博士学位論文
Study on the mode of action of waldiomycin
近畿大学大学院
農学研究科 バイオサイエンス専攻
Md. Fakhruzzaman
Doctoral Dissertation
Study on the mode of action of waldiomycin
January 8
th2016
Graduate School, Kinki University
Division of Agricultural Science
(Major: Bioscience)
Md. Fakhruzzaman
ABBREVIATIONS
AA acrylamide
a.a amino acid
ADP adenosine 5’-diphosphate
Ala alanine
Ap ampicillin
Asp (D) aspartic acid
ATP adenosine 5’ -triphosphate
Bp base pair
BPB bromophenol blue
B. subtilis Bacillus subtilis
BSA bovine serum albumin
C cytosine
cDNA complementary DNA
Cm chloramphenicol
conc. concentration
DMSO dimethyl sulfoxide
DNA deoxyribonucleic acid
DNase deoxyribonuclease
EDTA ethylenediaminetetraacetate
E. faecalis Enterococcus faecalis
G guanine
g gravity
Glu glutamic acid
Hr hour (s)
HK histidine kinase
IC50 50% inhibitory concentration
IPTG isopropyl β-D-1-thiogalactopyranoside
Kd dissociation constant
LB medium Luria-Bertani medium
Lys lysine
M molar
Min minute (s)
mM millimolar
MRSA methicillin-resistant Staphylococcus aureus
MW molecular weight
Ng nanogram
nM nanomolar
NMR nuclear magnetic resonance
OD optical density
PAGE polyacrylamide gel electrophoresis
PBS phosphate-buffered saline
PCR polymerase chain reaction
RNA ribonucleic acid
Rpm revolutions per minute
RR response regulator
RT-PCR reverse transcription polymerase chain reaction
S. aureus Staphylococcus aureus
SDS sodium dodecyl sulfate
Sec second (s)
TCS two-component signal transduction system
Tris tris (hydroxymethyl) aminomethane
TSB trypticase soy broth
V volume
VRE vancomycin-resistant Enterococcus faecalis
CONTENTS
CHAPTER I Introduction 1
CHAPTER II In vitro mode of action 19
CHAPTER III In vivo mode of action 26
CHAPTER IV
Effects of waldiomycin on WalK sensor histidine kinase localization to the cell
division septum 40 CHAPTER V Summary 49 REFERENCES 51 PUBLICATION 66 CHAPTER I Introduction
Two-component signal transduction system in bacteria
Microorganisms are the most fluctuating and flexible organisms. Life in the microbial world is characterized by continuous interactions between the bacterial cell and its environment. Bacteria inhabit soil, water, acidic hot springs, radioactive waste, and the deep portions of Earth’s crust. To stay alive and compete for resources bacteria must quickly and efficiently respond to an ever-changing environment. The ability of a bacterium to monitor environmental parameters, including osmotic activity and ionic strength, pH, temperature, and the concentrations of nutrients and harmful compounds, is a prerequisite for survival. For that purpose, bacteria have evolved surface-exposed signal transduction systems, typically comprised of transmembrane (TM) proteins that channel the input from sensory modules to intracellular responses. To sense environmental conditions bacteria use a variety of systems, which include secondary
metabolites and ions, such as cyclic adenosine monophosphate and Fe2+, and regulatory
proteins, such as catabolite regulatory protein (CRP) and ferric uptake regulator (Fur), whose affinity for DNA changes on ligand binding. However, two-component systems (TCSs) are the predominant form of signal transduction used by bacteria to respond to environmental stresses (Hoch and Silhavy, 1995; Stock et al., 2000).
TCSs are the most widespread signal transduction systems found in prokaryotes and regulate many bacterial behaviours such as virulence, pathogenicity, motility, nodulation, nutrient uptake, secondary metabolite production, metabolic regulation, cell division, etc. (Galperin, 2004). They have also been found in yeast and plants (Chang et
al., 1993; Maeda et al., 1994), but are absent in mammals (Thomason and Kay, 2000),
which makes them potential targets for antimicrobials drug design (Watanabe et al., 2008). They are relatively simple signalling pathways, first identified as a distinct class of signalling pathway 30 years ago (Nixon et al., 1986), tens of thousands of TCSs have now been identified in sequenced bacterial genomes and metagenomes (Ulrich and
Zhulin, 2007; Barakat et al., 2009). According to the microbial signal transduction
database MiST2, 1087 sequenced bacterial genomes encode for 63,259 TCS proteins, and hence, the average genome features roughly 29 individual signalling systems
(Szurmant and Hoch, 2010; Ulrich and Zhulin, 2010). The average eubacterial genome
encodes 50-60 TCS proteins (5-15 in Archaebacteria and Eukaryotes), but some organisms possess more than 250 (Ashby, 2006; Whitworth, 2008; Wuichet et al., 2010).
TCSs have global roles in bacterial functions such as growth, drug resistance, virulence, biofilm formation and quorum sensing (Watanabe et al., 2008). Therefore,
inhibitors of TCS can be expected to work as antibacterial agents as well as inhibitors for drug resistance, biofilm formation, quorum sensing and virulence. Interrupting these critical signalling pathways may provide an alternative strategy for the development of novel classes of preventive and/or therapeutic agents (Barrett and Hoch, 1998). TCSs are ideally suited to function in the integration of multiple signals because the membrane location of the sensor protein allows transduction of environmental cues, and the high degree of specificity of each sensor/regulator pair ensures a physiological response. Different TCSs can process multiple signals into a particular response by interacting with one another and/or by phosphorylating from small molecular weight phosphodonors (Lawhon et al., 2002; McCleary and Stock, 1994).
Histidine Kinases (HKs)
TCSs are comprised of a sensor histidine kinase (HK) and its cognate response regulator (RR). Depending on a particular environmental signal, the sensor HK autophosphorylates a conserved histidine residue in the dimerization domain, and then transfers the phosphoryl group to the conserved aspartic acid residue in the regulatory domain of its cognate RR. The HK catalyses its own auto-phosphorylation followed by the transfer of the phosphoryl group to the receiver domain on RR; phosphorylation of the RR usually activates an attached output domain, which can then effect changes in cellular physiology, often by regulating gene expression. Some HKs are bifunctional, catalysing both the phosphorylation and dephosphorylation of their cognate RR. The input stimuli can regulate either the kinase or phosphatase activity of the bifunctional HK.HKs are multifunctional, typically transmembrane, proteins of the transferase class that play a role in signal transduction across the cellular membrane (Wolanin et al., 2002). The vast majority of HKs are homodimers that exhibit autokinase,
phosphotransfer, and phosphatase activity. HKs can act as cellular receptors for signalling molecules in a way analogous to tyrosine kinase receptors (RTK). Multifunctional receptor molecules such as HKs and RTKs typically have portions on the outside of the cell (extracellular domain) that bind to hormone- or growth factor-like molecules, portions that span the cell membrane (transmembrane domain), and portions within the cell (intracellular domain) that contain the enzymatic activity. In addition
to kinase activity, the intracellular domains typically have regions that bind to a
secondary effector molecule or complex of molecules that further propagate signal transduction within the cell.
Response Regulators (RRs)
RRs are typically found at the ends of phosphotransfer pathways where they function as phosphorylation-activated switches that regulate output responses. These proteins usually have a two- (or more) domain structure with a conserved N-terminal regulatory domain and a variable C-terminal effector domain (s). The regulatory domains of RRs have three activities. First, they interact with phosphorylated HKs and catalyze transfer of a phosphoryl group to one of their own Asp residues. Second, they catalyze autodephosphorylation. And third, they regulate the activities of their associated effector domains in a phosphorylation-dependent manner. The conserved regulatory domains, often called receiver domains, can also be found within hybrid HKs or as isolated proteins within phosphorelay pathways. In these contexts, the receiver domains are not physically connected to effector domains and play no direct role in regulating effector domain function. RRs are mostly DNA-binding transcription factors whose affinities for their target promoters are modulated by phosphorylation. Subsequently, alteration of the phosphorylated state of RRs results in different gene expression profiles (Gao and Stock, 2009). Phosphorylation of a RR changes the biochemical properties of its output domain, which can participate in DNA binding and transcriptional control, perform enzymatic activities, bind RNA, or engage in protein– protein interactions (Gao et al., 2007).
