J. Brew. Soc. Japan. Vol.107, No. 12, p. 931~939(2012) 研 究 報 文
セミヨン種ワイン発酵醪から分離した乳酸菌の醸造学的性質
岸本宗和
1・塩原貫司
1・萩原健一
2・今井裕景
2・柳田藤寿
1(1山梨大学ワイン科学研究センター,2株式会社サドヤ)
平成 24 年 4 月 11 日受理
Characterization of Lactic Acid Bacteria Isolated from Semillon Grape Must
Munekazu Kishimoto1, Kanji shiohara1, Kenichi hagiwara2, Yukyo imai2, and Fujitoshi Yanagida1
(1The Institute of Enology and Viticulture, University of Yamanashi, 1-13-1 Kitashin, Kofu, Yamanashi 400-0005, Japan,
2Sadoya Co., Ltd, 3-3-24 Kitaguchi, Kofu, Yamanashi 400-0024, Japan)
Fifty-six lactic acid bacterial strains were isolated from Semillon grape must and winery equipment of Sadoya Co., Ltd., and malic acid degradation ability was confirmed in 43 of these strains. To identify these 43 strains, the polymerase chain reaction-restriction fragment length polymorphism method and 16S rDNA sequencing were carried out. All the strains isolated from the early stage of fermentation and the late stage of fermentation were identified as Lactobacillus plantarum and Oenococcus oeni, respectively. A strain isolated from winery equipment was identified as Pediococcus pentosaceus. The malic acid degrada-tion abilities of the two strains selected as the representatives of the former two species were compared to that of the commercially available starter O. oeni PN4 (Lallemand) in pH, ethanol, sulfur dioxide, and temperature resistance tests. The 09Se-A1-4 that was isolated from the early stage of fermentation showed a higher degradation rate than the others at pH 2.9. This strain also rapidly degraded malic acid in 4 ℓ scale winemaking trials. These findings suggest that 09Se-A1-4 is a useful strain for malolactic fer-mentation in grape musts at a low pH.
Key words: 白ワイン,マロラクティック発酵,Lactobacillus plantarum ─────────────────
緒言 乳酸菌が関与するマロラクティック発酵(MLF)は, 酵母によるアルコール発酵が完了した後に生起し,リ ンゴ酸を乳酸と二酸化炭素に変換することによってワ インの酸味を低減するとともに,微生物学的な安定性 を付与する。また,乳酸菌の生成する微量成分が製成 ワインの香味品質に大きな影響を及ぼし,ワインの香 りの複雑性を高める効果がある1,2,3)。これらのことか ら,MLF は多くの赤ワインや一部の白ワイン,ある いは,伝統的な発泡性ワインにおいて,極めて重要な 工程の一つとされている4)。 MLF に関与する乳酸菌は,球菌の Oenococcus oeni や Leuconostoc mesenteroides, 桿 菌 の Lactobacillus plantarum,Lb. hilgardii,Lb. brevis など多くがある が,主要となるのはこの内の O. oeni と Lb. plantar-um であるとされている5,6,7,8)。乳酸菌は生育に複雑な 栄養要求性を持つほか,生育環境の変化に対しても影 響を受けやすい。MLF の生起に及ぼす種々の要因の 中でも pH,亜硫酸濃度,アルコール濃度,温度の 4 つが最も大きな影響を与えるとされている4,9)。自然 発生により行われる MLF では,ワインの低 pH,高 アルコール濃度,高亜硫酸濃度あるいは低温などの厳 しいワイン環境の中で乳酸菌の生存や増殖が抑制され
