DOI: http://dx.doi.org/10.14947/psychono.34.9
条件づけにおける時間 II
―オペラント計時行動,および時間学習の神経機構―
藤 巻 峻
a, b・新 保 彰 大
a・松 井 大
a・
時 暁 聴
a・神 前 裕
a, c*
a 慶應義塾大学社会学研究科心理学専攻 b 日本学術振興会 c 慶應義塾大学先導研究センターRoles of temporal information in conditioning II:
Operant conditioning and neural substrates of timing
Shun Fujimaki
a, b, Akihiro Shimbo
a, Hiroshi Matsui
a, Xiaoting Shi
a, and Yutaka Kosaki
a, c*
a Department of Psychology, Keio University b Japan Society for the Promotion of Science c Advanced Research Centers, Keio University
In the previous article (Kosaki, Shi, Matsui, Shimbo, & Fujimaki, 2015) we reviewed studies regarding how animals represent temporal information in classical conditioning. In this article, we first review various issues pertaining to interval timing in operant conditioning, with the main goal of providing a concise summary of procedural and theoretical developments in interval timing research. In the remainder of the article, we will review recent empirical findings and theories about the neural substrates underlying timing mechanisms both in classical and operant conditioning, and discuss how the hippocampus and striatum might contribute to different aspects of temporal information processing during conditioning. We then argue that the potentially different timing mechanisms implemented by the hippocampus and the striatum, as evidenced by the recent discovery of time cells in the hippocampus and consistent findings regarding involvement of the striatum in interval timing, might each be considered to constitute a part of functionally dissociable multiple memory systems that have been described elsewhere in the literature, particularly in the context of spatial learning and the organisation of voluntary behaviour.
Keywords: operant conditioning, interval timing, striatal beat frequency model, striatum, hippocampus, multiple
memory systems は じ め に 本稿では,まず第1章において動物の学習場面で切り 離すことのできない「時間」という変数が,オペラント 条件づけの文脈においてどのように研究されてきたのか を紹介する。第 2章では,別稿(神前・時・松井・新 保・藤巻,2015)で紹介した古典的条件づけと,第1章 で紹介するオペラント条件づけ,双方の計時学習場面に おける神経基盤について概観する。これらを通じて, 様々な条件づけ学習の場面において時間という変数が果 たす役割の統合的な理解を試みるとともに,時間を扱う 条件づけ研究の今後の方向性を探っていく。 第1章 オペラント条件づけにおける時間 1.1 オペラント条件づけにおける計時行動研究 オペラント条件づけにおいて,時間という変数に関す る研究は,オペラント条件づけ研究それ自体の発展と歩 みをともにしてきた。その主たる理由は,個体の行動や それを制御する環境中の事象がともに,時間という共通 の枠組みの中で生じているということに求めることがで Copyright 2015. The Japanese Psychonomic Society. All rights reserved. * Corresponding author. Department of Psychology, Keio
University, 2–15–45 Mita, Minato-ku, Tokyo, 108–8345, Japan. E-mail: [email protected]
きるだろう。
条件づけ研究で扱われる時間は,秒から分単位までの 比較的短い時間間隔である。このような短い時間間隔に おける動物個体の反応はインターバル計時と呼ばれ,概 日リズムなど周期的な内因性のリズムとは区分されてい る(Carr & Wilkie, 1997; Gibbon, Fairhurst, & Goldberg, 1997)。このインターバル計時に関する手続きは当座的 計時(immediate timing),回顧的計時(retrospective tim-ing),展望的計時(prospective timing)という3つに分類 することができる(Killeen & Fetterman, 1988)。これらの 手続きのうち,展望的計時に相当する手続きは計時行動 研究というよりも選択行動研究の文脈で主に用いられて きたという背景があるため,本稿では説明を省略する。 以下の節ではまず,当座的計時と回顧的計時における代 表的な手続きを概説するとともに,そうした計時場面に おける動物の行動を予測すべく提唱されてきた,いくつ かの計時理論について簡単に紹介する。 1.2 当座的計時 当座的計時とは,現に経過中の時間を手がかりとして 何らかの反応を行うような場面を指す。このような場面 では現在経過中の時間を関数とした反応パターンの様相 が主要な従属変数として扱われる。以下では代表的な当 座的計時の手続きを紹介する。 1.2.1 固定時隔スケジュール 計時行動に関する初 期の研究から現在に至るまで,最も広く使われている強 化スケジュールが固定時隔(fixed interval: 以下FIとす る)スケジュールである。FIスケジュールとは,強化子 呈示終了後から一定のインターバルが経過した後の初発 反応に対してのみ強化子が呈示され,それ以前に行われ た反応には強化子が随伴しないというものである。この スケジュールのもとでは,FIスキャロップという特徴的 な反応パターンが見られる(例えば Ferster & Skinner, 1957)。これは強化子呈示直後には反応休止期間が続き, 時間経過とともに加速的に反応率が上昇していく反応パ ターンである。 Schneider (1969)は,このFIスケジュール下で長い間 訓練したハトのキイつつき反応のパターンを精緻に観察 し,ブレーク・アンド・ランという特徴的なパターンを 発見した。これは,一定期間の低反応率時期が続いた後 に急激な高反応率時期が始まるというものである。この パターンにおいて,低反応率から高反応率への切り替わ り時点はブレークポイントと呼ばれている。Schneider (1969) の 実 験 で は,16 秒 か ら 512 秒 ま で の FI ス ケ
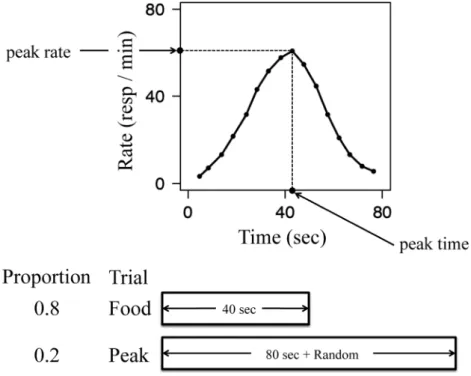
Figure 1. A typical peak function (top panel) obtained by the peak-interval procedure with FI 40-s training (bottom panel). On food trials, the first responding after 40 s results in the food presentation. On peak trials, the stimulus presentation is prolonged for 80 s plus random time, during which food presentation is omitted (re-plotted from Roberts, 1981, Figure 1).