Phosphorelays
Phosphorelays are a more complex version of the TCS in which a sensor kinase first transfers the phosphoryl group to a RR possessing the domain with the conserved aspartate but no output domain (Appleby et al., 1996; Perraud et al., 1999). The RR subsequently transfers the phosphoryl group to a histidine-containing phosphotransfer protein, and it is the latter protein that serves as a phosphodonor to the terminal RR, which possesses an output domain mediating a cellular response. In some phosphorelays, the sensor HK and the RR lacking the output domain (and sometimes also the histidine-containing phosphotransfer protein) are fused in a single polypeptide (Appleby et al., 1996).
The vast majority of RRs are active only when phosphorylated (Hoch, 2000; Gao
et al., 2007). Therefore, any condition or product that affects the phosphorylated state of
a RR will impact its ability to exert its biological functions. Consequently, the output of a RR is determined not only by the presence of the specific signals sensed by its cognate sensor kinase but also by gene products that stimulate or inhibit its phosphorylation.
Such products can, in principle, target any one of the various steps leading to phosphorylation of the RR, including sensor kinase autophosphorylation, phosphotransfer to the RR, dephosphorylation of a phosphorylated RR, and the activity of the output domain. The presence of multiple stages in a phosphorelay provides additional potential targets for control.
TCS connectors
The TCS connectors are an emerging group of proteins that modulate the activity of sensor kinases and RRs at the post-translational level (Eguchi et al., 2012). Because connector proteins are typically synthesized in response to signals that are different from those sensed by the cognate sensor, they often establish regulatory links between otherwise independent signal transduction pathways (in other words, they “connect” a TCS to the signal (s) controlling a different regulatory system). Bacterial TCS connectors play the critical roles in a variety of cellular functions, including the adaptation to nutrient-limiting conditions, sporulation, competence, antibiotic resistance, and the transition to stationary phase.
The WalRK two-component signal transduction system
Two-component systems have long been known to serve as a primary means of bacteria for niche adaptation to ever changing environmental conditions. Upon completion of the first bacterial genome sequencing projects, the global importance of TCSs to the bacteria became increasingly clear when some genomes were found to feature more than 100 such signalling systems (Galperin, 2005). It also became evident that mutations in most systems had not been identified in genetic screens to confer a desired phenotype. Hence no function had been assigned to the majority of these systems. This was true for even the most scrutinized model organisms, such as
Escherichia coli and Bacillus subtilis. The lack of knowledge on most systems fuelled
systematic deletion studies in an attempt to identify roles for systems of yet unknown
function (Fabret and Hoch, 1998;Zahrt and Deretic, 2000).
In B. subtilis, a systematic deletion study revealed a single system, the WalRK system, could not be inactivated and was thus deemed a possibly essential signalling system (Fabret and Hoch, 1998). Subsequent work with conditional mutants and with strains where the system was placed under IPTG inducible control proved the essentiality of the system in the model organism (Fabret and Hoch, 1998; Fukuchi et al.,
2000).
The identification of an essential signalling system was perhaps somewhat surprising. It did not conform to the common perception of a role for TCSs in niche adaptation. Rather it became clear that the system has to be embedded in important processes regulating the life cycle of the cell. A phylogenetic analysis of a limited number of available sequenced organisms at the time suggested that orthologous systems are widespread and perhaps even ubiquitous among the low-G+C Gram-positives, also known as the phylum Firmicutes (Fabret and Hoch, 1998). The initial identification of this system as being essential in B. subtilis fuelled a flurry of studies with various different aims. First, it became imperative to identify the cellular processes that the WalRK system is regulating, with the expectation that such knowledge would lead to an increased understanding of bacterial physiology. Second, the identification of orthologous systems fuelled the question whether the system was also essential and conserved a role in other organisms, in particular important human pathogens, such as Staphylococcus aureus and Streptococcus pneumoniae. Lastly, the initial identification of these systems as essential and the lack of similar systems in the animal kingdom, suggested that the WalRK system might be a good potential drug target.
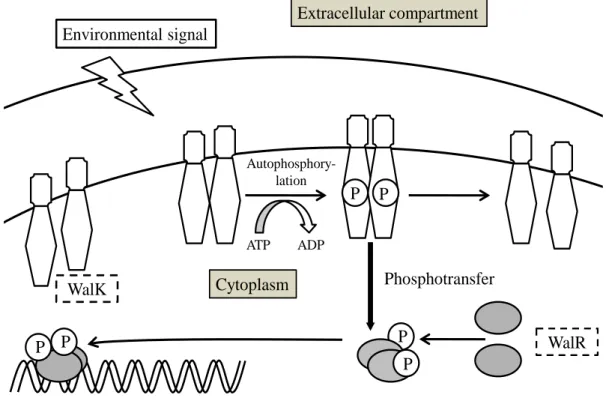
The WalK (HK)/WalR (RR) two-component signal transduction system (Fig. 1)
is highly conserved among Gram-positive bacteria having a low GC content, including
B. subtilis, S. aureus, Enterococcus faecalis, Listeria monocytogenes, Streptococcus mutans, S. pneumoniae, and Streptococcus pyogenes (Dubrac et al., 2008). This TCS is
also specific to Gram-positive bacteria, such as B. subtilis and S. aureus, in which it exerts an effect as a master regulatory system for cell wall metabolism and is essential for cell viability (Bisicchia et al., 2007; Dubrac et al., 2007; Dubrac and Msadek, 2004; Fabret and Hoch, 1998; Martin et al., 1999). Thus, inhibitors directed against WalK/WalR may have a bactericidal effect and may be active against an array of clinically important Gram-positive pathogens. Originally identified in B. subtilis, the WalK/WalR system has since been extensively studied, and in these studies it has various designations (YycG/YycF, VicK/VicR or MicA/MicB).
TABLE 1. Essential TCSs for cell growth (Gotoh et al., 2010)
TCS (HK/RR) Bacterium Function Regulated gene Disease Ref.
WalK/WalRa S. aureus Cell wall metabolism isaA, ssaA, lytM Opportunistic infection Martin et al. (1999)
S. pneumoniae Cell wall metabolism pcsB, lytN, fabK Pneumoniae Lange et al. (1999)
S. mutans Biofilm formation gtfBCD, ftf, gdpB Caries Senadheera et al. (2005)
S. pyogenes Cell wall metabolism Unknown Necrotizing fasciitis Liu et al. (2006)
S. epidermidis Cell wall metabolism Unknown Opportunistic infection Qin et al. (2006)
L. monocytogenes Unknown Unknown Listeriosis Kallipolitis and Ingmer (2001)
E. faecalis Unknown Unknown Urinary tract infection Hancock and Perego (2002)
Bacterial endocarditis
YhcS/YhcR S. aureus Unknown Unknown Opportunistic infection Sun et al. (2005) HP165/HP166 H. Pylori Unknown Unknown Chronic gastritis Beier and Frank (2000);
Dietz et al. (2002)
MtrB/MtrA M. tuberculosis Unknown dnaA Tuberculosis Zahrt and Deretic (2000); Fol et al. (2006)
S. aureus: Staphylococcus aureus; S. pneumoniae: Streptococcus pneumoniae; S. mutans: Streptococcus mutans; S. pyogenes: Streptococcus pyogenes; S. epidermidis: Staphylococcus epidermidis; L. monocytogenes: Listeria monocytogenes; E. faecalis: Enterococcus faecalis; H. pylori: Helicobacter pylori; M. tuberculosis: Mycobacterium tuberculosis.
aalso known as YycG/YycF, VicK/VicR, MicA/MicB.
The first identified member of the WalK/WalR regulon was ftsAZ in B. subtilis, signalling a potential role in cell division (Fukuchi et al., 2000). Recent studies indicate that cell wall metabolism genes predominate in the WalK/WalR regulon for the three bacteria where this system has been extensively studied. In B. subtilis, WalK/WalR activate expression of yocH, yvcE, and lytE, which encode autolysins, and ydjM, which is predicted to encode a cell wall-associated protein (Bisicchia et al., 2007; Howell et al., 2003; Stapleton et al., 2007). Additionally, WalK/WalR negatively regulate the expression of yoeB and yjeA, which encode proteins that modulate autolysin activity (Bisicchia et al., 2007; Salzberg and Helmann, 2007). There are nine cell wall metabolism genes which potentially belonging to the WalRK regulon in S. aureus (Dubrac et al., 2007). These genes encode two major S. aureus autolysins (atlA and
lytM) and lytic transglycosylases (isaA and sceD) (Dubrac et al., 2007; Dubrac and
Msadek, 2004; Stapleton et al., 2007) and five proteins that contain a CHAP amidase domain (ssaA, SA0620, SA2097, SA2353, and SA0710) (Dubrac et al., 2007, 2008). Direct binding of WalR to the promoter regions of ssaA, isaA and lytM was shown, indicating that genes involved in cell wall metabolism are controlled directly by the WalK/WalR system (Dubrac and Msadek, 2004; Dubrac et al., 2007).
