M I Y A D I A W A R D
Nobuyuki Kutsukake
Complexity, dynamics and diversity of sociality
in group-living mammals
Received: 3 May 2008 / Accepted: 7 October 2008 / Published online: 19 November 2008 ÓThe Ecological Society of Japan 2009
AbstractNumerous studies in group-living animals with stable compositions have demonstrated the complex and dynamic nature of social behaviour. Empirical studies occasionally provide principles that cannot be applied directly to other group-living species. Because of this, researchers are required to address fine-scaled conceptual questions and to incorporate species-specific character- istics of the study species. In this paper, I raise three key topics that will promote our understanding of animal sociality: the effects of heterogeneous social relationships on the pattern, distribution, and function of social interactions; conflict management for maintaining group living; and meta-dyad-level perspectives for understand- ing dyadic social relationships and behaviours. Through the discussion of these topics together with examples of group-living mammals, I emphasise the importance of direct behavioural observations and functional analyses in studies of species- or taxonomic-group-specific char- acteristics of social behaviour in a wide range of taxo- nomic groups. In addition to approaches focusing on specificity, another approach that examines the general principles or common characteristics found across different taxonomic groups could provide synthetic and reductive frameworks to understand divergent sociality. The complementary use of these two approaches will offer a comprehensive understanding of social evolution in group-living animals.
Keywords Sociality Æ Social relationship Æ Group living Æ Mammals Æ Social diversity
Introduction
Understanding the emergence and evolution of complex sociality is one of the central issues in ecology and evolutionary biology (Wilson1975; Maynard Smith and Szmathma´ry 1995). Theoretically, it is predicted that animals form groups when the benefits of group living exceed the costs. In general, the benefits of group living include a decreased probability of being preyed upon, the sharing of useful information, thermoregulation, and an increased probability of winning competitions with conspecific or competing species (Krause and Ruxton 2002). On the other hand, costs mainly result from competition for limited resources, such as food and reproductive opportunities. Additional costs include a high probability of parasitic infection and increased disease transmission (Krause and Ruxton2002).
Aggregation is widely seen in animals of various taxonomic groups (Wilson 1975; Krause and Ruxton 2002). In contrast to simply aggregated species with temporally fluid group composition, some group-living species form groups that have relatively stable compo- sitions of members with non-random structures (Whitehead 1997; Whitehead and Dufault 1999; indi- vidualized societies; de Waal and Tyack2003). In such groups, group members repeatedly engage in social interactions, and as a result, the histories of the social interactions shape the stable social relationships (Fig.1; Hinde1976). Sociality within these groups is regarded as an autonomous distributed system in which each group member employs social strategies to maximise the (inclusive) fitness. In addition, the strategy of each group member is decided by a complex feedback system that is affected by the social strategies of other group members. Such social environments are regarded as complex adaptive systems that produce distinct selective pres- sures and promote the formation of complex and dynamic social relationships, which cannot be observed in species with unstable group membership. Due to the complex and dynamic nature of animal sociality,
Nobuyuki Kutsukake is the recipient of the 12th Denzaburo Miyadi Award.
N. Kutsukake (&)
Department of Evolutionary Studies of Biosystems, The Graduate University for Advanced Studies, Hayama, Miura, Kanagawa 240-0193, Japan E-mail: [email protected] Tel.: +81-46-8581610
Fax: +81-46-8581544
DOI 10.1007/s11284-008-0563-4
researchers occasionally reach conclusions that cannot be applied to other taxonomic groups because of dif- ferences in ecology, life history, and other biological characteristics (Fig.1). In such situations, researchers are required to address fine-scaled conceptual questions regarding animal sociality by incorporating the specific characteristics of the study species.
In this paper, I review the previous literature and selectively identify three key topics that I believe will promote our understanding of the complexity and dynamics of sociality in mammals. These topics include the following questions: (1) how do heterogeneous social relationships affect the pattern, distribution, and func- tion of individual social behaviour, (2) how do group members regulate social relationships and maintain group living, and (3) how does the presence of other group members affect the relationships and interactions between two individual group members? Through the discussion of these questions, I point out that long-term comparative data of social behaviour are lacking in a wide range of taxonomic groups and emphasise the importance of direct behavioural observations and functional analyses of social interactions. In the latter part, I show the direction of future studies by discussing the contrasts between approaches focusing on specificity
and generality for understanding animal sociality. Note that this paper does not aim to provide a comprehensive review of animal sociality. Rather, the cited examples are biased to long-lived mammals for the following two reasons. First, few attempts have been made to review social behaviour across different taxonomic groups of mammals, despite the fact that mammalian sociality has received attention in previous studies (Silk2007). A lack of reflection on mammalian biological characteristics (i.e., long life history and the complexity of social structures) and research investigations might bias the comprehensive theorisation of social evolution in ani- mals. Second, it is usually difficult to evaluate the fitness benefits of a single social interaction or social relation- ships in long-lived mammals (Silk2002, 2007). Because of this problem, it has been difficult to estimate the importance of selection pressures caused by social behaviour (i.e., social selection) relative to the magni- tude of natural selection and sexual selection. As a re- sult, the investigations of evolutionary aspects of animal sociality remain one of the most challenging topics in current evolutionary ecology.
Effects of heterogeneous social relationships on social behaviour
Even in simply aggregated species with unstable com- positions, the nature of temporal social relationships among individuals is heterogeneous because of differ- ences in individual intrinsic factors, such as age and sex. In contrast to these species, the heterogeneity of social relationships is pronounced in group-living animals with structured societies, both because of the variation in relatedness between individuals and by the history of the repeated social interactions between group members. The characteristics of the social relationships shape the individual strategies and interactions between two indi- viduals. Because the group structure is determined by the sum of the social relationships within a group, het- erogeneity in social relationships ultimately results in heterogenic variation at the group level. The formation of stable and heterogeneous social relationships makes animal sociality increasingly complicated, such that principles found in aggregated species may not apply to group-living species with stable memberships and re- peated social interactions. Therefore, it is vital to investigate how the heterogeneity of social relationships affects the pattern, distribution, and function of the so- cial interactions. At the behavioural level, however, de- tailed analyses of social interactions have been conducted in limited species, such as primates, which prevents comparison of mammalian socialities.
