Efficient reduction of Cys110 thiyl radical by glutathione in human myoglobin
34
0
0
全文
(2) Abstract Human myoglobin (hMb) possesses a cysteine (Cys) residue which is rare among mammalian Mbs. To investigate the effects of this unique Cys residue at the amino acid position 110 (Cys110) on hMb reactions, we studied the reactions of wild type (WT) methMb and its alanine mutant at Cys110 (C110A) with H2O2, particularly in the presence of reduced glutathione (GSH) which is well known as a reducing agent. The formation rates of the ferryloxo (Fe(IV)=O) species by H2O2 under air were about the same for WT and C110A methMbs, whereas the protein decomposed more in the case of WT than C110A hMb. With the addition of GSH, hMb consumed H2O2 faster and decomposition of the protein decreased, where the effects were more prominent in WT than C110A hMb. The radicals produced by the reaction with H2O2 decreased significantly due to the addition of 1 mM GSH in the case of WT hMb, but not in the case of C110A hMb. These results show that GSH reduces H2O2–induced protein decomposition due to reduction of the C110-thiyl radical in WT hMb by electron transfer.. Keywords: human myoglobin; hydrogen peroxide; thiyl radical; glutathione; electron transfer.. Abbreviations Cys. cysteine. Mb. myoglobin. WT. wild type. GSH. reduced glutathione. GSSG. glutathione disulfide. DMPO. 5,5-dimethyl-1-pyrroline N-oxide. 2.
(3) TEMPO. 2,2,6,6-tetramethylpiperidine-N-oxyl. MALDI-TOF. matrix-assisted laser desorption/ionization-time-of-flight. 3.
(4) 1.. Introduction Mb is an oxygen carrier hemoprotein, consisting of eight α-helices and seven nonhelical. segments constructed of 153 amino acids [1]. The heme is coordinated with the nitrogen atom of the histidine residue, and the oxygen binds to the heme iron at the opposite position of the histidine. Human Mb (hMb) possesses a unique cysteine at position 110 (Cys110), while most other mammalian Mbs do not [2]. Ligand binding has been studied extensively in Mb [3], and the effects of the Cys residue in hMb on the reactions with small molecules, such as NO and CO, have been investigated [4-6]. The reaction of the Cys residue in hMb with NO yields S-nitroso hMb, which is proposed to be a bio-active NO pool [4]. We have recently shown that the heme in Cys-introduced mutant sperm whale Mb can be reduced by electron transfer from the Cys residue to the heme, where the Cys thiyl radical was produced under CO atmosphere [5]. The reaction of Mb with H2O2 has also been investigated extensively [6-13]. The ferryloxo (Fe(IV)=O) species is generated by the reaction of metMb with H2O2, where an electron is provided to the heme iron from the porphyrin or nearby amino acid and thus a porphyrin π-cation radical or amino acid radical is produced, respectively. The ferryloxo species with and without the π-cation is referred to as compound I and compound II, respectively [7-10]. Locations of the amino acid radicals in compound II of sperm whale and human Mbs have been investigated with EPR using the spin-trapping technique [14-16]. In the reaction with H2O2, sperm whale Mb forms a dimer through the covalent bond between Tyr103 of one Mb and Tyr151 of another Mb, while horse Mb does not due to the lack of Tyr151 [17]. For the reaction of hMb with H2O2, the Cys thiyl radical is generated by electron transfer from Cys110 to the Tyr103 radical, resulting in the formation of the Mb dimer through the disulfide bond [16]. Recently, it has been shown that the thiyl radical produced by the reaction with H2O2 in hMb oxidizes dopamine to yield a reactive quinone. 4.
(5) species and this quinone species reacts with another thiyl radical to produce a cysteinyl-dopamine adduct [6]. Amino acid radicals are reactive and are usually quenched by antioxidants and enzymes, such as superoxide dismutase, glutathione peroxidase, and catalase. Reduced glutathione (GSH) serves as a radical scavenger [18], eliminating reactive oxygen species [19, 20] and amino acid radicals [14, 21]. In fact, rat liver contains about 10 mM GSH in its cells [22]. GSH reduces the Tyr radical, produced by the reaction of horseradish peroxidase with H2O2 [21]. GSH also reduces the peroxyl radical of the amino acid residue produced by the reaction of horse Mb with H2O2 [14]. When GSH reduces the radical, a glutathione thiyl radical is produced and the produced thiyl radical reacts with the thiolate anion of another GSH, thus forming a disulfide anion radical [21]. This anion radical produces a superoxide and a glutathione disulfide in the presence of O2 [21]. However, the effect of GSH on the reaction of hMb with H2O2 has not been investigated in detail. To investigate the effects of the Cys residue on hMb reactions in the presence of GSH, we compared the reactions of WT and C110A hMbs with H2O2. We found that GSH protects methMb from decomposition by reduction of the Cys thiyl radical.. 2.. Experimental. 2.1. Material and methods Enzymes for site-directed mutagenesis were obtained from Takara Shuzo Co. (Kyoto, Japan). Oligonucleotide primers were purchased from Sigma-Aldrich Co. (Tokyo, Japan). The cDNA of hMb was purchased from Toyobo Co. (Osaka, Japan). Using the cDNA of hMb as a template, the DNA segment was amplified by PCR with hMbNdeI-F and hMbBamHI-R primers (Table 1) [23]. The PCR product was purified and digested with NdeI and BamHI. The digested DNA segment was inserted into the NdeI-BamHI site of the pET29b expression. 5.
