大豆タンパク質由来ペプチドの多面的生理機能の解 明に関する研究
中森, 俊宏
https://doi.org/10.15017/1441352
出版情報:Kyushu University, 2013, 博士(農学), 論文博士 バージョン:
権利関係:Fulltext available.
大豆タンパク質由来ペプチドの 多面的生理機能の解明に関する研究
中森 俊宏 2014
大豆タンパク質由来ペプチドの多面的生理機能の解明に関する研究
目次
第1章
緒論 ... 1
第2章 脂肪組織における大豆ペプチドの生理機能 第1節 緒言 ... 21
第2節 材料及び方法... 23
第3節 結果... 27
第4節 考察... 38
第5節 小括 ... 41
第3章 肝臓における大豆ペプチドの生理機能 第1節 緒言 ... 43
I
目 次
第2節 材料及び方法... 46
第3節 結果及び考察... 55
第4節 小括 ... 86
第4章 消化管内での大豆ペプチド(β-コングリシニン分解物)の生理機能 第1節 緒言 ... 88
第2節 材料及び方法... 91
第3節 結果及び考察... 95
第4節 小括 ... 101
第5章 脳の認知に及ぼす大豆ペプチドの効果 第1節 緒言 ... 102
第2節 材料及び方法... 104
第3節 結果... 110
第4節 考察... 119
第5節 小括 ... 121
II
第6章
飲料中でのペプチドの安定性に関する研究
第1節 緒言 ... 122
第2節 材料及び方法... 123
第3節 結果... 126
第4節 考察... 138
第5節 小括 ... 140
第7章 総括 ... 141
謝辞 ... 146
参考文献 ... 148
III
第1章 緒論
第1章 緒論
世界最大の油糧資源である大豆
大豆は縄文時代から栽培されている記録があり1-3)、今日に至るまで日本の食文 化の創成に深く貢献している。現在においては北海道から九州まで栽培されおり、
2012年度の国内の生産実績は236千㌧である4)。一方、2012年度の海外からの輸
入実績は2,727千㌧で主な輸入国はアメリカ、カナダ、ブラジル等である(Fig.1-1)
5)。輸入大豆のうち 932 千㌧は食品用途に使用され、残りは飼料用途である。輸
入大豆の主な食品用途は、みそ、醤油、豆腐・油揚げ、納豆、高野豆腐、豆乳等 で、食品用途に占める国産大豆の使用比率は 25%程度に留まっている。2012 年 度の世界の大豆生産実績は291百万㌧となり、油糧種子原料の中でも現在最も多 く生産されている。このうち飼料向けの採油用途が 80%であるが、食品用途は 20%で51百万㌧にとどまっている4)。
世界の 2012 年度の牛肉の生産量は 57 百万㌧、豚肉は 105 百万㌧、鶏肉が 78 百万㌧であり、食品向けの大豆の生産実績は牛肉の生産量と同程度である 6)。牛 肉と比べた時の大豆の生産性は、環境に対する負荷が少なく効率が良いとされる。
実際、大豆 1 ㌧を生産するために 1,000m3の水が必要とされるのに対して牛肉 1
㌧の生産には大豆に比べ16倍の水量が必要となる7)。また、農場の単位面積当た りのタンパク収量で比較すると大豆は牛に比べて17倍生産性に優れている8)。
1
大豆の組成
大豆の一般組成は、水分 12%、タンパク質 33%、脂質 21%、炭水化物 29%、
灰分5%である(Fig.1-2)9)。大豆タンパク質の主成分はグリシニンとβ-コングリ シニンであり10)、これ以外にリン脂質会合タンパク質、アルブミン、トリプシン インヒビターやアミラーゼなどの酵素、各種ペプチド11)及びアミノ酸を少量含ん でいる。大豆タンパク質は専ら Non-GMO(非遺伝子組み換え)大豆を搾油した 脱脂大豆より製造され(Fig.1-7)、畑の肉ともいわれるほどアミノ酸バランスに 優れた良質のタンパク質源であり9,10)、肉と比べてもアミノ酸組成に遜色はない。