Arguably the most influential hypothesis posited to explain heterogeneous social relationships is the kin selection hypothesis (Hamilton 1964; Maynard Smith 1964). The kin selection hypothesis predicts that related individuals engage in cooperative behaviour more often than unrelated individuals because of increased inclusive
Fig. 1 Scheme of sociality in an animal group with heterogeneous social relationships. Sociality is determined by the combination of selective pressures, including adaptation to ecological factors and constraints of phylogeny and life history. The social structure can be regarded as a social network (i.e., an entity of the social relationships within a group) composed of dyadic social relation- ships between group members. Dyadic relationships are largely determined by individual (e.g., age and sex) or relationship (e.g., relatedness) intrinsic factors. In the figure, dashed circles represent relatedness among group members. Differences in brightness and thickness of the lines represent social bonds with different magnitudes. Note that the characteristics of social relationships are not necessarily determined by the relationship intrinsic factors (see text). The characteristics of each social relationship are also affected by both (A) the history of dyadic social interactions between two group members and (B) other group members. A box (A) shows temporal sequences of social interactions between two individuals (t indicates the present social interaction, while the past interactions are indicated by t–n). A box (B) shows the influences of other group members on dyadic social relationships
fitness (Hamilton1964). Numerous studies have shown strong influences of relatedness on social behaviour, particularly on cooperation (reviewed in Dugatkin1997; Korb and Heinze 2008). However, recent studies high- light that relatedness does not always guarantee valuable and cooperative relationships (Griffin and West 2002; West et al. 2002) and that kin selection benefits have been overestimated (Clutton-Brock2002). For example, the costs of kin competition diminish the benefits of kin selection in viscous populations where individual dis- persal is limited (Frank 1998; West et al. 2002). Coop- eration among related individuals that superficially fits the kin selection hypothesis can often be explained by other ultimate mechanisms (Clutton-Brock 2002). Therefore, it is incorrect to assume a priori that relat- edness is always tightly linked to cooperative social relationships.
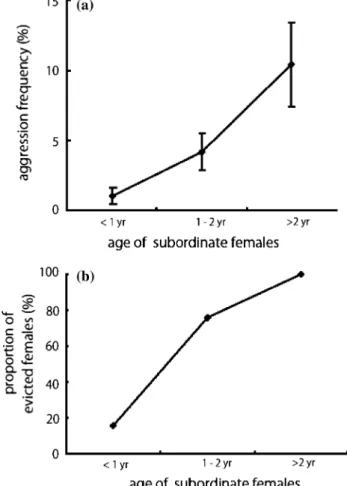
Observations of social behaviour are one of the most informative and direct ways to investigate and quantify the effects of heterogeneous social relation- ships. Studies in the cooperatively breeding carnivore, the meerkat (Suricata suricatta), provide one such example. This species forms multi-male, multi-female groups with high reproductive skew. Subordinates, usually the offspring of dominant pairs, do not repro- duce, but instead help in rearing the offspring of the dominant pairs. Like other cooperatively breeding species, help by subordinates enhances the reproductive output of dominants (Emlen 1997). At the same time, however, there is an intense intrasexual competition between same-sexed dominant and subordinate indi- viduals, irrespective of the degree of relatedness be- tween them. That is, intrasexual aggression is frequent among same-sexed group members in meerkats (Ku- tsukake and Clutton-Brock 2006a, b). This aggression is particularly pronounced among females (Clutton- Brock et al. 2006). Dominant females attack older subordinate females who are likely to reproduce and evict them from the group at the later stages of the dominant female’s pregnancy (Fig. 2; Kutsukake and Clutton-Brock 2006a). Evicted subordinate females show increased stress levels, decreased fertility, and abortion if pregnant (Young et al.2006). Although the peaceful and cooperative aspects of cooperatively breeding species have received attention in previous studies, these results reveal the understudied competi- tive aspects of cooperatively breeding societies, i.e., relatedness does not guarantee peaceful relationships in cooperatively breeding species, and high reproductive skew is achieved as a result of intense intrasexual competition (Kutsukake and Clutton-Brock 2006a, b, 2008a, b; Clutton-Brock et al. 2006).
The study of heterogeneous social relationships occasionally provides an opportunity to test general ecological principles and the function of social behav- iour in sophisticated ways. One such example is the reconsideration of group size effects in vigilance behaviour. Vigilance is used mainly for anti-predator purposes in group-living animals (Caro 2005). Indi-
vidual vigilance rates are known to decrease as group size or local gregariousness increases due to dilution and the presence of many ‘eyes’ (reviewed in Krause and Ruxton 2002; Caro 2005). However, recent careful studies have begun to notice social influences on vigi- lance and have clarified that group size effects are not universal (reviewed in Treves 2000). For example, in the wild chimpanzee (Pan troglodytes) the presence of other group members increases, not decreases, indi- viduals’ levels of vigilance (Fig.3a, b; Kutsukake2006, 2007). Given that chimpanzees are a highly social species; this result indicates that vigilance may be di- rected toward competitive group members. If vigilance functions as a way to monitor other group members, it is predicted that the characteristics of social relation- ships affect vigilance level, with individuals being more vigilant toward competitive group members. The above prediction is supported by studies indicating that chimpanzees are more vigilant when non-association group members are nearby than when other categories
Fig. 2 Intrasexual reproductive conflicts among female meerkats. a Aggression by dominant females in relation to the age of subordinate females. Aggression frequency indicates the proportion of encounters with aggression divided by the total number of encounters for each dyad of the same-sexed dominant and subordinate females. b The proportion of females evicted by a dominant female from a group for each category of subordinate female age. The figures are redrawn from Kutsukake and Clutton- Brock (2006a). Individual mean ± SE is shown
of group members are nearby (Fig. 3c; Kutsukake 2006). Likewise, behavioural observations on wild chimpanzees show that the rate of self-directed behav- iour, a behavioural indicator of emotional state and individual stress level (Maestripieri et al.1992), shows a similar pattern (Kutsukake 2003). Specifically, females increased the rate of self-directed behaviour more when other group members were nearby than when a non- affiliative group member was in their proximity (Ku- tsukake 2003). Influences of social context and rela- tionship quality have been reported in other mammals (e.g., brown capuchin monkeys Cebus apella, Hirsch 2002; giraffe Giraffa camelopardalis Cameron and du Toit2005; elk Cervus elaphus Lung and Childress2007; European rabbits Oryctolagus cuniculus; Monclus and Roedel 2008), suggesting that social influences of vigi- lance are common in group-living mammals.
These studies suggest that intrinsic factors of rela- tionships, such as relatedness, do not always determine the characteristics of social relationships. At the same time, the consideration of heterogeneous social rela- tionships may facilitate a more fine-scaled understand- ing of ecological principles. The quantification of heterogeneous social relationships cannot be performed from long-term demographic data or life history data. Therefore, detailed functional analyses of social behav- iour in individual animals are indispensable for under- standing animal sociality.
Conflict management for the regulation of social relationships and maintenance of group living
Although animals form groups when the benefits exceed the costs, it is too simplistic to assume that this payoff is passively determined with no room for group members to affect its outcome. Some animals actively maximise the net benefits by reducing the costs in order to main- tain group living. Social behaviours that reduce the costs of aggression (i.e., consumption of energy and time and damage of social relationships between opponents) are referred to as conflict management (Aureli and de Waal 2000). Here, I exemplify three behavioural options of conflict management (dominance, greeting, and recon- ciliation) and discuss what is known and unknown for each behaviour.