(6) vector. Amino acid substitution of hMb at the Cys110 position was introduced by PCR-based in vitro mutagenesis of the pET29b expression vector using hMbC110A-F and hMbC110A-R primers (Table 1). Plasmid DNAs were prepared using the QIAprep spin Mini prep kit (QIAGEN, Tokyo, Japan). DNA sequencing was carried out with the BigDye Terminator v1.1 cycle sequencing kit (Applied Biosystems, Inc., Foster City, CA) and an ABI PRISM 310 genetic analyzer sequencing system (Applied Biosystems, Inc., Foster City, CA). The obtained plasmids were introduced into competent cells of E. coli BL21(DE3) . Recombinant WT and C110A hMbs were overproduced in E.coli BL21(DE3) cells. The cells were grown at 37oC in 5 L flasks containing 2 L of LB broth (Sigma-Aldrich Co., Tokyo, Japan) (20 g/L) until the OD600 value became about 0.5. The expression of recombinant hMbs was induced by addition of isopropyl β-D-1-thiogalactopyranoside (WAKO, Tokyo, Japan) (0.25 mM, final concentration). Cultures were incubated subsequently for 12 h at 20oC and harvested. MethMb was purified using the previous method [5]. The purity of WT and C110A hMb was confirmed by the SDS-PAGE analyses where no dimer of these proteins was detected, and the ratio of the absorbances (Abs) at 280 and 409 nm (Abs409/Abs280 > 4.1). The concentrations of the proteins were calculated from the absorbance at 409 nm and adjusted to desired concentrations [24]. Diluted H2O2 solution was prepared from 30% H2O2 solution (WAKO, Tokyo, Japan). The concentration of H2O2 was calculated from the absorbance at 230 nm (ε230nm = 62 M-1cm-1) [25].. 2.2. Optical absorption measurements The optical absorption spectra of methMb were recorded on a UV-2450 spectrophotometer (Shimadzu Co., Kyoto, Japan) with a quartz cell at 20oC. MethMb (2 μM) was prepared in 20 mM potassium phosphate buffer, pH 7.0, both with and without GSH (1 mM). To obtain N2 atmosphere, the cell was degassed and refilled with N2 using a vacuum. 6.
(7) line. The reaction solution was mixed after the addition of H2O2 (20-120 μM), then all measurements were started within 30 s after the mixture.. 2.3. EPR measurements The EPR spectra were obtained with a JES-FA100N spectrometer (JEOL Ltd., Tokyo, Japan) at room temperature or 77 K. The spin-trap reagent, 5,5-dimethyl-1-pyrroline N-oxide (DMPO),. was. purchased. from. Labotech. (Tokyo,. Japan).. 2,2,6,6-Tetramethyl-piperidin-1-oxyl (TEMPO) was obtained from Sigma-Aldrich Co. (Tokyo, Japan). WT and C110A methMb (500 μM) solutions in 20 mM potassium phosphate buffer, pH 7.0, were degassed. The DMPO solution (5 mM, final concentration) was added anaerobically to the Mb solution. For measurements with 1 mM GSH, the GSH solution was degassed separately and added to the sample solution in a vial under N2 atmosphere. The mixed solution (250 μL, final volume) was incubated for 2 min after the addition of H2O2 (2.5 mM, final concentration). After incubation, 200 μL of the solution was transferred into a flat EPR quartz cell (JEOL DATUM, Tokyo, Japan) and the spectra were recorded immediately at room temperature. For measurements at 77 K, 50 μL of the mixed solution was transferred into a 5 mm EPR quartz cell (JEOL DATUM, Tokyo, Japan) under N2 atmosphere. The sample tube was transferred into a fingertip liquid N2 Dewar flask (JEOL Ltd., Tokyo, Japan) filled with liquid N2. The EPR spectrometer settings were as follows: Microwave frequency, 9.41 GHz at room temperature or 9.05 GHz at 77 K; microwave power, 5 mW. Each spectrum was an average of 4 scans. The signal intensity of each spectrum was normalized with that of the Mn marker spectrum. The peak areas of the signals were calculated by the double-integration of the spectra. The radical concentrations were estimated by comparison of the calculated peak areas with that obtained for 100 μM TEMPO.. 7.