Fig. 1. The WalK/WalR two-component signal transduction system.
WalK/WalR is attractive as a target for antimicrobials for the following reasons: (i) None has been found in mammalian genomes. The WalK/WalR signal transduction system is distinct from serine/threonine and tyrosine phosphorylation in higher eukaryotes. (ii) WalK and WalR inhibitors are expected to be a new class of antibiotics
for multi-drug-resistant bacteria such as MRSA and VRE.
Architecture and structures
Both the WalR RR as well as the WalK kinase, feature some of the most common domains found for TCSs proteins. The WalR RR is of the OmpR-type, featuring two domains, the RR receiver domain and the winged helix-turn-helix DNA-binding domain, also known as the trans_reg_c domain (Galperin, 2006) (Fig. 2). No structure of the full-length WalR protein is currently available. High-resolution structures are available for isolated RR domains of WalR from B. subtilis and S. pneumoniae (Bent et al., 2004; Zhao et al., 2009). Similarly, high-resolution structures are available for isolated DNA-binding domains of E. faecalis, B. subtilis and S. aureus WalR proteins (Trinh et
al., 2007; Okajima et al., 2008). These structures have revealed no surprises and ATP ADP
Regulation of gene expression
Phosphotransfer WalR Autophosphory-lation P P P P P P Environmental signal Extracellular compartment Cytoplasm WalK
generally conform to the typical OmpR-type RR architecture. Perhaps these structures might in the future be useful for structure-based drug design. Based on extensive microarray, gel shift and DNAase I foot printing data from B. subtilis, S. aureus and S.
pneumoniae a cross-organismal consensus DNA binding sequence for WalR has been
determined as 5′-TGTNDH-N5-BKBWRN-3′ (Dubrac et al., 2008). OmpR-type RRs typically bind six basepair direct repeats with a four to five base pair spacing, and the WalR consensus sequence follows this common pattern (Blanco et al., 2002).
The WalK kinase features a complex domain-architecture (Fig. 2). Two subtypes can be distinguished, that of the streptococcal WalK and that of all other WalK proteins. The former appears to be an N-terminal truncation of the full-length WalK protein. The full-length protein is ~600 amino acids large and features two transmembrane helices separated by an extracytoplasmic domain. No structure of this domain is available, but fold prediction programs unambiguously assign this sequence stretch to fold as a PAS domain (Santelli et al., 2007; Chang et al., 2010). PAS domains are very versatile lignad binding modules and are utilized by a large number of signalling proteins across all kingdoms of life (Moglich et al., 2009). The N-terminal transmembrane helix and the extracytoplasmic PAS domains are absent in streptococcal WalK proteins indicating that this protein might respond to different signals. The remainder of the protein architecture is identical in all WalK proteins. It features a cytoplasmic HAMP domain as an immediate extension of a second transmembrane helix, followed by a cytoplasmic PAS domain and by the catalytic kinase core. HAMP domains are common structural features in transmembrane signal transduction proteins with a perceived function in transmitting a conformational change from one cellular compartment to another (Hulko
et al., 2006; Szurmant et al., 2007c). The catalytic core comprises the HisKA domain,
which harbours the site of autophosphorylation and the ATP-binding HATPase_c domain (Szurmant et al., 2007c).
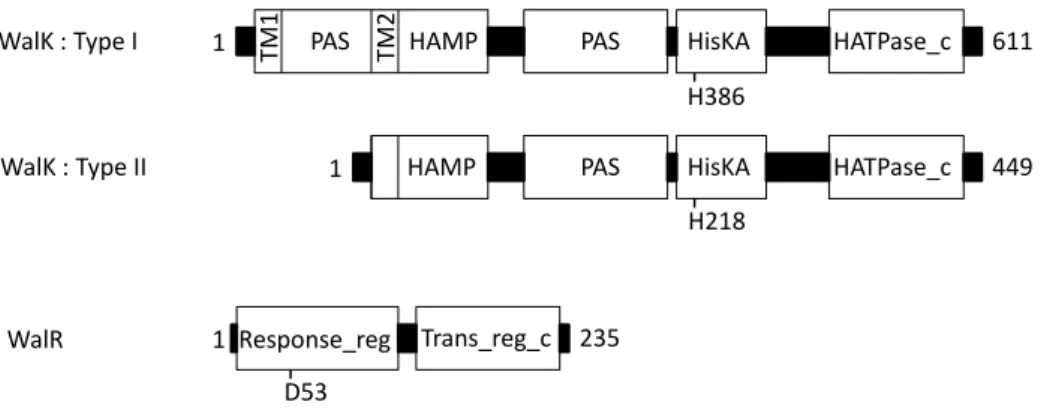
Fig. 2. Domain architectures for WalK and WalR proteins.
Two architectures have been recognized for WalK kinases. Type I is the prevalent type and includes two transmembrane helices (TM1, TM2), an extracytoplasmic PAS domain, cytoplasmic HAMP and PAS domains and the catalytic core comprising HisKA and HATPase_c domains. Type II WalK kinases are found in the Streptococcaceae and lack N-terminal TM1 and the extracytoplasmic domain. The amino acid lengths and the autophosphorylation site histidine are indicated for the representative WalK from B.
subtilis (type I) and from S. pneumoniae (type II). The WalR RR is an OmpR-type
regulator with a response_reg domain and a winged helix-turn-helix Trans_reg_c domain. The amino acid length and the aspartate site of phosphorylation are indicated for the representative WalR protein from B. subtilis.
The whole WalK crystal structure has yet to be resolved, however, a three-dimensional model of the conserved ATP-binding subdomain of Staphylococcus
epidermidis WalK was constructed in silico and used in a structure-based virtual screen
for specific inhibitory compounds (Qin et al., 2006).
Output: the WalR regulon
To rapidly gain significant insight into the role of the WalRK signalling system and to explain why this system was essential for viability it was important to define the genes whose expression was modulated by the WalR transcription regulator, also known as the WalR regulon. The typical approach to identify a regulon for a transcription factors is the measuring of global expression changes between a wild type and the transcription factor deletion strain by transcriptome analysis. The essential nature of the WalRK system dictated the design of slightly more sophisticated strategies. Approaches ranged from the generation of hybrid proteins to strains depleted for the WalRK system and in silico approaches assuming similar binding sites across different species of bacteria. These approaches were supplemented with in vitro gel shift and DNAase I foot
PAS
TM1 TM2 HAMP PAS HisKA HATPase_c
WalK : Type I 611
HAMP PAS HisKA HATPase_c
WalK : Type II 449 1 1 Response_reg Trans_reg_c 1 235 WalR H218 H386 D53
printing assays, and systematic progress was made resulting in the identification of genes directly regulated by the WalRK system. To date, the regulon of WalR has been studied in some detail in three organisms, B. subtilis, S. pneumoniae and S. aureus (Table 2). The consensus that has emerged from these studies is a global theme in the regulation of genes whose products are involved in cell wall homeostasis and cell wall restructuring (Dubrac et al., 2008). Perhaps somewhat surprisingly though, the regulons across the three different organism have proven to be divers, possibly reflecting changing requirements on cell wall restructuring based on cell size, shape and living environments.
TABLE 2. List of experimentally validated genes under direct regulation by the WalR transcription factor
Organism Gene/operon Function Regulation Evidence Reference
B. subtilis ftsAZ Cell division + GS, TR, FP Fukuchi et al. (2000), Howell et al. (2003) yocH CW hydrolase + MA, NA, FP, Howell et al. (2003)
GS
ykvT CW hydrolase NK FP Howell et al. (2003)
lytE CW hydrolase + NA, GS Bisicchia et al. (2007) ydjM CW associated protein + MA, NA, GS Bisicchia et al. (2007) yvcE (cwlO) CW endopeptidase + MA, NA, GS Bisicchia et al. (2007) tagAB, tagDEF Teichoic acid biosynthesis – FP Howell et al. (2003) yoeB Autolysin inhibitor – MA, NA, GS Bisicchia et al. (2007) yjeA Peptidoglycan deacetylase – MA, NA, GS Bisicchia et al. (2007) S. aureus ssaA CW amidase + GS, FP Dubrac and Msadek (2004)
isaA CW transglycosylase + FP Dubrac and Msadek (2004) lytM CW endopeptidase + FP Dubrac and Msadek (2004) S. pneumoniae pcsB CW amidase + MA, GS, FP Ng et al. (2003, 2005)
pspA Virulence + MA, GS, FP Ng et al. (2005) lytN CW hydrolase + MA, GS, FP Ng et al. (2005) spr1875 CW binding/unknown + MA, GS, FP Ng et al. (2005)
CW, cell wall; +, positive regulation by WalR; –, negative reulation by WalR; NK, not known; FP, DNase I foot printing; GS, gel shift mobility assay; MA, microarray; TR, transcriptional reporter; NA, Northern analysis.