The most common form of conflict management is the mediation of dominance relationships by dominant and submissive interactions and signals (Pusey and Packer 1997; Kutsukake2000; Preuschoft and van Schaik2000; Searcy and Nowicki 2005). Although dominance rela- tionships reduce unnecessary conflicts over limited re- sources (Pusey and Packer 1997), relatively little is known regarding the function of submission as a mech- anism of conflict management. Submissive signals are used to convey relative dominance relationships among group members and act to reduce the probability of
Fig. 3 Influences of group members nearby (within 3 m) on individual vigilance level in wild chimpanzees. a Vigilance duration measured by a 2-min focal observation method in relation to the number of group members nearby. b Vigilance level measured by point sampling during a 5-min interval in relation to the number of group members nearby. c Effects of social relationships with proximate group members on individual vigilance level measured by point sampling during a 5-min interval. The figures are redrawn from Kutsukake (2006, 2007). Individual mean ± SE is shown
aggression by dominant individuals (Preuschoft and van Schaik 2000). However, closer examination of these functions has been rare (Flack and de Waal2007). Some studies have found that social signals that were previ- ously regarded as submissive are not (Stevens et al.2005) and that submission does not function as a form of conflict management (Fig. 4a; Kutsukake and Clutton- Brock2008b). Mathematical theories of submission have been formulated only recently (Matsumura and Hayden 2006). Therefore, it is necessary to functionally test submissive signals.
To facilitate social harmony, animals engage in a ritualised pattern of non-aggressive behaviour that usually occurs during a reunion. This is referred to as greeting behaviour (Colmenares et al. 2000). For example, in the black-and-white colobus (Colobus guer- eza) individuals engage in ‘over-head mounting’ and other types of contact behaviour. Functional and con- textual analyses have shown that this behaviour is per- formed mainly by subordinates to dominant individuals and facilitates the occurrence of affiliation between in- teractants (Fig. 4b; Kutsukake et al. 2006). This sug- gests that greeting behaviour functions as conflict management for this species. In other species, greeting behaviour may have different functions, including the reaffirmation of social bonds (guinea baboons Papio papio, Whitham and Maestripieri2003), suggesting that ritualised contact among group members has evolved for divergent functions in different species.
The most explicit form of conflict management is
‘reconciliation’. Reconciliation is the affiliation of opponents following an aggressive interaction (de Waal and van Roosmalen 1979). Reconciliation functions to
reduce the probability that the victim will suffer further attack by the aggressor (Fig. 4c; Aureli et al. 2002). In addition, the behaviour reduces the post-aggression stress of the opponents (Aureli et al. 2002). These observations indicate that this behaviour reduces the costs associated with aggression and resolves conflict among individuals (conflict resolution). Reconciliation does not occur after all cases of aggression, and occurrence rates show inter-species, intra-species, and within-group variation (Arnold and Aureli 2006). Rec- onciliation is particularly effective in repairing damaged social relationships between group members with strong social bonds because these dyads experience higher levels of post-aggression stress than ones with weak social bonds (long-tailed macaques Macaca fascicularis: Aureli 1997; Japanese macaques Macaca fuscata: Kutsukake and Castles2001; chimpanzees: Koski et al.2007). Aureli et al. (2002) predicted that reconciliation should be common among species that live in stable social groups, have individualised relationships, and experience hostil- ity after aggression, particularly among species in which aggressive interactions disturb biologically valuable so- cial relationships. Reconciliation is widely observed in group-living primates (Arnold and Aureli 2006) and in other group-living mammals (reviewed in Schino 2000; domestic goats Capra hircus: Schino 1998; spotted hye- nas Crocuta crocuta: Wahaj et al.2001; bottlenose dol- phins Tursiops truncatus: Samuels and Flaherty 2000; Weaver 2003; Tamaki et al.2006; domestic dogs Canis familiaris: Cools et al.2008; wolves Canis lupus: Cordoni and Palagi2008), suggesting that conflict resolution is a common behavioural option for group-living animals. However, reconciliation is not always demonstrated in
Fig. 4 Effects of a submission, bgreeting, and c reconciliation on the occurrence of aggression or affiliation after each type of social behaviour. a The effects of submission during aggression on the probability of aggression reoccurrence in the next social interaction in meerkats. The figure is redrawn from Kutsukake and Clutton-Brock (2008b). Individual mean + SE are shown. b Effects of greeting behaviour on the probability of affiliation between interactants in black and white colobus. The figure is redrawn from Kutsukake et al. (2006). The mean probability based on pooled data is shown. c Effects of reconciliation on the probability of aggression reoccurrence in Japanese macaques. The figure is redrawn from Kutsukake and Castles (2001). Individual means are shown
group-living animals that fit the aforementioned criteria set by Aureli et al. (2002) (red-bellied tamarins Saguinus labiatus: Schaffner and Caine2000; Schaffner et al.2005; common marmosets Callithrix jacchus jacchus: Westlund et al. 2000; black lemurs Eulemur macaco: Roeder et al. 2002; meerkats: Kutsukake and Clutton-Brock 2008b; naked mole-rats Heterocephalus glaber: Kutsukake, personal observation; see also Kappeler 1993 and Rol- land and Roeder2000for ring-tailed lemurs Lemur catta: van den Bos1998 for domestic cats Felis catus). Social factors associated with the evolution of reconciliation require further discussion (Aureli et al.2002; Kutsukake and Clutton-Brock2008b), but it is likely that no single factor can explain the absence of reconciliation in all species.
As discussed, group-living animals have behavioural mechanisms that manage and resolve conflicts in order to cope with the temporal disturbance of social rela- tionships. These types of behaviours are important in controlling the dynamics of social relationships and act to maximise the net benefits of group living. At the same time, the forms of behaviour and their relative impor- tance vary among different societies. Furthermore, behavioural distributions across species are currently unknown. Further empirical studies in species with divergent socialities will facilitate our understanding of the selective pressures shaping conflict management.
Understanding of social behaviour from the meta-dyad-level perspective
One important lesson from previous studies of animal sociality is that animals live in complex social networks within which group members are embedded (Krause et al.2007). Since a single social interaction between two individuals is affected by the social behaviours and relationships among other individuals in their social network, understanding the effects of social interactions should not be restricted to a local scale (i.e., specifically between the interactants), but should instead be exam- ined at a meta-dyad or more global level (i.e., between third parties and within a social network; Cheney and Seyfarth 1990, 2007; Harcourt and de Waal 1992; McGregor 2005; Conradt and Roper 2005). Here, I outline two examples of why meta-dyad-level perspec- tives are necessary and how the consideration of meta- dyad-level perspectives promotes our understanding of animal sociality.