以上、大豆は生産性及び栄養性に優れているものの、食品用途への利用は世界 大豆産量の2割に留まっていることも事実であり、環境負荷軽減や食糧資源確保 の面からも食品用途の割合を向上させることが喫緊の課題であるとされている。
2
第1章 緒論
出典:United States Department of Agriculture 「World Markets and Trade」
Fig.1-1 2011-2012年の世界の大豆生産量6)
3
Fig.1-2 大豆の組成12)
4
第1章 緒論
生活習慣病予防素材としての大豆タンパク質
世界保健機関(WHO)が発表した「2012 年世界保健統計」によると、先進国 を中心に肥満度が上昇し、世界の男性 10%、女性 14%がBMI30 以上の肥満と報 告されている13)。特に北米、中南米、中東、アフリカの一部(北アフリカ、南ア フリカ)は、BMI 30 以上の肥満者の割合が 30%を超えており(Fig.1-3)肥満に よる生活習慣病が増加している。日本は比較的肥満が少ない地域に入るものの、
男性の肥満者数が経年的に増加している傾向が見受けられる。死亡理由を疾病別 に見ると、2008 年度の世界の総死亡数 5,700 万人の63%にあたる 3,600万人が、
脳卒中や心疾患、糖尿病、高血圧症、動脈硬化などの生活習慣病及び癌が原因で 死亡している13)。特に、生活習慣病は中年から高齢者の間で急増しており、WHO は2030年までに生活習慣病を成因とする死亡数は5,500万人に増加し、心疾患に よる年間死亡数は 2,500 万人に増加すると予測している。心疾患予防としては血 中中性脂肪(TG)やコレステロールのコントロールによる動脈硬化の予防が奨励 されている13)。しかしながら、国民栄養調査によると日本人には血中TG濃度が
150 mg/dL 以上の高TG者が多く見受けられ、日本人の男性45%、女性33%が該
当する14,15)。特に日本人に多い内臓脂肪型肥満においては、血中高 TGによる動
脈硬化の進展が危惧されている16-18)。
大豆タンパク質の生理機能研究に関しては、Carroll らが血漿 LDL 濃度低下作 用を示すことを1975年に報告19)して以来多くの研究がなされ、Suganoらは難消 化性の高分子画分(HMF)が腸管内において過剰な胆汁酸と結合し体外に排泄す
5
るメカニズムを提唱している(Fig.1-4)20-26)。このような大豆タンパク質の積極 的な生体調節機能に基づき、米国食品医薬品局(FDA)は大豆タンパク質のコレ ステロールの低減効果に注目し、大豆タンパク質を1日あたり25 g含む食品につ いて「心臓病のリスクを低減する食品」という表示を認めている27)。また、我が 国においてもヒトでのコレステロール低減効果が認められ、特定保健用食品の関 与成分の一つとして認可されている28)。
大豆タンパク質によるトリグリセリド濃度調節作用に関しては、カゼインと比 較して血清及び肝臓のトリグリセリド濃度を有意に低下させるとの報告がある
29-30)。また、大豆タンパク質の長期摂取により脂肪組織重量も減少することから
肥満を改善する効果も期待されている31)。本抗肥満作用については、直接的作用 と し て の 血 中 イ ン ス リ ン 濃 度 の 低 下 と 、 そ れ に 伴 う 脂 肪 酸 合 成 制 御 転 写 因
子,SREBP-1 の発現抑制による肝臓での脂肪酸合成能の低下並びに肝臓脂肪の蓄
積抑制であると報告されている32-34)。しかしながら、肝臓での脂肪酸合成抑制作 用や白色脂肪細胞の肥大化抑制作用が大豆タンパク質そのものの作用か、あるい はその派生物(ペプチド、イソフラボン類)によるものかは解明されていない35-37)。
6
第1章 緒論
Fig.1-3 世界の肥満者の状況(BMI30以上の状況)
