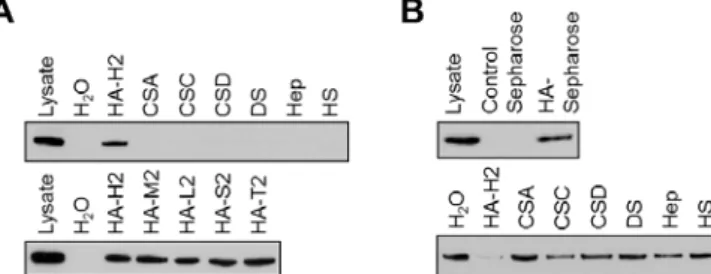
で評価した.第一に,KIAA1199/HEK293 細胞の溶出液と GAG をインキュベートし,生 じた GAG-タンパク質複合体を CPC で沈殿させ,KIAA1199 と GAG との共沈を抗 KIAA1199 抗体を用いたウェスタンブロット法により評価した.その結果,高分子 HA-H2 (1,452 kDa),CSA,CSC,CSD,DS,Hep,HS のうち,KIAA1199 は HA-H2 とのみ 共沈し,他の GAG とは共沈しなかった(Fig. 2-2A,上のパネル).また,KIAA1199 は HA-H2 だけでなく,それより低分子の HA(HA-M2(1,039 kDa),HA-L2(219 kDa), HA-S2(52 kDa),HA-T2(28 kDa))とも共沈した(Fig. 2-2A,下のパネル).次に, HA-Sepharose ビーズを用いて,KIAA1199 の HA への結合活性を評価した.Fig. 2-2B(上 のパネル)に示した通り,KIAA1199 はコントロールのビーズには吸着せず,HA-Sepharose ビーズに吸着した.また,この吸着はKIAA1199 を前もってフリーの HA とインキュベー トすることで競合的に阻害されたが,他のGAG では阻害されなかった(Fig. 2-2B,下の パネル).以上の結果より,KIAA1199 は種々の分子量の HA と特異的に結合する性質を有 すると考えられた. 2-3-2 KIAA1199 安定発現細胞の HA 分解様式
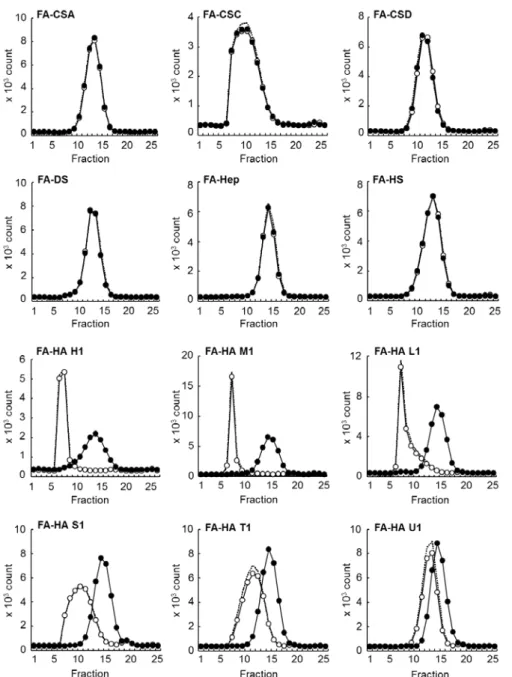
KIAA1199 安定発現細胞(KIAA1199/HEK293 細胞)を用い,KIAA1199 を介した HA 分解機構の基質特異性を調べた.KIAA1199/HEK293 細胞と種々の GAG をインキュベー トした結果,KIAA1199/HEK293 細胞は FA-HA H1(1,760 kDa)を選択的に分解したが, 他の FA-GAG(FA-CSA,FA-CSC,FA-CSD,FA-DS,FA-Hep,FA-HS)は全く分解し なかった(Fig. 2-3).また,KIAA1199/HEK293 細胞は FA-HA H1 だけでなく,それより 低分子のFA-HA(M1(907 kDa),L1(197 kDa),S1(56 kDa),T1(28 kDa),U1(9.8 kDa))も分解した(Fig. 2-3).また,KIAA1199/HEK293 細胞により分解された HA の還 元末端糖および非還元末端糖は,それぞれN-アセチルグルコサミン(GlcNAc)(Fig. 2-4A)
Fig. 2-2. HA-specific binding of KIAA1199 in KIAA1199/HEK293 cells. (A) GAG-binding assay
とグルクロン酸(Fig. 2-4B)だったことから,HA の切断部位はβ-エンド-N-アセチルグル コサミン結合であることが示された.以上の結果より,KIAA1199 が司る HA 分解機構は, HA 特異的なエンド-β-N-アセチルグルコサミニダーゼ様式であり,高分子だけでなく低分 子量のHA も分解し得ると考えられた.
Fig. 2-3. HA-specific depolymerization by KIAA1199/HEK293 cells. Control HEK293 (open
2-3-3 HA のエンドサイトーシス経路および分解の場の同定
これまでに,HARE(HA receptor for endocytosis)依存的 HA 分解機構と CD44 依存的 HA 分解機構は,それぞれクラスリン被覆ピットとカベオラを介していることが報告されて
いる2, 11).そこで,KIAA1199 依存的 HA 分解機構への,クラスリン経路およびカベオラ
経路の関与を調べた.まずは,クラスリン重鎖(CHC)およびクラスリンのアダプタータ ンパク質であるα-アダプチン複合体のサブユニットの一つ AP-2 の発現を siRNA により発 現抑制したところ,KIAA1199/HEK293 細胞の培養系 HA 分解は顕著に低下した(Fig. 2-5A, B).一方,カベオラの構成因子の一つであるカベオリン-1 の siRNA 依存的発現抑制は HA 分解に影響を与えなかった(Fig. 2-5C).これらのことから,KIAA1199 依存的 HA 分解に はクラスリン経路が関与していることが示唆された.
