The
JaPanese
Jottntat
of Psychonomic SCience2003
,
VoL22
,
No
.
1,
132−
136講 演論 文
領 域
固
有 性
・一
般 性
と
脳機 能
川 畑 秀 明
鹿 児 島大 学
Domain
−
specificity
and
generality
in
the
brain
Hideaki
KAwABATA
Kαgoshima University*
Domain −
specificity and generality of cognition are exciting subjects relating that can shedlight Qn thc processes of human cognitive evo !ution
.
Their theorization,
however,
is
very compli・
cated
,
and the re]ationshipsbetween
domain
specificity arldbasic
architecture ofthe
brain
have
not yet been fully clarified
、
This paper reviews recent studies on the relationshipbetween
domain
−
specificity/generality andbrain
functions.
Among visual cognition,
in which the authoris
specialized
,
facial
cognition necds special attention asit
is
connectedto
the
basic
social cognitionand should have had an important adaptive value
.
The
studies with neuroimaging show that faceperception
,
cognition of facial expression,
and that of facial attractiveness are processedin
severalparticular brain areas
including
fusiform face area,
amygdala and some else.
Moreover,
recent studies of neuroimaging suggest that thereis
a neuralbasis
for
not on】ydomain
specificity but alsofor
the
general intelligence as dornain−gencrality
Qf cognition.
Key words :
domain
specificity,
domain
gcncrality,
face perception,
social cognition,
brain
L
は じめに ヒ ト は 生命 活動 ヒ, 生存や繁殖の有 利さ にっ な が る よ うに,
様々 な適 応 課 題に応 じて特 化し た 認知機構を持っ て い る.
そ れで は,
ヒ ト はどの程 度,
領 域 固 有の心的・
神経シ ス テム を 進 化 させ てきたの だ ろうか (Elman et al.
1996),
ま た,
ヒ トは どの よ うに認 知 的 流動性を もっ た一
般 化し た心を持っ よ うに進 化 して き たの だ ろ う か (Mithen,
1996).
こ の ようなt 心の スイス アー
ミー
ナ イ フ的な特化さ れ た脳機能と, 認 知の様々 な領 域 固 有 性を っ なぐ一
般 性に関す る脳 機能の両側面の理解は発 達心 理 学や進 化 心理学の視 点を 応用し な が ら発 展すべ く,
今 後 の 認知神経 科 学の重 要な課 題 とな る,
で は,
生存や繁 殖 上で有 利にな る よ うに構 成された認 知 機 能とは何で あ ろ う か,
Pinker (1994
)はヒ トの 心の 進 化 的な適 応 性 を 考 慮したか た ちで い くっかの領 域 固 有 な 心 を 示 し た.
た と え ば,
知 覚や言語,
食べ 物 (食べて*
Department
ofPsychology,
Faculty
ofEduca−
tion,
Kagoshima University,1−20−6
Kurimoto,
Kagoshima
−
shi 890−
0065 い いものは何か),
臼 己概 念,
正 義 感,
配 偶 (性 的 魅 力な ど),
危 険へ の認知な ど が その適 応 的 心の 例 として挙 げ られている.
確 かに,
これ らの領 域 固 有な認 知機能は,
ヒ トが生 存や繁 殖 上で 有 利に働 くべ く構 成され た認 知機 能であ る と と もに,
人 間 ら し さの 源 と な るよ うな認 知 機 能を含んで い る といえ る.
現 在,
特 定の 情 報が脳の ど こ で処理 されるか とい う 「エ リア X 」を脳 内に見っ けよ う とする とい う作 業が,
神経生 理学や脳イ メー
ジ ング 法の発 展と相まっ て世 界 的 に盛ん である.
本 稿で は,
これ まで明ら かになっ てい る 認知の領 域 固 有 性・一
般 性と脳 機 能の関 係につ い て,
特 に顔認 知にお け る領 域 固 有 性の問 題 を 中心に扱 う.
2.