ジュールでこのブレークポイントを求めたところ,イン ターバルの絶対的な長さには関わらず,インターバル開 始からおよそ3分の2の時点にブレークポイントが集中 することを見出している。 1.2.2 ピーク法 ピーク法は上述のFIスケジュール を離散試行型にしたものであり,強化試行(food trial) と強化子が呈示されない試行であるピーク試行で構成さ れる(Catania, 1970; Roberts, 1981)。強化試行では,通常 のFIスケジュールと同様に刺激呈示から一定時間経過 後の最初の反応に対して強化子が呈示される。これに対 し,ピーク試行では強化子が呈示されず,あらかじめ設 定された時間が経過した時点で自動的に試行が終了す る。ピーク試行の持続時間は,通常,強化試行の3倍程 度に設定されることが多い。また,強化試行とピーク試 行では同一の弁別刺激が用いられるため,個体が試行を 弁別することはできないようになっている(Figure 1, bottom panel)。 この手続きでは,ピーク試行において反応率が頂点に 達する時点(ピーク時間)や,経過時間に伴う反応率の 変化が主要な従属変数である。ピーク試行では,通常の FIスケジュールと同様に,試行開始からの時間経過に伴 い次第に反応率が上昇していくパターンが見られるが, 強化試行で用いられるインターバルが経過した後は次第 に反応率が減少していき,典型的には FI時間を頂点と した正規分布のような山型の曲線を描く(Figure 1, top panel)。 1.3 回顧的計時 回顧的計時とは,過去に呈示された刺激の持続時間を 手がかりとして反応を行うような場面を指す。この場面 における主な手続きとしては,刺激の持続時間を継時的 に弁別させる Go/No-go型課題,持続時間を相対的に弁 別させる課題,そして間隔二分手続きが挙げられる。上 述した当座的計時場面では,現在において経過中の時間 に基づく反応パターンの様相が主な従属変数となってい た。これに対し,刺激持続時間に関する弁別課題では, 過去に呈示された刺激の持続時間を手がかりとした個体 の反応が従属変数となる。 1.3.1 Go/No-go 型課題と相対的持続時間弁別課題 最も単純な刺激持続時間の弁別課題としては,Church & Gibbon (1982)による Go/No-go 型の手続きが挙げられ る。彼らの実験では,刺激がある持続時間で呈示された 場合(以下,正刺激とする)に被験体であるラットがレ バー押し反応を行えば,強化子として が呈示された。 その一方で,正刺激とは異なる持続時間(以下,負刺激 とする)で刺激が呈示された場合には,レバー押し反応 を行っても強化子は呈示されなかった。例えば4秒を正 刺激とした場合,刺激の持続時間がそれよりも長い場合 や短い場合には反応しても強化子は呈示されず,刺激が 4秒間呈示された場合の反応に対してのみ,強化子が呈 示された。この手続きでは,横軸に様々なテスト刺激の 持続時間,縦軸に反応確率を取ることで,刺激の持続時 間に対する般化勾配を得ることができる。すなわち,般 化勾配の頂点やその広がりが主要な従属変数となる。
Church & Gibbon (1982)以降の研究では,2種類の刺 激の相対的な持続時間を弁別させる課題もしばしば用い られている(例えば Dreyfus, Fetterman, Smith, & Stubbs, 1988; Stubbs et al., 1994)。ハトを被験体とした一般的な 手続きでは,例えば赤刺激をt1秒呈示した後に緑刺激を t2秒呈示し,赤刺激の持続時間が長かった場合は一方の 選択肢へのキイつつき反応が,その逆の場合は他方の選 択肢へのキイつつき反応が強化される。この課題では弁 別の正確さが主な従属変数となる。 1.3.2 間隔二分課題 回顧的計時の手続きとして最 も広く用いられてきた手続きが,間隔二分課題である (Church & Deluty, 1977; Platt & Davis, 1983; Stubbs, 1968)。
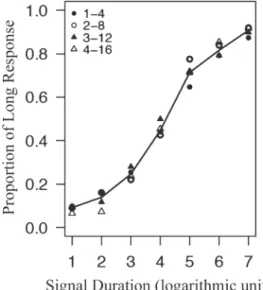
Figure 2. A psychometric function obtained by the temporal bisection task (re-plotted from Church & Deluty, 1977, Figure 2). The proportion of “long” re-sponse is plotted as a function of the durations of test stimuli represented in normalised logarithmic scale. Each symbol represents different training condition using a particular set of stimulus; 1 vs. 4, 2 vs. 8, 3 vs. 12, and 4 vs. 16 s (see text for further details). Line segments connect the medians of four conditions.
Church & Deluty (1977)の実験では,ラットが被験体と して用いられ,2種類の刺激持続時間を弁別する訓練が 行われた。呈示された刺激が短い(例えば2秒)場合に は左レバー,長い(例えば8秒)場合には右レバーに反 応すれば,強化子として が呈示された。弁別獲得後の テストでは,長短それぞれの訓練刺激の間に属する複数 の持続時間を持った刺激が呈示され,それに対する左右 のレバーへの反応割合が算出された。この反応割合が 50%となる刺激持続時間,すなわち二分点(bisection point)がこの課題の主要な従属変数となる(この二分 点は主観的等価点に相当する)。彼らの実験では4種類 の刺 激 持 続 時 間 の 組 み 合 わ せ(1 秒–4 秒,2 秒–8 秒, 3 秒–12 秒,4 秒–16 秒)で弁別訓練を行ったが,いず れの組み合わせでも二分点は長短それぞれの刺激持続 時間の幾何平均に近くなることが明らかとなっている (Figure 2)。
Stubbs (1968)はハトを被験体として用い,Church & Deluty (1977)とは異なる方法で間隔二分課題を行って いる。Stubbs (1968)の手続きでは,1秒から10秒まで を1秒間隔で区切り,計10種類の刺激持続時間が用意さ れた。これらのうち,1秒から5秒までのいずれかの刺 激が呈示された場合は一方のキイ(短選択肢)を,6秒 から10秒までのいずれかの刺激が呈示され場合は他方 のキイ(長選択肢)を選べば によって強化された。こ の実験では横軸に対数化した刺激持続時間を取り,縦軸 には長選択肢を選択した割合をプロットした結果,シグ モイド型の関数を描くことが示された。 1.4 スカラー性 当座的計時,回顧的計時の両場面を通じて,計時行動 に関する様々な特性が多くの研究によって報告されてき た。その中でもとりわけ重要視されてきたのがスカラー 性(scalar property)である。スカラー性とは,インター バル開始からの経過時間と計時行動の指標をそれぞれ相 対化したうえでプロットした際に,様々な長さのイン ターバルのもとで得られた指標が重なりあうことであ る。例えばDews (1970)では,FI30秒,FI300秒,FI3000 秒という 3つの条件でハトのキイつつき反応を訓練し た。そして,各条件におけるインターバル開始からの経 過時間を相対化して横軸に,最大反応率で相対化した反 応率を縦軸にとってプロットした結果,条件間で反応パ ターンが重なり合うことが見出された。1.2.2項で紹介 したピーク法で得られる反応分布に関しても,ピーク時 間とピーク反応率に対する相対値としてプロットすると スカラー性が見られる。スカラー性はこうした当座的計 時場面に限らず,回顧的計時場面である間隔二分課題で も成り立つことが知られている(Gibbon, 1981)。 1.5 計時行動に関する諸理論 動物の計時行動に関するモデルはこれまで数多く提唱 されてきた。Wearden (1999)によれば,それらの多く は個体の内部に体内時計(internal clock)の存在を仮定 している点で共通している。こうした体内時計モデル は,計時部,記憶部,比較・決定部という3つの要素で 構成されている点は理論間で共通しているものの,特に 計時部における仮定がモデル間で大きく異なっている。 具体的に言えば,「個体が何を手がかりにして計時を 行っているか」という仮定に,モデル間の主要な違いを 求めることができる。こうした計時部における仮定の違 いから,計時に関するモデルを1)ペースメーカ–アキュ ムレータタイプ(pacemaker–accumulator type),2)減衰 過 程 タ イ プ(process-decay type),3)発振器同時検出 タイプ(oscillator/coincidence detection type)という3つ のタイプに分類することが可能である(Matell & Meck, 2000)。 1.5.1 ペースメーカ–アキュムレータタイプ ペース メーカ−アキュムレータタイプでは,個体内にペース メーカが存在すると仮定しており,そのペースメーカが 発するパルス数が計時の手がかりになっているとされ る。このペースメーカ–アキュムレータモデルにおいて 最も強い影響力を誇ってきた理論は,スカラー期待理論 (scalar expectancy theory: Gibbon, 1977)を計時行動に応 用したスカラー計時理論(Gibbon, 1991; Gibbon, Church, & Meck, 1984)である(以下SETとする)。
SETでは,動物個体が訓練を繰り返すことで参照記憶 内に計時開始から強化子呈示までの時間が蓄積パルス数 の確率分布として表現される。計時場面では,この参照 記憶内の分布から1つのパルス数が抽出され,それが計 時開始とともにアキュムレータに蓄積されていくパルス 数と比較され,これらのパルス数の差分が一定の閾値に 達すると反応が生起すると仮定される。 このようにSETが記憶表象を重視したモデルであるの
に対し,Killeen & Fetterman (1988)による行動的計時理 論(behavioral theory of timing)では,個体が行う行動を 計時の手がかりとして重視している。このモデルでは, ある間隔で強化子が呈示されるような実験事態において 個体が自発する行動(付随行動)を行動ステートと呼ん でいる。行動ステートは時間経過とともに次のステート へと移行していくため,個体は自身の現在の行動ステー トがどのような状態にあるかを手がかりに,計時を行う