Among group-living mammals, social interactions involving more than three individuals are commonly seen. Such polyadic interactions can be frequently ob- served within the context of aggression. Group-living mammals, such as primates, spotted hyenas, bottlenose dolphins, wild dogs (Lycaon pictus), and coatis (Nasua nasua), use coalitionary aggression, i.e., joint aggression toward a third party, to gain social benefits such as dominance and reproductive advantages (de Waal1982; Nishida 1983; Harcourt and de Waal 1992; Kutsukake
and Hasegawa 2005; Engh et al. 2005; Romero and Aureli2008). Third parties who are not involved in the aggressive bout may interact with the opponents fol- lowing the behaviour (e.g., consolation, appeasement, solicited consolation; Watts et al.2000; Das2000; Wittig and Boesch 2003; Kutsukake and Castles 2004; Koski et al.2007; Fraser et al.2008), or dominant individuals can intervene in ongoing aggression, thus terminating the aggression (Petit and Thierry 2000; Flack et al. 2006). Social interactions in a given dyad influence the nature and occurrence of social interactions among bystanders: the third-party individuals interact with each other following aggression (Cheney and Seyfarth 1986, 1989; Aureli et al. 1992; Judge and Mullen 2005). The function of these social behaviours following aggression have been regarded as conflict management because they reduce the probability of escalated aggression and reg- ulate the social relationships; however, only a few empirical studies have investigated the function of these types of social behaviour (Palagi et al.2006; Koski and Sterck 2007). These polyadic interactions increase the complexity and dynamics of animal sociality because the possible combinations of interactants should increase in species with polyadic interactions relative to ones with- out these interactions.
The presence of group members affects social behaviour and relationships in animals even if these individuals are not directly involved in the social inter- actions. The biological market effect (Noe¨ et al. 1991; Noe¨ and Hammerstein 1994, 1995) is one theory that formulates how other group members affect the rela- tionships between two individuals. This theory proposes that exchanges of social behaviour (e.g., cooperation) are affected by the ratio of asymmetric actor and receiver individuals. Previous studies have shown that the market effect operates in various ecological conditions (e.g., interspecific mutualism, Bshary 2001; Hoeksema and Schwartz 2001), including social behaviour (the econ- omy of coalition formation, Noe¨ 1992; allogrooming exchanges, Barrett and Henzi 2006; obtaining female sexual information by males, Stopka and Macdonald 1999; attitude toward ex-group members, Schaffner and French 1997). Studies in meerkats have shown that similar principles might operate in cooperatively breed- ing species. In meerkats, a low offspring-to-helper ratio reduces the per capita work burden of group members and is positively related to the reproductive success of dominant individuals (Clutton-Brock et al. 2001). Due to the Allee effect, groups with few numbers of helpers do not succeed in reproduction (Clutton-Brock et al. 2001). Therefore, it is predicted that the value of each subordinate is high for dominant males in small groups compared to large groups. Accordingly, it is predicted that the aggressiveness of a dominant individual is positively correlated with the number of subordinates within a group. Indeed, analyses of aggression by dominant males in meerkats have demonstrated that the number of subordinates in a group is positively related to the rate of intrasexual aggression by a dominant male
(Fig.5; Kutsukake and Clutton-Brock 2008a). At the same time, dominant males attack non-natal males who can be reproductive within a group more frequently than natal males, suggesting that reproductive conflict is re- flected in social behaviour. Furthermore, the difference in aggression toward non-natal and natal males is slight in small groups, but exaggerated in large groups (Fig.5; Kutsukake and Clutton-Brock 2008a). This result sug- gests that aggressiveness by dominant males is moder- ated by the number of subordinates within a group, including reproductive rivals, which is in agreement with the predictions of the biological market effect.
These examples show that social interactions and relationships between two individuals are under the influence of other group members. These examples fur- ther posit the necessity of meta-dyad-level perspectives in predicting and correctly interpreting animal social behaviour. Furthermore, proper statistics that would enable researchers to analyse social interactions at the meta-dyad level without dividing groups into dyads have not been fully developed. Recent advances in analytical techniques (e.g., Krause et al. 2007; Wey et al. 2008; Whitehead2008) are expected to overcome this problem in future studies. For example, social network analysis (Scott 2000), which is based on mathematical ideas of graph theory, is a powerful analytic method that quan- tifies the metric of the social structure; a graph (i.e., network) is simply a set of lines (i.e., dyadic relationship
between group members) connecting nodes (i.e., indi- viduals), and social network analysis calculates the properties of the patterns formed by the lines, such as the density of the network or centrality of each node (Scott2000). Social network analysis has recently been applied to investigate the dynamics of the social struc- ture and relationships of animals (e.g., Krause et al. 2007; Wey et al. 2008; Whitehead 2008). By analyzing the structure of social networks without breaking them down to individual relationships, social network analysis could successfully quantify sociality across taxonomic groups and could provide unique insights that cannot be gained from analyses of dyadic relationships alone.
Conclusions and future directions: toward a comprehensive understanding of social diversity
In this paper, I have highlighted three conceptual questions that promote our understanding of sociality in animals: (1) the effects of heterogeneous social rela- tionships on the pattern, distribution, and function of social behaviour, (2) conflict management for the maintenance of group living, and (3) meta-dyad-level perspectives in analysing social behaviour. Unfortu- nately, detailed analyses on sociality, as well as contex- tual and functional analyses on social behaviour, have been mainly conducted in limited taxonomic groups (e.g., primates and eusocial insects). Additional data on the structure and functions of sociality in various taxo- nomic groups is necessary. Although this paper mainly focused on mammals, the questions addressed here will be useful in analysing sociality in other vertebrates (de Waal and Tyack 2003; Korb and Heinze 2008; e.g., birds: Marler1996; Emery et al.2007; Seed et al.2007). Some invertebrates also show social behaviour similar to that discussed in this paper (e.g., the individual recog- nition in the paper wasp Polistes fuscatus: Tibbetts2002; ritualized greeting in crayfish Procambarus clarkii: Issa and Edwards 2006; coalition formation in fiddler crab Uca mjoebergi: Backwell and Jennions2004; aggression and submission in paper wasp Polistes dominulus: Cant et al.2006; allocleaning in the ocypodid crab Macroph- thalmus banzai: Ueda and Wada1996). Investigations on the functional and fitness consequences of these inter- actions will aid in clarifying whether these social behaviours have similar functions to those found in phylogenetically distant mammals and will help us understand the selective forces that promote the evolu- tion of particular social behaviours. The similarities may be superficial because of the critical differences in bio- logical characteristics between invertebrates and verte- brates. For example, the cognitive ability of study species may differentiate the function of the social behaviour, and researchers must consider the function of social behaviour separately according to taxonomic group. Although we do not have enough long-term and comparative data to discuss this idea, it seems that there
Fig. 5 Intrasexual reproductive conflicts reflected in aggression between dominant and subordinate male meerkats. Aggression frequencyindicates the proportion of aggressive encounters divided by the total number of encounters for each dyad of same-sexed dominant and subordinate males. Subordinate males are catego- rised according to their relationship with dominant males. Non- natal males, both unrelated and non-sibling, can be reproductive rivals for dominant males. Figures are redrawn from Kutsukake and Clutton-Brock (2008a). Individual mean ± SE is shown
is no a priori reason to employ such classification. More importantly, such a classification may not allow us to test the interesting hypothesis that the exhibition of so- cially complex behaviour requires sophisticated cogni- tive ability.