次に,HA 分解の場の同定を目的とし,各種阻害剤が KIAA1199/HEK293 細胞の HA 分
Fig. 2-4. Determination of HA cleavage sites. (A and B) Determination of the reducing and
non-reducing terminal sugars of depolymerized HA. HPLC pattern of pyridylaminated N-acetylglucosamine (PA-GlcNAc) was obtained from HA depolymerized by KIAA1199/HEK293 cells (A). Sephadex G-25 column chromatogram of depolymerized [3H]HA after digestion withβ-N-acetylglucosaminidase (open circles) or sequential digestion withβ-glucuronidase digestion and then β-N-acetylglucosaminidase (closed circles) (B).
Fig. 2-5. Clathrin-specific HA depolymerization in KIAA1199/HEK293 cells. (A-C) Clathrin
Fig. 2-6. Cellular localization of KIAA1199 and exogenously added HA in KIAA1199/HEK293 cells. (A) Effects of inhibitors on HA depolymerization. KIAA1199/HEK293
KIAA1199 抗体による免疫沈降実験では,CHC は KIAA1199 と共沈し(Fig. 2-6B), KIAA1199/HEK293 細胞を用いた KIAA1199 と CHC の二重染色では,両分子のシグナル は原形質膜付近のベシクルに観察され,いくつかのベシクルで KIAA1199 と CHC が近接 していることを見出した(Fig. 2-6C, D).また,KIAA1199/HEK293 細胞に添加したビオ チン標識高分子 HA は,共焦点顕微鏡観察により,原形質膜付近のベシクルに認められ, そのシグナルはHA をStreptomyces hyaluronidase で前もって消化した場合には認められ なかった(Fig. 2-6E, F).一方,KIAA1199/HEK293 細胞に添加した外因性[3H]HA は,そ の大部分が培養上清に検出され,細胞内および細胞表面への蓄積は認めらなかった(Table 2-1 ). 加 え て , [3H]HA は KIAA1199/HEK293 細胞の培養 上清とインキュベートしても分解 されなかった(結果省略).これら の結果から,KIAA1199 を介した HA 分解は,素早いベシクルエン ドサイトーシスとリサイクリング 経路を介し,HA 分解産物の細胞 内の蓄積や,リソソームにおける 分解は関与しないと考えられた. 2-3-4 KIAA1199 の細胞内局在制御における N 末端 30 アミノ酸の役割 KIAA1199 の N 末端 30 アミノ酸はシグナル配列と予想されている(Fig. 2-1)28). KIAA1199 の細胞内局在制御における N 末端 30 アミノ酸の機能を調べるため,C 末端に V5 タグを有する全長 KIAA1199(KIAA1199-V5)と,その N 末端 30 アミノ酸を欠失した KIAA1199-V5(KIAA1199(∆30aa)-V5)を,それぞれ HEK293 細胞で発現させた(Fig. 2-7A). 抗KIAA1199 抗体を用いたウェスタンブロット法の解析により,KIAA1199-V5 は 160 kDa 付近に検出されたのに対し,KIAA1199(∆30aa)-V5 はそれより 10 kDa 程度小さい位置に検 出された(Fig. 2-7B).同様の結果が,抗 V5 抗体を用いたウェスタンブロット法でも得ら れたことから,KIAA1199 の C 末端はその成熟過程でプロセシングを受けないと考えられ
Table 2-1. Distribution of [3H]HA in extracellular, cell surface-associated and intracellular
compartments after addition to KIAA1199/HEK293 and control HEK293 cells.
た(Fig. 2-7B).次に,KIAA1199-V5 および KIAA1199(∆30aa)-V5 発現細胞の HA 分解能 を調べたところ,KIAA1199-V5 発現細胞は HA を分解したのに対し,KIAA1199(∆30aa)-V5 発現細胞はHA を全く分解しなかった(Fig. 2-7C).また,KIAA1199-V5 発現細胞より単 離した KIAA1199-V5 の N 末端アミノ酸配列は T31VAAGCPDQS であり,成熟した KIAA1199 は,N 末端の 30 番目のセリンと 31 番目のスレオニンの間で切断されることが 示された(Fig. 2-7D).これらの結果から,HA 分解活性には全長KIAA1199 cDNA の発現 が必要であり,KIAA1199 タンパク質の成熟の過程において N 末端 30 アミノ酸は切断され ると考えられた.