視 覚 認 知の領 域 固 有 性と脳機
能 : 顔 と場 所の認 知 知覚研究 特に視覚の レ ベ ル で は,
サルを 用いた神 経 生理学に よっ
て,
色や奥行き,
運動な どの視覚情報が脳 の ど の部 位で専 門 的に処理 されて い るのか が明ら かに なっ てきてい る,
こ の よう な詳 細 な機 能 単位の処 理は,
知 覚が高度に専 門 化 (特殊化 )さ れ たメカニ ズムによっThe Japanese Psychonomic Society
NII-Electronic Library Service The Japanese Psyohonomio Sooiety
川 畑:領 域 固 有 性
・一
般 性と脳 機 能133
OA0.
20 一.
14L4C
響
¶.
5 t 0。
5 4β 0 卩1● 駒 働鵬 ● 卩隔ceFigure 1
.
Result ofbrain
areas activated byface and place stimuli
in
a single subject.
Sta−
tistical analysis was
done
with SPM99 (Well・
colne
DepartInent
of Imaging Neuroscience,
London ,
U .
K .
〕.
〔a)Brain activity at fusiform
face
arca (FFA )produced by contrast of’
face
vs
.
place’
(Tarailach coordinates,
38,−
50,
−
22).
(b)Brain activity at para−hippocampal
place area (PPA )
by ‘
p!ace vs.
face’
(Tarailach
coordinates
,
20,−
60,−
4).
Thesc images were superimposed on SPM standard template.
(c),
〔
d
):Parameter
estimatcs for face stimuli andplace stimuli at the peak point in FFA (c)and
PPA {d)
.
て構成さ れて い ることを示 して いる(
Zeki,1978,1993
>.
た とえ ば, V4 が色 知覚の セ ン ター
で あ ること (た と えば
,Zeki,1973
;Bartels
&Zeki,
2000a),
運 動 情 報は,
網 膜か ら外側 膝状体 (LGN )を経 由し て V1
,
V2,
V5 へ と 到 達 する とい う.一
連の情 報 処理の流れ が存 在し (Zeki,
1971},
V1
が局 所 的な運 動 方 向検出を する の に比べて,
V5
がグロー
バ ル な 運 動 知 覚 や 速度 知 覚に選 択 的 応 答 す るこ と が知ら れて いる (StQner & Albrlght,1993
; ffyche et a ].
,
1995).
さ らに最 近,
物体や顔,
場 所 (風 景) な どの より高 次な 視 覚 情報に も専門化さ れ た脳 機 能が存 在して い るこ と が わ かっ て き た.
顔や場 所の知 覚は,
ヒ ト が適 応 的に行動 する上で必 要となる領 域 固有な認 知で あ る といえ よ う.
Figure 1は顔 知 覚,
場 所 知 覚の 脳機能の局在性を表し た 機能的 磁 気 共 鳴 画 像 診 断 法 (fMRI )に よる計 測 事 例であ る.
た と え ば顔の知 覚で は,
紡 錘 状 [り1
(fusiform gyrus> で局 所 的な活 動が見ら れ (Figure la},
こ の顔 刺激に対 して特 異 的に反 応す る領域は
FFA
(fusifor
皿face
area )と呼ば れて い る (Kanwisher et al
.
,
1997).
また風 景の よう な場 所の知覚で は
,
傍 海 馬 回 (parahippocampal gy−
rus )が局 所 的に活 動 し (
Figure
lb
),
こ の 領域はPPA
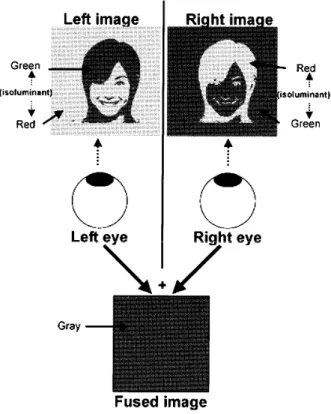
Green
争
「1501umm 画nl ウ Red 1∈i//、
宅
/
/
t
争
●
Left
eye
Grey/
/
/
/
tttua\
!
/
争
A
‘戛
ノ
Right
eye Red 争 etuinlnant ) Ψ GreenFused
image
Figure 2
.
A schernatic presentation of the
dichoptic
stimulatiQn method usedin
Moutoussis &
Zeki
〔2002).