醸 協( ) やすく,MLF の生起を制御することは極めて困難で あった。近年,優良な乳酸菌が選抜されて MLF スタ ーターとして利用されるようになり10),簡便に MLF を生起できるようになりつつある。しかしながら,接 種するワインによっては,pH,アルコール濃度,亜 硫酸濃度,発酵温度あるいは栄養状態などの環境に適 応できない場合には,接種したスターターが死滅して MLF が生起しないこともある。不安定な MLF の生 起を高めるために,海外では不足する栄養分を含む促 進剤の利用なども進められているが,白ワインの醸造 は pH が低い場合が多く,低温で発酵が行われるため に赤ワインに比べて MLF の生起が困難である。 他方,MLF において乳酸菌が生成する微量の香味 成分にジアセチル,アセトイン,酢酸,乳酸エチルな どがある。これらの成分はいずれも含有量が増加する とワインの品質低下を招く恐れがあるが,適度に存在 する場合には香味に複雑性を付与する。例えば,ジア セチルは 1 ~ 4ppm の濃度範囲では好ましい影響を与 えるものの,5ppm を超えるような場合には品質に悪 影響を及ぼすとされており1,11),ワインの種類によっ ても品質に影響を及ぼす濃度は異なる12)。 このようなことから,MLF を安定的に生起し,酒 質の向上や多様化に寄与するためには,ワインの pH, アルコール濃度,亜硫酸濃度,発酵温度あるいは栄養 状態などの環境要因,さらには,ブドウ品種,醸造方 法,求めるワインの酒質など様々な状況に応じて選択 的に使用できる特徴の異なった乳酸菌の入手が不可欠 である。本研究では,多様な乳酸菌の収集を目的とし て,白ワイン醸造において頻繁に MLF が生起するワ イナリーのワイン発酵醪および醸造設備より乳酸菌を 分離し,その醸造学的性質について調査したので結果 を報告する。 実験方法 1.ワイン発酵醪および醸造設備からの乳酸菌の分 離 株式会社サドヤにおいて,甲府市善光寺で収穫され た 2009 年セミヨン種ブドウ果汁(初発糖度 23 度, pH 3.3,総酸 5.62g/ℓ,遊離亜硫酸濃度 14 mg/ℓ,総 亜硫酸濃度 48 mg/ℓ)を原料として仕込みを行った ワイン発酵醪から一部を発酵の経過に合わせて採取し た。なお,醪の澱引きと亜硫酸の添加は実施しなかっ た。また,醸造設備からの分離は,滅菌したガーゼを 用いた拭き取りにより行った。採取した試料を生理食 塩水にて適宜希釈した後に,BM 平板培地6)に塗布し て 30℃で 3 ~ 5 日間アネロパック・ケンキ(三菱ガ ス化学)を使用して嫌気的に培養を行った。出現した 性状の異なるコロニーから,カタラーゼ陰性かつグラ ム陽性を示す菌株を乳酸菌として単離した。 2.16S rDNA の解析
供試菌株のゲノム DNA 抽出には Nucleospin Tis-sue(Macherey-Nagel)を使用した。抽出した DNA を 鋳 型 と し, プ ラ イ マ ー に 27F(5'-AGAGTTT-GATCCTGGCTCAG-3)および 1492R(5'-GGTTAC-CTTGTTACGACTT-3)を用いて 16S rDNA を増幅 した。PCR 反応は,変性 95℃で 30 秒,アニーリング 60℃で 30 秒,伸長 72℃で 90 秒を 30 サイクルの条件 で行い,電気泳動にて増幅産物を確認後,一部を制限 酵素 Hae Ⅲで処理しアガロースゲル電気泳動解析を 行った。 また,上述の方法で抽出した DNA を鋳型として, プライマーに 27F,1492R および WLAB1(5'-TCCG-GATTTATTGGGCGTAAAGCGA-3),WLAB2(5' -TCGAATTAAACCACATGCTCCA-3)13)を用いて同 条 件 で PCR を 行 っ た。 得 ら れ た 増 幅 産 物 を QIAquick PCR Purification Kit(Qiagen 社製)を用 いて精製し,株式会社ファスマック DNA 合成事業部 にて塩基配列解析を行った後,データベース(BLAST program of the National Center for Biotechnology In-formation14))にて相同性検索を行った。 3.分離株のリンゴ酸分解能の測定 山梨大学ワイン科学研究センター付属育種試験場で 収穫したセミヨン種ブドウを亜硫酸無添加で発酵した ワイン(リンゴ酸 1.98 g/ℓ)を培地に用い,アルコー ル 9.2 %,pH 3.50,総亜硫酸 8 mg/ℓ,発酵温度 25℃ を基本条件とした。試験に応じて酒石酸を用いて pH を 2.9 あるいは 3.1 に,アルコール濃度を 12.0 %ある いは 14.5 %に,総亜硫酸濃度を 20 mg/ℓあるいは 30 mg/ℓに調整し,発酵温度を 15℃あるいは 20℃とした。 これに,BM 培地で前培養した菌株の洗浄菌体懸濁液 (109 cfu/mℓ)を試験培地に対して 1 %量になるよう に接種し,25℃で 4 週間培養後に残存するリンゴ酸量 を HPLC 有機酸分析システムにより測定した。