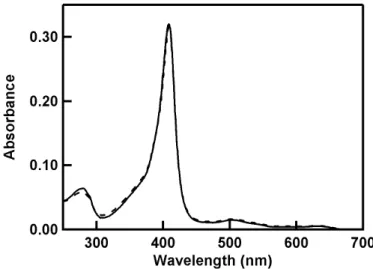
(8) 2.4 Mass spectrometry analysis Matrix-assisted laser desorption/ionization-time-of-flight (MALDI-TOF) mass spectra of hMbs were recorded on an autoflex II instrument (Bruker Daltonics, Bremen, Germany) with 3,5-dimethoxy-4-hydroxycinnamic acid (WAKO, Tokyo, Japan) as a matrix. WT methMb was dissolved in 50 mM potassium phosphate buffer, pH 7.0. The WT methMb (100 μM, final concentration) solution was mixed with the GSH (100 μM or 50 mM, final concentration) solution. H2O2 (500 μM, final concentration) was added to the mixed solution (100 μL, final volume), and the solution was incubated for 10 min at 20oC. After incubation, gel filtration chromatography was performed for the protein solution with a PD-10 column (GE Healthcare UK Ltd., Amersham Place, United Kingdom). The eluted protein solution was concentrated with VIVASPIN 500 (Sartorius Stedim Biotech S.A., Aubagne Cedex, France). The concentrated protein solution was mixed with the matrix and placed on the mass plate. All mass spectra were calibrated with Protein Calibration Standard II (Bruker Daltonics, Bremen, Germany).. 3. Results 3.1. Formation of the Fe(IV)=O species in hMb by reaction with H2O2 The Soret band of C110A methMb was detected at 409 nm, similar to that of WT hMb, which showed that the structure around the heme in hMb was not perturbed significantly by the removal of the Cys residue (Fig. S1). The addition of 20 μM H2O2 to methMb under air caused decreases in the absorbances at 409, 505, and 632 nm (Abs409, Abs505, and Abs632) and increases at 421, 549, and 589 nm (Abs421, Abs549, and Abs589) (Fig. 1). These new absorption bands are attributed to the ferryloxo (Fe(IV)=O) species [8, 9]. When the Fe(IV)=O species is produced in methMb by H2O2, the porphyrin π-cation radical is generated concomitantly [8-10, 12] and amino acid radicals are formed by intramolecular. 8.
(9) electron transfer from the amino acid residue to the porphyrin π-cation radical [11]. The pseudo-first-order rate constants (kobs(Fe(IV))) of Fe(IV)=O formation for WT and C110A methMbs under air were obtained by fitting the absorbance changes at 421 nm with single-exponential functions. The obtained kobs(Fe(IV)) values exhibited a linear dependence on H2O2 concentration (Fig. 1, inset), and the apparent rate constant (k1) was obtained from equation 1.. kobs(Fe(IV)) = k1[H2O2]. (1). The k1 values of WT and C110A hMbs were 510 and 430 M-1s-1, respectively. The k1 values of WT and C110A hMbs were similar, but both were slightly higher than the values in the previous report (310 and 350 M-1s-1 for WT and C110A hMbs, respectively) [26]. We used low H2O2 concentrations (20-120 μM) to avoid decomposition of Mb by H2O2, and the difference in the k1 values may be due to protein decomposition in the previous experiments, which were performed at high H2O2 concentration (~1000 μM). The similar k1 values, however, between WT and C110A hMbs indicate that Cys110 does not have a significant effect on formation of the Fe(IV)=O species in hMb, which is in agreement with the previous report [26]. To investigate the effect of O2 on formation of the Fe(IV)=O species by H2O2, WT and C110A hMbs were mixed with H2O2 under N2 atmosphere. The kobs(Fe(IV)) values on formation of the Fe(IV)=O species for WT and C110A hMbs did not change significantly in the absence of O2 (Fig. S2). These results show that O2 does not affect formation of the Fe(IV)=O species. WT and C110A hMbs were mixed with H2O2 under air in the presence of 1 mM GSH to investigate the effects of GSH on formation of the Fe(IV)=O species. The kobs(Fe(IV)) values of. 9.
(10) Fe(IV)=O formation for WT and C110A hMbs were not affected by the addition of GSH (Fig. S3). These results demonstrate that GSH also does not affect formation of the Fe(IV)=O species.. 3.2. Decomposition of hMb by the reaction with H2O2 The intensities of the Soret and visible bands of the heme of hMb decreased gradually during the reaction with H2O2 under air (Fig. 2), which is attributed to slow protein decomposition by the amino acid radicals produced [27, 28]. The pseudo-first-order rate constant for decomposition (kobs(dec)) of hMb by H2O2 was estimated from the initial change in absorbance at 421 nm after formation of the Fe(IV)=O species. The kobs(dec) values of WT hMb were about twice as large as those of C110A hMb for all the H2O2 concentrations investigated (Fig. 2, inset). For example, kobs(dec) for WT and C110A hMbs by 20 μM H2O2 were 3.6 x 107 and 1.6 x 107 s-1, respectively. These results show that H2O2-induced protein decomposition of WT hMb was faster than that of C110A hMb under air. A decrease at about 421 nm and an increase at about 409 nm were observed in the absorption spectra of both WT and C110A hMbs after consumption of H2O2 under air, which suggested conversion of the Fe(IV)=O hMb species to methMb (Fig. 3), although not all of the proteins converted back to methMb even after 10 h of reaction. The autoreduction of Fe(IV)=O hMb to methMb could be caused by intra- or inter-molecular electron transfer between the ferrylheme and amino acid [29]. To estimate the amount of protein decomposition, we calculated the ratios of Abs409 before (Abs409(before)) and after (Abs409(after)) consumption of H2O2 (Abs409(after)/Abs409(before)). The Abs409(after)/Abs409(before) ratios were 0.53 and 0.65 for WT and C110A hMbs, respectively (Figs. 4A and 4B, Table 2), showing that decomposition by H2O2 occurred slightly more in WT than C110A hMb under air. The reaction of hMb with H2O2 was performed under N2 atmosphere to investigate the. 10.
