ASCONE2010 渡辺正峰先生講義レポート
意識の神経メカニズムをめぐって (基礎編)
大 泉 匡 史
∗2,3理化学研究所脳科学総合研究センター脳数理研究チーム∗2,日本学術振興会特別研究員∗3
Masafumi Oizumi∗2,3
Laboratory for Mathematical Neuroscience, RIKEN Brain Science Institute∗2 Research Fellow of the Japan Society for the Promotion of Science∗3
本稿は2010 年 11 月に行われた Autumn School for Computational Neuroscience(ASCONE) に おける,渡辺正峰先生の講義「意識の神経メカニズムをめぐって(基礎編)」の講義レポートである.こ の講義は合宿の一番最初の講義として,現在までの意識研究に関する基礎的な知識を提供する目的を主 として行われた.講義の最後には,渡辺先生自身が提案する意識の神経モデルについての解説も行われ た.本稿ではこの講義の内容を踏まえて,現在までの意識研究の概観および渡辺先生が提案する意識モ デルに関して紹介する.
1. 温度計に意識はあるか?
「温度計に意識はあるか?」と聞かれたらほとんどの 人は即座に,ないと答えると思う.それどころか,そ のような質問を真面目な顔をして聞いてくる人がいた としたら,ひどく滑稽に思うかもしれない.それでは, 内部に温度センサがあって,温度が高い時には「今日 は暑いなー」等と言ってうちわであおぎ,温度が低い 時には「うー寒い」等と言ってこたつに入るロボット には意識があるだろうか?
単純に考えると意識があるということの意味とは, 自分が今どういう状態かが分かる,例えば今何を見て いるのか,暑いのか寒いのか,楽しいのか悲しいのか 等,こういったことが自分で分かっているということ だろう.それでは先ほどの温度センサを持ったロボッ トはどうかと考えると,暑いのか寒いのかという自分 の状態を自分で「分かっている」ように見える.わざわ ざ人間らしいロボットの例を持ち出さなくても,自分 の状態を監視するシステムを持った機械は現在の我々 の生活の中でもいくらでも例があるし,容易に作れて しまう.従って,「客観的な体験」として今自分が暑い のか寒いのかといったことを自分で分かっているとい う意味では我々人間と先ほどのロボットに差異はない.
それでは,こういった自分の状態を監視するシステ
∗2〒351–0198 埼玉県北和光市広沢 2–1
∗3
ムを内部に持った機械が,我々と同じように「主観的 な体験」として「暑い」と思ったり「寒い」と思ったり しているのだろうか?この問に対しては多くの人がそ んなことは有り得ないと言うだろうし,私自身もそう 思っている.しかし,なぜこのようなロボットに主観 的な体験がなくて人間にはあるのかと問われると,私 は明確な答えを持っていないことに気づく.
2. 意識のハードプロブレム
物質及び電気的・化学的反応の集合体に過ぎない脳 から,一体どのようにして主観的体験が生じ得るのか というこの問題こそが,Chalmersによって提唱された 意識のハードプロブレムと呼ばれる問題である
6)
.大 多数の人が一つの神経細胞が意識を持っていないとい うことには同意すると思うが,なぜそのような一つ一 つは意識のない単純素子が大勢集まることによって主 観的体験が生まれるのかという問題は,McGinnが言 うところの「水からワインが生じている」ようなもの に感じられる
15)
.
意識のハードプロブレムに対する人々の態度は様々 である.提唱者のChalmers自身は,現代の物理学の 範囲内では解決できない問題だととらえており,新た な物理法則が必要だと考えている.同様に,物理学者の
Penroseも正しく定式化された量子重力理論が必要で
あると主張している
17)
.さらには,Dennettのように そもそも意識というものは錯覚であり,意識のハード
プロブレムは疑似問題であると解釈するものもある
8)
. 3. 意識が生じるための必要条件とは?