ことができると仮定されている。行動的計時理論では, ステート間の移行は個体内のペースメーカが発するパル スによって決定されると仮定しているため,ペースメー カ–ア キ ュ ム レ ー タ タ イ プ に 分 類 さ れ る。Machado (1997)は,この行動ステートという考え方を継承しつ つ,行動的計時理論をさらに発展させた計時学習モデル (learning-to-time model)を提唱している。 1.5.2 減衰過程タイプ 上述したモデルは個体内に ペースメーカが存在することを仮定しており,そのペー スメーカが発するパルス数で計時行動を説明するという 点で共通していた。これに対し,減衰過程タイプのモデ ルでは,計時開始を示す何らかの事象や刺激変化(タイ ムマーカ)が起こった後,時間経過に伴い記憶痕跡や神 経活動の減衰が生じると考える。減衰過程モデルでは, こうした減衰過程が計時部の役割を担っており,個体が 計時を行う際の手がかりになると仮定されている。減衰 過程タイプに属する代表的なモデルとしては,Staddon & Higa (1999)による多重時間尺度モデル(multiple time scale model)が挙げられる。そのほかにも,Grossberg & Schmajuk (1989)によるスペクトル計時モデル(spectral timing model)もこの減衰過程タイプとして位置づけら れることがある(Matell & Meck, 2000)。ただしMatell & Meck (2004)では,次項で述べる発振器同時検出タイプ と対比して,これらのモデルをともにマルチニューロン タイプとして扱っている。
1.5.3 発振器同時検出タイプ 発振器同時検出タイ
プのモデルとしては,Church & Broadbent (1990)による 多重発振器モデル(multiple oscillator model)がよく知ら れている。このモデルは異なる周期で発振を繰り返す複 数の発振器を仮定したコネクショニストモデルである。 しかしながら,多重発振器モデルは神経機構からみた妥 当性を欠いているとMatell & Meck (2000)により指摘さ れており,その問題点を克服するモデルとして線条体振 動頻度モデル(striatal beat frequency model: 以下SBFとす る)が彼らにより提唱されている(Matell & Meck, 2004 も参照)。このモデルでは,各細胞による短い周期の独 立した発振が統合されることによって生じる長い発振周 期が,計時の手がかりとして機能すると仮定している。 その具体的なメカニズムに関しては,2章で詳述する。 1.6 結論 本章では,オペラント条件づけにおける計時行動の手 続きやそのもとで確認されてきた現象について紹介する とともに,計時に関わる様々な現象を包括的に説明すべ く提唱されてきた理論について簡単に紹介した。本稿で 扱った内容はオペラント条件づけにおける膨大な計時行 動研究の中のごく一部であり,扱いきれていない問題も 数多く存在する。特に各モデルの挙動や今後の方向性な どについては考察しきれていない。これらの問題につい ては,石井(2008a, 2008b)による包括的な総説を参照 されたい。 第2章 条件づけにおける時間の神経機構 本章では,前章までで紹介した様々な条件づけ課題に おいて時間情報の符号化および表出を担う神経機構を探 ることを目的とする。紙数の制限から,ここでは海馬と 線条体の時間学習における機能に焦点を絞って議論を進 める。より包括的な議論は,以下の総説を参照されたい (Buhusi & Meck, 2005; MacDonald, 2014; MacDonald, Fortin,
Sakata, & Meck, 2014; Meck, Church, & Matell, 2013)。
2.1 時間表象の神経基盤
2.1.1 時間細胞 近年,海馬 CA1 の神経細胞群が
時間経過を表現していることを示す研究がMacDonaldら によって報告されている。MacDonald, Lepage, Eden, & Eichenbaum (2011)は,ラットに走路上の異なる地点で 10秒の遅延を挿んで物体と匂い刺激を継時的に呈示し, これらの組み合わせによってGo反応またはNo-Go反応 を要求する課題を行った。ラットは 10秒の遅延の間, 最初の物体が何であったかを記憶しておく必要があっ た。課題遂行中に海馬 CA1の錐体細胞活動を記録する と,遅延中のそれぞれの時間に特異的に発火する細胞が 見つかった。これらの細胞活動を時間順に並べ替える と,10秒の遅延期間を順々に埋めていくような,すな わち時間の経過を表現するような活動を細胞集団として 行っていることが見いだされた。MacDonaldらはこれら の細胞を,空間内の特定の位置を符号化する場所細胞に なぞらえて「時間細胞」と呼んだ。このような時間特異 的な活動は,空間情報の関与を除外するためにラットの 頭部を固定した課題においても確認されている(Mac-Donald, Carrow, Place, & Eichenbaum, 2013)。
時間細胞の特徴としては,遅延時間の変化や,試行が どの刺激によって開始されたかによっても個々の細胞の 反応パターンが変化することが挙げられる。すなわち, これらの細胞は無差別に時間の経過を表現するのではな く,ある特定の事象間の決まった時間間隔を一つのまと まりとして符号化し,その中における時間経過を表現し ていると考えられる。こうした課題の変化に対する個々 の細胞の「時間受容野」の変化は,空間内の刺激配置あ るいは課題要求の変化に伴って場所細胞の場所受容野が
変化する再配置(remap)とよく似ており,MacDonald らはこれを時間的再配置(retime)と呼んでいる。さら に,個々の細胞は遅延中の特定の時点に発火頻度分布の ピークを示すが,遅延の後半にピークを持つ細胞ほどそ のピークの分散が大きい,すなわちスカラー性を持つこ ともその特徴として挙げられる。また,場所細胞につい ては,隣り合う場所受容野を持つ細胞が θ 位相の異なる 位置で発火し,ラットがある細胞の受容野中心に近づく につれ当該細胞の θ 位相内での発火位置が前進する θ 位 相歳差(theta phase precession)と呼ばれる現象が知られ ている(O’Keefe & Recce, 1993)。この位相歳差現象は空 間的に隣り合う事象同士を連続的に結合する可塑性のも ととなるため,ナビゲーションにおける未来への予測を 含む時間順序情報の生起に関与することが示唆されてき たが(Johnson & Redish, 2007; Yamaguchi, 2003),現在の ところ時間細胞にこのような θ 位相歳差があることは確 認されていない。また場所細胞の θ 位相歳差と時間細胞 の特性との関連性についても明らかにはされていない。 こうした事象間の時間的欠損を埋めるような神経活動 が海馬で見られることは,後節で述べるように海馬が痕 跡条件づけにおいて重要であるという従来の損傷研究か らの知見(例えば Solomon, Vander Schaaf, Thompson, & Weisz, 1986)によく一致する。一方で,海馬の細胞がこ うした時間特異的な反応を生み出すのに必要な時間情報 は,どこで生み出されるのであろうか? この問題に関 してEichenbaum (2014)は,場所細胞の連鎖的な活性に よる結合を例にとり,それが海馬内部において生み出さ れる可能性を指摘する一方,皮質部のオシレーションパ ターンを線条体で統合して時間情報として符号化する線 条体振動頻度モデル(SBF: Matell & Meck, 2000)に触れ, 海馬外にその起源がある可能性を併記している。次節で は,このSBFについて記述する。 2.1.2 線条体振動頻度モデル(SBF) Miall (1989)が 提唱した振動頻度モデルでは,各細胞が比較的短い周期 で独立して発振する場合,これらを統合することで各周 期の公倍数にあたる長い発振周期を得ることができるた め,数秒から数十秒にわたる事象間隔のクロックとなり うることが示唆されてきた。Matell & Meck (2000)の SBFでは,この考えをもとに,線条体の中型有棘神経細 胞(medium spiny neuron; 以下MSNとする)に皮質細胞 における発振パターンを統合する同期検出器としての役 割を想定し,そこで検出されたパターンが強化子の呈示 に対応する黒質緻密部からのドーパミン入力によって 「強化」されるという機構をその基本としている。 より詳しく見て行くと,線条体のMSNは皮質からそ れぞれ1万から3万にのぼる数の入力を受けている。皮 質の細胞はそれぞれ異なる周期で発火しており,MSN においてはこれらのうち一定数以上の入力(150ほどと 見積もられる)が同時に生起したときのみ,細胞が発火 しやすい状態になると考えられる。これはすなわち
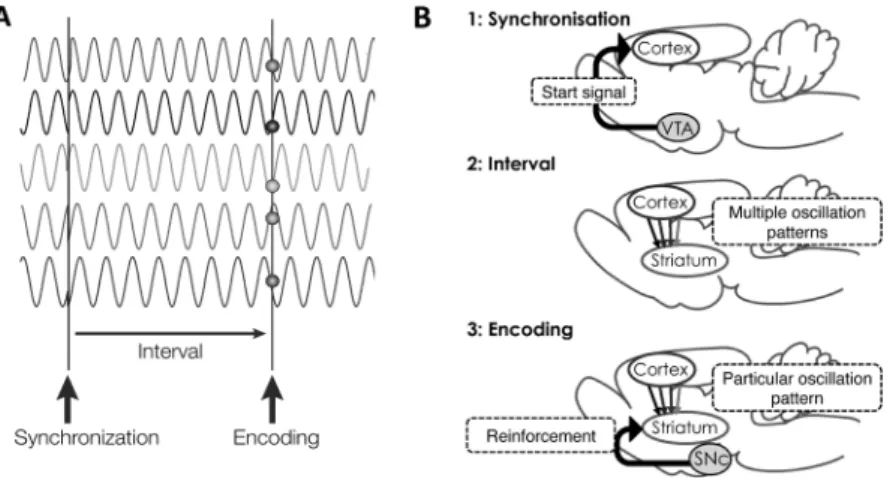
Figure 3. Schematic description of the SBF model. (A) After being synchronised by the ‘start’ signal, cortical neurons os-cillate independently at various rates and become progressively de-synchronised. At the time of reward presentation, the distinct pattern of multiple oscillations is encoded as a specific time record. (B) Neural system mediating the time encod-ing. Mesocortical dopaminergic projections provide ‘start’ signal to cortical neurons. MSNs in the striatum encode time by means of integrating cortical oscillation patterns. The phasic nigro-striatal dopaminergic transmission reinforces the time code represented in the MSNs (Panel A is adapted and modified from Buhusi & Meck, 2005, with permission of the pub-lisher).