また,KIAA1199 はN型糖鎖修飾されることが予想されているため,N-グリカナーゼお よびEndo H を用いて,KIAA1199-V5 および KIAA1199(∆30aa)-V5 が糖鎖修飾を受けて いるか検討した.脱糖鎖処理後のKIAA1199 の分子量変化を,抗 KIAA199 抗体を用いた ウェスタンブロット法により解析したところ,KIAA1199-V5 は分子量が低下したことから, 糖鎖修飾を受けていると考えられた.一方,KIAA1199(∆30aa)-V5 は分子量に変化が認め ら れ ず, 糖 鎖修 飾を 受け な いと 考 えら れた (Fig. 2-8A).次に免疫細胞染色にて,
Fig. 2-7. HA depolymerization by HEK293 cells expressing KIAA1199-V5 and
KIAA1199(∆30aa)-V5. (A) A schematic diagram showing constructs of KIAA1199-V5 and
KIAA1199-V5 および KIAA1199(∆30aa)-V5 の細胞内局在を調べたところ,KIAA1199-V5 は原形質膜付近のベシクル以外に,PDI(小胞体マーカー)(Fig. 2-8B)や GM130(ゴル ジマーカー)(Fig. 2-8C)と一部共局在していたのに対し,KIAA1199(∆30aa)-V5 はどちら のオルガネラマーカーとも共局在は認められず,サイトゾルに検出された(Fig.2-8B, C). 2-3-5 マウスKiaa1199 ホモログ遺伝子(mKiaa1199)の機能解析 mKiaa1199 はヒトKIAA1199とアミノ酸配列で91%の相同性を示し,高分子多糖の加 水分解に関与すると考えられている4 つの PbH1 ドメインは両者で完全に保存されている
Fig. 2-8. Deglycosylation and cellular localization of KIAA1199-V5 and
KIAA1199(∆30aa)-V5. (A) Deglycosylation of KIAA1199. The lysates of KIAA1199-V5
(Fig. 2-9)27, 28).mKiaa1199の全長cDNA を HEK293 細胞に導入した結果,ウェスタン ブロット法ではmKiaa1199 は KIAA1199 と同じ位置にバンドが認められ,mKiaa1199 発
mouse 1 : MRASGRHDVS LKIVLATGCL LLANFSGASS AVATECPDQS PELQPWSPGH NRDYQVHIGH human 1 : MGAAGRQDFL FKAMLTISWL TLTCFPGATS TVAAGCPDQS PELQPWNPGH DQDHHVHIGQ
mouse 61 : GRKLLLTSSA TVHSITISGG GKLVIKDHHE HIVLRTRYIL IDDGGELHAG SALCPFEGNF human 61 : GKTLLLTSSA TVYSIHISEG GKLVIKDHDE PIVLRTRHIL IDNGGELHAG SALCPFQGNF
mouse 121 : SIVLYGRADE NILPDPYYGL KYIGVDKGGT LELHGQKKLS WTFLNKTLHP GGMQEGGYFF human 121 : TIILYGRADE GIQPDPYYGL KYIGVGKGGA LELHGQKKLS WTFLNKTLHP GGMAEGGYFF
mouse 181 : ERSWGHRGVI VHVIDAKLGT VVHSDRFDTY RSKKESERLV QYLNAVPDGR ILSVAVNDEG human 181 : ERSWGHRGVI VHVIDPKSGT VIHSDRFDTY RSKKESERLV QYLNAVPDGR ILSVAVNDEG mouse 241 : SRNLDDTARK AMTKLGSKHF LHLGFRHPWS FITVKGNPSS SVEDHIEYHG HKGSAAARVF human 241 : SRNLDDMARK AMTKLGSKHF LHLGFRHPWS FLTVKGNPSS SVEDHIEYHG HRGSAAARVF
mouse 301 : KLFQTEHGEH FNVSSSSEWV QDVEWTEWFD HDKVPQSKGG EKISDLRAAY PGKICNRPID human 301 : KLFQTEHGEY FNVSLSSEWV QDVEWTEWFD HDKVSQTKGG EKISDLWKAH PGKICNRPID
mouse 361 : IQATTMDGVA LSTEVVYKNG QDYRFACYTR GRACRSYRVR FLCGKPVRPK LTVSIDTNVN human 361 : IQATTMDGVN LSTEVVYKKG QDYRFACYDR GRACRSYRVR FLCGKPVRPK LTVTIDTNVN
mouse 421 : STILSLVDNV RSWRPGDTLV VASTDYSMYQ AEEFRVLPCK ACTSTQVKVA GKPQYLHIGE human 421 : STILNLEDNV QSWKPGDTLV IASTDYSMYQ AEEFQVLPCR SCAPNQVKVA GKPMYLHIGE
mouse 481 : EIDGVDMRAE VGLLTRNIVV MGEMEDRCYP YTNHICDFFD FDTFGGHIKF ALGFKAAHLE human 481 : EIDGVDMRAE VGLLSRNIIV MGEMEDKCYP YRNHICNFFD FDTFGGHIKF ALGFKAAHLE
mouse 541 : GVELKYMGQQ LVGQYPIHFH LAGDLDEQGG YDPPTYIRDL SIHHTFSRCV TVHGSNGLLI human 541 : GTELKHMGQQ LVGQYPIHFH LAGDVDERGG YDPPTYIRDL SIHHTFSRCV TVHGSNGLLI
mouse 601 : KDVVGYNSLG HCFFTEDGPE ERNTFDHCLG LLVKSGTLLP SDRDSRMCKV ITEDSYPGYI human 601 : KDVVGYNSLG HCFFTEDGPE ERNTFDHCLG LLVKSGTLLP SDRDSKMCKM ITEDSYPGYI
mouse 661 : PKPRQDCNAV STFWMANPNN NLINCAAAGS EETGFWFIFH HVPTGPSVGM YSPGYSEHIP human 661 : PKPRQDCNAV STFWMANPNN NLINCAAAGS EETGFWFIFH HVPTGPSVGM YSPGYSEHIP
mouse 721 : LGKFYNNRAH SNYRAGMIID NGVKTTEASA KDKRPFLSII SARYSPHQDA DPLKPREPAI human 721 : LGKFYNNRAH SNYRAGMIID NGVKTTEASA KDKRPFLSII SARYSPHQDA DPLKPREPAI
mouse 781 : IRHFTAYKNQ DHGAWLRGGD VWLDSCRFAD NGIGLTLASG GTFPYDDGSK QEIKNSLFVG human 781 : IRHFIAYKNQ DHGAWLRGGD VWLDSCRFAD NGIGLTLASG GTFPYDDGSK QEIKNSLFVG
mouse 841 : ESGNVGTEMM DNRIWGPGGL DHSGRTLPIG QNFPIRGIQF YDGPINIQNC TFRKFAALEG human 841 : ESGNVGTEMM DNRIWGPGGL DHSGRTLPIG QNFPIRGIQL YDGPINIQNC TFRKFVALEG