4.小規模試験醸造 2010 年に山梨大学ワイン科学研究センター付属育 種試験地で収穫したセミヨン種ブドウを原料とし,亜 硫酸を 25 mg/ℓの濃度で添加して一夜静置した清澄 果汁(糖度 21.6 度,pH 3.24,総酸 6.79 g/ℓ,リンゴ 酸 1.41 g/ℓ)を原料に,4ℓ規模で行った15)。なお,
酵母は乾燥酵母 MaurivinTM Cru-Blanc(AB Mauri)
を用い,資化性窒素が 150 mg・N/ℓになるようにリ ン酸水素二アンモニウムを加えて,発酵温度 18℃で 発酵を行った。アルコールの生成が緩慢になった発酵 17日目に,BM 培地で前培養した乳酸菌の洗浄菌体 を 106~ 107 cfu/mℓに な る よ う に 接 種 し て 22 ℃ で MLF を行った。 5.分析方法 試験ワインの一般成分分析は,国税庁所定分析法注 解16)に従って実施した。有機酸の分析は,島津製作 所 HPLC 有機酸分析システムにより行った。カラム は Rspack KC-811(8.0mm i.d. × 300mm,2 本 直 列 接続)を用い,5mM p- トルエンスルホン酸水溶液を 溶出液として,カラム温度 40℃,流速 0.8ml/min で 溶出し,5mM p- トルエンスルホン酸および 100μM EDTA を含む 20mM ビス(2- ヒドロキシエチル)イ ミノトリス(ヒドロキシメチル)メタン水溶液を用い たポストカラム pH 緩衝化電気伝導度検出法により分 析した。 香気成分の分析は,ガスクロマトグラフィーにより 行った。酢酸エチル,n- プロピルアルコール,イソ ブチルアルコールおよびイソアミルアルコールの低沸 点成分は GC-2014(島津製作所)を用いて Shinohara and Watanabe の直接導入法17)により行った。その他 の成分は,ペンタン:ジエチルエーテル= 1:1 で香気 成分を抽出した後,GCMS-QP2010Ultra(島津製作 所)にて分析した。分析カラムには Inert Cap Pure Wax(60m × 0.25mm i.d., 膜厚 0.25μm,GL サイエン ス)を使用した。注入口温度 230℃,イオン源温度 200℃,キャリアガス He 流量 1.0 mℓ/min,カラム温 度 80℃で 10 分間保持後 3℃ /min で 140℃まで昇温, さらに 5℃ /min で 220℃まで昇温して 10 分間保持の 条件で分析した。 また,試験ワインは,12 名の研究員により 20 点法 で官能評価した。 結果および考察 1.セミヨン種ワイン製品における MLF 生起状況 株式会社サドヤの 1970 年から 2008 年に醸造された セミヨン種ワイン製品 13 点について,一般成分およ び有機酸の分析を行い MLF 生起の状況を調査した。 結果を Table 1 に示した。セミヨン種ワインのアルコ ール濃度は 11.3 ~ 13.5 %,pH は 3.14 ~ 3.64,総酸は 4.55 ~ 7.64 g/ℓの範囲にあった。また,リンゴ酸は 未検出あるいは 0.20 ~ 1.62 g/ℓ,乳酸は 0.42 ~ 3.55 g/ℓの範囲にあった。一般的に乳酸菌が関与しないワ
Vintage (v/v %) pHAlcohol Titratable acidity(g/ℓ) Citric acid Tartaric acid Malic acid Lactic acid Succinic acid Acetic acid(g/ℓ)
1970 13.5 3.63 5.09 0.23 1.11 1.30 1.42 0.43 0.60 1972 12.5 3.64 5.09 0.17 1.03 0.20 3.11 0.45 0.95 1975 12.7 3.61 5.62 0.06 0.47 NDa) 3.55 0.57 1.21 1977 12.2 3.41 5.66 0.19 1.03 ND 2.75 0.54 0.98 1979 12.0 3.41 5.87 0.25 0.98 ND 2.83 0.38 0.73 1985 11.3 3.41 7.64 0.23 1.06 0.58 3.16 0.52 1.62 1986 12.4 3.47 6.79 0.13 0.59 ND 2.76 0.42 0.81 1989 11.4 3.28 6.40 0.27 1.89 1.62 1.16 0.29 0.39 1990 12.4 3.35 4.94 0.04 1.61 ND 1.51 0.47 0.54 1992 12.5 3.50 5.01 0.25 1.26 0.76 0.60 0.40 0.27 1997 12.6 3.57 4.55 0.23 1.47 0.84 1.29 0.28 0.52 2004 12.5 3.14 6.95 0.31 3.76 0.65 0.42 0.20 0.68 2008 12.7 3.53 5.68 0.37 1.15 1.40 0.62 0.78 0.35 a)Not detected.