Thus far, I have stressed the importance of enhancing long-term behavioural observations of iden- tified individuals in a wide range of species for the de- tailed analyses of social behaviour. In each species, sociality is determined by the combined effects of dif- ferent ecological (e.g., predation pressure and food re- sources) and evolutionary (e.g., constraints of phylogeny and life history) factors (Fig. 1). In addition, social traits are shaped by complex frequency-depen- dent interactions and feedback loop systems among social strategies by conspecific individuals, which makes the social behaviour peculiar to a particular species. Based on these backgrounds, the uniformed fitting of ecological or evolutionary principles found in the majority of species may overlook species or taxonomic group-specific characteristics. In the examples cited in this paper, polyadic interactions and reconciliation do not occur in all group-living species. The investigation of such specific social behaviour is useful for elucidating the magnitude of the social complexity in each study species, and it is vital to address fine-scaled questions that incorporate species-specific or taxonomic-group- specific characteristics of social complexity and dynamics. At the same time, however, increasing numbers of empirical studies show divergent patterns of animal sociality and provide a synthetic view of social diversity. Previous studies have focused on key bio- logical parameters that can be commonly confirmed in animals of different taxonomic groups and formulate the synthetic view of animal social diversity (e.g., reproductive skew: Vehrencamp 1983a, b; Emlen1997; Johnstone 2000; Kutsukake and Nunn 2006, 2009; so- cial network: Krause et al. 2007; Wey et al. 2008; Whitehead 2008; genetic structure: Ross 2001; life his- tory: Arnold and Owens 1998, 1999; Hatchwell and Komdeur 2000; Ligon and Burt 2004; Blumstein and Armitage1998; Helms Cahan et al.2002; food type and distribution or socioecology: van Schaik 1983, 1989; Sterck et al. 1997; Isbell and Young 2002). An advan- tage of such a synthetic approach is that researchers can directly compare social characteristics across taxo- nomic groups in a quantitatively similar manner, which could reveal the broad evolutionary pattern in animal sociality and provide powerful predications that can be applied to broad ranges of taxonomic groups. Attempts to synthesise social diversity are still in the early stages, and it is questionable whether these attempts are suffi- cient to explain social diversity in animals. Therefore, more emphasis must be placed on the building and testing of synthetic theories of social evolution. It should be also noted that synthetic theories occasion- ally miss species- or taxon-specific traits. Too much emphasis on general principles will prevent our under- standing of species-specific traits (‘‘the devil is in the
details’’), while too much emphasis on specific questions will miss the broad perspectives of social diversity (‘‘cannot see the forest for the trees’’); thus, I emphasise the importance of the complementary use of both ap- proaches for a comprehensive understanding of animal socialities.
AcknowledgmentsThis paper is based on the presentation for the Miyadi Award at the 55th Annual Meeting of the Ecological Society of Japan, March 2008. I would like to thank all colleagues who supported my previous studies. Special thanks go to my supervisors, mentors, and colleagues, particularly Toshikazu Hasegawa, Duncan L. Castles, Toshisada Nishida, Noyuri Suet- sugu, Takafumi Ishida, Tim H. Clutton-Brock, Charlie L. Nunn, Kazuo Okanoya, Mariko Hasegawa, and Keiko K. Fujisawa. Masayo Soma, Masakado Kawata, Kazuhiro Eguchi, and Dan Blumstein gave critical comments on the manuscript. My studies were supported by JSPS Research Fellowships, RIKEN Special Postdoctoral Researchers Program, Hayama Center for Advanced Studies and Grant-in-Aid for Young Scientists B and Start-up (no. 18870025 and 20770023).
References
Arnold KE, Owens IPF (1998) Cooperative breeding in birds: a comparative test of the life history hypothesis. Proc R Soc Lond B Biol Sci 265:739–745. doi:10.1098/rspb.1998.0355
Arnold KE, Owens IPF (1999) Cooperative breeding in birds: the role of ecology. Behav Ecol 10:465–471. doi:10.1093/beheco/ 10.5.465
Arnold K, Aureli F (2006) Postconflict reconciliation. In: Campbell CJ, Fuentes A, MacKinnon KC, Panger M, Bearder SK (eds) Primates in perspective. Oxford University Press, New York, pp 592–608
Aureli F (1997) Post-conflict anxiety in nonhuman primates: the mediating role of emotion in conflict resolution. Aggress Behav 23:315–328. doi:10.1002/(SICI)1098-2337(1997)23:5<315:: AID-AB2>3.0.CO;2-H
Aureli F, Cords M, van Schaik CP (2002) Conflict resolution fol- lowing aggression in gregarious animals: a predictive frame- work. Anim Behav 64:325–343. doi:10.1006/anbe.2002.3071 Aureli F, Cozzolino R, Cordischi C, Scucchi S (1992) Kin-oriented
redirection among Japanese macaques: an expression of a re- venge system? Anim Behav 44:283–291. doi:10.1016/0003- 3472(92)90034-7
Aureli F, de Waal FBM (2000) Natural conflict resolution. Uni- versity of California Press, California
Backwell PRY, Jennions MD (2004) Coalition among male fiddler crabs. Nature 430:417. doi:10.1038/430417a
Barrett L, Henzi SP (2006) Monkeys, markets and minds: biolog- ical markets and primate sociality. In: Kappeler PM, van Schaik CP (eds) Cooperation in primates and humans: mech- anisms and evolution. Springer, Berlin, pp 209–232
Blumstein DT, Armitage KB (1998) Life history consequences of social complexity: a comparative study of ground-dwelling sciurids. Behav Ecol 9:8–19. doi:10.1093/beheco/9.1.8
van den Bos R (1998) Post-conflict stress response in confined group-living cats (Felis silvestris catus). Appl Anim Behav Sci 59:323–330. doi:10.1016/S0168-1591(98)00147-6
Bshary R (2001) The cleaner fish market. In: Noe¨ R, van Hooff JARAM, Hammerstein P (eds) Economics in nature: social dilemmas, mate choice and biological markets. Cambridge University Press, Cambridge, pp 146–172
Cameron EZ, du Toit JT (2005) Social influences on vigilance behaviour in giraffes, Giraffa camelopardalis. Anim Behav 69:1337–1344. doi:10.1016/j.anbehav.2004.08.015
Cant MA, English S, Reeve HK, Field J (2006) Escalated conflict in a social hierarchy. Proc R Soc Lond B Biol Sci 273:2977–2984. doi:10.1098/rspb.2006.3669
Caro T (2005) Antipredator defenses in birds and mammals. Uni- versity of Chicago Press, Chicago
Cheney DL, Seyfarth RM (1986) The recognition of social alliances by vervet monkeys. Anim Behav 34:1722–1731. doi:10.1016/ S0003-3472(86)80259-7