Input: the WalK signal
In order to truly understand the role an individual signal transduction system plays in bacterial physiology, it is not sufficient to know the genes whose expression is
regulated by the transcription factor. Rather knowledge on the molecular signals that regulate the activity of the sensor kinase is required to comprehend the purpose of the response. Unfortunately, the identification of an input signal is not as straightforward as the identification of the output regulon. Even for the best-studied signalling systems the activation signal has often remained elusive. Commonly, growth conditions are known that alter the activity of individual sensor kinases in vivo, but small molecular ligands have been identified for only a handful of signalling systems. Based on these isolated cases, the assumption is that most TCS sensor kinases modulate their kinase activity in response to a specific small molecular ligand.
No such molecular ligand has to date been identified that modulates the activity of the WalK sensor HK. Some reports have suggested that WalK might directly sense molecular oxygen or redox potential. Such reports, however, are commonly based on the observation that the WalK kinase contains a cytoplasmic PAS domain and rely on phenotypic evidence that supports a function in redox or oxygen sensing. PAS domains were originally associated with a role in oxygen or redox response in bacteria (Zhulin et
al., 1997). With the accumulation of sequenced genomes it has become clear that the
PAS domain is a very versatile ligand-binding domain and that it can accommodate and respond to a large variety of molecular signals (Moglich et al., 2009). In addition, PAS domains were shown to be involved in protein-protein interactions. It thus appears ill advised to suggest that a protein is directly involved in redox sensing based in large part on the observation that it contains such a domain.
In the lieu of structural information of the WalK sensing domain, which may provide direct insight in the signals perceived, several approaches have been undertaken in order to identify the input of the WalK system. As mentioned previously a transposon
mutagenesis screen using a WalRH215P temperature sensitive strain aimed to identify
mutations in genes that result in the activation of the WalRK system. This study revealed that YycH and YycI proteins serve to inactivate the WalK kinase (Szurmant et
al., 2007b). The molecular details of this process have been addressed and resulted in
the picture that WalK kinase forms a ternary complex with YycH and YycI proteins of reduced kinase activity (Szurmant et al., 2007b, 2008; Fukushima et al., 2011).
An immunofluorescence based approach was undertaken for B. subtilis to identify the subcellular localization of the WalK kinase expecting that such information might reveal insights as to how, when and where the WalK kinase receives its activation signal (Szurmant et al., 2007a; Fukushima et al., 2008). Fortunately, the WalK localization pattern turned out to be insightful, although such studies with other sensor kinases in other organisms frequently were disappointing, since the proteins did not localize to any
specific compartment in the cell. In contrast, the WalK kinase specifically localized to the septum in rapidly dividing cells. This septum localization was dependent on the master regulator of cell division, FtsZ. In a strain depleted for ftsZ, WalK no longer localized to the septum. Tested by semi-quantitative RT-PCR, depletion of either ftsZ or the walRK operon had equivalent effects on the expression of the WalR-dependent genes, suggesting that WalK requires a properly assembled divisome for localization and activation (Fukushima et al., 2008). In support of this notion, immunofluorescence studies on the WalR RR showed that it co-localizes with DNA in rapidly dividing cells indicating a high level of phosphorylation. In contrast, in ftsZ depleted cells WalR no longer exclusively localized to the DNA, suggesting a reduced level of phosphorylation.
Although these studies failed to identify a molecular ligand, they did identify the apparently essential purpose that the WalRK system serves in bacterial physiology. By combining knowledge on the activation of WalK at division sites with knowledge about the WalR regulon, a picture emerges where the WalRK system serves to coordinate cell wall restructuring with cell division (Fukushima et al., 2008). Under actively dividing conditions, the WalK kinase localizes to division sites, where it is activated and induces the expression of genes whose products are required for cell wall lysis and restructuring. These activities are most certainly unwelcome under non-dividing conditions. Under these conditions, WalK no longer localizes to division sites and is not activated, resulting in the de-repression of inhibitors of the enzymes required for cell wall restructuring. The WalRK system thus represents a growth-dependent regulator of cell wall homeostasis, at least in B. subtilis.
A subsequent study aimed at identifying the molecular factors that direct the WalK kinase to the septum and those that are involved in its activation (Fukushima et al., 2011). This problem was addressed by two approaches: On the one hand, WalK kinase constructs were expressed in B. subtilis that harboured deletions of individual or multiple domains. When testing activity and localization of the various constructs it became apparent that constructs deleted for any or all of the N-terminal domains down to the cytoplasmic PAS domain were constitutively active and no longer associated with the YycH and YycI proteins. Nevertheless, these WalK constructs retained the ability to localize to the septum, thus leaving only the three cytoplasmic domains, the PAS domain or the catalytic domains, as possible interaction partners with the divisome proteins. The isolated PAS domain as well as a catalytic fragment of WalK proved to be highly unstable in vivo. Still, experiments with the latter suggested that the PAS domain is the determinant for septum localization.
restructuring, it was suggested that the WalK kinase might sense specific cell wall determinants (Dubrac et al., 2008). For B. subtilis a detailed analysis of changes in expression of the WalR regulon in response to sublethal concentrations of several cell wall acting antibiotics suggested an involvement of the D-Ala-D-Ala moiety in the peptidoglycan precursor lipid II as a possible WalK activation signal. This hypothesis was tested in a study where the E. faecalis vancomycin resistance cassette was introduced into B. subtilis (Bisicchia et al., 2011). The genes encoded by this cassette catalyse the replacement of the D-Ala-D-Ala depsipeptide with D-Ala-D-Lac. The latter has a reduced affinity for Vancomycin resulting in resistance. In exponentially growing cells no changes in the expression of the WalR regulon were observed upon introduction of the Vancomycin resistance cassette making an involvement of the D-Ala-D-Ala depsipeptide as a possible WalK signal unlikely.
Role in antibiotic resistance
Methicillin-resistant S. aureus (MRSA) are an increasingly relevant threat to human health. Infections are generally treated with last line of defence antibiotics such as vancomycin and daptomycin. In recent years clinical isolates showing reduced or no susceptibility to either of these antibiotics have emerged following failed drug treatments. A large number of such isolates have now been genome sequenced to identify the genetic determinants responsible for lack of susceptibility by comparative genomics.
Vancomycin is a glycopeptide antibiotic produced by Amycolatopsis orientalis that exerts its function by binding to the D-Ala-D-Ala moiety of lipid II and by inhibiting peptidoglycan polymer biosynthesis and cross-linking. In contrast, daptomycin is an acidic cyclic lipopeptide antibiotic produced by Streptomyces roseosporus and is believed to act primarily by inserting in and depolarizing the cytoplasmic membrane. Despite these entirely different modes of action, mutations in the walRK system have been linked to non-susceptibility to either antibiotic following drug treatment.
Regarding vancomycin non-susceptible S. aureus, also known as VISA (for
vancomyicn-intermediate S. aureus), initially two mutations had been reported to promote Vancomycin resistance. A strain was found that carried an insertion mutation in the yyc promoter resulting in vast overexpression of the entire yyc operon (Jansen et al., 2007). Another isolate was described featuring a mutation in the yycH gene that introduced a premature stop codon (Mwangi et al., 2007). Such a mutation was suspected to result in an overactive unregulated WalRK system in analogy to the
function YycH plays in B. subtilis (Szurmant et al., 2007b). Both reports thus suggested that an overactive WalRK system might have contributed to the VISA phenotype. Mutations originally described in daptomycin-resistant isolates included point mutations in WalK (S221P and R263C) (Friedman et al., 2006). Whether these mutations increased or reduced the activity of WalK or had an entirely different effect remains unknown.