MSNが各皮質細胞の発振周期を統合することでより長 い発振周期のクロックとして機能することを意味する。 強化子呈示による位相性のドーパミン伝達(Schultz, Dayan, & Montague, 1997)は,MSNにおける皮質入力近 傍へのシナプス結合を通じて,強化子が呈示された時点 での皮質細胞群の特定の活動パターン(すなわち時間符 号)をLTPによって選択的に「記録」することになる。 また,試行開始刺激の呈示によって黒質および腹側被蓋 野(ventral tegmental area; 以下VTA)のドーパミン細胞 が発火し,黒質からのドーパミン入力によりMSNの膜 電位が初期値にリセットされる一方,VTAから皮質へ の入力により皮質細胞群の発振がリセットされると仮定 される。その後,皮質の細胞群は独立にその位相がずれ て行くと想定される。これらにより,特定のMSNは一 度「強化」された後,皮質細胞の同じ活動パターンが起 きた時点,すなわち試行開始から同じ時間が経過した時 点でのみ発火する,入力フィルターとしての性質を獲得 することになる(Figure 3)。
Matell, Meck, & Nicolelis (2003)は,実際にMSNの活動 記録を行い,10秒と40秒の2ピーク法課題において,あ る細胞は10秒に活動ピークを,別の細胞は40秒に活動 ピークを示すことを見いだした。また Matell & Meck (2004)は,SBFによるシミュレーションを用いてMSN の活動にスカラー性が生じることを報告している。これ らの結果から,SBFは計時学習の神経モデルとしての妥 当性が高いといえる。 2.2 計時に関する損傷,薬理研究―当座的計時場面― これまで述べた海馬における時間特異的な神経活動や SBFによる予測は,海馬および線条体が計時学習に重要 であることを示唆しているが,損傷研究や薬理研究から もこの考えは概ね支持されている。ここでは1章で概説 した当座的および回顧的オペラント計時課題を中心とし て,それらにおける線条体と海馬の役割を中心に概観す る。 2.2.1 ピーク法 ピーク法を用いた研究からは,当 座的インターバル計時における線条体の役割が示されて いる(MacDonald, Cheng, & Meck, 2012; Meck, 2006b)。例
えばMeck (2006b) は,ピーク法による訓練後に黒質緻 密部もしくは背側線条体を損傷したラットでは反応分布 が平板化し,ピークが出現しなくなることを報告してい る。さらに黒質緻密部を損傷されたラットにドーパミン を含むカテコールアミンの前駆体であるL-DOPAを投与 すると反応のピークが出現したが,背側線条体を損傷し たラットではL-DOPA投与の効果は無く,反応は平板化 したままであった。この結果は,背側線条体のドーパミ ン受容体が当座的計時に必要であることを示唆してい る。また,ピーク時間に向けて反応が増加に転じる「開 始点」およびピーク時間が過ぎた後に反応が減少に転じ る「終止点」の出現が,それぞれ背側線条体と腹側線条 体の可塑性に依存することが,タンパク合成阻害剤アニ ソマイシンをそれぞれの部位に訓練前投与した実験から 示されている(MacDonald et al., 2012)。 一方ピーク法における海馬の役割に関しては,Meck, Church, & Olton (1984)が,海馬采–脳弓経路(fimbria– fornix; 以下FFXとする)を切断したラットでピーク時間 が早まることを初めて報告し,その後同様の結果が数多 く報告されている(Balci, Meck, Moore, & Brunner, 2009; Meck, 1988; Olton, Meck, & Church, 1987; Olton, Wenk, Church, & Meck, 1988; Yin & Meck, 2014: ただしDietrich & Allen, 1998も参照)。また古典的条件づけを用いたピー ク法においても,背側海馬の損傷がピーク時間を早める ことが報告されている(Tam & Bonardi, 2012; Tam, Jen-nings, & Bonardi, 2013)。
ピーク法におけるこうした海馬と線条体の役割は,互 いにどう関係しているのだろうか? Yin & Meck (2014) は,以下に述べるような研究から,海馬損傷の影響が線 条体への影響を介した副次的なものである可能性を提案 している。 彼らは背側海馬損傷マウス(背側の CA1–3 および 歯 状 回 を 含 む) と δ-オ ピ オ イ ド 受 容 体 欠 損 マ ウ ス (Oprd1−/−)において同様のピーク時間減少を認めた。 δ-オピオイド受容体は主に背側海馬の GABA 作動性神 経細胞に発現し,この細胞の抑制に関与する。また Oprd1−/−マウスでは海馬依存性の学習が障害され,線 条体依存の学習が促進されることから(Le Merrer, Rezai, Scherrer, Becker, & Kieffer, 2013),背側海馬損傷マウスお
よびOprd1−/−マウスにおけるピーク時間の減少は,海
馬から線条体への抑制の低下による線条体の過活動が原 因であるとしている(Yin & Meck, 2014)。
これを支持する例として,ピーク法課題中の背側海馬 CA1と背側線条体の局所活動電位(LFP)を記録した研 究において,30秒にピークを示すレバー押しとLFPの相 関が高かったのは CA1ではなく背側線条体であったこ とが示されている(Hattori & Sakata, 2014)。また,海馬 を含む側頭葉損傷患者において20秒までのインターバ ル計時が正常である一方(Shaw & Aggleton, 1994),回顧 的な時間間隔判断に基づく意思決定に変化がみられると いう報告(Melgire et al., 2005)からも,ピーク法におけ る海馬の副次的機能説は支持される。
またピーク法において刺激が数秒間中断されるギャッ プ試行を用いた研究からは,通常のラットではピーク位 置がギャップ期間に相当する時間だけ右側へシフトす る,すなわち作業記憶に保持されたギャップ前の刺激 呈示時間とギャップ後の刺激時間と合計することで強化 時間を予測するのに対し,FFX損傷(Meck, et al., 1984; Olton et al., 1987)あるいは海馬へのコリン性投射の主要 な起始部位である内側中隔野の 損傷(Meck, Church,
Wenk, & Olton, 1987)により,ピーク位置がギャップ期 間とギャップ前の刺激呈示期間を足し合わせた時間だけ 右側へシフトする,すなわちギャップ後に計時がリセッ トされることが示されている。こうした研究からは,海 馬がインターバル計時における反応ピークの産出そのも のではなく,計時における作業記憶の側面に関与するこ とが示唆される。 2.2.2 中枢作動薬の役割 当座的計時課題における 線条体の関与を示唆する研究として,ドーパミン作動薬 の投与がピーク時間に与える影響を調べた研究がある。 Meckらの一連の研究によって,ラットに対するメタン フェタミンあるいはコカインの全身投与がピーク法に おけるピーク時間を早めること(Buhusi & Meck, 2002; Maricq, Roberts, & Church, 1981; Matell, King, & Meck, 2004; Meck, 1996),またドーパミン拮抗薬の投与によってピー ク時間が後退すること(Buhusi & Meck, 2002; Meck, 1986) が明らかになっている。ピーク時間へのドーパミン作動 薬の影響は,詳細な生理学的メカニズムは不明ながら も,ピーク時間のシフト量が計時する時間の長さに対し て相対的であることや,幅広い計時課題において同様の 効果が見られることから,計時におけるクロック速度そ のものへの影響によると考えられている(Matell, Bates-on, & Meck, 2006; Meck, 1983, 1996)。また,ドーパミン作 動薬のクロック速度への効果は,前頭葉の損傷,あるい は前頭葉へコリン性投射を送る基底核大細胞部の損傷に よって消失するが,FFXの損傷あるいは海馬へコリン性 投射を送る内側中隔野の損傷では消失しないことが示さ れており(Meck, 2006a),前頭葉を含む皮質部位から線 条体への収束性入力によってクロックが生成されるとす るSBFの仮定を支持する結果であるといえる。 2.2.3 古典的条件づけにおける条件反応のタイミン グ 長い条件刺激(CS)を用いた遅延条件づけや痕跡 条件づけでは,無条件刺激(US)までの時間的距離に 基づいて条件反応(CR)が表出されることから,計時 機能のみに着目した場合,上で述べた当座的オペラント 計時との類似性を見いだすことができる。痕跡条件づけ は海馬の損傷によって成立しなくなることがよく知られ