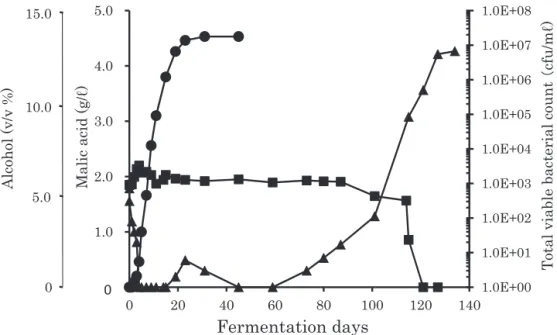
醸 協( ) インの乳酸含有量は 0.5 g/ℓ程度であり18),1992 年, 2004年,2008 年の製品においては MLF が生起しな かったと考えられる。一方,乳酸が 1.0 g/ℓ以上含ま れる 10 点のワインでは MLF が生起したものと推察 され,株式会社サドヤのセミヨン種ワインでは,高頻 度で MLF が自然生起することが示された。 2.セミヨン種ワイン発酵醪および醸造設備からの 乳酸菌の分離 前述において,株式会社サドヤのセミヨン種ワイン では頻繁に MLF が自然生起することが確認された。 そこで,2009 年にセミヨン種ワイン発酵醪および醸 造設備から乳酸菌の分離を試みた。ワイン発酵醪のア ルコール濃度,リンゴ酸含有量,バクテリア生菌数の 推 移 を Fig. 1 に 示 し た。 発 酵 開 始 時 に は 約 1 × 103cfu/mℓの菌数が認められたが,アルコールの生成 とともに急激に減少した。その後,発酵 60 日を過ぎ たころから再び菌数の増加が認められ,MLF が自然 生起してリンゴ酸が減少した。なお,2009 年の仕込 みでは,MLF が自然生起し,完了するまでに 134 日 を要したが,ワイン発酵醪のアルコールが 13.6 %と 比較的高いことが長期化した要因の一つと考えられる。 このセミヨン種ワイン発酵醪から乳酸菌の分離を試 みた結果,発酵の開始からアルコールの生成が緩慢に なりはじめた発酵 23 日目までの期間(前期醪)に 22 菌株が分離され,大部分が発酵 3 日目までの期間であ った。一方,発酵 60 日目以降からリンゴ酸が完全に 消費された発酵 134 日目までの期間(後期醪)に 24 菌株が分離された。さらに,醸造設備から 10 菌株が 分離された。これらの分離株の内,前期醪から分離さ れた 18 菌株,後期醪から分離された 24 菌株および醸 造設備から分離された 1 菌株の合計 43 菌株にリンゴ 酸の分解能が認められた。 3.分離株の分類学上の位置
O. oeni, Leu. mesenteroides, Lb. plantarum および 関連の乳酸菌の識別に制限酵素 Hae Ⅲにより処理し た 16S rDNA の PCR-RFLP 解析が有効である19)。分 離された菌株の中のリンゴ酸分解能を有する 43 菌株 について制限酵素 Hae Ⅲにより処理した 16S rDNA の電気泳動解析を行った結果を Fig. 2 に示した。前期 醪から分離された 09Se-A1-4 株,09Se-A3-1 株のバン ドパターンは Lb. plantarum(NRIC 1067T)のバンド 1.0E+00 1.0E+01 1.0E+02 1.0E+03 1.0E+04 1.0E+05 1.0E+06 1.0E+07 1.0E+08 0.0 1.0 2.0 3.0 4.0 5.0 0 20 40 60 80 100 120 140
Fermentation days
Malic acid (g/ℓ)
Total
viable
bacterial
count
(c
fu
/mℓ)
Alcohol
(v/v
%)
5.0 15.0 10.0 00
Fig. 1 Changes in malic acid content and viable bacterial count during fermentation of Semil-lon grape must.
Symbols: (●),alcohol content in must; (■), malic acid content in must; (▲), total viable bacterial count.