A recent comprehensive report identified distinct mutations in either WalR or WalK
in 10 out of 13 additional clinical isolates from patients following failed Vancomycin treatment (Howden et al., 2011). Subsequent genetic analysis demonstrated that these mutations were indeed responsible for the VISA phenotype. Individual mutations were not localized to any specific domain. WalR mutants harboured point mutations in either the RR domain (A96T) or in the DNA-binding domain (K208R). Mutations in WalK occurred in the N-terminal transmembrane helix (L14F), the HAMP domain (G223D), the PAS domain (V268F; D337-339+I340V), the HisKA domain (N382S) or the HATPase_c domain (D469-70; A567D; T595I). Another equally comprehensive study found mutations in WalK in 24 out of 39 clinical VISA isolates, making walK the most prevalent gene to feature a mutation (Shoji et al., 2011). In addition, the mutations found in this study were similarly diverse to the above described set of mutations and included point mutations and amino acid insertions dispersed across all domains of WalK.
Both studies documented phenotypes and transcriptional profiles for several of the
WalK mutant strains and independently observed similar effects. The mutated strains featured a reduced autolysis activity. Microscopic analysis revealed a thicker cell wall, a phenotype that was already known to be associated with the VISA phenotype (Howden
et al., 2011; Shoji et al., 2011). Microarray analysis demonstrated global changes in the
expression of a large number of metabolic proteins, perhaps a secondary effect due to the drastic changes in cell wall metabolism.
Somewhat troubling, many of the walK and walR mutations studied by Howden and coworkers also promoted non-susceptibility to daptomycin, despite that fact that these strains had never been exposed to daptomycin (Howden et al., 2011). This study thus conclusively demonstrated that identical mutations in the WalRK system can promote cross-resistance to the most commonly used antibiotics against MRSA infections, despite the distinct modes of action of these two antibiotics. It remains unclear whether all the diverse mutations exert similar effects on WalRK activity. The observed phenotypes of reduced autolytic activity and thickened cell wall are more consistent with reduced WalRK activity, which is in contrast to earlier studies that
suggested an overactive WalRK system to be responsible for the VISA phenotype (Howden et al., 2011; Shoji et al., 2011). Additional studies are clearly warranted to
clarify the effects of these mutations on WalRK activity.
Drug discovery efforts
TCSs have long been recognized as potential broad range targets for antimicrobials. An original idea was that drugs that specifically target TCS chemistry should show little toxicity to humans, thanks to the absence of similar chemistry in the mammalian protein complement. Before the identification of essential systems, it was suggested that such drugs should render a microbe unresponsive to its environment and thereby attenuate its virulence and ability to survive in a host organism. Unfortunately, broad range inhibitors with specificity towards two-component chemistry have not been identified. Instead, most inhibitors proved to be semi-specific protein denaturing agents and often had haemolytic activity (Stephenson et al., 2000).
The discovery of the essential WalRK TCS and its wide distribution across the phylum Firmicutes fuelled several targeted approaches to discover specific inhibitors for the WalK kinase or the WalR RR. Such compounds can be expected to serve as specific antimicrobial drugs against important human health threats such as S. aureus, E.
faecalis or the streptococci. Several reports outlined below have described the discovery
of WalK kinase and WalR inhibitors with bactericidal activities. None of the described inhibitors has passed clinical trials and we are thus still far away from the therapeutic use of any of the reported compounds or their derivatives. Nevertheless, the ongoing efforts suggest that targeting the WalRK system remains a valuable approach and that an effective antimicrobial drug might emerge in the future.
WalK kinase inhibitors
A screen to identify natural compounds from microbial acetone extracts with specific activity towards the WalRK system was designed based on a B. subtilis strain
carrying the temperature sensitive WalRH215P mutation (Watanabe et al., 2003). It was
argued that any compound with increased antimicrobial activity against the temperature sensitive strain over the parental wild type strain might specifically target the WalRK system. From seven acetone extracts that followed this notion, a single compound was extracted, aranorosinol B, which was already known to exhibit bactericidal activity against some Gram-positives. In vitro analysis of this compound demonstrated that it
inhibited WalK kinase activity at relatively high concentrations. Utilizing this same screen along with methanol extracts from another 1368 microbes resulted in the isolation and identification of a single compound with broad antimicrobial activity against low G+C Gram-positives with a maximal inhibitory concentration below 1 µg/ml (Okada et al., 2010). Consistent with the notion that this compound specifically targets the WalRK system, B. subtilis phenotypes observed upon exposure to this compound matched those that were observed upon depletion of the WalRK system. These phenotypes included cellular filamentation and the emergence of cells that had lost their cytoplasmic content (Ghost-cells) (Fabret and Hoch, 1998; Okada et al., 2010).
An in vitro analysis demonstrated that this compound directly binds to WalK with a Kd
in the low micromolar range and that it inhibits WalK autophosphorylation with a
similar IC50. It remains to be seen whether this compound or its derivatives might serve
as a new antimicrobial, but these initial studies look promising. In 2006, a virtual drug screen was described that utilized a homology-modelled structure of the WalK HATPase_c domain of S. epidermidis as a target along with an in silico database consisting of 85,000 compounds (Qin et al., 2006). 76 potential hit compounds were tested for their ability to inhibit S. epidermidis growth at a concentration of 200 µM. By this approach seven compounds could be singled out. These compounds were shown to inhibit bacterial growth of several Gram-positive strains, but none of them had any inhibitory effect against Gram-negatives. In vitro assays suggested that some of these compounds have a high affinity for the WalK kinase and inhibit its autophosphorylation activity. Some specificity could be demonstrated when these compounds were tested against another kinase, S. epidermidis SrrB, with no or reduced inhibitory effect against this kinase. Preliminary tests using a Vero cell line and human erythrocytes ruled out general cytotoxic or haemolytic effects on eukaryotic cells for most of these compounds. Despite these promising results no follow up reports have surfaced that suggest that any of these compounds are being developed as antimicrobial drugs.
WalR response regulator inhibitors
Given the fact that the WalK kinase is not essential in the streptococci and that cross-phosphorylation of response regulators can be acquired relatively easily suggests that the WalR RR represents a better target for antimicrobials than the WalK kinase. Despite this fact the first attempt at specifically targeting the WalR protein has only recently been reported.
WalR homodimerization. In this screen the WalR RR domain was fused to a DNA-binding domain of the Thermotoga maritima transcriptional repressor IclR. IclR is known to act as a repressor in its dimeric form (Zhang et al., 2002). By replacing its dimerization domain with the RR domain of either WalR or of several other RRs, an in
vivo screen was devised that extracted compounds that specifically inhibited
dimerization of the WalR-IclR hybrid, but not of the other RR–IclR hybrids. The hybrid proteins were expressed in E. coli in the presence of the eGFP gene under the control of an IclR responsive promoter. It was shown that all RR–IclR fusion proteins inhibited eGFP expression, in contrast to a negative control strain harbouring the isolated IclR DNA-binding domain.
Upon screening an extensive chemical library the authors identified two compounds, named walrycin A and walrycin B, that inhibited the activity of WalR-IclR but not of the ArcA-IclR and OmpR–IclR hybrid proteins. Subsequent assays attempted to prove their specificity for WalR by their ability (1) to inhibit phosphorylation by WalK in vitro, (2) to inhibit expression of WalK responsive genes in S. aureus and B.
subtilis as determined by quantitative RT-PCR, (3) to inhibit bacterial growth of both
wild-type and WalR-overexpressing strains of S. aureus and B. subtilis and (4) to affect WalR homodimerization. These assays in their entity suggested that the two compounds target WalR specifically. The data indicated that rather than preventing WalR dimerization the compounds might act by promoting dimerization of a nonnative and inactive conformation (Gotoh et al., 2010). Again, while promising, it remains to be seen whether these compounds or their derivatives might be useful as antimicrobials or whether they prove to be toxic or ineffective.
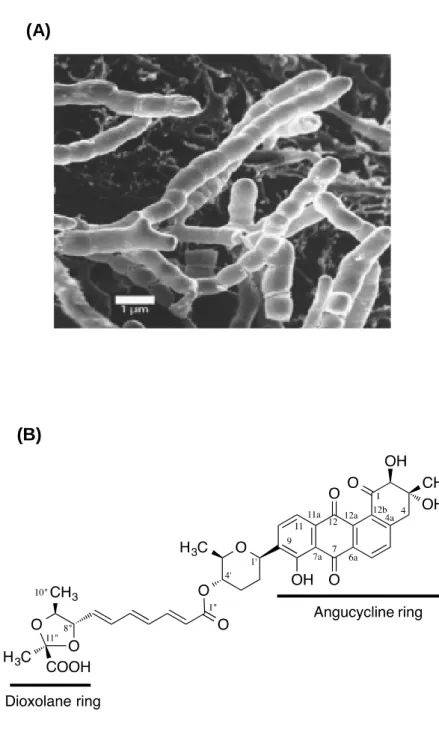
In this study, we have focused on the WalK inhibitor, waldiomycin (Fig. 3A and B) which we discovered a new WalK inhibitor, by screening metabolites from actinomycetes (Igarashi et al., 2013). This inhibitor belongs to the family of angucycline antibiotics and is structurally related to dioxamycin. Here, we have employed in vivo and in vitro strategies to further characterize the novel WalK-histidine kinase inhibitor, waldiomycin.