(11) effects of O2 on protein decomposition (Figs. 4C and 4D). The Abs409(after)/Abs409(before) ratios were 0.88 and 0.65 for WT and C110A hMbs, respectively (Table 2). Decomposition of WT hMb decreased significantly with the removal of O2, whereas that of C110A hMb showed no significant change. Since removal of O2 decreases decomposition of WT hMb but not C110A hMb, Cys110 increases decomposition of the protein in the presence of O2. To investigate the effects of Cys110 in more detail, the reaction of hMb with H2O2 was performed under air in the presence of 1 mM GSH. Consumption of H2O2 was faster in WT than C110A hMb in the presence of 1 mM GSH. After the reactions of WT and C110A hMbs with H2O2 in the presence of 1 mM GSH, absorption bands appeared around 541 and 582 nm, which are attributed to oxy hMb (Figs. 4E and 4F) [24]. The ratio of oxy hMb compared to methMb was estimated by subtracting the oxy hMb spectrum from the spectrum obtained after H2O2 consumption. Arbitrary ratios were applied to the subtraction in order to remove the oxygen-related peaks in the difference spectrum at 541 and 582 nm, where the amount of oxy hMb was consequently estimated as 10 and 30% for WT and C110A hMbs, respectively. Taking the ratio of oxy hMb production into consideration, the intrinsic Abs409(after) was calculated from equation 2,. Abs409(after) = Abs409(obs) – Abs409(oxy hMb - methMb) x R. (2). where Abs409(obs) and R represent the observed absorbance at 409 nm and the ratio of the oxy species, respectively. The Abs409(after)/Abs409(before) ratios were estimated as 0.89 and 0.78 for WT and C110A hMbs, respectively (Table 2). The Abs409(after)/Abs409(before) ratio increased from 0.53 to 0.89 in WT hMb, whereas it increased only from 0.65 to 0.78 in C110A hMb by the addition of 1 mM GSH. These results show that GSH suppresses H2O2-induced protein decomposition more effectively in WT than C110A hMb.. 11.
(12) The reactions of WT and C110A hMbs with H2O2 were also performed under N2 atmosphere in the presence of 1 mM GSH (Figs. 4G and 4H). The Abs409(after)/Abs409(before) ratio did not change from 0.89 in WT hMb by removal of O2, whereas it increased from 0.78 to 0.89 in C110A hMb (Table 2). These results indicate that GSH reduces the protein radical of WT hMb efficiently, but not that of C110A hMb.. 3.3. Radicals of WT and C110A hMbs produced by the reaction with H2O2 WT and C110A hMbs were incubated with H2O2 and DMPO for 2 min at room temperature, and their EPR spectra were recorded at room temperature and 77 K. The EPR spectrum of WT hMb at room temperature (Fig. S4) was in agreement with the reported spectrum [16]. The EPR spectrum of WT hMb at 77 K exhibited four peaks at 319.6, 321.2, 323.3, and 326.0 mT in the g = ~2 region (Fig. 5a). The signals observed at room temperature and 77 K are due to the DMPO adducts of the Cys110 thiyl radical. Four peaks were also detected at 319.4, 321.5, 322.9, and 325.9 mT in the spectrum of C110A hMb at 77 K (Fig. 5b). The signals detected in C110A hMb at 77 K are in agreement with the reported spectrum [15] and are attributed to the DMPO adducts of the Tyr103 phenoxyl radical. The peak areas of the EPR signals of WT hMb were about ten times larger than those of C110A hMb. With the addition of 1 mM GSH, the radical concentration estimated from the signals of WT hMb decreased by about 90% (Fig. 5c), whereas that for C110A hMb decreased by about 50% (Fig. 5d). These results show that the thiyl radical in WT hMb was reduced efficiently by electron transfer from GSH. However, the decrease in the radical by addition of GSH to C110A Mb suggests that GSH also reduces the tyrosine radical at Tyr103.. 3.4. Mass spectrometry of hMb reacted with H2O2 in the presence of GSH The MALDI-TOF mass spectra were obtained for WT methMb and WT hMb after the. 12.
(13) reaction with H2O2 in the presence of 100 μM or 50 mM GSH (Fig. S5). The mass determined for WT hMb after the reaction with H2O2 was 17,185 atomic mass unit, which was about the same as that of WT methMb (17,182 atomic mass unit). These results indicate that GSH does not bind to Cys110 significantly when WT hMb reacts with H2O2 in the presence of GSH.. 4. Discussion 4.1. Effect of O2 on the reaction of hMb with H2O2 Decomposition of the protein by H2O2 was suppressed significantly by the removal of O2 for WT hMb but not for C110A hMb. The Cys110 thiyl radical is generated by the reaction of WT hMb with H2O2, whereas the tyrosine phenoxyl radical has been observed in C110A hMb when it reacted with H2O2 [16, 30]. It has been suggested that the Cys110 thiyl radical is generated by electron transfer from the thiolate group of Cys110 to the initial tyrosine phenoxyl radical of Tyr103 [16, 30]. The thiyl radical of WT hMb forms a dimer through a disulfide bond [15, 16]. For C110A hMb, the protein cannot form a dimer with a disulfide bond and decomposes under N2 atmosphere due to the Tyr103 radical. Therefore, protein decomposition was suppressed more in WT than C110A hMb by removing O2 (Figs. 4A-D). The rate of protein decomposition by H2O2 under air was about twice as fast for WT hMb when compared with C110A hMb, although the rates for formation of the ferryloxo species were about the same between the two proteins (Figs. 1 and 2). It has been reported that the reactions of Cys thiyl radicals of organic compounds with O2 produce the reactive Cys thiol peroxyl radicals [31, 32]. The Cys thiyl radical of WT hMb may produce the Cys thiol peroxyl radical in the presence of O2. The unique Cys residue in hMb is unfavorable for the stability of the protein in the presence of O2 due to the high reactivity of the Cys thiyl radical, but Cys110’s electron transfer ability may have an important role in hMb.. 13.