パターンと一致した。なお,前期醪から分離されたリ ンゴ酸分解能を有するその他の菌株もすべて Lb. plantarum(NRIC 1067T)と同一のバンドパターンを 示した。一方,後期醪から分離された 09Se-B3-2 株, 09Se-B9-1 株は O. oeni(JCM 6125T)のバンドパター ンと一致し,後期醪から分離されたリンゴ酸分解能を 有するその他の菌株もすべて O. oeni(JCM6125T)と 同じバンドパターンを示した。また,ワイナリー醸造 設 備 か ら 分 離 さ れ た 09SAD-5-2 株 は O. oeni, Lb. plantarum 双方のバンドパターンと一致せず,異なる 種であると判断される。 前期醪から分離された 09Se-A1-4 株,09Se-A3-1 株, 後期醪から分離された 09Se-B3-2 株,09Se-B9-1 株お よびワイナリー醸造設備から分離された 09SAD-5-2 株について 16S rDNA の塩基配列に基づく解析を行 った。NCBI BLAST を利用して,アセンブルした塩 基配列について既知配列との相同性を検索した結果, 前期醪から分離された 09Se-A1-4 株,09Se-A3-1 株は GenBank accession number HM218736 あ る い は HQ259243 の Lb. plantarum の 16S rDNA 配列と 99% 以上の相同性を示し,後期醪から分離された菌株は GenBank accession number HM013951 の O. oeni と の相同性が 99 %以上を示した。また,ワイナリー設 備より分離された 09SAD-5-2 株は GenBank accession number HQ286591 の Pediococcus pentosaceus との相 同性が 99 %以上を示し,ワイン発酵醪から分離され た菌株とは異なる種であった。以上の PCR-RFLP 解 析および相同性の結果を踏まえ,前期醪から分離され た菌株は Lb. plantarum に,後期醪から分離された菌 株は O. oeni に,ワイナリー醸造設備から分離された 菌株は P. pentosaceus に分類される可能性が極めて高 いことが明らかになった。Lafon-Lafourcade ら7), Fleet ら20)はボルドー地方のワインについて調べ,発
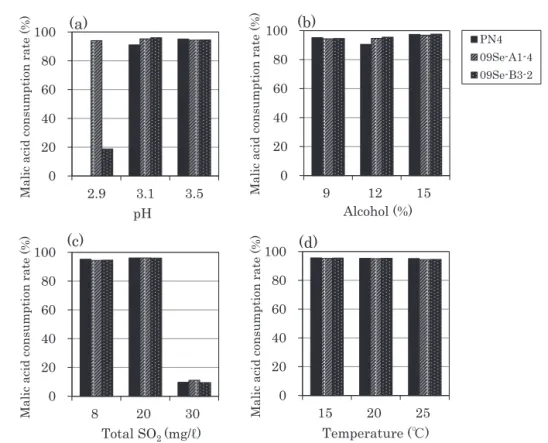
酵 初 期 に は Lactobacillus, Leuconostoc, Pediococcus 属の乳酸菌が多く分離され,アルコール発酵後には O. oeni が分離されたことを報告した。柳田ら21)は国 内の赤ワインについて調査し,発酵初期には Lb. plantarum が,後期には O. oeni が多く分離されたこ とを報告しており,本研究の結果はこれらの報告を支 持するものであった。 4. 分離株の MLF 生起に及ぼす pH,アルコール 濃度,亜硫酸濃度および発酵温度の影響 セミヨン種ワイン培地を用いて分離株の MLF の生 起に及ぼす pH,発酵温度,アルコール濃度,亜硫酸 濃度の影響を検討した。試験には,前期醪および後期 醪から分離されたそれぞれの菌株の中から予備試験に おいて最もリンゴ酸分解能が高かった 09Se-A1-4 株お よび 09Se-B3-2 株を用い,市販 MLF スタータ O. oeni PN4(Lallemand)と比較した。結果を Fig. 3 に示し た。PN4 株は pH 3.1 および pH 3.5 において 90 %以 上のリンゴ酸分解率を示したが,pH 2.9 では MLF が
600
1000
300
Size(bp)
100
1
M
2 3 4 5 6 7 M
Fig.2 Hae Ⅲ restriction patterns of PCR-amplified 16S rDNA. Lanes: M, size marker; 1, O. oeni (JCM 6125T); 2, Lb. plantarum(NRIC