(A)
(B)
CHAPTER II
In vitro mode of action.
Introduction
The rapid emergence of antibiotic resistance in pathogenic bacteria has emphasized the need for an accelerated approach to the discovery of new antibacterial agents. Bacterial genomics, bioinformatics and gene manipulation studies have led to the discovery of novel protein targets for antibacterial agents (Moir et al., 1999). The TCSs of bacteria have received increasing attention for their potential as novel antibacterial drug targets for the following reasons (Barrette and Hoch, 1998; Macielag and Goldschmidt, 2000; Matsushita and Janda, 2002). First, TCSs are essential for coordinated expression of stress-response genes including those for virulence factors. Second, TCSs regulate the expression of antibiotic resistance determinants including drug-efflux pumps (Hirakawa et al., 2003; Kato et al., 2000; Eguchi et al., 2003). Third, protein-histidine phosphorylation in the signal transduction pathway in bacteria is distinct from serine/threonine and tyrosine phosphorylation in higher eukaryotes. Finally, the high degree of structural homology in the catalytic domain of HKs and in the receiver domain of RRs suggests that multiple TCSs within a single bacterium could be inhibited simultaneously, potentially leading to a lower frequency of drug-resistant bacteria. Recently, a number of TCS autophosphorylation inhibitors with inhibitory activity against multi-drug resistant bacteria have been developed (Barrette and Hoch, 1998; Macielag and Goldschmidt, 2000; Matsushita and Janda, 2002; Barrette et al., 1998; Yamamoto et al., 2000).
The majority of bacteria use a phosphotransfer mechanism termed a TCS,
comprised of a HK and RR, to sense environmental conditions and bring about appropriate changes in cellular behavior (Stock et al., 1988; Stock et al., 2000; West and Stock, 2001). TCSs are important in regulating the virulence and propagation of pathogenic bacteria (Barrett and Hoch, 1998; Stephenson and Hoch, 2002; Stephenson and Hoch, 2002). This is a fundamental system of bacterial response to environmental stresses in both Gram-positive and Gram-negative bacteria, and most TCSs are not necessarily essential for growth. However, a small number of TCS-encoding genes have been found to be essential; for instance, a set of WalK (HK) and WalR (RR) is the essential TCS in B. subtilis and S. aureus. Bacterial two-component regulatory systems play a pivotal role in the process of infection. Interest in these systems as novel bacterial targets has been rekindled by the recent discovery of several essential systems in
important Gram-positive and Gram-negative pathogens. Several series of TCS inhibitors derived from broad screening approaches have been reported in the literature, however, most appear to suffer from poor selectivity, excessive protein binding and/or limited bioavailability. Consequently, pharmaceutical chemists have turned to alternate strategies, such as the design of substrate-based inhibitors, the generation of combinatorial libraries and the isolation of natural products, to identify inhibitors with more desirable properties. Although TCSs have been known since 1986, the full range of biochemical and physiological processes under their control has only recently been recognised, with the advent of bacterial genomic analysis and follow-up gene inactivation studies.
Assays to screen for inhibitors of TCS have been based either on the biochemical reactions carried out by HKs and their cognate RRs, or on the ability of the latter to activate expression of particular genes. Biochemical screens for inhibitors have measured the ability of agents to interfere with: i) Autophosphorylation of a HK ii) Transfer of the phosphate group from a HK to the cognate RR iii) Both of these activities. From the initial search of the inhibitors targeting the WalK (C-terminal cytoplasmic HK region) of B. subtilis, the inhibitors found were imidazole derivatives (e. g. NH125) (Yamamoto et al., 2001). Imidazole derivatives had antibacterial activity in drug-resistant S. aureus, E. faecalis, and S. pneumoniae as well as B. subtilis with
MICs of 0.39-6.25 µg ml-1. However, the inhibitors have caused structural alteration of
HK leading non-specifically to aggregation.
The specific hypersensitivities shown by temperature-sensitive mutants indicate that use of these mutants in whole-cell screening provides a rapid method to develop target-specific screens for the identification of novel compounds (Numata et al., 1986; Kamogashira et al., 1988). Previously we developed a differential growth assay targeting WalK/WalR two-component signal transduction system, in which a temperature-sensitive walR mutant (CNM2000) of B. subtilis was supersensitive to inhibitors of HK, in comparison with wild-type 168 strain (Watanabe et al., 2003; Okada et al., 2007). As a result, an antibacterial agent, aranorosinol B, was isolated as an HK inhibitor by this differential growth assay (Watanabe et al., 2003). Using this screening method, we isolated three new inhibitors; walkmycin B (Okada et al., 2010), signermycin B (Watanabe et al., 2012) and waldiomycin (Igarashi et al., 2013).
The emergence and spread of hospital acquired multi-drug resistant bacteria present a need for new antibiotics with innovative mode of action. Advances in molecular microbiology and genomics have led to the identification of numerous bacterial genes coding for proteins that could potentially serve as targets for
antibacterial compounds. Since signal transduction in mammals occurs by a different mechanism, inhibition of HKs could be a potential target for antimicrobial agents.
In this chapter, I have employed in vitro strategies to examine the inhibitory effects
of waldiomycin against WalK sensor HKs which are the key elements in
two-component signal transduction systems.
Materials and Methods
Bacterial strains and plasmids
Bacterial strains and plasmids used in this study are listed in Table 1. B. subtilis
and Escherichia coli cells were grown under aerobic conditions at 37⁰C in
Luria-Bertani (LB) medium (1% polypeptone, 0.5% yeast extract, and 0.5% NaCl) with antibiotic added when required. S. aureus cells were grown in Trypticase Soy Broth (TSB) medium (1.7% Bacto tryptone, 0.3% Bacto soytone, 0.5% NaCl, 0.25%
dipotassium phosphate, and 0.25% dextrose) at 37⁰C with aeration.
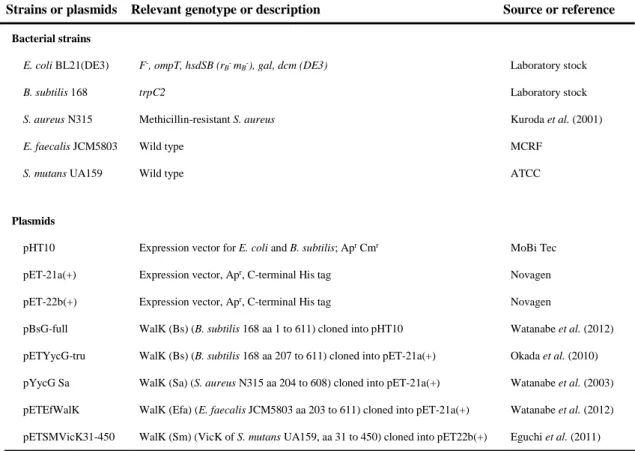
TABLE 3. Bacterial strains and plasmids used in this study
Strains or plasmids Relevant genotype or description Source or reference Bacterial strains
E. coli BL21(DE3) F-, ompT, hsdSB (r
B- mB-), gal, dcm (DE3) Laboratory stock
B. subtilis 168 trpC2 Laboratory stock
S. aureus N315 Methicillin-resistant S. aureus Kuroda et al. (2001)
E. faecalis JCM5803 Wild type MCRF
S. mutans UA159 Wild type ATCC
Plasmids
pHT10 Expression vector for E. coli and B. subtilis; Apr Cmr MoBi Tec
pET-21a(+) Expression vector, Apr, C-terminal His tag Novagen
pET-22b(+) Expression vector, Apr, C-terminal His tag Novagen
pBsG-full WalK (Bs) (B. subtilis 168 aa 1 to 611) cloned into pHT10 Watanabe et al. (2012) pETYycG-tru WalK (Bs) (B. subtilis 168 aa 207 to 611) cloned into pET-21a(+) Okada et al. (2010) pYycG Sa WalK (Sa) (S. aureus N315 aa 204 to 608) cloned into pET-21a(+) Watanabe et al. (2003) pETEfWalK WalK (Efa) (E. faecalis JCM5803 aa 203 to 611) cloned into pET-21a(+) Watanabe et al. (2012) pETSMVicK31-450 WalK (Sm) (VicK of S. mutans UA159, aa 31 to 450) cloned into pET22b(+) Eguchi et al. (2011) MCRF, Microbial Chemistry Research Foundation; Apr, ampicillin resistance; Cmr, chloramphenicol resistance.
