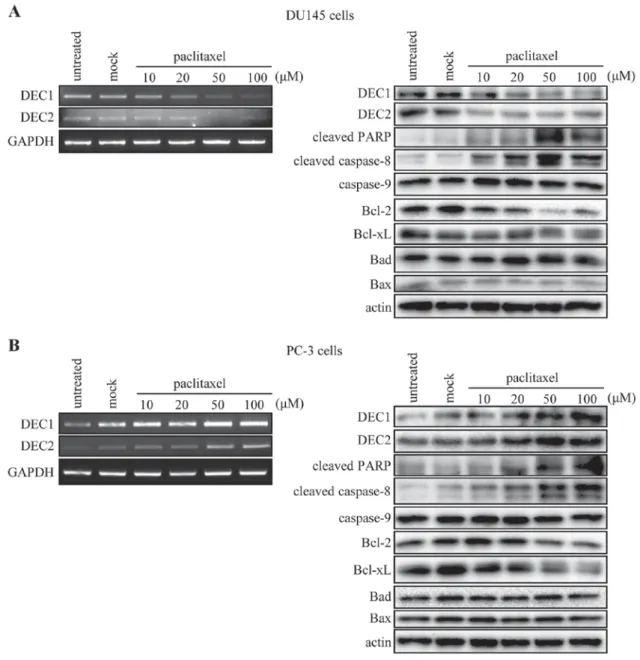
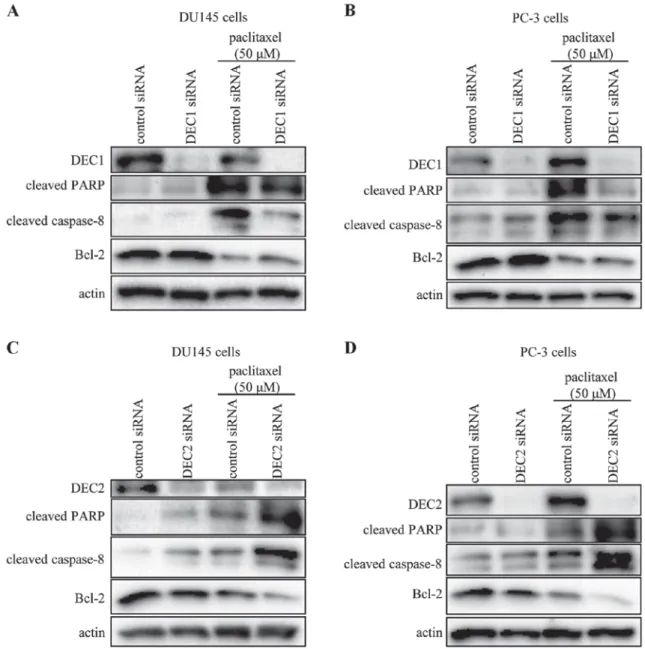
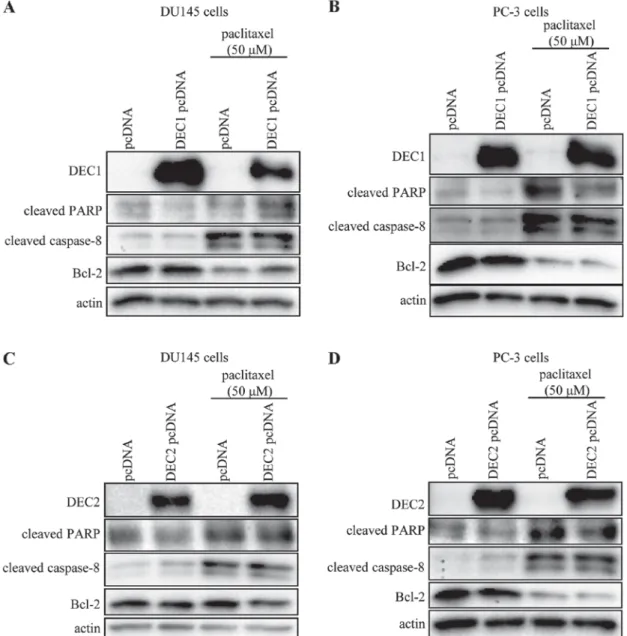
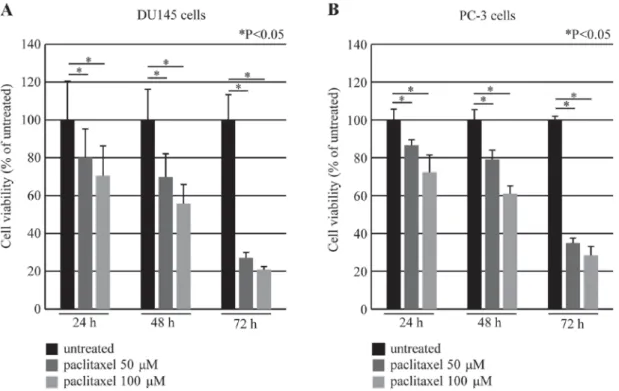
DEC2 exerted an anti-apoptotic effect in paclitaxel-induced apoptosis of human prostate cancer cells.
7
0
0
全文
(2)
(3)
(4)
(5)
(6)
(7)
図
関連したドキュメント
Effect of gemcitabine on the expression of apoptosis-related genes in human pancreatic cancer cells.
reported that gemcitabine-mediated apoptosis is caspase- dependent in pancreatic cancers; Jones et al [14] showed that gemcitabine-induced apoptosis is achieved through the
induced CXCL8 mRNA expression (data not shown) and CXCL8 protein expression in pancreatic cancer cells in xenografted tumor tissues (Fig.. We next administered anti-CXCL8 antibody
Accordingly, our results showed that kahweol acetate and cafestol inhibited the proliferation and migration of both ACHN and Caki-1 cells, with their synergistic effects apparent
(14) Similarly, as observed on other members of Pim family, (15,16) we demonstrated that aberrantly expressed Pim-3 can inactivate a pro-apoptotic BH3-only Bcl-2-like molecule,
To determine the promoter activities of 5 ′ -flanking region of the human Pim-3 gene, we subcloned various deleted 5 ′ -flanking regions of the human Pim-3 gene into a
Pim-3, a proto-oncogene with serine ⁄ threonine kinase activity, is aberrantly expressed in human pancreatic cancer and phosphorylates Bad to block Bad-mediated apoptosis in
In this study, we attempt to show the cell fate of proliferating Müller cells after MNU treatment to address the question “Are proliferating Müller cells indeed the source
The impact of rotational error regarding the treatment of prostate cancer with VMAT has not yet been evaluated, and our preliminary study showed that pitch angle error affected