(14) 4.2. Reduction of amino acid radicals in hMb by GSH The rate of H2O2 consumption by WT hMb increased significantly with the addition of 1 mM GSH, although the effects of GSH were less significant for C110A hMb. With the addition of GSH, decomposition of WT hMb by H2O2 was also suppressed, but that of C110A hMb was suppressed less (Figs. 4E and 4F). Furthermore, the intensity of the EPR signals of the thiyl radical in WT hMb decreased significantly with the addition of GSH, while those of the phenoxyl radical in C110A hMb did not (Fig. 5). Therefore, the increase in the rate of H2O2 consumption and decrease in decomposition of the protein were attributed to reduction of the thiyl radical by GSH [14, 19-21]. Although a small amount of the oxy species of WT and C110A hMbs was formed by reactions with H2O2 in the presence of 1 mM GSH, reduction of methMb to oxy hMb was not significant. It has been reported that GSH does not directly reduce metMb to ferrous oxyMb but reduces ferryloxo Mb to metMb producing the glutathione radical [33, 34]. WT hMb reacted with H2O2 faster than C110A hMb in the presence of GSH, suggesting that conversion of ferryloxo hMb to methMb by GSH is faster in WT hMb than in C110A hMb. These results indicate that GSH may largely affect conversion of ferryloxo hMb to methMb through reduction of the Cys thiyl radical. It has been reported that the Cys thiyl radical is generated by intramolecular electron transfer from Cys110 to the Tyr103 radical in hMb by the reaction with H2O2, depending on the protein concentration [16]. Since GSH reduced the amount of radicals produced by the reaction of C110A hMb with H2O2 (Fig. 5), GSH may also suppress production of the Cys110 thiyl radical of hMb by inhibition of intermolecular electron transfer between the tyrosyl radical at Tyr103 and the thiol at Cys110 on another molecule, in addition to the direct reduction of the thiyl radical by GSH. Formation of the cross-linked heme (Tyr103-heme) by. 14.
(15) the reaction of horse Mb with H2O2 has also been reported [35]. The cross-linked heme could be produced in the present reaction between hMb and H2O2, because hMb also possesses Tyr103. Since the EPR measurements show that GSH reduced the amounts of the amino acid radicals of WT and C110A hMbs (Fig. 5), GSH is likely to suppress the formation of the cross-linked heme by the reduction of amino acid radicals. No peak was observed at the glutathione-bound WT hMb position in the MALDI-TOF mass spectrum after the reaction of WT hMb with H2O2 (Fig. S5). A glutathione radical is produced when GSH donates an electron to the thiyl radical. It has been proposed that the glutathione radical is decomposed through several steps in the presence of O2 [19, 33]. The glutathione disulfide (GSSG) is generated when two glutathione radical molecules react with each other, whereas a glutathione disulfide anion radical is produced when the glutathione radical reacts with GSH. This disulfide anion radical, in turn, reacts with O2, and a glutathione disulfide and a superoxide are generated, in which the superoxide is quenched by GSH [19]. For the reaction of hMb with H2O2, the GSSG concentration increased about two-fold (40 μM) compared to the initial H2O2 concentration by incubation of hMb with H2O2 for 2 h (Fig. S6). These results indicated production of the glutathione radical. When the ferryloxo species of WT hMb is generated by the reaction with H2O2, the thiyl radical is produced simultaneously. The produced thiyl radical reacts with O2 rapidly and the Cys thiol peroxyl radical is produced. It is presumed that the produced Cys thiol peroxyl radical reacts with the protein and decomposes it. Actually, the rate of decomposition by H2O2 under air was twice as fast for WT hMb when compared with C110A hMb (Fig. 2). In the presence of GSH, effective electron transfer from GSH to the thiyl radical directly or indirectly through the tyrosyl radical of another molecule may protect the protein from decomposition even under air. It is noteworthy that the Abs409(after)/Abs409(before) ratio of WT hMb was larger than that of C110A hMb in the presence of 1 mM GSH under air, although. 15.
(16) decomposition of C110A hMb by H2O2 was suppressed with addition of 1 mM GSH under N2 atmosphere (Table 2, Figs. 4E-H). After consumption of H2O2, electron transfer from the Cys residue to the ferryloxo site may produce methMb. In conclusion, the C110 thiyl radical receives an electron from GSH efficiently. The electron transfer prevents protein decomposition caused by amino acid radicals and the reactive oxygen species produced during the reaction of hMb with H2O2, although the thiyl radical enhances decomposition of the protein with O2 in the absence of GSH.. Acknowledgements We thank Ms. Akemi Shodai and Ms. Chie Nakayama for their help in WT and C110A hMb preparations. We thank Mr. Steven Nishida and Mr. Leigh McDowell for their advice during manuscript preparation. This work was partially supported by a Grant-in-Aid for Scientific Research from the Japan Society for Promotion of Science (Category B, No. 21350095, S.H.) and Japan Science and Technology Agency (Research Seeds Quest Program, S.H.).. Appendix A. Supplementary data Supplementary data associated with this article can be found, in the online version, at doi:XXXXXX/XXXXXX.. References [1]. J.C. Kendrew, R.E. Dickerson, B.E. Strandberg, R.G. Hart, D.R. Davies, D.C. Phillips, V.C. Shore, Structure of myoglobin: A three-dimensional Fourier synthesis at 2 Å resolution, Nature 185 (1960) 422-427.. [2]. S.R. Hubbard, W.A. Hendrickson, D.G. Lambright, S.G. Boxer, X-ray crystal. 16.