Protein purification
E. coli BL21(DE3) cells containing the pETSa315VR, pETYycG-tru, pYycG Sa,
pETSMVicK31-450 and pETEfWalK plasmids were grown in 2x YT medium at 30⁰C
with aeration. When the cell density reached at OD600 of 0.5, IPTG was added at the
final concentration of 1 mM, and the cell suspension was further cultivated for 3 hrs before the cells were harvested by centrifugation, washed with lysis buffer (50 mM
Tris-HCl [pH 8.0], 100 mM NaCl) and then stored at -80⁰C until use. For purification of
His-tagged proteins, frozen cells were resuspended in lysis buffer with 1 mM phenylmethylsulphonyl fluoride and then lysed by sonication. The lysates were cleared
by centrifugation at 10,000 rpm for 10 min at 4⁰C. The supernatant was further
centrifuged at 14,000 rpm for 10 min at 4⁰C and affinity purified by Ni (II)-NTA
agarose (Qiagen, Los Angeles, CA). Eluted protein samples were then dialyzed against storage buffer (10 mM Tris-HCl [pH8.0], 0.1 mM EDTA, 0.1 mM dithiothreitol and
10% [v/v] glycerol) and stored at -20⁰C. Protein concentration was determined by using
the Bradford method and the purity was checked by SDS-PAGE. The cytoplasmic WalK domains (with C-terminal His tag) of Gram-positive B. subtilis, E. faecalis, and S.
mutans were expressed and purified as described previously using E. coli BL21 (DE3)
containing pETYycG-tru (Okada et al., 2010), pETEfWalK (Watanabe et al., 2012), and pETSMVicK31-450 (Eguchi et al., 2011), respectively.
Autophosphorylation and phosphotransfer assays
Purified HKs were autophosphorylated by incubation at 30⁰C in 50 mM Tris-HCl
(pH 8.5), 100 mM KCl, 100 mM NH4Cl, 5 mM MgCl2, 12.5 µM ATP solution (2.5 µM
ATP, and 16.7 nM [γ-32P] ATP) in a total volume of 10 µl. The reaction was terminated
by adding an SDS-PAGE sample buffer {300 mM Tris-HCl (pH 8.0), 10% SDS, 25% β-mercaptoethanol, 50% glycerol, 0.25% bromophenol blue}. For phosphotransfer assay,
cognate RRs of HKs were added to the autophosphorylated HKs and incubated at 30⁰C,
and the reaction was terminated by the SDS-PAGE sample buffer. These samples were then separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). The gel was then dried, exposed onto an imaging plate, and analyzed with FLA-7000, a laser scanner optimized for quantitative phosphorimaging (Fuji Film,
Inhibition of autophosphorylaion activity of WalK HK
The purified WalK cytoplasmic domains were preincubated in kinase buffer {50
mM Tris-HCl (pH 8.5), 100 mM KCl, 100 mM NH4Cl, 5 mM MgCl2} at 30⁰C, as
previously reported (Eguchi et al., 2011; Okada et al., 2007, 2010; Watanabe et al., 2003). Then 1 µl of drug solution was added to the mixture. After a 5-min incubation,
ATP solution (2.5 µM ATP and 16.7 nM [γ-32P] ATP) was added, and the reaction
mixture was incubated for an additional 10 min. The reaction was stopped 10 min later using 5×sodium dodecyl sulfate (SDS) sample buffer {300 mM Tris-HCl (pH 8.0), 10% SDS, 25% β-mercaptoethanol, 50% glycerol, 0.25% bromophenol blue}. The samples were separated by SDS-PAGE, the gel was then dried, exposed to an imaging plate, and analyzed using FLA-7000 a laser scanner optimized for quantitative phosphorimaging (Fuji Film, Tokyo, Japan). The signal bands detected by FLA-7000 were analyzed by Multi Gauge version 3.0 software (Fuji Film, Tokyo, Japan), and Prism 4.1 (GraphPad Software, CA) was used to calculate the 50% inhibitory concentrations (IC50s).
Results and Discussion
Autophosphorylation and phosphotransfer activity of WalK HK
As shown in Fig. 4A, the purified WalK HK (S. aureus) became
autophosphorylated which is evidenced by the appearance of distinct signal bands. The band intensity increased depending on the incubation time and a relatively stronger band of WalK autophosphorylation was observed after 2 hrs of incubation. Next, the cognate RR was added to the autophosphorylated HK in order to confirm that the phosphorylated bands obtained in Fig. 4A were the HKs in concern. As shown in Fig. 4B, WalR RR was phosphorylated by their cognate HK, confirming that the purified sensor was indeed WalK. Compared to autophosphorylation, the phosphotransfer reaction proceeded very rapidly, and the phosphorylated RR bands appeared as early as 30 min after the addition of the RR to the autophosphorylated HK.
Fig. 4. Autophosphorylation and phosphotransfer of WalK histidine kinase.
(A) Autophosphorylation of WalK. The purified WalK HK was incubated with [γ-32P]
ATP, subjected to SDS-PAGE, and analyzed by autoradiography. (B) Phosphotransfer from HK to its cognate RR WalR. Histidine kinase was autophosphorylated prior to the addition of its cognate RR. After incubation, the reaction mixture was subjected to SDS-PAGE and analyzed by autoradiography.
Inhibitory effects of waldiomycin against WalK HKs
The inhibitory effect of waldiomycin against the autophosphorylation activity of the cytoplasmic regions of WalK was analyzed. Waldiomycin or an equivalent volume
of 100% DMSO as the diluents was incubated with the HKs for 5 min at 30⁰C before
initiating the autophosphorylation reaction. As shown in Fig. 5, the bands for the autophosphorylated WalK clearly disappeared in a dose-dependent manner. The half
maximal inhibitory concentrations (IC50s) against WalK HKs purified from B. subtilis, S.
aureus, E. faecalis, and S. mutans were 10.2, 8.8, 9.2, and 25.8 µM, respectively. These
results demonstrate that waldiomycin inhibits WalK HKs from different sources with similar efficacy, indicating that waldiomycin is a broad-acting general inhibitor of HK. Simocyclinones, a group of antibiotics structurally related to waldiomycin, are bifunctional antibiotics that inhibit bacterial DNA gyrase (Edwards et al., 2009). It has been shown that the aminocoumarin and angucycline moieties in simocyclinone are indispensable for potent inhibitory activity by interacting independently to the binding pocket of DNA gyrase. Similarly, the dioxolane carboxylic acid and angucycline moieties in waldiomycin are considered to be important for WalK HK inhibitory activity (Igarashi et al., 2013, Fig. 3B).
120 60 30 5 1 10 30 10 5 2 1 0.5 WalK-P WalK-P WalR-P min min (A) (B)
Fig. 5. Waldiomycin inhibits autophosphorylation activity of WalK HK.
Histidine kinase was treated with waldiomycin for 5 min before autophosphorylation reaction was initiated. The residual activity was calculated as the percentage (%) of activity after treatment with waldiomycin against the diluents-treated (100% DMSO) control (waldiomycin, 0 µg/ml). Error bars represent the standard errors of the mean (SEM) of three independent assays.
The results obtained from our in vitro study of the mode of action of waldiomycin demonstrate that waldiomycin targets essential WalK HK (Fig. 5) and inhibits autophosphorylation activity of this essential HK, thereby hindering the WalK/WalR signal transduction pathway involved in cell growth and division.
Waldiomycin (µg/ml) 0 0.9 1.9 3.9 7.8 15.6 31.3 62.5 125 P-WalK235-608 R es id u a l a ct iv it y ( % ) Log10[Waldiomycin (µg/ml)] 100 50 0 0
CHAPTER III
In vivo mode of action.
