Protein II
Tertiary Structure of Proteins
- The combinations of secondary structures
- Arrange in space of all atoms in one polypeptide chain
o However, to draw a clear distinction difference is quite difficult - Two large classes
o Fibrous proteins o Globular proteins
Fibrous Proteins
- Polypeptide chains arranged parallel along the axis - Either in long fibres or large sheets
- Deals mainly with structures o Mechanically strong
o Insoluble in water and dilute salt solutions - Examples:
o Keratin – Hair and wool o Collagen
Main component of connective tissues in animal
Stress-bearing Collagen
Structures
- 3 polypeptide chains wrapped around each other o Ropelike twist
o Three coils form one coil (Triple helix) o Coils of the coils
o Known as tropocollagen
- They do not form α-helix but EACH POLYPEPTIDE CHAIN IS A HELIX WITH α-chain. Amino Acid content (X - Pro/Hyp - Gly) – X is any amino acids
- 30 % Proline and hydroxyproline - Hydroxylysine (Hyl)
- Every 3rd residue is glycine (Actually only glycine can fit in the CROWDED centre of the triple helix because GLYCINE has the SMALLEST structure)
Properties
- The polypeptide chains hold together by hydrogen bonding
- Covalent crosslinks between Lys and His residues will strengthen the structure o Crosslinks will increase with age
o Younger meat is better to be eaten
- If there is a failure in hydroxyproline (Hyp), a fragile collagen will result o Take in more vitamin C to synthesis Proline Hydroxyproline
Inner coil
Outer coil
Globular Proteins
- Polypeptide chains fold in spherical-like shape - Wide biological functions
o Enzymes, transport proteins, regulatory proteins, etc - Structure can be determined by
o X-ray crystallography (Using the concept of diffraction, particle wave) o Nuclear magnetic resonance (Determine 2-D structure)
- Water soluble
o Most Polar amino acids are folded on the surface of the globular protein, which allow the interacting of hydrogen bonding and ion-dipole interactions with aqueous environment o Most non-polar groups are folded inside the protein, minimizing contact of aqueous
environment
- Contains secondary structures o α-helix and β-sheet Super-secondary structure
- Known as motifs
o βαβ-units – 2 parallel strand connected with one α-helix o αα-unit – 2 antiparallel α-helices
o β-Meander - antiparallel β-sheets o Greek key
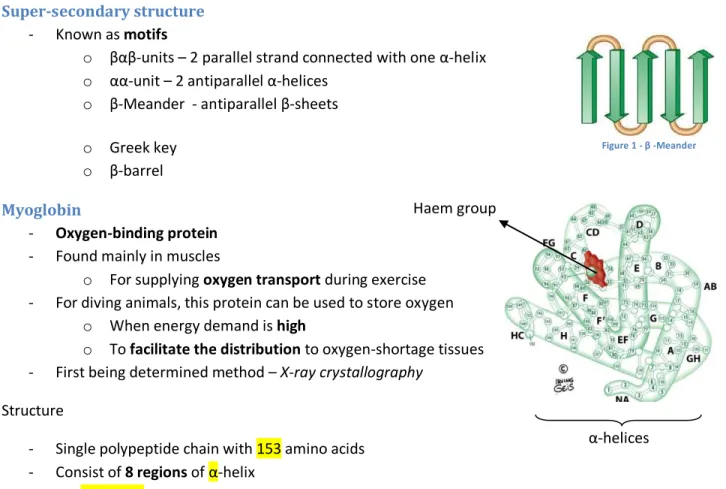
o β-barrel Myoglobin
- Oxygen-binding protein - Found mainly in muscles
o For supplying oxygen transport during exercise - For diving animals, this protein can be used to store oxygen
o When energy demand is high
o To facilitate the distribution to oxygen-shortage tissues - First being determined method – X-ray crystallography
Structure
- Single polypeptide chain with 153 amino acids - Consist of 8 regions of α-helix
- NO β-SHEETS
- Connected by bends and polar groups on the protein surface - The Haem group is buried in th interior of the protein
Figure 1 - β -Meander
Haem group
α-helices
Protein Folding and the Denaturation of Protein
- Protein folding include: o Hydrogen bonding
Between polar R Groups (Ser and Thr)
Hydrogen bond can form between two polar amino acids which causes folding o Hydrophobic interaction
Two Non-polar R groups will repel each other due to the hydrophobic properties of amino acids
Val and Ile o Electrostatic attraction
Between two opposite charged amino acids
Lys and Glu o Electrostatic repulsion
Between two same charged amino acids
Lys and Arg, Glu and Asp o Forming of disulphide bonds
2 Cysteine can form one Cystine by forming disulphide bond
o Association and packing of secondary structure to achieve most stable structure o This can have a variety of formation of tertiary structures
- When the structure order of the protein had lost. The protein starts to denature o Results in loss of biological activity
- Denaturation often occurs in:
o Non-covalent interactions because the bond energy is low o The disulphide bond is disrupted
- Ways to denature proteins o Heating
Increase of temperature = Increase vibration of molecules
Disrupt weak interaction in the tertiary structure o Large pH variation
This will disrupt the electrostatic interactions o Binding of detergents
Disrupt hydrophobic interactions
Sodium dodecyl sulphate o Chaotropic reagents