て い る(例 え ば McEchron, Bouwmeester, Tseng, Weiss, & Disterhoft, 1998; Moyer, Deyo, & Disterhoft, 1990; Quinn, Oommen, Morrison, & Fanselow, 2002; Solomon et al., 1986) が,計時との関係でより重要なのはCRの獲得そのもの ではなく,CRのタイミングへの影響であろう。上記の 研究では電撃や目へ空気を吹き付けるといった嫌悪刺激 をUSとして用いているが(CRは瞬目反射,瞬膜反射, すくみ反応,活動量低下など),食 USを用いた痕跡条 件づけでは海馬を損傷してもCRを獲得すること自体は 可能であり(Jarrard & Davidson, 1990; Kyd, Pearce, Hasel-grove, Amin, & Aggleton, 2008),重要な点として,背側海 馬損傷により遅延条件づけおよび痕跡条件づけの両方で CRの表出時点がUSの到来が予測される時間よりも早く なる影響が示されている(Tam & Bonardi, 2012)。このこ とはインターバル計時に関して,オペラント条件づけと 古典的条件づけとで同様の神経回路による時間情報処理 がなされている可能性を示唆する。 2.3 計時に関する損傷,薬理研究―回顧的計時場面― ここでは間隔二分課題を中心に,回顧的計時における 神経基盤を概観する。これらの課題では共通して,過去 に経験した時間の長さに基づいて反応を表出することが 要求される。 2.3.1 間隔二分課題 間隔二分課題では,FFXを損傷 したラットで二分点が減少することが示されている (Meck et al., 1984)。しかし歯状回に限局した損傷では二 分点の位置に影響がないことが報告されており(Bueno & Bueno-Júnior, 2011),間隔二分課題には歯状回以外の 海馬領域が関与することが示唆される。 これに対し線条体の関与については,ドーパミンD2 受容体を線条体に過剰発現させたマウスにおいて,比較 刺激が2秒と8秒の持続音の場合には障害が見られない が,より長い刺激(6秒と24秒)を用いると弁別が低下 するという報告がある(Ward et al., 2009)。しかしこの 課題では長い刺激を用いた場合にのみ障害が見られ,さ らに呈示されるテスト刺激が長くなるに従って障害が大 きくなったことから,Wardらは線条体におけるD2受容 体過剰発現の影響を,計時そのものに対する障害ではな く,作業記憶あるいは持続的注意の障害であると解釈し ている。またドーパミン作動薬メタンフェタミンが二分 点をより短い方向へ,拮抗薬ハロペリドールがより長い 方向へ移動させることが示されており,ドーパミン投射 の主要な標的である線条体の関与が示唆される(Maricq
& Church, 1983; Meck, 1983)。ただし,これらの効果は時 間間隔の記憶への影響ではなく,投与時の個体内のク
ロック速度への影響によるとされる。
時間弁別における複数領域の関与を直接比較するため に,Onoda & Sakata (2006)はラットを用いて刺激間間 隔(0.5秒または2秒)の弁別課題を行い,その間の事象 関連電位(ERP)の計測を行った。その結果,線条体お よび前頭皮質,そして視床において時間弁別に関連する 活動が見られるが海馬では見られないことが明らかに なった。ただし,2秒と8秒の刺激間隔を用いて行われ た同様の弁別においては海馬の関与が示されている (Onoda, Takahashi, & Sakata, 2003; Sakata & Onoda, 2003)。
2.3.2 文脈的経過時間弁別 回顧的計時には,エピ ソード記憶のようにある事象が過去のどれくらい遠くの 時点で生じたかを弁別する事態も含まれる。これまで述 べた回顧的計時研究が主に数秒から数十秒の範囲での刺 激持続時間の弁別を要求したのに対し,文脈的経過時間 弁別では過去に事象が生じてから刺激の存在しない期間 がどれくらい経過したかを,数分から数十分のより長い 範囲に渡って弁別することが要求される。すでに別稿 で,古典的条件づけを用いた研究においてラットが数 分単位の ITI を弁別できることを紹介した(Bouton & Garcia-Gutierez, 2006; Bouton & Hendrix, 2011; Todd, Win-terbauer, & Bouton, 2010)。こうしたより長い時間の文脈 的経過時間判断に海馬が重要であることを示唆する結果 が,Jacobs, Allen, Nguyen, & Fortin (2013)によって報告 されている。 Jacobs et al. (2013)はラットを用いて,前試行からの 経過時間を弁別刺激として,その経過時間と結びついた 特定の匂い刺激を,2つの匂い刺激の中から選択させる 課題を行った。この課題において,ムシモル投与による 海馬の一時的非活性化は,2種類の匂い選択刺激と結び ついた時間の比が大きい場合(例えば1 : 8)には影響を もたらさないが,比が小さい場合(2 : 3)に課題遂行を 障害すること,さらにこの障害は比較時間の絶対値が大 きい場合(8分 : 12分)でのみ見られ,絶対値が小さい 場合(60秒 : 90秒)では逆に統制群に比べ弁別を促進す ることが明らかになった。これらの結果は,一般的に海 馬が類似した刺激パターンを分離することに重要である という理論(例えばTreves & Rolls, 1994)に一致し,さ らにより長い時間間隔は文脈性条件性刺激としての役割 を果たすことから(Bouton & Hendrix, 2011),海馬の文 脈刺激処理における役割とも一致する。ただし,比較時 間の絶対値が小さい場合の促進効果に関しては,海馬と 他の計時における責任部位,例えば線条体との機能的競 合が取り除かれた結果である可能性が考えられるもの の,詳細なメカニズムは不明である。いずれにせよ, Jacobsらの結果は回顧的計時に対する海馬の役割を明確 に示す重要なものであるといえる。 2.4 計時における線条体と海馬の役割: 今後の展望 これまで本章では計時行動における神経機構を,線条 体と海馬の役割を中心として簡単に見てきた。これらの 研究からは,詳細においては不明な点も多いながらも, ピーク法に代表される当座的計時手続きには線条体を中 心とした系が,間隔二分課題や経過時間弁別のような回 顧的計時手続きには海馬がより重要であるという傾向が 示されている。このことはより具体的には何を意味する のであろうか? 一つの見方は,線条体は当座的計時に必要とされる現 在時間の知覚に基づく予測的計時,海馬は過去の時間の 記憶および比較に必要であるというものである。この当 座的計時における線条体の役割は,強化が常に一定の間 隔後に呈示されるという課題の「定常性」を符号化する ことにあると考えることができる。つまり,線条体はあ る決まった間隔で自動的に反応を産出することに必要で あり,そこでは将来の強化時間に対する複雑で目的的な 予測や比較は含まれないと考えられる。これは線条体の より一般的な機能に即して考えると,刺激–反応連合に 基づいた習慣(habit)型オペラント行動に背側線条体が 重要であり(Yin, Knowlton, & Balleine, 2004),また反応 比率スケジュールよりも時隔スケジュールの方が線条 体依存の習慣を生成しやすく(Dickinson, Nicholas, & Adams, 1983),さらに反応間の選択を伴う並立時隔より も単一時隔スケジュールの方が習慣を形成しやすい (Kosaki & Dickinson, 2010)という知見と一致するもので
ある。 これに対し海馬は,過去に経験した事象間の時間間隔 を符号化し,それらを分離,比較することによって目的 的な反応選択を行うことに重要であると言える。この考 えは,先に述べた時間細胞の柔軟な表象特性からも支持 される。このような線条体と海馬の計時における仮説的 機能区分は,これらの部位のより一般的な機能区分と一 致する。例えば空間学習においては,線条体が特定の刺 激–反応連合によって目的地に り着く学習に必要であ る一方,海馬は複数の目印と目的地の関係性を符号化 し,より柔軟で正確な空間探索行動を可能にすることが 知られている(例えばKosaki, Lin, Horne, Pearce, & Gilroy, 2014; O’Keefe & Nadel, 1978; Packard & McGaugh, 1996)。 また課題によっては海馬と線条体の機能が学習において 競合的に働くことが知られており(Kosaki, Poulter, Aus-ten, & McGregor, 2015; Poldrack & Packard, 2003),時間学