Cheney DL, Seyfarth RM (1989) Redirected aggression and rec- onciliation among vervet monkeys, Cercopithecus aethiops. Behaviour 110:258–275. doi:10.1163/156853989X00501 Cheney DL, Seyfarth RM (1990) How monkeys see the world:
inside the mind of another species. University of Chicago Press, Chicago
Cheney DL, Seyfarth RM (2007) Baboon metaphysics: the evolu- tion of a social mind. University of Chicago Press, Chicago Clutton-Brock TH (2002) Breeding together: kin selection and
mutualism in cooperative vertebrates. Science 296:69–72. doi: 10.1126/science.296.5565.69
Clutton-Brock TH, Russell AF, Sharpe LL, Brotherton PNM, McIlrath GM, White S et al (2001) Effects of helpers on juvenile development and survival in meerkats. Science 293:2446–2449. doi:10.1126/science.1061274
Clutton-Brock TH, Hodge SJ, Spong G, Russell AF, Jordan NR, Bennett NC, Sharpe LL, Manser MB (2006) Intrasexual com- petition and sexual selection in cooperative mammals. Nature 444:1065–1068. doi:10.1038/nature05386
Colmenares F, Hofer H, East ML (2000) Greeting ceremonies in baboons and hyenas. In: Aureli F, de Waal FBM (eds) Natural conflict resolution. University of California Press, California, pp 94–96
Conradt L, Roper TJ (2005) Consensus decision making in animals. Trends Ecol Evol 20:449–456. doi:10.1016/j.tree.2005.05.008 Cools AKA, van Hout AJM, Nelissen MHJ (2008) Canine rec-
onciliation and third-party-initiated postconflict affiliation: do peacemaking social mechanisms in dogs rival those of higher primates? Ethology 113:53–63
Cordoni G, Palagi E (2008) Reconciliation in wolves (Canis lupus): new evidence for a comparative perspective. Ethology 114:298– 308. doi:10.1111/j.1439-0310.2008.01474.x
Das M (2000) Conflict management via third parties: post-conflict affiliation of the aggressor. In: Aureli F, de Waal FBM (eds) Natural conflict resolution. University of California Press, California, pp 263–280
Dugatkin LE (1997) Cooperation among animals. Oxford University Press, Oxford
Emery NJ, Seed AM, von Bayern AM, Clayton NS (2007) Cog- nitive adaptations to social bonding in birds. Philos Trans R Soc Lond B Biol Sci 362:489–505. doi:10.1098/rstb.2006.1991 Emlen ST (1997) Predicting family dynamics in social vertebrates. In:
Krebs JR, Davies NB (eds) Behavioural ecology: an evolutionary approach, 4th edn. Blackwell Scientific, Oxford, pp 228–253 Engh AL, Siebert ER, Greenberg DA, Holekamp KE (2005) Pat-
terns of alliance formation and post-conflict aggression indicate spotted hyenas recognize third party relationships. Anim Behav 69:209–217. doi:10.1016/j.anbehav.2004.04.013
Flack JC, de Waal F (2007) Context modulates signal meaning in primate communication. Proc Natl Acad Sci USA 104:1581– 1586. doi:10.1073/pnas.0603565104
Flack JC, Girvan M, de Waal FBM, Krakauer DC (2006) Policing stabilizes construction of social niches in primates. Nature 439:426–429. doi:10.1038/nature04326
Frank SA (1998) Foundations of social evolution. Princeton Uni- versity Press, Princeton
Fraser ON, Stahl D, Aureli F (2008) Stress reduction through consolation in chimpanzees. Proc Natl Acad Sci USA 105:8557–8562. doi:10.1073/pnas.0804141105
Griffin AS, West SA (2002) Kin selection: fact and fiction. Trends Ecol Evol 17:15–21. doi:10.1016/S0169-5347(01)02355-2 Hamilton WD (1964) The genetical evolution of social behaviour I
and II. J Theor Biol 7:1–16, 17–52. doi:10.1016/0022-5193 (64)90038-4
Hatchwell BJ, Komdeur J (2000) Ecological constraints, life history traits and the evolution of cooperative breeding. Anim Behav 59:1079–1086. doi:10.1006/anbe.2000.1394
Harcourt AH, de Waal FBM (1992) Coalitions and alliances in humans and other animals. Oxford University Press, Oxford Helms Cahan S, Blumstein DT, Sundstro¨m L, Liebig J, Griffin A
(2002) Social trajectories and the evolution of social behavior. Oikos 96:206–216. doi:10.1034/j.1600-0706.2002.960202.x Hinde RA (1976) Interactions, relationships, and social structure.
Man (Lond) 11:1–17. doi:10.2307/2800384
Hirsch BT (2002) Social monitoring and vigilance behavior in brown capuchin monkeys (Cebus apella). Behav Ecol Sociobiol 52:458–464. doi:10.1007/s00265-002-0536-5
Hoeksema JD, Schwartz MW (2001) Modelling interspecific mu- tualisms as biological markets. In: Kappeler PM, van Schaik CP (eds) Cooperation in primates and humans: mechanisms and evolution. Springer, Berlin, pp 173–183
Isbell LA, Young TP (2002) Ecological models of female social relationships in primates: similarities, disparities, and some directions for future clarity. Behaviour 139:177–202. doi: 10.1163/156853902760102645
Issa FA, Edwards DH (2006) Ritualized submission and the reduction of aggression in an invertebrate. Curr Biol 16:2217– 2221. doi:10.1016/j.cub.2006.08.065
Johnstone RA (2000) Models of reproductive skew: a review and synthesis. Ethology 106:5–26. doi:10.1046/j.1439-0310.2000. 00529.x
Judge PG, Mullen SH (2005) Quadratic postconflict affiliation among bystanders in a hamadryas baboon group. Anim Behav 69:1345–1355. doi:10.1016/j.anbehav.2004.08.016
Kappeler PM (1993) Reconciliation and post-conflict behavior in ringtailed lemurs, Lemur catta, and redfronted lemurs, Eulemur fulvus rufus. Anim Behav 45:901–915. doi:10.1006/anbe.1993. 1110
Korb J, Heinze J (2008) Ecology of social evolution. Springer, Heidelberg
Koski SE, Koops K, Sterck EHM (2007) Reconciliation, rela- tionship quality and post-conflict anxiety: testing the integrated hypothesis in captive chimpanzees. Am J Primatol 69:158–172. doi:10.1002/ajp.20338
Koski SE, Sterck EHM (2007) Triadic postconflict affiliation in captive chimpanzees: does consolation console? Anim Behav 73:133–142. doi:10.1016/j.anbehav.2006.04.009
Krause J, Croft DP, James R (2007) Social network theory in the behavioural sciences: potential applications. Behav Ecol So- ciobiol 62:15–27. doi:10.1007/s00265-007-0445-8
Krause J, Ruxton GD (2002) Living in groups. Oxford University Press, Oxford
Kutsukake N (2000) Matrilineal rank inheritance varies with absolute rank in Japanese macaques. Primates 41:321–335. doi: 10.1007/BF02557601
Kutsukake N (2003) Assessing relationship quality and social anxiety among wild chimpanzees using self-directed behaviour. Behaviour 140:1153–1171. doi:10.1163/156853903322589687 Kutsukake N (2006) The context and quality of social relationships
affect vigilance behaviour in wild chimpanzees. Ethology 112:581–591. doi:10.1111/j.1439-0310.2006.01200.x
Kutsukake N (2007) Conspecific influences on vigilance behaviour in wild chimpanzees. Int J Primatol 28:907–918. doi:10.1007/ s10764-007-9156-2