新しい物理法則を模索するというのもそれはそれで 重要な研究かもしれないが,とりあえず現在我々が知っ ている自然法則の範囲内でできる実践的な研究はない のだろうか?その代表例の一つがCrickとKochが取る アプローチの仕方で,意識の神経相関,Neural Corre- late of Consiousness(NCC)を実験的手法により明ら かにするという方法である
4, 11)
.意識が神経細胞の活 動によって生まれているというのは疑いようのない事 実なのであるから,意識の神経相関をまず明らかにし ようという態度は極めて建設的でありかつ実践的ある. 彼らはこのアプローチに基づいて一次視覚野がNCC か否かといったような問題を研究している
5)
.もちろ ん,NCCが十分明らかになったとしても,意識のハー ドプロブレムが解けるかどうかは全く保証がないのだ が,現状ではNCCが何かといった問題すら分かって いないのだからまずここからスタートするというのは 健全な態度であると感じる.
本講義において渡辺先生が取る研究態度も,意識の ハードプロブレムは取り扱わずに現状で何ができるか を考えるというものである.渡辺先生から受講者に対 して出された課題は以下のようなものであった(原文 ママ).
本日のお題「NCCの少し上を目指して」
•意識のハードプロブレムに真っ向から挑む のは現段階では非常に困難である.そこで, これをとりあえず棚にあげ,現象的意識の 神経表象とそれを支える神経システムが満 たすべき条件について考察してください.
•余力があるなら,上記条件を満たすモデル を提案してください.
•さらに余力があるなら,上記モデルを検証 する実験を考えてください.
現時点では意識が生じるための「十分条件」を特定 することは困難であろうが(十分条件が分かるという ことは,原理的には意識のある機械を作ることができ るということを意味する!),意識が生じるために神経 システムが満たすべき,「必要条件」を実験的にいくつ か見出すことは可能かもしれない.そういった意味で, 渡辺先生が受講者に与えた課題は現代の意識研究にお いていわば中心的な課題と言える.それでは具体的に, 一体どういったものが「必要条件」として考えられて いるのかを次節で解説する.
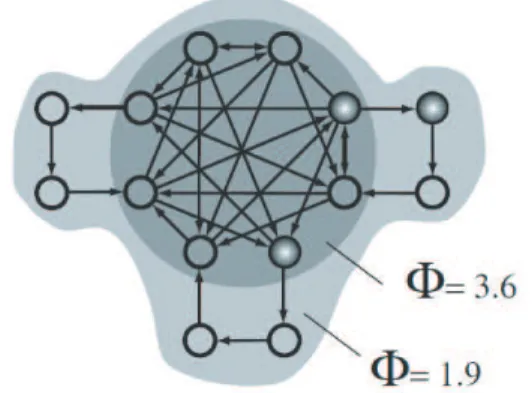
Fig. 1 Integrated information theoryとダイナミッ クコア.Tononi(2008)より.図のシステム全 体のintegrated informationはΦ = 1.9であ り,回路中央の部分システムはΦ = 3
.6であ ることを示す.この場合,中央の部分システム がダイナミックコアに相当する(ダイナミック コアに関しては6節を参照のこと.).
4. 意識と情報の統合
意識が生じるために必要な条件を一つ,極めて単純 に考えてみよう.まず,単体の神経細胞が意識を持っ ていないということは仮定する.そして,神経細胞が 大多数集まった脳というシステムが意識を持っている とする.両者の違いは何かと言えば,単体か,多数かと いう違いだ.しかしながら,いくら多数を集めたとし ても,神経細胞同士がお互いにつながっておらず,そ れぞれが独立に活動するシステムには意識は生じない だろう.従って,神経細胞が多数集まっていること,そ して複雑につながって相互作用し,情報をやりとりし ているということが意識が生じるための必要条件であ ると言える.
ここまでは,少し考えれば誰でも思いつく必要条件 であるが,ただ単にシステム内の素子間で相互作用し, 情報をやりとりしているだけでは意識は生じないはず で,より詳細にどのような相互作用があれば意識が生 じるかを明確化できるかどうかが問題となる.