Fused image of
dichoptic
presentation is invisible becausematched areas have opposite color cQntrast
between
the two eyes.
Single image present−
ed to the eye
,
includes objectimage
whichhas isoluminant color
between
object andbackground
.
(para
−hippocampal
place area )と呼ば れて い る (Ep−
stein & Kanwisher,
1998),
F{gurelc
と1d
は顔刺激,
風 景 刺 激に対す る FFA
,
PPA の 活 動 変化量 を示して い る.
こ の よ うに, 顔の知 覚一FFA ,
場 所の知 覚一
PPA,
とい う刺激一
脳 内 部 位の対 応 関 係が存 在 することが わ か る,
また FFA の活動は, 同じ人 種, 異な る人 種の顔の記 憶 再 認 時に も起こ るが,
同じ人 種の顔の方が異な る人 種 の 顔 再 認 時より もFFA の活 動が高 くなるとい う,
記 憶 による FFA 活 動のモ ジュ レー
シ ョ ンを 示 す 知 見も あ る (Golby et al.
2001 ).
こ の 認知の違い の根 底には自分の 仲間を 見分け る とい う社 会 的 認 知 機 能の適 応が,
脳 内 処 理に影響を及ぼ して い る と考え る こ と が で き る.
顔の部 分 的な要 素におい て も, 特に対人 関係に必 要と さ れる視 線や 口の動 きのよ う な情報の場合は,
その知 覚 認知に特定の脳機 能が存 在する.
視 線 知 覚や 口 の動 きの 知 覚,
口元を読む作 業の 場 合で は,
側 頭 葉にある上 側 頭溝 (superior temporal sulcus ;
STS
)が活 動を示す (Puce
134
基 礎 心 理 学 研 究 第22 巻 第1号 et al.
,
1998:Hoffman &Haxby ,2000
).
ま た, 視線知 覚 はSTS
の活 動を引 き起こすだけ でな く,
顔 自体の知覚 中 枢で あ るFFA
の 活 動をモ ジュ レー
トするこ とも わ か っ て い る (George et al.
,
2001 ).
視 線 知 覚やllの動 き に よ っ て活 動が起こ る STS は,
そ の 他に も光 点で体の 動 き を 表 すバ イ オ ロ ジカ ル モー
シ ョ ン 〔Senior
et aL,
2000 ),
手の部 分 的な動き (Rizzolatti et al.
,
1996>に も 特 異 的に応 答する.
こ の よ うな刺 激に対 する局 在 的脳 活 動は情 報が入 力さ れて い ること が重要な の で あ り,
必 ずしも意 識される必 要が ない.
た と えばFigure
2
に示される ように,
背 景と そ のL
に ある図 形が反対色で等 輝度に なっ て い る刺 激 を,
背 景と図 形の色の対 応 を 逆に して 2っ 左 右 両 眼 別に 同時提 示す る と,
そ れぞれの 眼で は刺 激が何であるか が 同 定で きるが,
両眼で融合して み る と2
っ の刺 激の 背 景 と図 形がそれ ぞ れ反 対 色であ るこ と か ら刺激は一
様な色 に知 覚さ れ,
背 景の上の図 形が知 覚で き なくなる.
この よ う な刺 激呈示で 背景の上に顔や建物のパ タ ンを配 置し て提 示 し,
その時の脳 活動をfMRI
で調べ る と,
見え て い な い顔 刺 激に対して もFFA が活 動し,
ま た見えて い な い建物 刺激に対し て もPPA
が 活 動 するこ とが 明 らか になっ て い る (Moutoussis &Zeki,
2002 ).
っ ま り,
脳に イ ン プ ッ トさ え あ れ ば見え ない顔や場 所に対して脳は活 動 するこ と を 意 味 し,
認 知 以 前に脳は自動 的に機 能して いっ で も認 知で き るよ うに準 備し て い る と言え る.
3.
社 会 的情 報と して の顔認知と 脳機 能: 表 情 視 線 知 覚や表情認知を含め,
顔の認 知は,
ヒ ト の ヒ ト ら しさ た る高 度な社 会 性の入 り口 と しての 機能を 果 た す と同 時に,
生 得 的な機 能で あ る と知ら れて い る (Slater
&Kirby,1998
).