Introduction
The ability of a bacterial cell to respond to changing environmental conditions necessitates the presence of signal transducing systems that respond to specific cues and initiates stimulus-specific cellular responses, usually through differential gene expression. TCSs are versatile regulatory networks which are critical components involved in environmental sensing and adaptation of microbes to the dynamic outside environment (Stock et al., 2000). The spectrum of stimuli and the range of responses coordinated by TCSs are as broad as the environmental niches to which bacteria have adapted. The conspicuous absence of TCS proteins in mammalian genomes makes them interesting potential targets for the development of novel anti-bacterial drugs.
Staphylococcus aureus is a Gram-positive bacterium with low GC content
(Fermicutes) and grouped into Bacillus-Clostridium group on the basis of the 16S ribosomal DNA sequence. S. aureus widely inhabits on the skin surface and nasal cavity of warm-blooded animals. The organism produces many toxins and is highly efficient at overcoming antibiotic effectiveness. In 1961, it developed resistance to methicillin,
invalidating almost all antibiotics including the most potent β-lactams (Jevons, 1961).
Antimicrobial agents are used to treat patients infected with bacteria. However, treatment of such patients by antimicrobial agents often produces drug resistant bacteria. Multidrug resistant bacteria have emerged after use of multiple antimicrobial agents, and have spread all over the world. Infections with multidrug resistant bacteria are serious clinical problems nowadays. Patients infected with multidrug resistant bacteria such as methicillin-resistant Staphylococcus aureus (MRSA) and vancomycin-resistant enterococci (VRE) are difficult to treat because only a few antimicrobial agents are effective on such multidrug resistant bacteria. MRSA easily infect
immuno-compromised hosts. The continuing increase of hospital-and
community-associated MRSA infections highlights an urgent need for the development of alternative potent antibacterial agents (Grundmann et al., 2006; Kazakova et al., 2005; Rice, 2006). The ability of this organism to resist current antibiotic therapies and cause infection is partially due to the coordinated regulation of gene expression allowing the bacteria to survive in different stress conditions.
TCSs are important sensory units and allow microbial organisms to adapt to different niches, as well as play a significant role in pathogenesis and biofilm formation
for various bacterial species (Gross, 1993; Stock, 2000; Li, 2002; Novick, 2006). Therefore, interrupting these critical signalling pathways may provide an alternative strategy for the development of novel classes of preventive and/or therapeutic agents (Barrett and Hoch, 1998). Genome scanning has predicted 17 potential TCSs in S.
aureus, some of which have been reported to be regulators of bacterial virulence. The
AgrCA TCS is a well-known global regulatory system controlling numerous genes involved in S. aureus virulence, such as cytotoxin- and hemolysin-encoding genes, but other TCSs, such as ArlSR, SaeSR, VraSR, and SsrAB, have also been reported to play roles in S. aureus virulence (Benton et al., 2004; Kuroda et al., 2003; Novick, 2003).
Until now, vancomycin had remained the weapon of last resort, but the recent appearance of the vanA vancomycin resistance gene cluster in S.aureus highlights the growing threat this bacterium poses to human health and the urgent need to develop novel therapeutic approaches (Chang et al., 2003). It has been suggested that WalK or WalR may serve as potential targets for the development of novel antimicrobial agents (Fukushima et al., 2011; Szurmant et al., 2005; Turck and Bierbaum, 2012).
The WalK/WalR TCS has been found to be conserved in most of the Firmicutes but is apparently absent from the genomes of Gram-negative bacteria (Fabret et al., 1999; Ng and Winkler, 2004). Following the initial observation that this system was the only one of 34 two-component signal transduction systems required for cell survival by B. subtilis, the essentiality of orthologs has been confirmed for the important
Gram-positive pathogens S. aureus, S. pneumoniae and E. faecalis (Fabret et al.,
1999; Hancock and Perego, 2004; Martin et al., 1999; Ng et al., 2003). S. aureus is a prominent agent of nosocomial disease and has shown an unusual aptitude in acquiring resistance to antimicrobial agents. A search for essential novel antibiotic targets in S.
aureus revealed the existence of an essential two-component regulatory system similar
to the genes walRK from B. subtilis (Martin et al., 1999; Fabret and Hoch, 1998).
The WalR regulon in B. subtilis comprises proteins involved in cell wall metabolism and turn-over (Howell et al., 2003; Bisicchia et al., 2007). In S. aureus, a search using the Bacillus consensus WalR recognition site yielded 31 loci with a WalR binding motif (Dubrac and Msadek, 2004) among them nine proteins with proven autolytic function or similarity to autolysins, which are upregulated upon induction of walRK (Dubrac et al., 2007).
Recent studies have revealed conserved functions for WalK/WalR TCS, defining this signal transduction pathway as a crucial regulatory system for cell wall metabolism.
A combination of approaches, including the use of hybrid RRs, conditional mutants, or
genetic, phenotypic and transcriptomic analyses, has recently shown that WalKR controls cell wall metabolism in all of the bacteria studied. This TCS also plays a role in cell membrane synthesis/fluidity, cell division, exopolysaccharide synthesis and oxidative stress in some cases. As this essential TCS constitutes a highly attractive target for antimicrobial therapy, this aspect has also been the subject of much investigation for the design of specific WalK inhibitors as novel antimicrobial compounds. The WalKR system has attracted considerable interest recently, yielding insights into its physiological function, the nature of its essentiality and its potential
activating signal. This TCS has attracted considerable interest as a probable
antimicrobial target against low GC % Gram positive pathogens since it has been found to be essential for the viability in most of the organisms studied (Fabret and Hoch, 1998; Dubrac and Msadek, 2004; Ng et al., 2003; Senadheera et al., 2005; Gotoh et al., 2010).
The emergence of multi-drug resistant staphylococcal isolates, especially, MRSA, is generating enormous public health concern and highlights an urgent need for new, alternative agents for treating multi-drug-resistant pathogens. Previously we showed that waldiomycin inhibits WalK HKs from both of S. aureus and B. subtilis and shows the antimicrobial activity against MRSA as well as B. subtilis.
In this chapter, I described the in vivo effects of HK inhibitor, waldiomycin in B.
subtilis and S. aureus in order to further characterize this inhibitor by applying several in vivo strategies.
Materials and Methods
Bacterial strains, plasmids and growth conditions
Bacterial strains and plasmids used in this study are listed in Table 3. B. subtilis
cells were grown under aerobic conditions at 37⁰C in Luria-Bertani (LB) medium (1%
polypeptone, 0.5% yeast extract, and 0.5% NaCl) with antibiotic added when required.
S. aureus cells were grown in Trypticase Soy Broth (TSB) medium (1.7% Bacto
tryptone, 0.3% Bacto soytone, 0.5% NaCl, 0.25% dipotassium phosphate, and 0.25% dextrose) at 37⁰C with aeration.
Extraction of total RNA and complementary DNA (cDNA) synthesis
For quantitative reverse transcription PCRs (qRT-PCRs), the cells of S. aureus N315 and B. subtilis 168 were grown with aeration to an optical density at 660 nm
(OD660) of 0.3 at 37⁰C. After addition of waldiomycin, the cultures were incubated for
another 5 min. The cells were then harvested by centrifugation and immediately suspended in RNA-later (Ambion, Austin, TX). RNA was then purified with the SV Total RNA Isolation System (Promega, Madison, WI), followed by DNase I treatment (Turbo DNA-free kit; Ambion) to eliminate residual contamination of genomic DNA. The purity and concentration of the RNA were determined by spectrophotometry and gel electrophoresis. Reverse transcription reactions were carried out with the High-capacity cDNA Reverse Transcription Kit (ABI, Foster City, CA).
qRT-PCRs
PCR primers (Table 4) were designed with Primer Express Software Version 3.0 (ABI). Amplification of cDNA (1 ng) was carried out with 250 nM gene-specific primers and PowerSYBR Green Master Mix (ABI). PCR amplification, detection, and analyses were performed using the StepOne Real-Time PCR System (ABI). PCR
conditions included an initial denaturation step at 95⁰C for 10 min, followed by
40-cycle amplification (95⁰C for 15 sec and 60⁰C for 60 sec). The specificity of the
amplified product and absence of primer dimer formation were verified by generating a melting curve. Melting curve analysis was performed with an initial denaturation step of
95⁰C for 15 sec followed by 60⁰C for 1 min and 95⁰C for 15 sec. Fluorescence was
measured continually during the melting curve cycle (a stepwise 0.3⁰C temperature)
beginning at 60⁰C. The absence of contaminating genomic DNA was verified by testing
each sample in control reactions without a previous reverse transcription step. The critical threshold cycle (CT) was defined for each sample. The expression levels were