この問題に対する一つのアプローチとして,Tononi が 提 唱 す る integrated information theory(IIT)が あ る
2, 21)
.IIT は 脳 内 の 領 野 間 ,ニュー ロ ン 間 に お い て ど の 程 度 情 報 の や り と り,統 合 が 行 わ れ て い る かを情報理論によって定量化する理論である(図1). IITにおいて定義された情報理論的指標,integrated
(A)
(B)
Fig. 2 Motion phosphene と TMS induced sco- toma.(A) MT+/V5野にTMSを適用する とmotion phospheneと呼ばれる,動きのあ る閃光が知覚される.(B)視覚刺激呈示時に V1野 にTMS を 適 用 す る と 視 野 欠 損(TMS induced scotoma)が生じる.
information(Φ)が大きければ,意識の程度が高く,Φ が小さければ意識の程度が低い.例えば,覚醒時におけ るΦより睡眠時におけるΦの方が小さいのではないか といったことが予想されている
2)
.それではどのくらい Φが大きければ意識があると言えるのかといった問題 に対しては,明確な答えが用意されてはいないのだが
(Tononi自身は意識はあるかないかといった離散的な
量ではなく連続的な量ではないかと考えている
21)
.), 一つの判断基準として有用かどうかを調べることは今 後の重要な研究課題であろう.
5. 高次領野から低次領野へのフィードバック
意識が生じるための必要条件としてもう一点,高次 領野から低次領野へのフィードバックが重要か否かと いう問題を考える.この問題を視覚的意識に関して調 べた実験として,Pascual-LeoneとWalshの実験が ある
16)
.彼らの実験は,MT+/V5野にtranscranial magnetic stimulation(TMS)を適用した際に生じる motion phosphene及び,刺激呈示時にV1野にTMS を適用した際に生じる視野欠損(TMS induced sco-
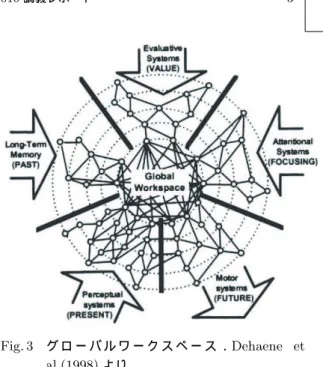
Fig. 3 グ ロ ー バ ル ワ ー ク ス ペ ー ス .Dehaene et al.(1998)より.
toma)を利用している(図2).彼らはV5野を磁気刺 激した後にV1野を磁気刺激して,motion phosphene に視野欠損が生じるかどうかを調べた.仮にmotion
phospheneに何ら影響がないということになれば,視
覚的意識は高次領野が担っていて,低次視覚系は網膜 同様,視覚情報の単なる中継基地に過ぎないというこ とになる.逆に,motion phospheneに影響があると すれば高次からフィードバックによるV1野の活動が 視覚的意識に必要ということになる.結果としては, V5野を磁気刺激した5-45ms後にV1野を磁気刺激 をすると,motion phospheneに影響が出ることが分 かった.この結果は,高次領野から低次領野へのフィー ドバックが視覚的意識に必要であるということを示唆 する.
6. 意 識 の 場 所
意識が生じるために神経システムが満たすべき必要 条件に関連して,脳内のどこに意識を生み出す神経シ ステムが存在するのかという問題を考えよう.
まず分かりやすい考え方として,BaarsやDehaene らによって提案されたグローバルワークスペース理論 がある
1, 7)
.グローバルワークスペースとは感覚処理 を行う神経システム,価値判断を行う神経システム, 記憶を担う神経システム,注意をコントロールするシ ステム,運動処理を行う神経システム等,モジュール 化された神経システムの全てがアクセス可能な神経回
路網のことを指す(図3).グローバルワークスペース は様々な情報を統合し,それに基づいて次の行動を実 行する司令塔の役割を果たす場所であり,グローバル ワークスペースで処理された情報が意識に相当すると いう考え方である.