さらに視 線や身 体 部 分の知 覚はコ ミュ ニ ケー
ショ ン の ため に必要と さ れ る機能で ある.
それゆ え に,
社 会 的 情 報と して の顔 情 報は他の視 覚刺激と区 別 して認識されるべ く領 域1
司有 件を持っ
た認 知と して発達 し, 専 門化し た脳機能が 必要と さ れた と考え る こと がで きる.
顔が持 っ て い る形態特 徴以外の情 報と して表情を挙げ ること が で き る.
表 情 認 知の脳 機 能で は,
大 き く3っ の 領 域の 役 割が 重 要視さ れて いる,
〔1)扁 桃 体 (amygdala ) は基 本 感 情の ほぼ すべての表 情 認知に関与して い るとさ れる 〔Adolphs,
2002〕.
特に恐 怖 表 情の よ うに脅威や危 険信 号と しての感情理解につ い て は,
こ の領 域の活 動が ±曽加する 〔Morris
et al.
1996
).
(2
)前頭 前野(prefrontal area )は怒 りな どの表 情 認 知において活動を示し(Kesler一
West
et aL,
2001
),
rTMS (連 続 経 頭 蓋 磁 気 刺 激 )を前 頭 前野 部に当て た研 究で は,
怒り表 情 認 知が反 応 時 間レ ベ ル で他の表 情の認 知に比べ て遅 く な る とい う結果が得ら れて い る (Harmer et aL,2001
>.
さ らに, 眼窩 前頭 野(orbito
−
frontal area )で は喜びの表 情 認 知 時に活 動するとの結 果も あ り(
Kesler・
West
et al.
2001
),
また こ の領域の障 害は ポジ テ ィ ブ感情の 認知
・
生 成に影 響 を 及ぼし
,
そ の結 果,
気 分 障 害と結びつ く との知 見 も あ る(Da −
masio et aL
,1994
).
(3
)体 性 感 覚 皮 質 (sQmato−
sensory cortex )は嫌悪表情刺激に対して活 動 を 示 す (Calder et al.
,
2000 )ほ か,
あ ら ゆ る感 情 認 知の表象の場と して考 え られて いる (Da皿 asio,
1994).
4. 社
会 的情報
から得 ら れ る 印 象と脳 機 能 : 魅力Pinker
〔1994
)の リス ト にある ように,
進 化 適 応に必 要と さ れ る 配偶 決 定に関して い う と顔 情 報に は魅 力の要 因も含ま れ,
ま た 乳 児 は 発 達の初 期か ら魅 力の あ る顔に 対 する選 好を示 すこ と か ら (Rubenstein
et aL,1999
), 魅 力に対す る感 情 形 成は領 域 固有な認 知機能で ある と考 え ら れ る.
顔に魅 力を感じる際に,
活動す る脳 部 位と して は眼 窩 前頭 野な ど が挙げられ る 〔Aharon et al.
,
2001 ;ODG −
herty
et aL,
2002
).
こ の領 域は,
他の サブ領 域 (島や前 頭前野,
前部 帯状回,
扁 桃 体な ど) を含め,
喜 びな どの ポ ジ ティ ブ感 情の生 起 や(Blood &Zatorre,
200
1
) , 報酬 を 得る際の脳 活 動 と一
致 する(Small et al.
,
2001 ).
この こと か ら顔魅 力 は 報 酬 価 と して認知 者に与え ら れ る情 報 であるとの考え も あ る.
また,
結 婚 柑 手や恋 人とい っ た恋 愛 感 情を向けてい る 人の写真刺激に対 する脳 活 動 測 定で も,
島や前 部 帯 状 回 な どの活動が見受け られ る(Bartels
&Zeki,
2000b ).
さ らに,
視 覚 刺 激か らの性 的覚 醒時に も,
島 や 前 部帯 状回,
眼窩 前 頭 野,
前 頭 前 野の活 動が見ら れる と さ れ る (Kar−
ama et al.
,
2002 ).