出典:WHO;http://gamapserver.who.int/mapLibrary/app/searchResults.aspx1
7
Fig.1-4 大豆タンパク質のコレステロール低下メカニズム21-26)
8
第1章 緒論
肥満予防素材としての大豆ペプチド
肥満は、過剰な摂取エネルギーが体脂肪として異常に蓄積することで起こる。
従って、肥満の治療や予防には、①エネルギー代謝を促進して体脂肪に蓄積した 脂肪を消費する、②脂肪合成を低減させ血中TG濃度を下げる、③食欲を抑え摂 取する食事量を制限することが肝要となる。Komatsuらは、肥満患者の食餌誘導 産熱に注目し、大豆ペプチドが大豆タンパク質やラクトアルブミン摂取と比較し て食餌誘導産熱の産生が高い傾向にあり、要因として交感神経系の調節作用並び に内分泌系の調節による可能性を指摘している38,39)。また、Toyoshimaらは、運 動後の大豆ペプチド摂取によって皮下脂肪の減少が特に誘導されることを報告
している40)。Tamaruらは、大豆ペプチドの低分子画分がラットの血清及び肝臓
TG濃度を低下させることを報告しているが41)、これまでに大豆タンパク質由来 のTG合成抑制ペプチドが単離同定されたとの報告はない。ペプチドの腸管吸収 に関しては、ジからトリまでの鎖長が吸収限界と考えられている42-45)。これまで に単離同定されている生理活性ジ、トリペプチド類をTable1-1にまとめた。大豆 由来のジ、トリペプチドに関してはは抗炎症作用を示すVPY46)や血圧低下ペプチ ドであるSVYが報告されている47)が、TG低下作用を示すジ、トリペプチドにつ いての報告例は見当たらない。
消化管ホルモンの一つであるコレシストキニン(CCK)は、CCK受容体に結合 し、迷走神経を通じて中枢の満腹感を刺激することが知られている。Haraらは大 豆タンパク質のペプシン分解物からCCKの分泌を促進するペプチドを同定し、
9
動物試験から摂食量を減らすことを明らかにした55)。CCKのみで満腹中枢を刺 激することは不可能であるが、肥満治療などにおいて食事量のコントロールに有 効な補助手段であると考えられる。
以上の学術的背景に鑑み、本研究の第2章~第4章では脂肪組織、肝臓及び消 化管での大豆ペプチドの生理作用を明らかにすることを目的として研究を遂行 した。
10
第1章 緒論
Table1-1 食品タンパク質由来の生理活性ジペプチド及びトリペプチド
Source Peptide sequence Function Reference
Soy protein VPY Anti-inflammative 46
〃 SVY Anti-hypertensive 47
Sardine muscle VY 〃 48
Fermented milk IPP 〃 49
〃 VPP 〃 〃
Wakame YH 〃 50
〃 KY 〃 〃
〃 FY 〃 〃
〃 IY 〃 〃
Rapeseed VW 〃 51
Spinach rubisco MRW 〃 52
Egg ovalbumin LW 〃 53
Rice albumin LPR Immunomodulating 54
11
脳に対する大豆ペプチドの効果
日本は長寿化と人口減少が同時に進行しており、世界で最も高齢化が進んでいる 先進国の一つである。今後65歳以上の人口比率が増加し2060年には40%に到達す るとされ、75歳以上の後期高齢者人口は2060年には2,336万人に到達すると予測さ れている(Fig.1-5)のに伴い認知症等の脳機能性疾患の増加が大いに懸念されてい る。65歳以上の高齢者人口に占める認知症高齢者は、2010年には280万人(65歳以 上人口の 6.3%)であったが、その後患者数は年々増加しており、2020 年には 289 万人(同8.4%)、2025年には323万(同9.3%)に推移すると予測されている(Fig.1-6)。