グローバルワークスペース理論のように,特定の脳 内の領野,神経細胞集団が意識を担っているという考 え方とは別の立場を取るものとしては,Tononiのダイ ナミックコア仮説がある20).ダイナミックコア仮説に おいては,意識を担うニューロン集団は時々刻々変わ り,脳全体に分散していると考えられている.脳全体と 言っても,全ての神経細胞が意識に貢献しているかと いうとそうではなくて,前の節で紹介したintegrated information, Φが大きい神経細胞の部分集合の活動の みが直接意識に貢献していると考えられている.この Φが大きい神経細胞の部分集合のことをダイナミック コアと呼んでいる(図1).例えば,小脳はダイナミッ クコアには含まれないと考えられている.どの神経細 胞集団がダイナミックコアかは時々刻々変化していて, ある神経細胞がある時間においてはダイナミックコア の一部を担うが,また別の時間ではダイナミックコア に含まれていないといったことが起こり得る.
グローバルワークスペースとダイナミックコアのど ちらが正しいのかは未解決の問題である.もちろん, これら以外にも様々な仮説は存在する(例えば
12, 13))
. 今後の実験的検証によってどの仮説が間違いで,どの 仮説が正しそうかということが徐々に明らかになって いくと考えられる.
7. 生成モデルと意識の仮想現実メタファー
最後に,前節までで紹介してきた前提知識を踏まえて, 渡辺先生自身が提案する意識モデルを紹介する
22)(
図 4).
まず第一に,渡辺先生はダイナミックコア仮説を採 用しており,図4に示される神経回路全体がダイナミッ クコアを形成しているとしている.ただし,渡辺先生 が言うダイナミックコアの定義はTononiのそれとは ちょっと違っていて,ダイナミックコア全体が意識の神 経表象というよりは,ダイナミックコアの中にも,意識 の担い手として本質的な役割を担っているものと,そ れらを「接着」する裏方の二つが存在すると考えてい る.これは,ニューロン集団の保持情報として私たちの 視覚的意識に一致するのは中低次領野に限られるとい うことから発想されている(intermediate level theory of consciousness9)).回路全体として情報の統合の程 度が大きい,すなわちintegrated information, Φが
Fig. 4 渡辺先生の意識モデル.
大きいという意味はTononiのダイナミックコアの定 義と一緒である.
第二に,生成モデルが脳内に存在するという仮定を 置いている
10)
.生成モデルとは,脳が持っている前提 知識から生み出した知覚世界の予測である.視覚で言 えば,網膜→LGN→V1→… といった順方向の視覚 情報処理過程に加えて(図4の上方向の矢印),高次領 野から視覚世界の予測信号が逆方向に流れて来て(図 4の下方向の矢印),実際の信号と予測信号との差分, 予測誤差が計算されると考えられている.そして,こ の予測誤差が小さくなるように,高次領野の予測が更 新されていく.
渡辺先生はこの高次領野が生み出す知覚世界の予測 こそが意識の中身であると考えている.渡辺先生のモ デルでは,知覚世界の予測を生み出している神経細胞 集団の活動こそが意識の神経表象であり,その他の神 経細胞の活動はΦを大きくするために必要ではあるけ れども,意識の中身ではなく,ダイナミックコアを接 着する裏方の役割を果たしていると考えている.さら に渡辺先生は,脳が生み出す知覚世界の予測とは言わ ば,脳が作り出す「仮想現実」であり,脳が覚醒状態 において「仮想現実」を生み出すシステムは夢をみる ためのシステムとしても使われているのではないかと 考えている(意識の仮想現実メタファー
18)). 8. 両眼視野闘争とモデルの関係と電極刺激による検
証実験
渡辺先生のモデルを両眼視野闘争時の猿のニューロン 活動に当てはめて考えてみよう.Logothetisのグルー
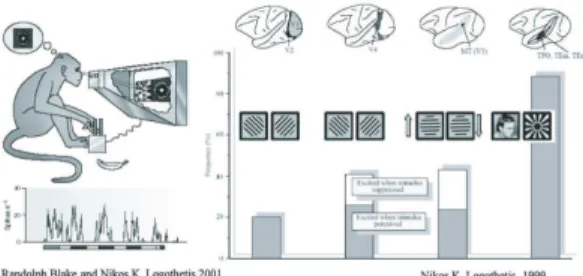
Fig. 5 両眼視野闘争時の猿のニューロン活動.Blake
& Logothetis(2002), Logothetis(1998)より 改変.