(17) structure of a recombinant human myoglobin mutant at 2.8 Å resolution, J. Mol. Biol. 213 (1990) 215-218. [3]. B.A. Springer, S.G. Sligar, J.S. Olson, G.N. Phillips, Mechanisms of ligand recognition in myoglobin, Chem. Rev. 94 (1994) 699-714.. [4]. P.K. Witting, D.J. Douglas, A.G. Mauk, Reaction of human myoglobin and nitric oxide. Heme iron or protein sulfhydryl (S) nitrosation dependence on the absence or presence of oxygen, J. Biol. Chem. 276 (2001) 3991-3998.. [5]. S. Hirota, K. Azuma, M. Fukuba, S. Kuroiwa, N. Funasaki, Heme reduction by intramolecular electron transfer in cysteine mutant myoglobin under carbon monoxide atmosphere, Biochemistry 44 (2005) 10322-10327.. [6]. S. Nicolis, M. Zucchelli, E. Monzani, L. Casella, Myoglobin modification by enzyme-generated dopamine reactive species, Chemistry 14 (2008) 8661-8673.. [7]. P. George, D.H. Irvine, The reaction between metmyoglobin and hydrogen peroxide, Biochem. J. 52 (1952) 511-517.. [8]. J.F. Gibson, D.J. Ingram, P. Nicholls, Free radical produced in the reaction of metmyoglobin with hydrogen peroxide, Nature 181 (1958) 1398-1399.. [9]. N.K. King, M.E. Winfield, The mechanism of metmyoglobin oxidation, J. Biol. Chem. 238 (1963) 1520-1528.. [10]. P.R. Ortiz de Montellano, C.E. Catalano, Epoxidation of styrene by hemoglobin and myoglobin. Transfer of oxidizing equivalents to the protein surface, J. Biol. Chem. 260 (1985) 9265-9271.. [11]. A. Wilks, P.R. Ortiz de Montellano, Intramolecular translocation of the protein radical formed in the reaction of recombinant sperm whale myoglobin with H2O2, J. Biol. Chem. 267 (1992) 8827-8833.. [12]. S.I. Rao, A. Wilks, P.R. Ortiz de Montellano, The roles of His-64, Tyr-103, Tyr-146,. 17.
(18) and Tyr-151 in the epoxidation of styrene and beta-methylstyrene by recombinant sperm whale myoglobin, J. Biol. Chem. 268 (1993) 803-809. [13]. C. Giulivi, E. Cadenas, Ferrylmyoglobin: formation and chemical reactivity toward electron-donating compounds, Methods Enzymol. 233 (1994) 189-202.. [14]. D.J. Kelman, J.A. Degray, R.P. Mason, Reaction of myoglobin with hydrogen-peroxide forms a peroxyl radical which oxidizes substrates, J. Biol. Chem. 269 (1994) 7458-7463.. [15]. P.K. Witting, D.J. Douglas, A.G. Mauk, Reaction of human myoglobin and H2O2. Involvement of a thiyl radical produced at cysteine 110, J. Biol. Chem. 275 (2000) 20391-20398.. [16]. P.K. Witting, A.G. Mauk, Reaction of human myoglobin and H2O2. Electron transfer between tyrosine 103 phenoxyl radical and cysteine 110 yields a protein-thiyl radical, J. Biol. Chem. 276 (2001) 16540-16547.. [17]. D. Tew, P.R. Ortiz de Montellano, The myoglobin protein radical. Coupling of Tyr-103 to Tyr-151 in the H2O2-mediated cross-linking of sperm whale myoglobin, J. Biol. Chem. 263 (1988) 17880-17886.. [18]. L.S. Harman, D.K. Carver, J. Schreiber, R.P. Mason, One- and two-electron oxidation of reduced glutathione by peroxidases, J. Biol. Chem. 261 (1986) 1642-1648.. [19]. C.M. Jones, A. Lawrence, P. Wardman, M.J. Burkitt, Electron paramagnetic resonance spin trapping investigation into the kinetics of glutathione oxidation by the superoxide radical: re-evaluation of the rate constant, Free Radic. Biol. Med. 32 (2002) 982-990.. [20]. G.V. Buxton, C.L. Greenstock, W.P. Helman, A.B. Ross, Critical-review of rate constants for reactions of hydrated electrons, hydrogen-atoms and hydroxyl radicals (·OH/·O-) in aqueous-solution, J. Phys. Chem. Ref. Data 17 (1988) 513-886.. 18.