今 後は,
眼 窩 前頭 野,
前部 帯状 回, 後 部 帯 状 回,
島,
扁 桃 体な どの領 域が,
報 酬「系 (reward system )と社会 的感 情 系 (social elnotion system )の双 方で どのよ う な振
舞いを す るの かにっ いて時間的明ら か に して い く必 要が あ る
.
5
.
領 域一
般 性と脳 機 能 このよ うに領 域 固 有 性に関す る 研究は比 較 的 あり (領 域 固 有 性を直 接 扱っ
て い る研 究 自体は少ない), そ の定 義やモ ジュー
ル との区 別は曖 昧な が らも強 調さ れて いてThe Japanese Psychonomic Society
NII-Electronic Library Service The Japanese Psyohonomio Sooiety
川 畑:領 域 固有性
・一
般 性と脳機能135
い る.一
方,
領域一
般性にっ い て は研 究が非 常に少な く,
その 定 義さ え非 常に曖 昧で ある.
Mithen (1996)は,
ヒ トは進 化の中で技 術 的,
言 語 的,
社 会 的,
博 物 的 知 能の 4つ の独 立 した知 能 を 獲 得 し,
そ れ が 現 代 人 の よ う な 高 度な知能を有す る よ うに なっ たの はそれ らの 4っ の知 能をっ なぐ認 知 的 流 動 性が発 達したか らで ある と した.
こ こ で は,
領 域一
般性を Mithen の い う認知 的 流 動性の よ う にメタモ ジュー
ル と して あ ら ゆ る領 域と も独立 し た 領 域と して定 義 して み る.
そ の領 域一
般 性の例として,
こ こ で は知 能の問 題 を 取 りF
.
げる..
般 能 力 として の 記 号 操 作 能 力の よ う なSpeaman
の一
般 囚 子と して表さ れて い る知能を必 要と する課 題で は,
前 頭 葉 外 側 部が活 動を示 し た (Duncan eta1.
,
2000 ),
ま た,
問 題 解 決 能 力で メ タ認知の よ うな認知制 御と しての
一
般 的流動性知能(generalnuid
intelli
・
gence)を必要と する課題で も前頭 葉外側
.
部や前部 帯 状 回の活 動が見ら れる こ と が明ら かになっ て い る 〔Gray etaL,
2003 ).
さ らに,
行 動コ ン トロー
ルやエ ラー
モニ タ リン グの よ う な 認 知 制 御の際の 脳 活 動 も前 部 帯 状 回 を 中 心 と してい る(Swick
&Turken,
2002 ).
6.
結 び と して 本 稿で は顔知覚を中心 に領域 固有性と脳 機能の話題を 中心 に展 開して き た.
社会 的認知や感情の脳 機 能は,
視 覚 情 報 処 理の よ うに単 純に 1っ の エ リア X の機 能 局 在 に よっ て成り立っ て い るとい うよ り,
複 数の領 域が関 与 して いる場合が.
多い,
表情認知で は,
表 情が何で あ る か にか か わ らず 扁 桃 体が活 勁 すること が多いが,
こ の よう に い くっ かの認 知 機 能で共 通して活 動 する領 域を 明 ら か にする こ と, 逆に眼窩前頭 野が魅 力と報.
酬の双方に関 与 して いる な ど,
特 定の領域が ど ん な 認知機 能に使わ れて い る のか を 理解 する ことは,
認 知 領 域 間の関 係を明ら か に して くれる.
引 用 文 献Adolphs,
R .2002
Recognizing emotionfro
皿fa
¢ial
expressions ;Psychological and neurological mech
−
anisms
.
Behavioralαnd Cognitivθ Neurosciencθ Re−
views
,
1,
21−
61,
Aharon ,1
.
, Etcoff,
N.
,
Ariely,
D.
,
Chabris,
C.
E,
0℃ on−
nor
,
E.
,
&Breiter,
H,
C.2001
Beautiful
faces
have
variable reward value :
fMRI
andbehavioral
evi−
dence.
Neuron,32,537−551.
Bartels
,
A.
& Zeki,
S.
S.