Yoshidaらは、加齢の影響を大いに受ける前頭前野を活性化することが可能となれ
ば、認知機能低下を遅延させることが可能であると報告している 58,59)。近年の食機 能研究により、食品成分による脳機能改善の可能性が提示されつつあり、脳機能を 改善する食品(ブレインフード)の研究が今後大いに進展することが期待されてい る。そこで、第5章ではブレインフードとしての大豆ペプチドの可能性について究 明を行った。
12
第1章 緒論
Fig.1-5 日本の人口の構成比の推移56)
13
Fig.1-6 介護領域での認知症患者数の推移57)
14
第1章 緒論
大豆ペプチド含有飲料での沈殿、保存安定に関する研究
大豆ペプチドは、脱脂大豆から調整された大豆タンパク質を原料に微生物由来酵 素を用いて加水分解により製造される。酵素分解された大豆タンパク質は、平均分
子量10,000程度の不溶性画分(以下、HMFと略)と平均分子量500程度の可溶性オ
リゴペプチド画分(以下、大豆ペプチドと略)に分離され、可溶性オリゴペプチド 画分は、最終粉末化されて大豆ペプチドとなる(Fig.1-7)。通常、大豆タンパク質を 酵素分解すると強い苦味を呈するが、大豆ペプチドは、使用する酵素及び原料の調 整により苦味が低減化されている。
大豆ペプチドの物理化学的特徴として、液性状は透明であり(Fig.1-8)、溶解時の 溶液粘度が低く、同組成のアミノ酸混合物と比べて浸透圧上昇が少ない。また、溶 解度の低いアミノ酸の水溶性が改善されるなどの利点もある(Fig.1-9)。これらの点 から、アミノ酸の補給源として飲料等に用いられるが、冷蔵、酸性域では沈殿が発 生するなどの問題があった。飲料用途には高い保存安定性が求められていることか ら、第6章では大豆ペプチド飲料で発生するオリ成分の同定を試み、保存安定性に 関する検討を行った。
15
Fig.1-7 大豆タンパク質及び大豆ペプチドの製造プロセス 大豆ペプチド
16
第1章 緒論
Fig.1-8 大豆ペプチドの液性状
8%大豆タンパク質 8%大豆ペプチド
17
Fig.1-9 大豆ペプチドと各アミノ酸の溶解性の比較
18
第1章 緒論
以上を勘案し、本研究では大豆タンパク質の高度利用を通して人々の健康や生活 の質向上に貢献することを命題として、大豆タンパク質由来ペプチドの多面的な生 理機能の解明を図った。以下、本研究の詳細な内容を列記する。
(1) 第2章では、脂肪組織のエネルギー代謝に関係するミトコンドリアの脱共役蛋 白質(UCP)-1 に対する大豆ペプチドの影響を検証した。その結果、大豆ペプ チドは、褐色脂肪細胞のUCP-1活性を高め、さらに白色脂肪細胞のUCP-1活性 化を通じて褐色化様作用を促進しエネルギーの消費を亢進させる可能性を示し た。また、大豆ペプチドはアディポネクチンの分泌を有意に高めることを見出 した。
(2) 第3章では、肝臓でのトリグリセリド合成における大豆ペプチドの効果を検証 し、作用成分の単離同定を試みた。その結果、大豆ペプチドに含まれるうま味 ペプチドは肝臓でのトリグリセリド合成活性を低下させることが判明した。さ らに、うま味ペプチド中からジペプチドSYを単離同定し、このSYがトリグリ セリドの合成活性化を低下させアポプロティンB100の分泌を抑制することを明 らかにした。
(3) 肥満を予防するには食事の摂取量を減らす必要がある。第4章では、β-コングリ
シニンの Bromelain 分解物がヒトでの実証試験において満腹感を誘発させるこ
とを示した。β-コングリシニンのBromelain分解物は呈味性も優れていることか ら、食事制限が必要な肥満患者や糖尿病患者の過食防止や空腹感を和らげる補
19
完的な使い方が期待出来る。
(4) 第5章では、脳の認知機能に対する大豆ペプチドの摂取効果をヒトで検証した。