プの実験によって両眼視野闘争時に低次視覚野及び高 次視覚野において,神経細胞が知覚交代に応じる神経 細胞がどれくらいの割合存在するかということが調べ られている
3, 14)
(図5).高次領野になるほど知覚交代 に応ずる神経細胞の割合は増え,また興味深いことに V4野やMT野では刺激が知覚されないと逆に興奮す る神経細胞も存在する.
これらの神経細胞を渡辺先生のモデル(図4)に従っ て分類すると以下のようになる.
a. 中低次視覚野(V1-V4)で知覚交代に応じないニ ューロン:ダイナミックコア外,生成モデルにお ける感覚入力
b. 中低次視覚野で知覚交代に応じ,かつポピュレー ションとしての保持情報が知覚に一致するもの: コア内,意識の本質的担い手,生成モデルにおけ る予測表象
c. 中低次視覚野で知覚交代に対する応答が逆のもの
(知覚していない時に活動が上昇):コア内,接着 役,生成モデルにおける予測誤差表象
d. 高次視覚野で知覚交代に応答するもの:コア内,接 着役,生成モデルにおける高次シンボリック表象 以上のような神経細胞の分類の正当性を検証するため にはどのような実験を行えば良いだろうか?渡辺先生 はfMRIで 猿 の 脳 活 動 を 取 り な が ら 微 小 な 電 気 刺 激 (electrical microstimulation)をすることによって19), 以下のようなことを確かめることを考えている.
まず第一に,microstimulationをごく微小の電流を 流すことによって生じる摂動ととらえた場合,コア内の 神経細胞を刺激すれば,その影響がコア全体に及ぶこ とが予想される.コア外でも,コアとの接点を通してコ
アに影響が及ぶことが考えられるが,より限定的であ ると考えられる.こういった予想をmicrostimulation 及びfMRIデータの解析によって確かめる.
第二に,microstimulationを上記a-dタイプの神経 細胞の表象自体を変化させる道具としてとらえ,猿に 知覚報告させる場合,以下のようなことが予想される.
1. aタイプの神経細胞への刺激の知覚的影響は限定 的: 左右眼の刺激に選択性をもつ二つのbタイプ の神経細胞群の活動強度が逆転しない程度であれ ば,知覚に影響しない.
2. bタイプの神経細胞への刺激の知覚的影響は直接 的:bタイプの神経細胞の活動強度が逆転すれば, 知覚も逆転する.
3. c,dタイプの神経細胞への刺激の知覚的影響は間
接的:aタイプの神経細胞への刺激よりbタイプ への神経細胞との相互作用が強いく,bタイプの 神経細胞の活動強度が逆転すれば,その時点で知 覚も逆転する.
今後上記したような予測を検証することによって, モデルの正当性,もしくは問題点が明らかになってい くことと思われる.
9. ま と め
本稿では意識を生み出すために神経システムが備え るべき必要条件は何かといった問題を中心として,こ れまで提案されてきた意識モデル及び渡辺先生が提案 されているモデルについて概要を記述した.本稿は渡 辺先生の講義で取り上げられた内容を,筆者自身の視 点,理解の範囲内から再構築したものであって,渡辺 先生の講義を忠実に再現したものとはなっていないこ とに注意されたい.講義の中で筆者が理解できなかっ た部分は割愛している.ただし,本稿に明らかな間違 いなどがないかは渡辺先生のチェックを受けている. 本稿では実際の実験事実の記述が少なく,モデルや 概念の記述がほとんどであるため,読者が意識研究に 対し間違った印象を持ってしまうのではないかと危惧 している.現在の意識研究は実験的検証が可能なモデ ルや概念の提案,そして実際の実験的検証に強く重き が置かれたものになっており,一昔前の意識が哲学者 の問題であった時代とは様相が異なっていることを強 調しておきたい.7,8節で紹介したように,渡辺先生 ご自身もモデルの提案だけでなく,それを検証する実 験を含め研究を行っておられる.もちろん,何か確定的 なことを言うためにはまだまだ実験事実は不足してい るが,今後,意識の神経機構に関する実験的検証がど のように進んでいくかは非常に楽しみなところである.