2000a The architecture of the colour centre in the human visual brain:newresults and a review
,
Eπm ρθαη/傭隅 α♂qプNeurt
)∫oゴー
ence
,
12,
172−
193.
Bartcls
,
A.
& Zeki,
S.
S.
2000b Neural basis of ro・
mantic
love.
Neuroreport
,
11,
3829−
3834.
Calder
,
A.
工,
Keane,
J.
,
Manes,
R,
Antoun,
N.
,
&Young
,
A.
W.
20001mpaired recognition and ex−
perience Qf
disgust
followingbrain
in
ゴury.
Nature
ハ偐πγosc歪6ηo¢
,3,1077−1078.
Damasio
,
A.
19941 )escarte’
s Error:Emotion,
1〜ation一
α傭yand the Human Brain
.
New York :Putnarn(
Grosset
Books
).
Damasio
,
H.
,
Grabowski,
T.
,
Frank,
R.
,
Galaburda,
A,
M
.
,& Damasio , A.
R.
1994 The return of PhineasGage :clues about the brain from the skull of a
famous patient
.
Science
,264,
1102−nO5 .
Duncan ,
1
,
,
Seitz
,
R .
」,
,
Kolodny,
」.
,
Bor,
D .
,
Hezog ,
H ,
,
Ahmed
,
A,
,
Newell,
F,
N.
,
& Emslie,
H,
2000 A neu−
ral basis of generalintelligence
.
Science,
289,
457−
460,
Elman ,
J
.
L.
,
Bates,
E.
A .
Johnson,
M.
H.
,
Kar 皿iloff
・
Smith
,
A.
,
Parisi,
D、
,
& Plunkett,
K.
1996 Rethinle−
ing
Innatene∬ ;ノ4 con ηectionistP
¢rsp¢ctive on devθt−
opment
.
MIT Press:Massachusetts.
Epstein,
R .
&Kanwisher,
N ,
1998A
cQrtlca !repre−
sentatjon of the
local
visual environment.
ムlt犲ure,
392
,
598−
601,
ffytche
,
D .
H .
,
Guy ,
C ,
N .
,
&Zeki,
S,1995
The
para工lel
visua !motion
inputs
into
areas Vl and V50f hu−
man cerebral cQrtex
.
Brain,
l l8,1375− 1394.
George , N
.
,
Driver,J.
,&Dolan, R.
J,
2001Seen
gaze−
direction modulates fusiform activity and its cou
−
pling with other
brain
areasduring
face
process・
ing.
Neur
()image,13,1102−1112.
Golby
,
A.
J.
,
Gabrieli,
D,
E.
,
Chiao,
」,
Y.
,
& Eberhardt,
J.
L
.
2001Differential responses in the fusiform re−
gion
to
same−
race and other−
racefaces,
ハlatureハ[eur ()science
,
4,
845−
850,
Gray,
J.
R,
Chabris,
C ,
F.
,
& Braver,
T,
S.
2003 Neu−
ral mechanisnls of general 且uid
intelligence
.
ハ[a−
ture Neuroscience
,
6,
316−322,
Harmer
,
C,
工,
Thilo,
K,
V 。
,
Rothwell,
J
,
C .
,
&Goodwin ,
G .
M ,2001
Transcranial
magnetic sti皿 ulation ofmedial
−frontal
corteximpairs
the processing of an−
gryfacial
expressions.
Na tu re neuroscience.
4,
17−
18.
Hoffman
,
E.
A.
& Haxby,
J,
V.
2000 Distinct repre−
sentations of eye gaze and
identity
in
thedistrib
−
uted
human
neural systemfor
face
perceptiQn,
〈[ature Neuroscienee
,
3,
80−
84,
Kanwisher
,
N.
,
McDermott,
J.
,
&Chun ,
M.
M,
1997The fし1siform face area :amodu !e in human ex
−
trastriate
cortex specializedfor
face
perception,
ノ
bumal
q厂ハleuroscience,17,
4302−4311.
Karama ,
S.
,
Lecours,
A .
R ,
Leroux,
J.
M ,
Bourgouin,
P
.
,
BeaudQin,
G.
,&Joubert
, S,
M.