mouse 901 : RHTSALAFRL NNAWQSCPHN NVTNIAFEDV PITSRVFFGE PGPWFNQLDM DGDKTSVFHD human 901 : RHTSALAFRL NNAWQSCPHN NVTGIAFEDV PITSRVFFGE PGPWFNQLDM DGDKTSVFHD
mouse 961 : LDGSVSEYPG SYLTKDDNWL VRHPDCINVP DWRGAICSGR YAQMYIQAYK SSNLRMKIIK human 961 : VDGSVSEYPG SYLTKNDNWL VRHPDCINVP DWRGAICSGC YAQMYIQAYK TSNLRMKIIK
mouse 1021 : NDFPSHPLYL EGALTRSTHY QQYQPVITLQ KGYTIHWDQT APAELAIWLI NFNKGDWIRV human 1021 : NDFPSHPLYL EGALTRSTHY QQYQPVVTLQ KGYTIHWDQT APAELAIWLI NFNKGDWIRV
mouse 1081 : GLCYPRGTTF SILSDVHNRL LKQTSKTGTF VRTLQMDKVE QSYPGRSHYY WDEDSGLLFL human 1081 : GLCYPRGTTF SILSDVHNRL LKQTSKTGVF VRTLQMDKVE QSYPGRSHYY WDEDSGLLFL
mouse 1141 : KLKAQNEREK FAFCSMKGCE RIKIKALLPR NAGISDCTAT AYPRFTERAI VDVPMPRKLF human 1141 : KLKAQNEREK FAFCSMKGCE RIKIKALIPK NAGVSDCTAT AYPKFTERAV VDVPMPKKLF
mouse 1201 : GAQLKTKDHF LEVKMESSRQ HFFHLRNDFA YIEVDGRRYP CSEDGIQIVV IDGSRGHVVS human 1201 : GSQLKTKDHF LEVKMESSKQ HFFHLWNDFA YIEVDGKKYP SSEDGIQVVV IDGNQGRVVS
mouse 1261 : HGSFRNAILQ GIPWQLFNYV AAIPDNSIVL MASKGRYITR GPWTRVLEKL GADKGLKLKE human 1261 : HTSFRNSILQ GIPWQLFNYV ATIPDNSIVL MASKGRYVSR GPWTRVLEKL GADRGLKLKE
mouse 1321 : KMAFVGFKGS FRPIWVTLET EDHKAKIFQV VPIPVVRKKK L human 1321 : QMAFVGFKGS FRPIWVTLDT EDHKAKIFQV VPIPVVKKKK L
Fig. 2-9. Sequence alignment of the mouse and human KIAA1199 proteins. Identical amino acids
現細胞は,ヒトKIAA1199 発現細胞と同様,[3H]HA を 10-100 kDa の中間サイズに分解し た(Fig. 2-10A).また,mKiaa1199 発現細胞により分解された HA の非還元末端糖はグ ルクロン酸(GlcA)だったことから(結果省略),HA の切断部位はβ-エンド-N-アセチルグ ルコサミン結合であると考えられた.
次に,mKiaa1199 安定発現細胞(mKiaa1199/HEK293 細胞)を樹立し,HA 分解にお けるmKiaa1199 の機能を調べた.まずは,mKiaa1199 タンパク質の GAG への結合活性 を調べたところ,mKiaa1199 は HA-H2(1,452 kDa),HA-M2(1,039 kDa),HA-L2(219 kDa),HA-S2(52 kDa),HA-T2(28 kDa)と結合し,他の GAG とは結合しないことが 示された(Fig. 2-10B, C).また,mKiaa1199/HEK293 細胞は培養系で,FA-HA(H1(1,760
Fig. 2-10 HA depolymerization by mKiaa1199 transfectants, HA-specific binding of recombinant mKiaa1199 protein, and peak size of the minimum degradates. (A) HEK293 cells
were transiently transfected with empty vector (Mock) or vector containing KIAA1199 or mKiaa1199 cDNA. The expression of KIAA1199 and mKiaa1199 proteins in each transfectant was assessed by immunoblotting using anti-KIAA1199 monoclonal antibody (Inset). Mock and mKiaa1199 transfectants were incubated with [3H]HA for 48 h, and HA depolymerization was then examined by size exclusion chromatography. GAPDH, a loading control. (B) Binding assay of mKiaa1199 protein to different GAGs. Lysates of mKiaa1199/HEK293 cells were incubated with H2O (negative control) or unlabeled GAGs including HA (HA-H2), CSA, CSC, CSD, DS, Hep and HS. The samples were precipitated with CPC, and analyzed by immunoblotting with anti-KIAA1199 antibody. (C) Binding of mKiaa1199 to HA species with different molecular sizes. The lysates were incubated with H2O (negative control) or HA with different sizes including HA-H2, HA-M2, HA-L2, HA-S2 and HA-T2, and then subjected to immunoblotting with anti-KIAA1199 antibody. (D) HPLC pattern of minimum degradates depolymerized from FA-HA H1 by mKiaa1199/HEK293 (chain line) and KIAA1199/HEK293 (dotted line) cells. The solid line indicates FA-HA H1 without incubation. (E) Peak sizes of minimum fragments from FA-HA H1 depolymerized by mKiaa1199/HEK293 and KIAA1199/HEK293 cells. Molecular weight of the fragments was determined using the following FA-HA species as a standard: FA-HA H1, M1, L1, S1, T1, U1 and 3K.