10. 謝 辞
本稿を詳細に読んでいただき,多くの重要なコメン トをくださった渡辺正峰先生に感謝する.特に7,8節 の内容に関しては詳細な解説をいただき,筆者が講義 では理解できなかった部分の理解を大きく助けていた だいた.また,図2,4,5は渡辺先生からご提供いた だいたものである.北園淳氏も本稿に関して多くのコ メントくださり,原稿を改善していただいた.北園氏 は筆者が意識の問題に興味を抱くきっかけを与えてく ださった人物であり,意識に関する色々な問題に関し て議論していただいた.あわせて深く感謝したい.
参 考 文 献
1) Baars, B. J. (1988): A Cognitive Theory of Consciousness. Cambridge, MA: Cambridge University Press.
2) Balduzzi, D. & Tononi, G. (2008): Integrated information in discrete dynamical systems: motivation and theoretical framework. PLoS Comp. Biol., 4, e1000091.
3) Blake, R. & Logothetis, N. K. (2002) Visual competition. Nat. Rev. Neurosci., 3, 13-21. 4) Crick, F. & Koch, C. (1998): Consciousness
and neuroscience. Cerebral Cortex, 8, 97-107. 5) Crick, F. & Koch, C. (1995): Are we aware of neural activity in primary visual cortex? Na- ture, 375, 121-123.
6) Chalmers, D. J. (1995): Facing up to the problem of consciousness. Journal of Con- sciousness Studies, 2, 200-219.
7) Dehaene, S., Kerszberg, M. & Changeux, J. P. (1998): A neuronal model of a global workspace in effortful cognitive tasks. Proc. Natl. Acad. Sci. USA, 95, 14529-14534. 8) Dennett, D. (1991): Consciousness Explained.
Boston: Little & Brown
9) Jackendoff, R. (1987) Consciousness and the Computation Mind. Cambridge, Mas- sachusetts: MIT Press
10) Kawato, M., Hayakawa, H. & Inui, T. (1993): A forward-inverse optics model of recipro- cal connections between visual cortical areas. Network-Computation in Neural Systems, 4, 415-422.
11) Koch, C. (2004): The Quest for Conscious- ness: A Neurobiological Approach. Engle- wood, CO: Roberts & Company.
12) Lau, H.C. & Passingham, R. E. (2006): Rel- ative blindsight in normal observers and the neural correlate of visual consciousness. Proc. Natl. Acad. Sci. U.S.A. 103, 18763‐ 18768.
13) Lau, H. C. (2008): A higher order Bayesian decision theory of consciousness. Prog Brain Res. 168, 35‐ 48.
14) Logothetis, N. K. (1998) Single units and con- scious vision. Phil. Trans. R. Soc. Lond. B, 353, 1801-1818.
15) McGinn, C. (1991): The Problem of Con- sciousness. Oxford, UK: Basil Blackwell 16) Pascual-Leone, A. & Walsh, V. (2001): Fast
backprojections from the motion to the pri- mary visual area necessary for visual aware- ness. Science, 292, 510-512.
17) Penrose, R. (1989): The Emperor’s New Mind. Oxford, UK: Oxford University Press 18) Revonsuo, A. (2000): Prospects for a scientific
research program on consciousness. In T. Met- zinger (Ed.), Neural correlates of conscious- ness. Cambridge, MA: MIT Press
19) Tolias, A. S., Sultan, F., Augath, M., Oel- termann, A., Tehovnik, E. J., Schiller, P. H.
& Logothetis, N. K. (2005) Mapping cortical activity elicited with electrical microstimula- tion using fMRI in the macaque. Neuron, 48, 901-911.
20) Tononi, G. & Edelman, G. M. (1998) Neuro- science - Consciousness and complexity. Sci- ence, 282, 1846-1851.
21) Tononi, G. (2008): Consciousness as inte- grated information: a provisional manifesto. Biol. Bull., 215, 216-242.
22) 渡辺正峰(2010):「意識」,イラストレクチャー 認知神経科学−心理学と脳科学が解くこころの 仕組み−村上郁也(編).197-214: オーム社