2002 Areas Qfbrain activation in males and females during view
−
ing
of eroticfilm
excerpts.
ffuman
Brain
MapPing ,
16
,
1−
13.
136
ges,diing\tfflnrg
22gij
1g-Kesler-West.M L.Andersen, A.H,,Smith,C.D.
Avi-son,
M.
J.
Davis,
C,
E,,
Kryscio,
R.
J,,
&
Blonder,
L,
X.
2001
Neural
substrates offacial
emotionproc-essing using
fMRI.
Cognitive
Brain
Researeh,
11, 213-226.Mithen,
S.
1996 The Ptehistoryof
Mind, London: Thames and Hudson.Morris,
J,
S. Frith,C.D. Perrett,D.I.Rowland, D. Young, A.W.Calder,
A.J.&
Dolan, R.J. 1996 A differentialneural response in thehuman
amyg-dala
tofearful
andhappy
facial
expressions, IVa-ture,383,
812-81s.
MQutoussis,
K,&
Zeki,S,
2002 The relationship be-tween cortical activation and perception investi-gated with invisiblestimuli. P)'oceedingsof
the IVdtionatAcademyof
Science,
USA,
99,
9527-9532.
O'Doherty,
J.,
Winston,J.
Critchley,
H,,Perrett,D.Burt,
D.
M.
&
Dolan,
R,
J.
2003
Beauty
in
asmile:the role of medial orbitofrontal cortex
in
facial
attractiveness, NeurQPsycholQgia,41,!47-155. Pinker,S, 1994 71heLanguageinstinct.London:Pen-guin,Puce,
A.
Allison,
T.
Bentin,
S.,
Gore,
J.
C.
&
McCar-thy,
G,
1998
Tempora]
cortex activationin
hu-mans viewing eye and mouth movements.f
IVeu-roscience,18,
2188-2199.
Rizzolatti,G. Fadiga, L,,Matelli,M,,Bettinardi,V. Paulesu, E. Parani,D. & Fazio,F. 1996
Localiza-tion
of grasp representationsin
humans
by
PET:
1.
Observation versus execution,
ExPerimental
BratnResearch,
111,
246-252,
Rodman,
H.R.
&
Albright,
T.D.
1989
Single-unit
analysis of pattem-motion selective propertiesin the middle temporal visual area
(MT).
ExPen'men-talBrain Researth,75,53-64.
Rubenstein, A.J.,Kalakanis, L.
&
Langlois,J.H.
1999
Infant
preferencesfor
attractivefaces:
a cog-nitive explanation.DevelQPmentat
Rsychology,
35,
848-855,
Senior,C. Barnes,
J.
Giampietro, V. Simmons, A,, Bu]lmore, E.T.,Brammer, M.,& David, A.S,2000 The functional neuroanatomy of implicit-motion perception or `representationalmomentum.'
CurL
rent
BiotQgy,
10,
16-22,
Slater,
A.,
&
Kirby,
R.
1998
Innate
andlearned
per-ceptual abilitiesin
thenewborn infant.ExPerimen-talBrain Research, 123,90-94.
SmalL D.M.,Zatorre,R.
J.,
Dagher, A.,Evans, A.C.,&Jones-Gotman,
M. 2001 Changes inbrainactivity relatedto
eating chocolate: from pleasure to aver-sion.Brain,
124,
1720-1733.
Stoner,
G,
R,
&
A]bright,
T.
D,
1993
Image
segmen-tationcues
in
motion processing: implicationsformodularity
in
vision.fou?"nat
of
Cognitive
IVleurosci-ence,5,
129-149.Swick, D. & Turken, A.U. 2002 Dissociation be-tween conflict
detectien
and error monitoringin
the
human
anterior cingulate cortex.P?'oceedings
of
theNdtional
Academy
of
Science,
USA,
99,
16354-16359.
Zeki,
S.
1978 Functional specialisationin
the visual cortex of the rhe$us monkey, IVdture,274, 423-428.Zeki,S,
1993
A
Vision
of
theBrain,
Oxford:
Black-wellScientific
Publications.
Zeki,
S,
1971Convergent
input
from
thestriatetex