(19) [21]. B.E. Sturgeon, H.J. Sipe, Jr., D.P. Barr, J.T. Corbett, J.G. Martinez, R.P. Mason, The fate of the oxidizing tyrosyl radical in the presence of glutathione and ascorbate. Implications for the radical sink hypothesis, J. Biol. Chem. 273 (1998) 30116-30121.. [22]. A. Wahllander, S. Soboll, H. Sies, I. Linke, M. Muller, Hepatic mitochondrial and cytosolic glutathione content and the subcellular distribution of GSH-S-transferases, FEBS Lett. 97 (1979) 138-140.. [23]. R.M. Horton, PCR Protocols: Current Methods and Applications, Humana Press Inc. Totowa, NJ 1993.. [24]. E. Antonini, M. Brunori, Hemoglobin and Myoglobin in Their Reaction with Ligands, North Holland Publishing Company, Amsterdam, 1971.. [25]. A. Dobrina, P. Patriarca, Neutrophil-endothelial cell interaction. Evidence for and mechanisms of the self-protection of bovine microvascular endothelial cells from hydrogen peroxide-induced oxidative stress, J. Clin. Invest. 78 (1986) 462-471.. [26]. P.K. Witting, A.G. Mauk, P.A. Lay, Role of tyrosine-103 in myoglobin peroxidase activity: kinetic and steady-state studies on the reaction of wild-type and variant recombinant human myoglobins with H2O2, Biochemistry 41 (2002) 11495-11503.. [27]. M.J. Davies, The oxidative environment and protein damage, Biochim. Biophys. Acta 1703 (2005) 93-109.. [28]. C. Schoneich, Mechanisms of protein damage induced by cysteine thiyl radical formation, Chem. Res. Toxicol. 21 (2008) 1175-1179.. [29]. O.M. Lardinois, P.R. Ortiz de Montellano, Autoreduction of ferryl myoglobin: discrimination among the three tyrosine and two tryptophan residues as electron donors, Biochemistry 43 (2004) 4601-4610.. [30]. S. Bhattacharjee, L.J. Deterding, J. Jiang, M.G. Bonini, K.B. Tomer, D.C. Ramirez, R.P. Mason, Electron transfer between a tyrosyl radical and a cysteine residue in. 19.
(20) hemoproteins: Spin trapping analysis, J. Am. Chem. Soc. 129 (2007) 13493-13501. [31]. M.D. Sevilla, M.Y. Yan, D. Becker, Thiol peroxyl radical formation from the reaction of cysteine thiyl radical with molecular oxygen: an ESR investigation, Biochem. Biophys. Res. Commun. 155 (1988) 405-410.. [32]. K. Schafer, M. Bonifacic, D. Bahnemann, K.D. Asmus, Addition of oxygen to organic sulfur radicals, J. Phys. Chem. 82 (1978) 2777-2780.. [33]. D. Galaris, E. Cadenas, P. Hochstein, Glutathione-dependent reduction of peroxides during ferryl- and met-myoglobin interconversion: a potential protective mechanism in muscle, Free Radic. Biol. Med. 6 (1989) 473-478.. [34]. O. Augusto, S. Lopes de Menezes, E. Linares, N. Romero, R. Radi, A. Denicola, EPR detection of glutathiyl and hemoglobin-cysteinyl radicals during the interaction of peroxynitrite with human erythrocytes, Biochemistry 41 (2002) 14323-14328.. [35]. C.E. Catalano, Y.S. Choe, P.R. Ortiz de Montellano, Reactions of the protein radical in peroxide-treated myoglobin. Formation of a heme-protein cross-link, J. Biol. Chem. 264 (1989) 10534-10541.. 20.
(21) Figure Captions Fig. 1.. Absorption spectra of WT methMb (2 μM) before (broken line) and after (solid line) the reaction with H2O2 (20 μM) for 10 min. The absorption spectra before and after the reaction correspond to those of the met and the Fe(IV)=O species, respectively. The inset shows the plot of the pseudo-first-order rate constants kobs(Fe(IV)) vs. H2O2 concentration for formation of the Fe(IV)=O species: WT (filled circle) and C110A (open circle).. Fig. 2.. Absorption spectra changes of WT methMb (2 μM) in the reaction with H2O2 (20 μM). The spectra after 10-200 min of reaction are shown. The spectral changes correspond to protein decomposition. The inset shows the plot of the pseudo-first-order rate constants kobs(dec) vs. H2O2 concentration: WT (filled circle) and C110A (open circle).. Fig. 3.. Absorption spectral changes of WT methMb (2 μM) in the reaction with H2O2 (20 μM) for 400-750 min. The spectral changes correspond to conversion of the Fe(IV)=O to the met species.. Fig. 4.. Absorption spectra of WT (A, C, E, and G) and C110A (B, D, F, and H) methMbs before (broken lines) and after (solid lines) the reaction with H2O2: Under air (A and B), under N2 atmosphere (C and D), and in the presence of GSH (1 mM) under air (E and F) and under N2 atmosphere (G and H). The spectra were recorded after completion of the reaction with H2O2 (700, 840, 400, 1200, 280, 360, 220, and 380 min for A, B, C, D, E, F, G, and H, respectively). The visible region (450-700 nm) of each spectrum is enlarged by seven.. Fig. 5.. EPR spectra of WT (a and c) and C110A (b and d) methMbs (500 μM) after the reaction with H2O2 (2.5 mM) and DMPO (5 mM) for 2 min. The spectra were recorded in the absence (a and b) and presence (c and d) of GSH (1 mM) at 77 K.. 21.
(22) The concentrations of the DMPO adducts of WT and C110A hMbs were estimated to be 0.14 and 0.011 spins/mol (in the absence of GSH) and 0.014 and 0.0075 spin/mol (in the presence of GSH), respectively.. 22.