0 2 4 6 8 10 1 5 10 15 20 Fraction A Mo c k m K ia a 1 199 GAPDH KIAA1199 Mock mKiaa1199 cDNA [3H]HA x 1 0 3dpm /m l KI AA1 1 9 9 100 kDa 10 kDa > 1,000 kDa HA -H2 CS A CS C CS D DS H2 O He p HS Ly s a te B HA -H2 HA -M2 HA -L2 HA -S2 HA -T2 H2 O Ly s a te C D E 3 4 5 6 7 5 10 15 20 25 Log m ol ec ul ar w ei g ht
Elution time (min) mKiaa1199 (M1) (H1) (S1) (L1) (T1) (U1) (3K) KIAA1199
Elution time (min)
kDa),M1(907 kDa),L1(197 kDa),S1(56 kDa),T1(28 kDa),U1(9.8 kDa)) を分解したが,他のFA-GAG(FA-CSA,FA-CSC,FA-CSD,FA-DS,FA-Hep,FA-HS) は分解しなかった(結果省略).これらの結果は,ヒト KIAA1199 の結果とよく一致した. 一方,mKiaa1199/HEK293 細胞の長期培養で得られる最小 HA 分解産物は約 4.1 kDa だっ たのに対し,ヒトKIAA1199/HEK293 細胞の長期培養で得られる最小 HA 分解産物は約 3.3 kDa だった(Fig. 2-10D, E).また,mKiaa1199/HEK293 細胞で得られた HA 分解産物の 分子量プロファイルには,約4.1 kDa のメインのピークに加え,2 kDa 付近にも小さなピ ークが認められた(Fig. 2-10E).
次に,mKiaa1199 を介した HA 分解への,クラスリンおよびカベオラ経路の関与を調べ た.mKiaa1199/HEK293 細胞において CHC および AP-2 の発現を siRNA により抑制した ところ,HA 分解は顕著に低下した(Fig. 2-11A, B).一方,カベオリン-1 の発現抑制は HA 分解に影響を与えなかった(Fig. 2-11C).また,mKiaa1199/HEK293 細胞を用いた mKiaa1199 と CHC の二重染色では,両分子のシグナルは原形質膜付近のベシクルに認め
Fig. 2-11 Clathrin-specific HA depolymerization and cellular localization of mKiaa1199 in mKiaa1199/HEK293 cells. (A-C) Changes in HA depolymerization in mKiaa1199/HEK293 cells by
カルはHA をランダムに分解し,種々の大きさの HA を産生することが知られているが34), KIAA1199 を介した HA 分解に,フリーラジカルの関与も含めた検討も必要である.
KIAA1199 の N 末端 30 アミノ酸はシグナル配列と予想されていることから 28), KIAA1199 の細胞内局在制御における N 末端 30 アミノ酸の役割について調べた.まずは全 長 KIAA1199(KIAA1199-V5)あるいは予め N 末端 30 アミノ酸を欠失した KIAA1199 (KIAA1199(∆30aa)-V5)を HEK293 細胞で発現させたところ,KIAA1199(∆30aa)-V5 発 現細胞はHA を全く分解せず,HA 分解活性には全長 KIAA1199 の発現が必要であった. 一方,機能的に成熟したKIAA1199 は,N 末端 30 番目のセリンと 31 番目のスレオニンの 間 で 切 断 さ れ て い た . 次 に , 脱 糖 鎖 後 の 分 子 量 変 化 に て KIAA1199-V5 と KIAA1199(∆30aa)-V5 のN 型糖鎖修飾の有無を検討した結果,KIAA1199-V5 は修飾を受 けるが,KIAA1199(∆30aa)-V5 は修飾を受けないと考えられた.N 型糖鎖は小胞体にて付 与されることから,KIAA1199(∆30aa)-V5 は正常な小胞体への移行,あるいは小胞体での 正常な折り畳みがなされず,小胞 体以降の輸送も正常に行われな いことが推測された.そこで,免 疫細胞染色にてKIAA1199-V5 お よび KIAA1199(∆30aa)-V5 の細 胞 内 局 在 を 調 べ た 結 果 , KIAA1199-V5 は原形質膜付近の ベシクルに加え,小胞体やゴルジ 体にも一部局在していたのに対 し,KIAA1199(∆30aa)-V5 は,正 常な細胞内局在性を失い,サイト ゾルに認められた.これらの結果から,KIAA1199 の N 末端 30 アミノ酸はシグナル配列と して機能し,KIAA1199 は翻訳時に小胞体に移行後,N 末端 30 アミノ酸の切断とN型糖 鎖修飾を受け,ゴルジ体を経由し,成熟したタンパク質として HA 分解に必須な原形質膜 近傍のベシクルへ輸送されると考えられた(Scheme 3).
3-2-4 RNA 干渉法およびウェスタン解析法 第1章で記載した方法に準じて実施した. 3-2-5 関節滑膜試料
関節滑膜組織はOA(n=10)および RA(n=8)患者より膝関節形成術時に採取した.非 炎症性関節疾患滑膜組織(n=3)は前十字靭帯の再建手術から約1 年後の関節鏡検査時に採 取した.OA と RA は The American College of Rheumatology35, 36)の基準に従い診断され た.滑膜組織の研究用途としての利用に関し,慶應義塾大学病院および㈱カネボウ化粧品 の倫理審査に従い,患者よりインフォームドコンセントを得た.
3-2-6 免疫組織染色法
KIAA1199 の検出にはホルマリン固定した RA 患者由来滑膜組織のパラフィン切片を用 いて実施した.発色はVector® DAB(茶色; Vector Laboratories 社製)で行い,その他は 第1章で記載した方法に準じて実施した.