醸 協( ) 生起せず,リンゴ酸の分解は認められなかった。これ に対して,前期醪から分離された 09Se-A1-4 株は試験 したいずれの pH においても MLF が生起し,リンゴ 酸の分解率は 90 %以上であった。一方,後期醪から 分離された 09Se-B3-2 株は pH 3.1 および pH 3.5 にお いて 90%以上のリンゴ酸分解率を示したが,pH 2.9 ではリンゴ酸の分解率は 20 %程度に止まった。ワイ ンの pH は MLF の生起に大きな影響を与え,pH3.3 以上では MLF の生起が容易であるが,pH 3.3 以下で は困難になることが報告されている22)。Bousbouras and Kunkee は23),ワインの pH は MLF に要する期 間に大きな影響を及ぼし,pH 3.15 のワインに乳酸菌 を接種した場合に MLF の完了に 164 日を要したのに 対し,pH 3.83 では 14 日で完了したことを報告した。 また,pH 3.5 以下のワインで自然生起する MLF の多 くが O. oeni によることが明らかにされている24)。こ のように,MLF の生起や優占種の決定にとってワイ ンの pH は重要な要因の一つである。本研究で前期醪 から分離された 09Se-A1-4 株(Lb. plantarum)が, MLF の生起が困難な pH2.9 において高いリンゴ酸分 解率を示した結果は非常に興味深く,低 pH 耐性を有 する有用な菌株であると考えられる。なお,発酵温度, アルコール濃度および亜硫酸濃度も MLF の生起に大 きな影響を及ぼす要因とされるが,リンゴ酸分解率に 及ぼすこれらの影響に大きな差異は認められなかった (Fig. 3)。 5.小規模試験醸造 2010 年にセミヨン種ブドウを原料として,4 ℓ規模 の小規模試験醸造を実施した。前期醪から分離された 低 pH 耐性を有する 09Se-A1-4 株および市販の MLF スターター O. oeni PN4 を供試し,乳酸菌を接種しな い試験区を比較対象とした。なお,乳酸菌は発酵 17 日目に接種した。発酵期間中のアルコール濃度および リンゴ酸含有量の推移を Fig. 4 に示した。09Se-A1-4 株は接種後に速やかな MLF の生起がみられ,約 2 週 間で大部分のリンゴ酸が消費された。一方,PN4 株 は 09Se-A1-4 株と同時期に接種したものの,MLF の
(a)
(b)
(d)
(c)
0 20 40 60 80 100 2.9 3.1 3.5 Malic acid consumption rate (%) pH 0 20 40 60 80 100 9 12 15 PN4 09Se-A1-4 09Se-B3-2 Malic acid consumption rate (%) Alcohol (%) 0 20 40 60 80 100 15 20 25 Malic acid consumption rate (%) Temperature (℃) 0 20 40 60 80 100 8 20 30 Malic acid consumption rate (%) Total SO2 (mg/ℓ)Fig. 3 Malic acid degradation ability in pH(a), alcohol(b), sulfur dioxide(c) and tem-perature(d) resistance test.
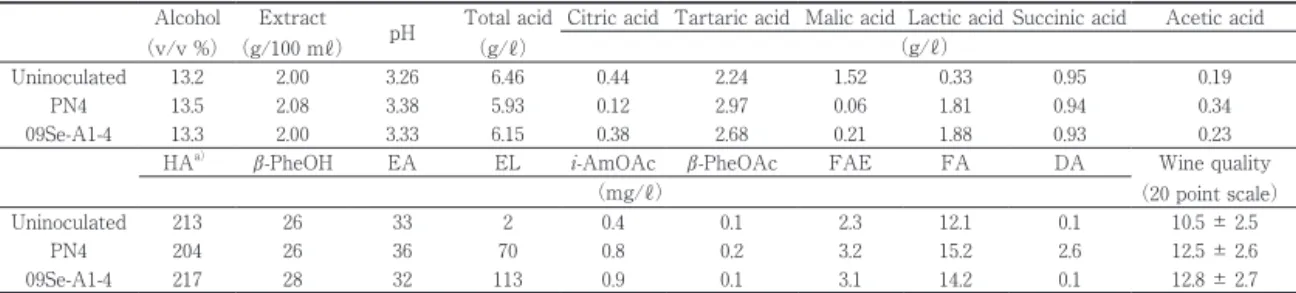
生起が遅れ,発酵 66 日目を過ぎたころからリンゴ酸 の消費が始まり,発酵開始から MLF が完了するまで に 86 日を要した。なお,乳酸菌を接種しない比較対 象区ではリンゴ酸の消費が認められず,MLF は生起 しなかった。乳酸菌の接種時期は,酒母の添加と同時 期,アルコール発酵の盛んな時期,アルコール発酵終 了直後がある25)。アルコール発酵終了直後は,乳酸 菌がワイン環境に適応して MLF が生起しやすく,最 も一般的な接種時期である。しかし,PN4 株では, 接種時のワインが比較的低い pH3.31 であったことや 酵母による栄養源消費に起因する栄養源不足などの要 因が相まって菌の増殖を抑制し,結果として MLF の 生起が大幅に遅れたものと推測される。 試験ワインの分析結果を Table 2 に示した。一般成 分分析における総酸値および pH,香気成分分析にお ける乳酸エチル,有機酸分析におけるリンゴ酸および 乳酸などの分析値から,比較対象区では MLF が生起 せず,09Se-A1-4 株および PN4 株では MLF が生起し
0.0
0.5
1.0
1.5
2.0
0.0
5.0
10.0
15.0
0
20
40
60
80
Malic acid (g/ℓ)
Alcohol(%)
Fermentation days
Addtion of lactic acid bacteria
Fig. 4 Evolution of malolactic fermentation in Semillon wine inoculated with an isolated strain and commer-cially available strain at 4 ℓ scale winemaking trials.