その結果、新しい情報を順次適切に処理する脳機能の維持に効果があることが 判明した。この効果は、大豆ペプチド摂取がやる気や情報処理効率の維持に資 する可能性を示唆する知見であった。
(5) 第6章では、大豆ペプチド飲料保存中に発生するオリの成分同定並びにその生 成機構の解明を行った。その結果、オリの構成成分は β-コングリシニンのαサ ブユニット310-362番を由来とするポリペプチドであった。これらポリペプチド を限外ろ過膜法により除去することによってオリ生成が発生しない高品質ペプ チド飲料の開発と提供が可能となった。
以上、本研究によって、大豆タンパク質由来ペプチドの多面的生理機能を明示す ることが可能となり、オリ発生のない高品質飲料の提供を通じて人々の健康や生活 の質向上に貢献出来ることが大いに期待される。
20
第2章 脂肪組織における大豆ペプチドの生理機能
第2章 脂肪組織における大豆ペプチドの生理作用
第1節 緒言
マウスやラットは、肥満の進行によって脂肪組織に過剰の脂肪が蓄積され、腹腔 内や皮下に存在する白色脂肪組織(WAT)の肥大化が促進される。他方、頚背部付 近に局在する褐色脂肪組織(BAT)では脂肪蓄積よりはむしろ熱産生系に傾くこと が知られている。
Saitoらは肥満マウスを用いた試験により、大豆ペプチドが交感神経系でのノルア
ドレナリン分泌の増大を促し、BAT での熱産生亢進によりエネルギー消費を促すこ とを報告している 62-64)。BAT の熱産生に重要な役割を担う脱共役タンパク質
(uncoupling protein:UCP)-1はミトコンドリアでの酸化的リン酸化を脱共役させて 酸化エネルギーを熱として発散させ、エネルギー消費に向かわせる 65,66)。UCP-1 の 発現と肥満には正の相関関係が成立し、UCP-1 発現を低下させたマウスでの肥満化 と高発現マウスでの抗肥満化が実証されている 67)。しかしながら、これまで大豆ペ プチドが脂肪組織のUCP-1発現に及ぼす影響に関しては報告例がなかった。そこで、
本章では大豆ペプチド摂取によるUCP-1発現挙動について、ラット及びラット腸管 由来初代内臓脂肪細胞を用いて詳細に検討した。
他方、腹腔内WATから分泌されるアディポサイトカインの中でも特にアディポネ クチンは、インスリン感受性を高め 68)、脂肪細胞の肥大に伴い産生が抑制されるこ
21
とが報告されている 69-73)。これまでに、大豆タンパク質摂取がアディポネクチンの 分泌量を増加させるとの興味ある知見が報告されている 74,75)。そこで、本章では大 豆ペプチド機能としてアディポネクチン分泌への影響についても検討を加えた。
22
第2章 脂肪組織における大豆ペプチドの生理機能
第2節 材料及び方法
1. 動物試験及び飼育方法
ラット(雄性Wistar系ラット, 6 週齢)は日本クレア㈱(東京)より購入した。ラ ットは、購入後、23±1℃、55±5%の管理環境下、ステンレス製個別ケージにて 1 週 間の予備飼育を行った。その後、1群 5 匹となるように CAS(Casein:オリエンタ ル酵母工業㈱、東京)、SPI(Soy protein isolate:不二製油㈱、大阪)、SPH(Soy peptide: ハイニュート-AM、不二製油㈱)の3グループに群分けし、AIN-76組成に基づき 各群のタンパク質含量が20%、油分10%、スクロース50%となるように調整した食 餌を作製した(Table2-1)。2週間の試験期間中、ラットには自由摂食及び自由摂水と させた。摂食量及び体重は毎日測定した。なお、動物試験は、「動物の愛護及び管理 に関する法律の一部を改正する法律(平成17年法律第68号)」による「実験動物の 飼養及び保管並びに苦痛の軽減に関する基準(平成 18 年環境省告示第 88号)」、及 び文部科学省が策定した「研究機関等における動物実験等の実施に関する基本指針