3-2-7 in situハイブリダイゼーション法
KIAA1199 の検出にはホルマリン固定した RA 患者由来滑膜組織のパラフィン切片を用 い,第1章で記載した方法に準じて実施した.
3-2-8 ミスセンス突然変異型KIAA1199遺伝子発現プラスミドの構築
4 種類のミスセンス突然変異型KIAA1199(R187C,R187H,H783R,V1109I)24)のcDNA は Takara Bio 社にて化学合成した.発現プラスミドは,これらの cDNA をそれぞれ pcDNA3.1(-)プラスミド(Invitrogen 社製)に挿入して構築した.組換え DNA 実験は㈱カ ネボウ化粧品の倫理審査で承認された.
3-2-9 統計処理
統計解析はSAS システムにより実施した.Dunnett’s multiple comparison test でコン トロールとの有意性を解析した.P値0.05 未満を統計的に有意とした.
第3節:実験結果
3-3-1 OA および RA 患者由来の滑膜細胞と滑膜組織における KIAA1199 の発現亢 進
人由来細胞に比べ,OA および RA 患者由来細胞の HA 分解活性は高く,それら活性は
Fig. 3-1. KIAA1199-mediated HA degradation in OA and RA synovial fibroblasts, and expression of KIAA1199 by synovial lining and sublining cells in the RA patients. (A)
KIAA1199 mRNA およびタンパク質の発現レベルとよく一致した(Fig. 3-1B).また, siRNA による KIAA1199 の発現抑制により,いずれの細胞の HA 分解活性もほぼ完全に消 失した(Fig. 3-1A).これらのことから,KIAA1199 は滑膜細胞の HA 分解においても必須 の因子であり,その発現亢進がOA および RA 細胞における HA 分解活性の亢進に寄与し ていることが考えられた. 次に,リアルタイムPCR 解析により,KIAA1199遺伝子の発現は非炎症性関節疾患滑膜 組織に比べ,OA 滑膜組織で上昇傾向にあり(P=0.088),RA 滑膜組織で有意に上昇(P<0.05) していることを示した(Fig. 3-1C).さらに,in situハイブリダイゼーション法および免疫 組織染色法により,KIAA1199 は RA 滑膜組織の主に lining 細胞といくつかの sublining 細胞に発現していることを見出し,そのシグナルはネガティブコントロールであるセンス プローブやnon-immune IgG では検出されないことを確認した(Fig. 3-1D, E).
3-3-2 難聴患者で見出されたKIAA1199 のミスセンス突然変異が HA 分解活性に与 える影響
KIAA1199 において難聴との関連性が指摘されている 4 種類のミスセンス突然変異24)が HA 分解活性に与える影響を調べた.4 種の変異型KIAA1199(R187C,R187H,H783R, V1109I)と野生型KIAA1199 cDNA を作製し,HEK293 細胞で一過的に発現させた.その 結果,いずれの変異型タンパク質も野生型タンパク質と同等のレベルで発現が認められた (Fig.3-2).一方,R187C および R187H 変異型を発現する細胞の HA 分解は,野生型, H783RおよびV1109I変異型を発現する細胞のHA 分解より著しく低下していた(Fig.3-2).
第4 節:考察
今回の研究において,siRNA 依存的な KIAA1199 の発現抑制により,健常人,OA およ びRA 患者由来のいずれの滑膜細胞の HA 分解もほぼ完全に消失することを見出した.こ
Fig. 3-2. KIAA1199-dependent depolymerization by cells expressing wild-type or mutant KIAA1199 proteins. HEK293 cells were transfected with vectors containing
総括 ヒアルロン酸(HA)は脊椎動物組織の細胞外マトリックスの主要な構成成分として広く 存在するグリコサミノグリカンである.皮膚には生体HA の約 50%が存在し,その代謝半 減期は1-1.5 日と言われている.一方,関節炎やガンなどの病態においては HA の分解が亢 進し,病態との関連性が報告されているが,HA 分解機構については未だ不明な点が多いの が現状である. 本申請論文では,生理的状態あるいは病態における組織の HA 分解機構を明らかにする ことを主眼として検討を進めた. 第1章では,正常ヒト皮膚線維芽細胞の HA 分解機構の解明に取り組んだ.皮膚線維芽 細胞は,培養系において外的に添加した HA を旺盛に分解した.しかし,その活性は細胞 溶出液では完全に消失することから,これまで HA 分解分解を担う分子の同定は困難であ った.一方,ヒトには6 つの HA 分解酵素(様)遺伝子が見出され,組織の HA 分解機構 として,CD44(HA 受容体)と 2 種類の HA 分解酵素(HYAL1 および HYAL2)が共同的 に働くというモデルが提唱された.しかしながら,皮膚線維芽細胞においてHYAL1遺伝子 の発現は認められず,またsiRNA による CD44 や HYAL2 の発現抑制も HA 分解に影響を 与えなかったことから,皮膚線維芽細胞にはこれらの分子が関与しない,新規な HA 分解 機構が存在すると考えられた.そこで,HA 分解に関わる未知の分子を同定すべく,まずは マイクロアレイ解析により,線維芽細胞の培養系 HA 分解活性と,その発現の挙動が一致 する遺伝子を包括的に抽出した.次に,得られた遺伝子について,siRNA 依存的に発現抑 制を行い,機能不明の難聴遺伝子として報告されていたKIAA1199 の発現抑制が,皮膚線 維芽細胞のHA 分解を顕著に抑制させることを見出した.さらに,培養系で HA を分解し ないHEK(human embryonic kidney)293 細胞にKIAA1199 cDNA を導入したところ, 皮膚線維芽細胞と同様のHA 分解能を新たに獲得したことから,KIAA1199 こそが HA 分 解を担う新規な分子であると考えられた.さらに,KIAA1199 は正常ヒト皮膚真皮の線維 芽細胞で発現していたことから,皮膚の生理的なHA 分解に寄与していると考えられた.