Symbols: (●), isolated strain 09Se-A1-4 ; (■), commercially available strain PN4; (▲), uninoculated (control). Straight line represents the concentration of alcohol, and dotted lines represent the concentration of malic acid.
Alcohol (v/v %) Extract (g/100 mℓ) pH Total acid (g/ℓ)
Citric acid Tartaric acid Malic acid Lactic acid Succinic acid Acetic acid (g/ℓ)
Uninoculated 13.2 2.00 3.26 6.46 0.44 2.24 1.52 0.33 0.95 0.19
PN4 13.5 2.08 3.38 5.93 0.12 2.97 0.06 1.81 0.94 0.34
09Se-A1-4 13.3 2.00 3.33 6.15 0.38 2.68 0.21 1.88 0.93 0.23
HAa) β-PheOH EA EL i-AmOAc β-PheOAc FAE FA DA Wine quality
(20 point scale) (mg/ℓ)
Uninoculated 213 26 33 2 0.4 0.1 2.3 12.1 0.1 10.5 ± 2.5
PN4 204 26 36 70 0.8 0.2 3.2 15.2 2.6 12.5 ± 2.6
09Se-A1-4 217 28 32 113 0.9 0.1 3.1 14.2 0.1 12.8 ± 2.7
a) HA, sum of n-propyl, i-butyl and i-amyl alcohols; β-PheOH, β-phenylethyl alcohol; EA, ethyl acetate; EL, ethyl lactate; i-AmO-Ac, isoamyl acetate;
β-PheOAc, β-phenylethyl acetate; FAE, sum of ethyl caproate, ethyl caprylate and ethyl caprate; FA, sum of caproic, caprylic and capric acid; DA, diacetyl.
醸 協( ) たことが確認された。MLF が生起した 09Se-A1-4 株 および PN4 株によるワインを比較すると,乳酸エチ ル含有量は 09Se-A1-4 株によるワインが 113 mg/ℓで あり,PN4 株によるワインに比較して 1.6 倍多い結果 であった。乳酸エチルは MLF が生起することによっ て増加するエステルであり,MLF が生起したワイン の香味に影響を及ぼす成分の一つであると考えられて いる25)。また,09Se-A1-4 株によるワインはクエン酸 の含有量が PN4 株によるワインに比べて多く,MLF の生起に伴う大きな減少は認められなかった。MLF では,リンゴ酸の消費の後に引き続いてクエン酸が消 費される。また,ジアセチルや酢酸の生成と密接に関 係し,消費されたクエン酸はこれらの成分に変換され ることが知られている3,4,27)。09Se-A1-4 株によるワイ ンはジアセチルや酢酸の含有量が PN4 株によるワイ ンに比較して少なく,クエン酸の消費量の違いが,菌 株間における成分の多少に影響を及ぼしている要因の 一つと推測される。研究員 12 名による官能評価の結 果,09Se-A1-4 株によるワインと PN4 株によるワイ ンの評点に有意な差異は認められなかった。しかし, PN4 株によるワインはわずかにバターやナッツ様の 複雑な香りが感じられたのに対して,09Se-A1-4 株に よるワインはフルーティーな香りを有し,両者は異な る酒質であるとする評価が得られた。 以上のことから,本研究でセミヨン種前期醪から分 離された 09Se-A1-4 株は,比較的低い pH のワイン発 酵醪においても速やかに MLF を生起できる乳酸菌と して有用であることが示唆された。 要約 1.セミヨン種ワイン発酵醪および醸造設備から乳酸 菌を分離し,そのリンゴ酸分解能を検討した。前期醪 から分離された 22 菌株中の 18 菌株に,後期醪から分 離された 24 菌株すべてに,醸造設備から分離された 10菌株中の 1 菌株,合計 43 菌株にリンゴ酸の分解能 が認められた。 2.16S rDNA の PCR-RFLP 解析および塩基配列解析 の結果から,前期醪から分離されたリンゴ酸分解能を 有する乳酸菌は,Lb. plantarum に,後期醪から分離 された乳酸菌は O. oeni に,ワイナリー醸造設備から 分離された乳酸菌は P. pentosaceus に分類される可能 性が極めて高いことが示された。 3.リンゴ酸分解率に及ぼす pH の影響について検討 したところ,前期醪から分離された 09Se-A1-4 株は pH 2.9 の条件下においても 90%以上の高い MLF 能を 有していた。 4.セミヨン種ブドウを原料とする小規模試験醸造に おいて,09Se-A1-4 株は速やかに MLF を生起し,さ らには,クエン酸の消費が少ない特徴を有する菌株で あることが認められた。 文献