習においても計時の範囲によっては海馬依存の計時と 線条体依存の計時が競合する可能性が示されている (Jacobs et al., 2013)点は興味深い。 このように見てくると,計時における線条体と海馬の 機能区分は,他の学習事態におけるこれらの領域の機能 区分とよく似ており,異なる学習・記憶システムの一部 として異なる種類の計時機能がそれぞれ備わっていると 考えることができる。もしこの考えが妥当なものであれ ば,他の学習事態においてこれまで研究されてきた問題 を計時学習にも適用することで,これらの複数計時機能 がどの程度独立であるのか,あるいは競合するのかと いった問題や,行動の目的性と計時の関係性といった, これまであまり検討されてこなかった問題に光が当たる ことが期待される。一つの例を挙げるならば,非空間性 のオペラント学習や空間学習においてはともに,訓練を 重ねるにつれて行動を制御する責任部位が海馬あるいは 前頭前野,および背内側線条体を中心とした回路から背 外側線条体を中心とした回路に遷移することが知られて いるが(近年の総説として例えば Balleine, Liljeholm, & Ostlund, 2009; Packard & Goodman, 2013),計時学習にお いてもこのような責任部位の経験依存的な変化が生じる かどうかを検証することが考えられる。このような問題 は,線条体と海馬における計時機能と他の学習機能との 関連性を調べるうえで,またこれらの部位が担う複数の 計時機構の間の機能的連携あるいは競合を明らかにする うえでも興味深い。こうした実験的研究を通じて,本稿 で紹介したSBFモデル(Matell & Meck, 2000, 2004)をも とに,計時における海馬の役割,さらには海馬と線条体 の相互作用をも説明するような包括的な計時神経モデル が発展することが期待される。また,上で述べたような 海馬が時間間隔をまとまりとして表象し,それらの比較 操作に関与するという可能性は,すなわち海馬が複数事 象間の時間的統合,例えば別稿で述べたMillerらの時間 的符号化仮説が提唱するような「時間地図」の生成に関 わることを予測する。現在のところ著者らが知る限りに おいてはこの予測は検証されておらず,今後の検証が待 たれるところである。 最後に,線条体と海馬の計時機能を結びつける基盤と して,前頭前野(prefrontal cortex; 以下PFCとする)の 役割が注目される。PFCは海馬から直接投射を受け(例 えばJay & Witter, 1991),またPFCと線条体との間には, SBFの主要構成要素である,視床を介したいくつかの並 列的回路が存在する(例えばAlexander, DeLong, & Strick, 1986)。したがって,当座的な計時と回顧的な計時を統 合するような場面,例えば上で述べた時間地図の生成の
ような機能においてPFCの関与が予想される。こうした 領域間での機能統合には,例えば海馬–PFC間の θ オシ レーションとVTA–PFC間の4 Hzオシレーションの同期 (Fujisawa & Buzsáki, 2011)などが関わっている可能性が 考えられる。また最近Meckらによって,領域間のオシ レーション同期によって計時情報と作業記憶を統合する 興奮–抑制オシレーションモデル(EIOモデル)が提出 されている(Gu, van Rijn, & Meck, 2015)。紙数の関係か ら本稿ではこれらを詳しく取り上げることはできない が,今後はこうした領域間の動的な相互作用を対象とし た研究が進展することが期待される。 結 論 本稿および別稿では,古典的条件づけおよびオペラン ト条件づけにおける時間の役割,時間情報の学習を広く 紹介した後,それらを担う神経機構を概観した。紙数の 制限から紹介できなかった現象や理論も多いが,それで も時間に関する研究が連合学習理論,行動分析,情報理 論,そして神経科学と多岐にわたる領域にまたがる大き な研究分野へと発展してきていることを見て取ることが できる。今後はこれらの領域間の融合した形での発展, さらに時間と空間に関する情報処理を統合するような大 きな理論的発展が期待されるところである。 謝 辞 本稿の準備・執筆にあたり,慶應義塾大学の渡辺茂名 誉教授に多くの貴重なご助言をいただきました。また本 稿の出版にあたり,日本学術振興会科学から責任著者へ の研究費補助金(課題番号: 26885079)による助成を受 けました。ここに謝意を表します。 引用文献
Alexander, G. E., DeLong, M. R., & Strick, P. L. (1986). Parallel organization of functionally segregated circuits linking bas-al ganglia and cortex. Annubas-al Review of Neuroscience, 9, 357–381.
Balci, F., Meck, W. H., Moore, H., & Brunner, D. (2009). Tim-ing deficits in agTim-ing and neuropathology. In J. L. Bizon & A. Woods (Eds.), Animal models of human cognitive aging. NJ: Humana Press, pp. 1–41.
Balleine, B. W., Liljeholm, M., & Ostlund, S. B. (2009). The in-tegrative function of the basal ganglia in instrumental con-ditioning. Behavioural Brain Research, 199, 43–52.
Bouton, M. E., & Garcia-Gutierez, A. (2006). Intertrial interval as a contextual stimulus. Behavioural Processes, 71, 307–317. Bouton, M. E., & Hendrix, M. C. (2011). Intertrial interval as a
contextual stimulus: Further analysis of a novel asymmetry in temporal discrimination learning. Journal of
Experimen-tal Psychology: Animal Behavior Processes, 37, 79–93. Bueno, J. L. O., & Bueno Júnior, L. S. (2011). Serial conditional
discrimination and temporal bisection in rats selectively lesioned in the dentate gyrus. Behavioural Processes, 86, 345–358.
Buhusi, C. V., & Meck, W. H. (2002). Differential effects of methamphetamine and haloperidol on the control of an internal clock. Behavioral Neuroscience, 116, 291–297. Buhusi, C., & Meck, W. H. (2005). What makes us tick?
Func-tional and neural mechanisms of interval timing. Nature Reviews Neuroscience, 6, 755–765.
Carr, J. A. R., & Wilkie, D. M. (1997). Ordinal, phase, and interval timing. In C. M. Bradshaw & E. Szabadi (Eds.), Time and behaviour: Psychological and neurobehavioural analyses. Amsterdam: Elsevier, pp. 265–327.
Catania, A. C. (1970). Reinforcement schedules and psycho-physical judgments: A study of some temporal properties of behavior. In W. N. Schoenfeld (Ed.), The theory of reinforce-ment schedules. New York: Appleton-Century-Crofts, pp. 1–42.
Church, R. M., & Broadbent, H. A. (1990). Alternative repre-sentations of time, number, and rate. Cognition, 37, 55–81. Church, R. M., & Deluty, M. Z. (1977). Bisection of temporal
intervals. Journal of Experimental Psychology: Animal Be-havior Processes, 3, 216–228.
Church, R. M., & Gibbon, J. (1982). Temporal generalization. Journal of Experimental Psychology: Animal Behavior Pro-cesses, 8, 165–186.