Kutsukake N, Castles DL (2001) Reconciliation and variation in postconflict stress in Japanese macaques (Macaca fuscata fus- cata): testing the integrated hypothesis. Anim Cogn 4:259–268. doi:10.1007/s10071-001-0119-2
Kutsukake N, Castles DL (2004) Reconciliation and post-conflict third-party affiliation among wild chimpanzees at the Mahale Mountains, Tanzania. Primates 45:157–165. doi:10.1007/ s10329-004-0082-z
Kutsukake N, Clutton-Brock TH (2006a) Aggression and sub- mission reflect reproductive conflict between females in coop- eratively breeding meerkats. Behav Ecol Sociobiol 59:541–548. doi:10.1007/s00265-005-0079-7
Kutsukake N, Clutton-Brock TH (2006b) Social function of al- logrooming in cooperatively breeding meerkats. Anim Behav 72:1059–1068. doi:10.1016/j.anbehav.2006.02.016
Kutsukake N, Clutton-Brock TH (2008a) The number of subor- dinates moderates intra-sexual competition among males in cooperatively breeding meerkats. Proc R Soc Lond B Biol Sci 275:209–216. doi:10.1098/rspb.2007.1311
Kutsukake N, Clutton-Brock TH (2008b) Do meerkats engage in conflict management following aggression? Reconciliation, submission, and avoidance. Anim Behav 75:1441–1453 Kutsukake N, Hasegawa T (2005) Dominance turnover between an
alpha and a beta male and dynamics of social relationships in Japanese macaques. Int J Primatol 26:775–800. doi:10.1007/ s10764-005-5308-4
Kutsukake N, Nunn CL (2006) Comparative tests of reproductive skew in male primates: the roles of demographic factors and incomplete control. Behav Ecol Sociobiol 60:695–706. doi: 10.1007/s00265-006-0213-1
Kutsukake N, Nunn CL (2009) The causes and consequences of reproductive skew in male primates. In: Hager R, Jones CB (ed) Reproductive skew in vertebrates: proximate and ultimate factors. Cambridge University Press, Cambridge (in press)
Kutsukake N, Suetsugu N, Hasegawa T (2006) Pattern, distribu- tion, and function of greeting behaviour among black-and- white colobus. Int J Primatol 27:1271–1291. doi:10.1007/ s10764-006-9072-x
Ligon JD, Burt DB (2004) Evolutionary origins. In: Koenig WD, Dickinson JL (eds) Ecology and evolution of cooperative breeding in birds. Cambridge University Press, Cambridge, pp 5–34
Lung MA, Childress MJ (2007) The influence of conspecifics and predation risk on the vigilance of elk (Cervus elaphus) in Yel- lowstone National Park. Behav Ecol 18:12–20. doi:10.1093/ beheco/arl066
Maestripieri D, Schino G, Aureli F, Troisi A (1992) A modest proposal: displacement activities as an indicator of emotions in primates. Anim Behav 44:967–979. doi:10.1016/S0003-3472 (05)80592-5
Marler P (1996) Social cognition: are primates smarter than birds? In: Nolan V, Ketterson ED (eds) Current ornithology. Plenum Press, New York, pp 1–32
Matsumura S, Hayden TJ (2006) When should signals of submis- sion be given?-A game theory model. J Theor Biol 240:425–433. doi:10.1016/j.jtbi.2005.10.002
Maynard Smith J (1964) Group selection and kin selection. Nature 201:1145–1147. doi:10.1038/2011145a0
Maynard Smith J, Szmathma´ry E (1995) The major transitions in evolution. Oxford University Press, Oxford
McGregor PK (2005) Animal communication networks. Cam- bridge University Press, Cambridge
Monclu’s R, Ro¨del HG (2008) Different forms of vigilance in re- sponse to the presence of predators and conspecifics in a group- living mammal, the European rabbit. Ethology 114:287–297. doi:10.1111/j.1439-0310.2007.01463.x
Noe¨ R (1992) Alliance formation among male baboons: shopping for profitable partners. In: Harcourt AH, de Waal FBM (eds) Coalitions and alliances in humans and other animals. Oxford University Press, New York, pp 285–321
Noe¨ R, Hammerstein P (1994) Biological markets: supply and demand determine the effect of partner choice in cooperation, mutualism and mating. Behav Ecol Sociobiol 35:1–11. doi: 10.1007/BF00167053
Noe¨ R, Hammerstein P (1995) Biological markets. Trends Ecol Evol 10:336–339. doi:10.1016/S0169-5347(00)89123-5
Noe¨ R, van Schaik CP, van Hooff JARAM (1991) The market effect: an explanation for pay-off asymmetries among collabo- rating animals. Ethology 87:97–118
Nishida T (1983) Alpha status and agonistic alliance in wild chimpanzees (Pan troglodytes schweinfurthii). Primates 24:318– 336. doi:10.1007/BF02381978
Palagi E, Cordoni G, Tarli SB (2006) Possible roles of consolation in captive chimpanzees (Pan troglodytes). Am J Phys Anthropol 129:105–111. doi:10.1002/ajpa.20242
Petit O, Thierry B (2000) Do impartial interventions occur in monkeys and apes. In: Aureli F, de Waal FBM (eds) Natural conflict resolution. University of California Press, California, pp 267–280
Preuschoft S, van Schaik CP (2000) Dominance and communica- tion: conflict management in various social settings. In: Aureli F, de Waal FBM (eds) Natural conflict resolution. University of California Press, California, pp 77–105
Pusey AE, Packer C (1997) The ecology of relationships. In: Krebs JR, Davies NB (eds) Behavioural ecology: an evolutionary approach, 4th edn. Blackwell Scientific, Oxford, pp 254–283 Roeder JJ, Fornasieri I, Gosset D (2002) Conflict and postconflict
behaviour in two lemur species with different social organiza- tions (Eulemur fulvus and Eulemur macaco): a study on captive groups. Aggress Behav 28:62–74. doi:10.1002/ab.90006 Rolland N, Roeder JJ (2000) Do ringtailed lemurs (Lemur catta)
reconcile in the hour post-conflict?: a pilot study. Primates 41:223–227. doi:10.1007/BF02557804
Romero T, Aureli F (2008) Reciprocity of support in coatis (Nasua nasua). J Comp Psychol 122:19–25. doi:10.1037/0735-7036. 122.1.19
Ross KG (2001) Molecular ecology of social behaviour: analyses of breeding systems and genetic structure. Mol Ecol 10:265–284. doi:10.1046/j.1365-294x.2001.01191.x
Samuels A, Flaherty C (2000) Peaceful conflict resolution in the sea? In: Aureli F, de Waal FBM (eds) Natural conflict resolu- tion. University of California Press, California, p 229
Schaffner CM, Aureli F, Caine NG (2005) Following the rules: why small groups of tamarins do not reconcile conflicts. Folia Pri- matol (Basel) 76:67–76. doi:10.1159/000083614
Schaffner CM, Caine NG (2000) Peacefulness in cooperatively breeding primates. In: Aureli F, de Waal FBM (eds) Natural conflict resolution. University of California Press, California, pp 155–169
Schaffner CM, French JA (1997) Group size and aggression:
‘recruitment incentives’ in a cooperatively breeding primate. Anim Behav 54:171–180. doi:10.1006/anbe.1996.0413
van Schaik CP (1983) Why are diurnal primates living in groups? Behaviour 87:120–144. doi:10.1163/156853983X00147 van Schaik CP (1989) The ecology of female social relationships.