(平成18年6月1日告示)」に基づき、日本学術会議が作成した「動物実験の適正 な実施に向けたガイドライン(平成18年6月1 日通知)」を踏まえて作成された社 内動物実験の実施基準に基づいて実施された。
2. 血液及び組織試料
2週間の試験期間終了後、各ラットを絶食させたのち、イソフルラン麻酔の無痛条 23
件下において腹部大動脈より採血を行った。臓器及び腹腔内脂肪、褐色脂肪を摘出 し、重量を測定後、実験に供するまで—80℃で保存した。
3. 脂肪組織での遺伝子発現量の測定
ISOGEN(日本ジーン㈱製、東京)RNA 抽出キットを用いて、ラットより摘出
した脂肪組織からtRNAを抽出した。その後、PrimeScriptR RT regent(TaKaRa BIO
㈱、京都)を用いて逆転写を行い、cDNA を得た。標的とする各遺伝子の発現は ABI 7300 real-time PCR system(Applied Bio Systems, USA)及びPremix Ex TaqTMキ
ット(TaKaRa㈱)による遺伝子プライマーセットを用いたSYBR Green検出によ
り行った(βアクチン(Actb)を指標)。用いたプライマーは以下の通りである。
UCP-1(F)– AATCAGCTTTGCTTCCCTCA, UCP-1(R) – GCTTTGTGCTTGCATTCTGA, UCP-2(F) – GCCACTTCACTTCTGCCTTC, UCP-2(R)– GAAGGCATGAACCCCTTGTA, UCP-3(F) – GAGAACCCAGGAGTGCAGAG, UCP-3(R) – GGGGGTGTAGAACTGCTTGA, Actb(F) – AGCCATGTACGTAGCCATCC, Actb(R) – CTCTCAGCTGTGGTGGTGAA
24
第2章 脂肪組織における大豆ペプチドの生理機能
4. 血液生化学項目と血中アディポネクチン量の測定
ラット血液は直ちに遠心分離(1,900×g, 15 min, 4℃)し、血漿を得た。なお、得 られた血漿は分析に供するまで—80℃にて保存した。アディポネクチン濃度は ELISA キット(mouse/rat adiponectin ELISA kit; Otsuka Pharmaceutical, Tokyo)にて 測定した。血漿中の中性脂肪、総コレステロール及び血糖は、Fuji DRI-CHEM 7000V auto-analyzer(Fuji Film Co., Tokyo)を用いて一斉解析した。
5. 内臓脂肪細胞の培養
ラット腸管膜由来の初代内臓脂肪細胞(㈱プライマリーセル: LotNo.ILKA-F)を購 入し、使用培地には内臓脂肪分化メディウムver. 1(以下「メディウム」)(㈱プライ マリーセル:LotNo.JCRA-M)を用いた。
また、被験試料である SPH は 10%濃度となるように 1%メディウム含有 DMEM 培地(ダルベッコ・フォークト変法イーグル培地)に溶解し、フィルター滅菌したもの を用いた。また、UCP-1発現誘導剤である(-)-ノルエピネフリン酒石酸(SIGMA社製、
以下NEと略)は、1×10-6mol/Lの濃度に調製して使用した。
培養は、初代内臓脂肪細胞(3.0×106cells)を12穴プレートに播種(2.4×105cells/well) した後、37℃、5%CO2条件下で4日間の分化誘導を行った。その後、SPH含有メデ ィウム培地(100μg/mL)にて培養を 8 日間継続した。なお、コントロールには 1% メディウム含有DMEM培地を用いた。最終培養日には、NEを1×10-6 mol/L含むメ ディウム培地に交換後、6時間の刺激培養を行った。
25
6. 形態観察
SPH 添加後、及び NE 添加前後に顕微鏡にて細胞形態観察を行い、代表例につい て写真撮影を行った。
7. UCP-1遺伝子発現の測定