(23) Fig. 1. 1.
(24) Fig. 2. 2.
(25) Fig. 3. 3.
(26) Fig. 4 4.
(27) Fig. 5. 5.
(28) Supporting Information. Efficient Reduction of Cys110 Thiyl Radical by Glutathione in Human Myoglobin Satoshi Nagaoa, Osamu Asamia, Hiroyuki Yasuib, and Shun Hirotaa,* a. Graduate School of Materials Science, Nara Institute of Science and Technology, 8916-5 Takayama-cho, Ikoma, Nara 630-0192, Japan, bDepartment of Analytical & Bioinorganic Chemistry, Kyoto Pharmaceutical University, 5 Nakauchi-cho, Misasagi, Yamashina-ku, Kyoto 607-8414, Japan. *. Corresponding author: Shun Hirota, Graduate School of Materials Science, Nara Institute of Science and Technology, 8916-5 Takayama-cho, Ikoma, Nara 630-0192, Japan. Tel.:+81-743-72-6110; Fax: +81-743-72-6119; E-mail: [email protected].. This section contains Fig. S1, Fig. S2, Fig. S3, Fig. S4, Fig. S5, and Fig. S6 as supporting information. 1.
(29) Fig. S1 Absorption spectra of WT (solid line) and C110A (broken line) methMbs (2 μM) in 50 mM potassium phosphate buffer, pH 7.0, at 20 °C.. 2.
(30) Fig. S2 Plot of the pseudo-first-order rate constants kobs(Fe(IV)) vs. H2O2 concentration for formation of the Fe(IV)=O species under N2 atmosphere: WT (filled circle) and C110A (open circle).. 3.
(31) Fig. S3 Plot of the pseudo-first-order rate constants kobs(Fe(IV)) vs. H2O2 concentration for formation of the Fe(IV)=O species under air in the presence of 1 mM GSH: WT (filled circle) and C110A (open circle).. 4.
(32) Fig. S4 EPR spectra of WT (a) and C110A (b) methMbs (500 μM) after the reaction with H2O2 (2.5 mM) and DMPO (5 mM) in the absence of GSH for 2 min. The spectra were recorded at room temperature. The concentrations of the DMPO adducts of WT and C110A hMbs were estimated to be 0.12 and 0.014 spin/mol, respectively.. 5.
(33) Fig. S5 MALDI-TOF mass spectra of WT methMb (100 μM) before (a) and after (b, c) the reaction with H2O2 (500 μM) in the presence of GSH (100 μM (b) and 50 mM (c)). The region of mass number 17050-17650 of each spectrum is enlarged by three. The peak around 17,400 atomic mass unit in spectra b and c are presumably due to the WT hMb-matrix adducts. Spectrum b was obtained for Mb after the reaction at molar ratio of hMb:GSH = 1:1 to avoid the replacement of the disulfide bond between Cys and GSH with that between two GSH molecules. No GSH adduct of hMb was detected in either spectrum b or c.. 6.
(34) Fig. S6 Generation of glutathione disulfide (GSSG) by the reaction between hMb and H2O2 in the presence of 1 mM GSH under N2 atomosphere: WT (filled circle) and C110A (open circle). The experiment was performed as follows1): methMb (2 μM) in 20 mM potassium phosphate buffer, pH 7.0, in the presence of 1 mM GSH was mixed with H2O2 (final concentration: 20 μM) under N2 atmosphere. The reaction was stopped by addition of an equal volume of 2 M HCl to the solution. The pH of the solution was neutralized with 1 M NaOH to obtain the sample solution. The sample solution (100 μL) was mixed with 100 μM NADPH (900 μL) and EDTA was added (0.6 mM, final concentration). Oxidation of NADPH was initiated by addition of glutathione reductase (0.6 μM, final concentration), and the GSSG concentration was calculated from the absorbance at 340 nm (ε340nm = 6.22 mM-1cm-1 ).1). References 1) T. P. M. Akerboom and H. Sies, Methods Enzymol. (1981) 77, 373-382; L. S. Harman et al., J. Biol. Chem. (1986) 261, 1642-1648.. 7.
(35)
図



+3


関連したドキュメント
In this paper we develop a general decomposition theory (Section 5) for submonoids and subgroups of rings under ◦, in terms of semidirect, reverse semidirect and general
(Construction of the strand of in- variants through enlargements (modifications ) of an idealistic filtration, and without using restriction to a hypersurface of maximal contact.) At
It is suggested by our method that most of the quadratic algebras for all St¨ ackel equivalence classes of 3D second order quantum superintegrable systems on conformally flat
This paper develops a recursion formula for the conditional moments of the area under the absolute value of Brownian bridge given the local time at 0.. The method of power series
Then it follows immediately from a suitable version of “Hensel’s Lemma” [cf., e.g., the argument of [4], Lemma 2.1] that S may be obtained, as the notation suggests, as the m A
Some new oscillation and nonoscillation criteria are given for linear delay or advanced differential equations with variable coef- ficients and not (necessarily) constant delays
Our method of proof can also be used to recover the rational homotopy of L K(2) S 0 as well as the chromatic splitting conjecture at primes p > 3 [16]; we only need to use the
In this paper we focus on the relation existing between a (singular) projective hypersurface and the 0-th local cohomology of its jacobian ring.. Most of the results we will present