Dews, P. B. (1970). The theory of fixed-interval responding. In W. N. Schoenfeld (Ed.), The theory of reinforcement sched-ules. New York: Appleton-Century-Crofts, pp. 43–61. Dickinson, A., Nicholas, D. J., & Adams, C. D. (1983). The
effect of the instrumental training contingency on suscepti-bility to reinforcer devaluation. The Quarterly Journal of Experimental Psychology Section B, 35, 35–51.
Dietrich, A., & Allen, J. D. (1998). Functional dissociation of the prefrontal cortex and the hippocampus in timing behav-ior. Behavioral Neuroscience, 112, 1043–1047.
Dreyfus, L. R., Fetterman, J. G., Smith, L. D., & Stubbs, D. A. (1988). Discrimination of temporal relations by pigeons. Journal of Experimental Psychology: Animal Behavior Pro-cesses, 14, 349–367.
Eichenbaum, H. (2014). Time cells in the hippocampus: A new dimension for mapping memories. Nature Reviews Neuroscience, 15, 732–744.
Ferster, C. B., & Skinner, B. F. (1957). Schedules of reinforce-ment. New York: Appleton-Century-Crofts.
Fujisawa, S., & Buzsáki, G. (2011). A 4 Hz oscillation adaptive-ly synchronizes prefrontal, VTA, and hippocampal activi-ties. Neuron, 72, 153–165.
Gibbon, J. (1977). Scalar expectancy theory and Weber’s law in animal timing. Psychological Review, 84, 279–325. Gibbon, J. (1981). On the form and location of the
psychomet-ric bisection function for time. Journal of Mathematical Psy-chology, 24, 58–87.
Gibbon, J. (1991). Origins of scalar timing. Learning and
Moti-vation, 22, 3–38.
Gibbon, J., Church, R. M., & Meck, W. H. (1984). Scalar tim-ing in memory. Annals of the New York Academy of Sciences,
423, 52–77.
Gibbon, J., Fairhurst, S., & Goldberg, B. (1997). Cooperation, conflict and compromise between circadian and interval clocks in pigeons. In C. M. Bradshaw & E. Szabadi (Eds.), Time and behaviour: Psychological and neurobehavioural analyses. Amsterdam: Elsevier, pp. 329–384
Grossberg, S., & Schmajuk, N. A. (1989). Neural dynamics of adaptive timing and temporal discrimination during asso-ciative learning. Neural Networks, 2, 79–102.
Gu, B. M., van Rijn, H., & Meck, W. H. (2015). Oscillatory multiplexing of neural population codes for interval timing and working memory. Neuroscience & Biobehavioral Re-views, 48, 160–185.
Hattori, M., & Sakata, S. (2014). Brain electrophysiological activity correlates with temporal processing in rats. Behav-ioural Processes, 101, 97–102.
石井 拓(2008a).動物のインターバル計時の諸理論と 論点 I ―静的特徴を中心として― 心理学評論,51, 526–546.
(Ishii, T. (2008a). Theories and moot points on animals’ interval timing I: Static properties. Japanese Psychological Review, 51, 526–546.)
石井 拓(2008b).動物のインターバル計時の諸理論と 論点II ―動的特徴を中心として― 心理学評論,51, 547–566.
(Ishii, T. (2008b). Theories and moot points on animals’ interval timing II: Dynamic properties. Japanese Psychologi-cal Review, 51, 547–566.)
Jacobs, N. S., Allen, T. A., Nguyen, N., & Fortin, N. J. (2013). Critical role of the hippocampus in memory for elapsed time. The Journal of Neuroscience, 33, 13888–13893. Jarrard, L. E., & Davidson, T. L. (1990). Acquisition of
concur-rent conditional discriminations in rats with ibotenate lesions of hippocampus and of subiculum. Psychobiology,
18, 68–73.
Jay, T. M., & Witter, M. P. (1991). Distribution of hippocampal CA1 and subicular efferents in the prefrontal cortex of the rat studied by means of anterograde transport of Phaseolus vulgaris-leucoagglutinin. The Journal of Comparative Neu-rology, 313, 574–586.
Johnson, A., & Redish, A. D. (2007). Neural ensembles in CA3 transiently encode paths forward of the animal at a decision point. The Journal of Neuroscience, 27, 12176–12189. Killeen, P. R., & Fetterman, J. G. (1988). A behavioral theory of
timing. Psychological Review, 95, 274–295.
Kosaki, Y., & Dickinson, A. (2010). Choice and contingency in the development of behavioral autonomy during instru-mental conditioning. Journal of Experiinstru-mental Psychology: Animal Behavior Processes, 36, 334–342.
Kosaki, Y., Lin, T. E., Horne, M. R., Pearce, J. M., & Gilroy, K. E. (2014). The role of the hippocampus in passive and active spatial learning. Hippocampus, 24, 1633–1652.
Dorsolateral striatal lesions impair navigation based on landmark-goal vectors but facilitate spatial learning based on a “cognitive map”. Learning & Memory, 22, 179–191. 神前 裕・時 暁聴・松井 大・新保彰大・藤巻 峻
(2015).条件づけにおける時間I―古典的条件づけ― 基礎心理学研究,34, 60–77.
(Kosaki, Y., Shi, X., Matsui, H., Shimbo, A., & Fujimaki, S. (2015). Roles of temporal information in conditioning I: Classical conditioning. The Japanese Journal of Psychonomic Science, 34, 60–77)
Kyd, R. J., Pearce, J. M., Haselgrove, M., Amin, E., & Aggleton, J. P. (2008). The effects of hippocampal system lesions on a novel temporal discrimination task for rats. Behavioural Brain Research, 187, 159–171.
Le Merrer, J., Rezai, X., Scherrer, G., Becker, J. A. J., & Kieffer, B. L. (2013). Impaired hippocampus-dependent and facili-tated striatum-dependent behaviors in mice lacking the delta opioid receptor. Neuropsychopharmacology, 38, 1050– 1059.
MacDonald, C. J. (2014). Prospective and retrospective dura-tion memory in the hippocampus: Is time in the foreground or background? Philosophical Transactions of the Royal Soci-ety of London, Series B, Biological Sciences, 369, 20120463. MacDonald, C. J., Carrow, S., Place, R., & Eichenbaum, H.
(2013). Distinct hippocampal time cell sequences represent odor memories in immobilized Rats. The Journal of Neuro-science, 33, 14607–14616.
MacDonald, C. J., Cheng, R. K., & Meck, W. H. (2012). Acqui-sition of “Start” and “Stop” response thresholds in peak-interval timing is differentially sensitive to protein synthesis inhibition in the dorsal and ventral striatum. Frontiers in Integrative Neuroscience, 6, 1–16.
MacDonald, C. J., Fortin, N. J. Sakata, S, & Meck, W. H. (2014). Retrospective and prospective views on the role of the hippocampus in interval timing and memory for elapsed time. Timing & Time Perception, 2, 51–61.
MacDonald, C. J., Lepage, K. Q., Eden, U. T., & Eichenbaum, H. (2011). Hippocampal “time cells” bridge the gap in memory for discontiguous events. Neuron, 71, 737–749. Machado, A. (1997). Learning the temporal dynamics of
behavior. Psychological Review, 104, 241–265.
Maricq, A. V., & Church, R. M. (1983). The differential effects of haloperidol and methamphetamine on time estimation in the rat. Psychopharmacology, 79, 10–15.
Maricq, A. V., Roberts, S., & Church, R. M. (1981). Metham-phetamine and time estimation. Journal of Experimental Psychology: Animal Behavior Processes, 7, 18–30.
Matell, M. S., Bateson, M., & Meck, W. H. (2006). Single-trials analyses demonstrate that increases in clock speed contrib-ute to the methamphetamine-induced horizontal shifts in peak-interval timing functions. Psychopharmacology, 188, 201–212.
Matell, M. S., King, G. R., & Meck, W. H. (2004). Differential modulation of clock speed by the administration of inter-mittent versus continuous cocaine. Behavioral Neuroscience,
118, 150–156.
Matell, M. S., & Meck, W. H. (2000). Neuropsychological mechanisms of interval timing behavior. Bioessays, 22, 94– 103.
Matell, M. S., & Meck, W. H. (2004). Cortico-striatal circuits and interval timing: Coincidence detection of oscillatory processes. Cognitive Brain Research, 21, 139–170.