1) C. R. Davis, D. Wibowo, R. Eschenbruch, T. H. Lee and G. H. Fleet:Am. J. Enol. Vitic., 36, 290-301(1985)
2) S. Maicas, J. V. Gil, I. Pardo and S. Ferrer: Food Res. Int., 32, 491-496(1999)
3) S. Q. Liu:J. Appl. Microbiol., 92, 589-601 (2002)
4) R. Bauer and L. M. T. Dicks:S. Afr. J. Enol. Vitic., 25 74-88(2004)
5) 乳酸菌研究集談会編:乳酸菌の科学と技術 pp.253-254,学会出版センター,東京(1996) 6) 柳田藤寿,鎌田 勉,篠原 隆,後藤昭二:醸
協,88,(3),238-244(1993)
7) S. Lafon-Lafourcade, E. Carre and P. Ribereau-Gayon:Appl. Environ. Microbiol., 46, 874-880 (1983)
8) L. Solieri, F. Genova, M. D. Paola and P. Giudu-ci:J. Appl. Microbiol., 108, 285-298(2010) 9) H. Vaillant, P. Formisyn and V. Gerbaux: J.
Appl. Bacteriol., 79, 640-650(1995)
10) J. C. Nielsen, C. Prahl and A. Lonvaud-Fu-nel:Am. J. Enol. Vitic., 47, 42-48(1996) 11) B. C. Rankine:Am. J. Enol. Vitic., 23, 152-158
(1972)
12) B. Martineau, T. E. Acree and T. Henick-Kling:Food Res. Int., 28, 139-143(1995) 13) I. Lopez, F. Ruiz-Larrea, L. Cocolin, E. Orr, T.
Phister, M. Marshall, J. VanderGheynst and D. A. Mills:Appl. Environ. Microbiol., 69, 6801-6807(2003) 14) http://blast.ncbi.nlm.nih.gov/Blast.cgi 15) 山梨県工業技術センター:葡萄酒醸造法,pp. 11-14,(2000) 16) 注解編集委員会編:第 4 回改正国税庁所定分析 法注解, pp. 62-79, 日本醸造協会,東京(1993)
17) T. Shinohara and M. Watanabe:Agric. Biol. Chem., 40, 2475-2477(1976)
18) 財団法人日本醸造協会:醸造物の成分,pp. 343-351,日本醸造協会,東京(1999)
19) H. Sato, F. Yanagida, T. Shinohara and K. Yokotsuka:J. Biosci. Bioeng., 90, 335-337 (2000)
20) G. Fleet, S. Lafon-Lafourcade and P. Ribereau-Gayon:Appl. Environ. Microbiol., 48, 1034-1038(1984)
21) 柳田藤寿,後藤亜紀人,芦沢 忠,内田多加夫, 篠 原 隆:J. Inst. Enol. Vitic. Yamanashi Univ., 30, 9-17(1995)
22) R. F. Kunkee: Am. J. Enol. Vitic., 18, 71-77 (1967)
23) G. E. Bousbouras and R. E. Kunkee:Am. J. Enol. Vitic., 22, 121-126(1971)
24) C. R. Davis, D. J. Wibowo, T. H. Lee and G. H. Fleet: Appl. Environ. Microbiol., 51, 539-545 (1986)
25) 柳田藤寿:ASEV Jpn. Rep., 6, 81-85(1995) 26) J. H. Swiegers, E. J. Bartowsky, P. A.
Hen-schke and I. S. Pretorius:Aust. J. Grape Wine Res., 11, 139-173(2005)
27) J. C. Nielsen and M. Richelieu:Appl. Environ. Microbiol., 65, 740-745(1999)