In: Standen V, Foley R (eds) Comparative socioecology. Blackwell Scientific, Oxford, pp 195–218
Schino G (1998) Reconciliation in domestic goats. Behaviour 135:343–356
Schino G (2000) Beyond the primates—expanding the reconcilia- tion horizon. In: Aureli F, de Waal FBM (eds) Natural conflict resolution. University of California Press, California, pp 225– 242
Scott J (2000) Social network analysis: a handbook, 2nd edn. Sage, London
Searcy WA, Nowicki S (2005) The evolution of animal communi- cation: reliability and deception in signaling systems. Princeton University Press, Princeton
Seed AM, Clayton NS, Emery NJ (2007) Postconflict third-party affiliation in rooks, Corvus frugilegus. Curr Biol 17:152–158. doi:10.1016/j.cub.2006.11.025
Silk JB (2002) Using the ‘‘F’’ word in primatology. Behaviour 139:421–446. doi:10.1163/156853902760102735
Silk JB (2007) The adaptive value of sociality in mammalian groups. Philos Trans R Soc Lond 362:539–559. doi:10.1098/ rstb.2006.1994
Sterck EHM, Watts DP, van Schaik CP (1997) The evolution of female social relationships in nonhuman primates. Behav Ecol Sociobiol 41:291–309. doi:10.1007/s002650050390
Stevens JMG, Vervaecke H, de Vries H, van Elsacker L (2005) Peering is not a formal indicator of submission in bonobos (Pan paniscus). Am J Primatol 65:255–267. doi:10.1002/ajp.20113 Stopka P, Macdonald DW (1999) The market effect in the wood
mouse, Apodemus sylvaticus: selling information on reproduc- tive status. Ethology 105:969–982. doi:10.1046/j.1439-0310. 1999.00485.x
Tamaki N, Morisaka T, Taki M (2006) Does body contact con- tribute towards repairing relationships? The association be- tween flipper-rubbing and aggressive behavior in captive bottlenose dolphins. Behav Processes 73:209–215. doi:10.1016/j. beproc.2006.05.010
Tibbetts EA (2002) Visual signals of individual identity in the wasp Polistes fuscatus. Proc R Soc Lond B Biol Sci 269:1423–1428. doi:10.1098/rspb.2002.2031
Treves A (2000) Theory and method in studies of vigilance and aggregation. Anim Behav 60:711–722. doi:10.1006/anbe. 2000.1528
Ueda K, Wada K (1996) Allocleaning in an intertidal ocypodid crab, Macrophthalmus banzai. J Ethol 14:45–52. doi:10.1007/ BF02350091
Vehrencamp SL (1983a) A model for the evolution of despotic versus egalitarian societies. Anim Behav 31:667–682. doi: 10.1016/S0003-3472(83)80222-X
Vehrencamp SL (1983b) Optimal skew in cooperative societies. Am Zool 23:327–355
de Waal FBM (1982) Chimpanzee politics—power and sex among apes. The Johns Hopkins University Press, London
de Waal FBM, van Roosmalen A (1979) Reconciliation and con- solation among chimpanzees. Behav Ecol Sociobiol 5:55–66. doi:10.1007/BF00302695
de Waal FBM, Tyack PL (2003) Animal social complexity. Har- vard University Press, Cambridge
Wahaj SA, Guse KR, Holekamp KE (2001) Reconciliation in spotted hyena (Crocuta crocuta). Ethology 107:1057–1074. doi: 10.1046/j.1439-0310.2001.00717.x
Watts DP, Colmenares F, Arnold K (2000) Redirection, consolation, and male policing: how targets of aggression interact with bystanders. In: Aureli F, de Waal FBM (eds) Natural conflict resolution. University of California Press, California, pp 281–301 Weaver A (2003) Conflict and reconciliation in captive bottlenose dolphins, Tursiops truncatus. Mar Mamm Sci 19:836–846. doi: 10.1111/j.1748-7692.2003.tb01134.x
West SA, Pen I, Griffin AS (2002) Cooperation and competition between relatives. Science 296:72–75. doi:10.1126/science. 1065507
Westlund K, Ljungberg T, Borefelt U, Abrahamsson C (2000) Post-conflict affiliation in common marmosets (Callithrix jac- chus jacchus). Am J Primatol 52:31–46. doi:10.1002/1098-2345 (200009)52:1<31::AID-AJP3>3.0.CO;2-Z
Wey T, Blumstein D, Shen W, Jorda’n F (2008) Social network analysis of animal behaviour: a promising tool for the study of sociality. Anim Behav 75:333–344. doi:10.1016/j.anbehav.2007. 06.020
Whitehead H (1997) Analysing animal social structure. Anim Be- hav 53:1053–1067. doi:10.1006/anbe.1996.0358
Whitehead H (2008) Analyzing animal societies: quantitative methods for vertebrate social analysis. University of Chicago Press, Chicago
Whitehead H, Dufault S (1999) Techniques for analyzing verte- brate social structure using identified individuals: review and recommendations. Adv Stud Behav 28:33–74. doi:10.1016/ S0065-3454(08)60215-6
Whitham JC, Maestripieri D (2003) Primate rituals: the function of greetings between male guinea baboons. Ethology 109:847–859. doi:10.1046/j.0179-1613.2003.00922.x
Wilson EO (1975) Sociobiology: the new synthesis. Belknap Press of Harvard University Press, Cambridge
Wittig RM, Boesch C (2003) The choice of post-conflict interac- tions in wild chimpanzees (Pan troglodytes). Behaviour 140:1527–1559. doi:10.1163/156853903771980701
Young AJ, Carlson AA, Monfort SL, Russell AF, Bennett NC, Clutton-Brock T (2006) Stress and the suppression of subor- dinate reproduction in cooperatively breeding meerkats. Proc Natl Acad Sci USA 103:12005–12010. doi:10.1073/pnas. 0510038103