謝辞
掲載論文
本研究の内容は以下の論文に発表した.
第1章,第2章,第3章
Yoshida H., Nagaoka A., Kusaka-Kikushima A., Tobiishi M., Kawabata K., Sayo T., Sakai S., Sugiyama Y., Enomoto H., Okada Y., Inoue S., KIAA1199, a deafness gene of unknown function, is a new hyaluronan binding protein involved in hyaluronan depolymerization., Proc. Natl. Acad. Sci. USA, 110, 5612-5617 (2013).
第2章
Yoshida H., Nagaoka A., Nakamura S., Tobiishi M., Sugiyama Y., Inoue S., N-terminal signal sequence is required for cellular trafficking and hyaluronan-depolymerization of KIAA1199., FEBS Lett., 588, 111-116 (2014).
第2章
引用文献
1. Fraser J.R., Laurent T.C., Laurent U.B., J. Intern. Med., 242, 27-33 (1997).
2. Pandey M.S., Harris E.N., Weigel J.A., Weigel P.H., J. Biol. Chem., 283, 21453-21461 (2008).
3. Ghosh P., Clin. Exp. Rheumatol. 12, 75-82 (1994).
4. Yoshida M., Sai S., Marumo K., Tanaka T., Itano N., Kimata K., Fujii K., Arthritis Res.
Ther., 6, R514-520 (2004).
5. Sugahara K.N., Murai T., Nishinakamura H., Kawashima H., Saya H., Miyasaka M., J. Biol.
Chem., 278, 32259-32265 (2003).
6. Vuorio E., Einola S., Hakkarainen S., Penttinen R., Rheumatol. Int., 2, 97-102 (1982). 7. Rooney P., Kumar S., Ponting J., Wang M., Int. J. Cancer, 60, 632-636 (1995). 8. West D.C., Hampson I.N., Arnold F., Kumar S., Science, 228, 1324-1326 (1985). 9. Csoka A.B., Scherer S.W., Stern R., Genomics, 60, 356-361 (1999).
10. Harada H., Takahashi M., J. Biol. Chem., 282, 5597-5607 (2007).
11. Bourguignon L.Y., Singleton P.A., Diedrich F., Stern R., Gilad E., J. Biol. Chem., 279, 26991-27007 (2004).
12. Csoka A.B., Frost G.I., Stern R. Matrix Biol., 20, 499-508 (2001).
13. Triggs-Raine B., Salo T.J., Zhang H., Wicklow B.A., Natowicz M.R., Proc. Natl. Acad. Sci.
U S A, 96, 6296-6300 (1999).
14. Rai S.K., Duh F.M., Vigdorovich V., Danilkovitch-Miagkova A., Lerman M.I., Miller A.D.,
Proc. Natl. Acad. Sci. U S A, 98, 4443-4448 (2001).
15. Martin D.C., Atmuri V., Hemming R.J., Farley J., Mort J.S., Byers S., Hombach-Klonisch S., Csoka A.B., Stern R., Triggs-Raine B.L., Hum. Mol. Genet., 17, 1904-1915 (2008).
16. Jadin L., Wu X., Ding H., Frost G.I., Onclinx C., Triggs-Raine B., Flamion B., FASEB J., 22, 4316-4326 (2008).
17. Nakamura T., Takagaki K., Kubo K., Morikawa A., Tamura S., Endo M., Biochem. Biophys.
Res. Commun., 172, 70-76 (1990).
18. Underhill C.B., Toole B.P., J. Cell Biol., 82, 475-484 (1979).
19. Sugiyama Y., Shimada A., Sayo T., Sakai S., Inoue S., J. Invest. Dermatol., 110, 116-121 (1998).
20. Michishita E., Garces G., Barrett J.C., Horikawa I., Cancer Lett., 239, 71-77 (2006).
21. Averbeck M., Gebhardt C.A., Voigt S., Beilharz S., Anderegg U., Termeer C.C., Sleeman J.P., Simon J.C., J. Invest. Dermatol., 127, 687-697 (2007).
23. Li L., Asteriou T., Bernert B., Heldin C.H., Heldin P., Biochem. J., 404, 327-336 (2007). 24. Abe S., Usami S., Nakamura Y., J. Hum. Genet., 48, 564-570 (2003).
25. Kohda D., Morton C.J., Parkar A.A., Hatanaka H., Inagaki F.M., Campbell I.D., Day A.J.,
Cell, 86, 767-775 (1996).
26. Yang B., Yang B.L., Savani R.C., Turley E.A., EMBO J., 13, 286-296 (1994).
27. He Q.Y., Liu X.H., Li Q., Studholme D.J., Li X.W., Liang S.P., Bioinformatics, 22, 2189-2191 (2006).
28. Birkenkamp-Demtroder K., Maghnouj A., Mansilla F., Thorsen K., Andersen C.L., Øster B., Hahn S., Ørntoft T.F., Br. J. Cancer, 105, 552-561 (2011).
29. Guo J., Cheng H., Zhao S., Yu L., FEBS Lett., 580, 581-584 (2006). 30. Armstrong S.E., Bell D.R., Anal. Biochem., 308, 255-264 (2002).
31. Reissig J.L., Storminger J.L., Leloir L.F., J. Biol. Chem., 217, 959-966 (1955). 32. Takemoto H., Hase S., Ikenaka T., Anal. Biochem., 145, 245-250 (1985). 33. Hase S., Tsuji Y., Matsushima Y., J. Biochem., 72, 1549-1555 (1972).