Matell, M. S., Meck, W. H., & Nicolelis, M. A. L. (2003). Inter-val timing and the encoding of signal duration by ensem-bles of cortical and striatal neurons. Behavioral Neurosci-ence, 117, 760–773.
McEchron, M. D., Bouwmeester, H., Tseng, W., Weiss, C., & Disterhoft, J. F. (1998). Hippocampectomy disrupts auditory trace fear conditioning and contextual fear conditioning in the rat. Hippocampus, 8, 638–646.
Meck, W. H. (1983). Selective adjustment of the speed of inter-nal clock and memory processes. Jourinter-nal of Experimental Psychology: Animal Behavior Processes, 9, 171–201.
Meck, W. H. (1986). Affinity for the dopamine D2 receptor predicts neuroleptic potency in decreasing the speed of an internal clock. Pharmacology, Biochemistry, and Behavior,
25, 1185–1189.
Meck, W. H. (1988). Hippocampal function is required for feedback control of an internal clock’s criterion. Behavioral Neuroscience, 102, 54–60.
Meck, W. H. (1996). Neuropharmacology of timing and time perception. Cognitive Brain Research, 3, 227–242.
Meck, W. H. (2006a). Frontal cortex lesions eliminate the clock speed effect of dopaminergic drugs on interval tim-ing. Brain Research, 1108, 157–167.
Meck, W. H. (2006b). Neuroanatomical localization of an in-ternal clock: A functional link between mesolimbic, nigros-triatal, and mesocortical dopaminergic systems. Brain Re-search, 1109, 93–107.
Meck, W. H., Church, R. M., & Matell, M. S. (2013). Hippo-campus, time, and memory ― A retrospective analysis. Behavioral Neuroscience, 127, 642–654.
Meck, W. H., Church, R. M., & Olton, D. S. (1984). Hippocam-pus, time, and memory. Behavioral Neuroscience, 98, 3–22. Meck, W. H., Church, R. M., Wenk, G. L., & Olton, D. S.
(1987). Nucleus basalis magnocellularis and medial septal area lesions differentially impair temporal memory. The Journal of Neuroscience, 7, 3505–3511.
Melgire, M., Ragot, R., Samson, S., Penney, T. B., Meck, W. H., & Pouthas, V. (2005). Auditory/visual duration bisection in patients with left or right medial-temporal lobe resection. Brain and Cognition, 58, 119–124.
Miall, C. (1989). The storage of time intervals using oscillating neurons. Neural Computation, 1, 359–371.
Moyer, J. R., Deyo, R. A, & Disterhoft, J. F. (1990). Hippocam-pectomy disrupts trace eye-blink conditioning in rabbits. Behavioral Neuroscience, 104, 243–252.
O’Keefe, J., & Nadel, L. (1978). The hippocampus as a cognitive map. Oxford: Oxford University Press.
O’Keefe, J., & Recce, M. L. (1993). Phase relationship between hippocampal place units and the EEG theta rhythm. Hippocampus, 3, 317–330.
Olton, D. S., Meck, W. H., & Church, R. M. (1987). Separation of hippocampal and amygdaloid involvement in temporal memory dysfunctions. Brain Research, 404, 180–188. Olton, D. S., Wenk, G. L., Church, R. M., & Meck, W. H.
(1988). Attention and the frontal cortex as examined by simultaneous temporal processing. Neuropsychologia, 26, 307–318.
Onoda, K., & Sakata, S. (2006). An ERP study of temporal dis-crimination in rats. Behavioural Processes, 71, 235–240. Onoda, K., Takahashi, E., & Sakata, S. (2003). Event-related
potentials in the frontal cortex, hippocampus, and cerebel-lum during a temporal discrimination task in rats. Cognitive Brain Research, 17, 380–387.
Packard, M. G., & Goodman, J. (2013). Factors that influence the relative use of multiple memory systems. Hippocampus,
23, 1044–1052.
Packard, M. G., & McGaugh, J. L. (1996). Inactivation of hip-pocampus or caudate nucleus with lidocaine differentially affects expression of place and response learning. Neurobiol-ogy of Learning and Memory, 65, 65–72.
Platt, J. R., & Davis, E. R. (1983). Bisection of temporal inter-vals by pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 9, 160–170.
Poldrack, R. A., & Packard, M. G. (2003). Competition among multiple memory systems: Converging evidence from ani-mal and human brain studies. Neuropsychologia, 41, 245– 251.
Quinn, J. J., Oommen, S. S., Morrison, G. E., & Fanselow, M. S. (2002). Post-training excitotoxic lesions of the dorsal hippo-campus attenuate forward trace, backward trace, and delay fear conditioning in a temporally specific manner. Hippocampus, 12, 495–504.
Roberts, S. (1981). Isolation of an internal clock. Journal of Experimental Psychology: Animal Behavior Processes, 7, 242– 268.
Sakata, S., & Onoda, K. (2003). Electrophysiological correlates of interval timing. In Meck, W. H. (Ed.), Functional and neural mechanisms of interval timing. FL: CRC Press, pp. 339–349.
Schneider, B. A. (1969). A two-state analysis of fixed-interval responding in pigeons. Journal of the Experimental Analysis of Behavior, 12, 667–687.
Schultz, W., Dayan, P., & Montague, P. R. (1997). A neural substrate of prediction and reward. Science, 275, 1593–1599. Shaw, C., & Aggleton, J. P. (1994). The ability of amnesic
subjects to estimate time intervals. Neuropsychologia, 32, 857–873.
Solomon, P. R., Vander Schaaf, E. R., Thompson, R. F., & Weisz, D. J. (1986). Hippocampus and trace conditioning of
the rabbit’s classically conditioned nictitating membrane response. Behavioral Neuroscience, 100, 729–744.
Staddon, J. E. R. & Higa, J. J. (1999). Time and memory: Towards a pacemaker-free theory of interval timing. Journal of the Experimental Analysis of Behavior, 71, 215–251. Stubbs, D. A. (1968). The discrimination of stimulus duration
by pigeons. Journal of the Experimental Analysis of Behavior,
11, 223–238.
Stubbs, D. A., Dreyfus, L. R., Fetterman, J. G., Boynton, D. M., Locklin, N., & Smith, L. D. (1994). Duration comparison: Relative stimulus differences, stimulus age, and stimulus predictiveness. Journal of the Experimental Analysis Behav-ior, 62, 15–32.
Tam, S. K. E., & Bonardi, C. (2012). Dorsal hippocampal involvement in appetitive trace conditioning and interval timing. Behavioral Neuroscience, 126, 258–69.
Tam, S. K. E., Jennings, D. J., & Bonardi, C. (2013). Dorsal hippocampal involvement in conditioned-response timing and maintenance of temporal information in the absence of the CS. Experimental Brain Research, 227, 547–559. Todd, T. P., Winterbauer, N. E., & Bouton, M. E. (2010).
Inter-stimulus interval as a discriminative Inter-stimulus: Evidence of the generality of a novel asymmetry in temporal discrimina-tion learning. Behavioural Processes, 84, 412–420.
Treves, A., & Rolls, E. T. (1994). Computational analysis of the role of the hippocampus in memory. Hippocampus, 4, 374– 391.
Ward, R. D., Kellendonk, C., Simpson, E. H., Lipatova, O., Drew, M. R., Fairhurst, S., Kandel, E. R., & Balsam, P. D. (2009). Impaired timing precision produced by striatal D2 receptor overexpression is mediated by cognitive and moti-vational deficits. Behavioral Neuroscience, 123, 720–730. Wearden, J. H. (1999). “Beyond the fields we know...”:
Explor-ing and developExplor-ing scalar timExplor-ing theory. Behavioural Pro-cesses, 45, 3–21.
Yamaguchi, Y. (2003). A theory of hippocampal memory based on theta phase precession. Biological cybernetics, 89, 1–9.
Yin, B., & Meck, W. H. (2014). Comparison of interval timing behavior in mice following dorsal or ventral hippocampal lesions with mice having δ-opioid receptor gene deletion. Philosophical Transactions of the Royal Society of London, Series B, Biological Sciences, 369, 20120466.
Yin, H. H., Knowlton, B. J., & Balleine, B. W. (2004). Lesions of dorsolateral striatum preserve outcome expectancy but dis-rupt habit formation in instrumental learning. European Journal of Neuroscience, 19, 181–189.