Urea, guanidine hydrochloride
They form stronger hydrogen bonding with the proteins compared to other
This will lead to affection of original hydrogen bonding
Disrupt hydrophobic interactions
- PRIMARY STRUCTURE of the protein is CRUCIAL in determining the TERTIARY STRUCTURE of a protein
Quaternary Structure
- Contains more than one polypeptide chains - Each set of polypeptide chain is called a subunit - Each subunit associate non-covalently most of the time
o Hydrogen bonds
o Hydrophobic interactions o Electrostatic interaction
- Sometimes, disulphide bond will be formed too
Reason of having one or more polypeptide chain
- Due to their non-covalent interactions, the small structural changes in one subunit will result in the drastic change in other subunits (ALLOSTERIC EFFECT(COOPERATIVITY))
o Haemoglobin – bind oxygen reversibly, exhibits positive cooperativity
When there is ONE O2 molecule bind in one subunit, it will be easier for the other subunits to be bound.
- Genetic economy and efficiency
o There will be using less genetic material to code for subunits o Each subunit of haemoglobin will be coded separately
o If there is one error in one subunit, it can be replaced with the CORRECT subunit INSTEAD OF CHANGING THE WHOLE PROTEIN
o This will result in smaller synthesis error - Functional efficiency
o There are many different reactions and transitions o Each subunit can perform one enzymatic reaction
o One quaternary structure of protein can catalyse different reactions
o Sometimes, one subunit will function in catalysis whereas another one is regulation
Haemoglobin
Main function: Oxygen transport
- Oxygen binds to the haemoglobin behave similarly with enzyme- substrate binding
o Including regulation of enzyme activity o Ho orary e zy e
- Four subunits : Tetramer
- 2 2 Conformation: Symmetrical o -chain: 141 amino acids o -chain: 146 amino acids
- One haemoglobin can bind to FOUR oxygen (It has 4 heme group) - Oxygen binding of haemoglobin is quite in sigmoidal curve
o This is because it has the positive cooperativity
Enzymes
- Biological catalyst
- All enzymes are protein EXCEPT for some RNAs
- The catalysis of enzyme involve the lower of activation energy (∆��≠) by providing an alternative pathway
o Free energy change still remains the same (∆��) o Increase in rate of reaction
- When there is a few transition states, the overall reaction rate will be determined by the step with the highest ∆��≠ ,i.e. the slowest transition states (This is known as the rate-limiting step) - Substrate binds to the active site of an enzyme by non-covalent forces - Formation of ES complex
Specificity of Enzyme
- Absolute specificity
o Catalyse only one unique substrate to a particular product - Relative specificity
o They can form structurally related substrate - Stereospecificity
o It will only bind to one stereoisomer of a substrate, not other
- Their high degree of specificity is due to the complimentarity between substrate and active site Models of Enzyme-Substrate binding
- Lock and key mechanism
o Binding only occurs when there is perfect complimentarity between enzyme and substrate o VERY RARE IN REALITY!
- Induced fit model
o Binding of substrates induces a change in conformation of enzyme resulting in a complimentary fit
Interactive process
Conformation change is induced
Enzyme-su strate affi ity fro ore pre ise fit between enzyme and substrate
Enzyme Kinetics
- We look at the initial velocity (�0) of the enzyme (Initial rate when ES formed)
- When the enzyme reached the plateau, the velocity is known as maximum velocity
- Michaelis Constant (��) o [S] when 12����
�0=
���� �
�� +�
1
�0
= ��
���� � + 1/����
Rate-limiting step
- Km can measure the affinity of enzyme for substrate
�� =�2+�−1
�1
- When k2 is rate-limiting (k2 is slower, �2 �−1) - However, when
o �2 �−1 and �2≈ �−1
o The Km does not measure the ES affinity anymore - ���� is related to turnover number
o kcat - measure of number of substrate converted to product by ONE enzyme molecule per unit time when the enzyme is fully saturated with substrate
- However, in our cells, [S] is seldom fully saturated
- Better way to measure enzyme efficiency under non-saturating condition o ����
��
o Second order rate constant for
E+S E+P reaction