34. Stern R., Kogan G., Jedrzejas M.J., Soltes L., Biotechnol. Adv., 25, 537-557 (2007). 35. Kuscu C, Evensen N, Kim D, Hu YJ, Zucker S, Cao J., PLoS One, 7, e44661. (2012). 36. Frewin D.B., Cleland L.G., Jonsson J.R., Robertson P.W., J. Rheumatol., 13, 13-14 (1986). 37. Anniko M., Arnold W., ORL. J. Otorhinolaryngol. Relat. Spec., 57, 82-86 (1995).
あったからことから,HA はエンド-β-N-アセチルグルコサミニダーゼ様式で分解されると 考えられた.次に HA 分解に関わるエンドサイトーシス経路の同定として,クラスリン経 路に関わるクラスリン重鎖(CHC)および AP-2 を siRNA により発現抑制したところ, KIAA1199/HEK293 細胞の HA 分解は顕著に低下した.しかし,カベオラ経路に関わるカ ベオリン-1 の発現抑制は HA 分解に全く 影響を与えなかった.さらに,細胞に添加 したビオチン標識 HA は,共焦点顕微鏡 観察により原形質膜付近のベシクルにシ グナルが認められた。これらのことから, 外的に添加した HA はクラスリン経路を 介し,一旦細胞内に取り込まれると考えら れた.次にHA 分解の場の同定を目的とし, 阻害剤を用いた解析では,Dynasore(エ ンドサイトーシスおよびベシクルの形成阻害),NH4Cl(エンドソーム-リソソーム内のアル カリ化),バフィロマイシンA1(液胞型プロトン ATPase 活性阻害),モネンシン(受容体 リサイクリング阻害)はKIAA1199/HEK293 細胞の HA 分解を低下させたが,ノコダゾー ル(初期エンドソームから後期エンドソームへの輸送阻害)はそれには影響を及ぼさなか った(Fig. 3).さらに,外的に添加した[3H]HA はそのほとんどが培養上清に回収された. 以上の結果から,細胞内に取り込まれた HA はエンドソームとリソソームの融合前の酸性 区画,すなわちクラスリン被覆小胞もしくは初期エンドソームで分解された後,リサイク ル経路を介して素早く細胞外に放出されると考えられた.また,KIAA1199 と CHC の二重 抗体染色では,両分子のシグナルはKIAA1199/HEK293 細胞の原形質膜付近にベシクル様 で観察された.今後,詳細な検討が必要であるが,HA 分解における KIAA1199 の機能と して,原形質膜近傍のベシクル生じる HA 分解に関わる可能性が推察された.また,全長 およびN 末端 30 アミノ酸を欠失したKIAA1199のcDNA を用いた解析により,KIAA1199 のN 末端 30 アミノ酸はシグナル配列として機能し,KIAA1199 の適切な細胞内輸送と培養 系HA 分解活性に必須であることを明らかにした.
と OA および RA 患者由来培養滑膜細胞の HA 分解活性を調べた.OA および RA 患者 由来細胞のHA 分解活性は健常人由来細胞に 比べて高く,加えてKIAA1199 mRNA およ びタンパク質の発現も上昇していた.また, siRNA による KIAA1199 の発現抑制は,い ずれの細胞のHA 分解活性もほぼ完全に消失 させた.次に,in situハイブリダイゼーショ ン 法 お よ び 免 疫 組 織 染 色 法 に よ り , KIAA1199 は RA 滑膜組織の主に lining 細胞 に発現していることを示した.さらにリアル タイムPCR により,KIAA1199遺伝子の発現は非炎症性関節疾患滑膜組織に比べ,OA お よびRA 滑膜組織で上昇していることを見出した(Fig. 4).これらの結果から,KIAA1199 は滑膜細胞のHA 分解においても必須の因子であり,滑膜組織での KIAA1199 の発現亢進 が,関節炎患者の滑膜における過剰なHA 分解に寄与している可能性が考えられた. KIAA1199 は内耳で高発現し,複数の非症候群性難聴家系にアミノ酸置換を伴う突然変 異(ミスセンス突然変異)が報告されていることから,これまで難聴への関わりが考えら れてきた.そこで本研究では、これらミスセンス突然変異が HA 分解活性に与える影響を 調べた.4 種類の変異型KIAA1199(R187C,R187H,H783R および V1109I)と野生型 KIAA1199 の cDNA をそれぞれ HEK293 細胞で一過的に発現させた結果,R187C および R187H 変異型を発現する細胞の HA 分解は,他の細胞に比べ著しく低下していることを見 出した.そのため,HA 分解の低下を伴う KIAA1199 の機能変化が,聴力の低下に何らか の影響を及ぼしていることが推察された. 今回の研究により,KIAA1199 は正常な皮膚真皮の生理的な HA 分解だけでなく,関節 炎滑膜における過剰な HA 分解においても重要な役割を担う新規な分子と考えられた.ま た,KIAA1199 の機能変化が,聴力の低下と何らかの関係がある可能性を示した.近年, 胃ガンや大腸ガンでの高発現も報告されており,今後,KIAA1199 をターゲットとした治 療薬の確立は有用であると考えられた. 本研究結果の掲載
1) Proc. Natl. Acad. Sci. U S A., 110, 5612-5617 (2013). 2) FEBS Lett., 588, 111-116 (2014).




![Table 2-1. Distribution of [ 3 H]HA in extracellular, cell surface-associated and intracellular compartments after addition to KIAA1199/HEK293 and control HEK293 cells.](https://thumb-ap.123doks.com/thumbv2/123deta/10126476.1960428/31.892.399.728.373.567/distribution-extracellular-surface-associated-intracellular-compartments-addition-control.webp)