細胞からのRNA抽出はTRI ReagentTMRを用いてマニュアルに準じて行った。逆 転写反応はRevertAid™First Strand cDNA Synthesis Kitを用いてcDNAを合成した後、
Real-time PCR 法により遺伝子発現量を求めた(GAPDH を指標)。プライマーは
Oligo@SIGMA GENOSYS-PCR(シグマアルドリッチジャパン㈱)を使用した。配列
情報は以下を通りである。
GAPDH(F)-AATGTATCCGTTGTGGATCTGA, GAPDH(R)– GCTTCACCACCTTCTTGATGT, UCP-1(F)− GCCTGCCTAGCAGACATCAT, UCP-1(R)− TGGCCTTCACCTTGGATCT
8. 統計処理
実験1~4の結果は平均値±標準誤差で示し、統計処理はTukey HSD検定を行った。
実験5~7の結果は平均値±標準誤差で示し、統計処理はStudent’s-t検定を行った。
26
第2章 脂肪組織における大豆ペプチドの生理機能
第3節 結果
第1項 Wistarラット脂肪組織に及ぼすSPHの作用
Wistar ラットの体重に及ぼすSPHの影響
摂食量において、SPI(Soy protein isolate)群は、期間を通じてコントロールであ るCAS(Casein)群、SPH(Soy peptide)群より高く、一日平均摂食量で有意に高か った。SPI群では体重増加も大きく終体重において他の群に比べ重かったが、飼料効 率ではCAS群と差がなく、SPH群の方が有意に低い値を示した(Table2-2)。
血液生化学項目
SPI群及びSPH群では血中コレステロールは本試験条件での高スクロース食では CAS群に比べて有意な低下が認められた。一方、血中中性脂肪では有意差は認めら れなかったものの、SPH群ではCAS群及びSPI群に比べて低い傾向にあった。血糖 値についてはSPH群及びSPI群ではCAS群に比べて低下傾向にあった(Fig.2-1)。
SPH摂取が脂肪組織重量に及ぼす影響
各飼料群が体内脂肪量に及ぼす影響をFig.2-2にまとめて示した。2週間の摂食期 間では体内白色脂肪合計量(睾丸周囲、腎周囲、腸管膜周囲)に有意差は認められ
27
なかったものの(Fig.2-2A)、頚背部褐色脂肪組織量(wt%)はSPI及びSPHの摂取 によって増加する傾向が認められた(Fig.2-2B)。
SPH摂取が褐色脂肪組織におけるUCPs mRNA発現におよぼす影響
UCPファミリーのmRNA発現量を測定したところ、UCP-1のmRNA発現におい てSPH群がコントロールであるCAS群、さらにSPI群に比べても高い傾向を示した
(Fig.2-3)。UCP-2及びUCP-3についてもUCP-1と同様にSPH群で高い傾向を示し た(Fig.2-3)。
SPH摂取が血中アディポネクチン量に及ぼす影響
血中アディポネクチン量はCAS食に比べて、SPI群及びSPH群で高い傾向にあり、
SPH群では有意に分泌量の増大が認められた(Fig.2-4)。
28
第2章 脂肪組織における大豆ペプチドの生理機能
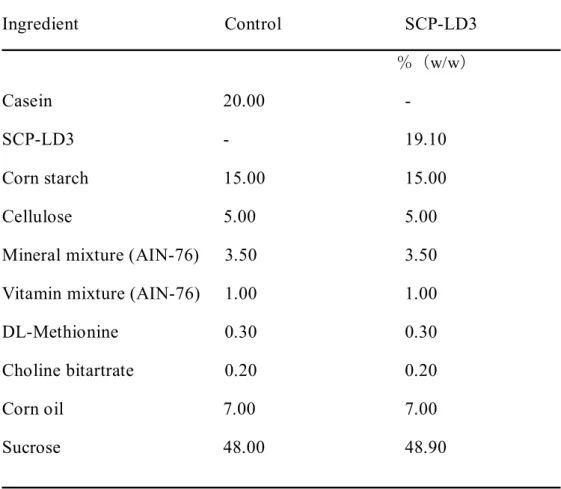
Table2-1 実験1での食餌組成
CAH SPI SPH
(w/w)%
カゼイン#1 22.70 - -
大豆タンパク質#2 - 23.39 - 大豆ペプチド#3 - - 23.07 スクロース 53.05 52.36 52.68
コーン油 10.00 10.00 10.00
セルロース 9.50 9.50 9.50
ミネラル#4 3.50 3.50 3.50
ビタミン#5 1.00 1.00 1.00
重酒石酸コリン 0.25 0.25 0.25
合計 100.00 100.00 100.00
#1;カゼイン(オリエンタル酵母工業㈱、東京) 粗タンパク質として88.1%含む。
#2; 大豆タンパク質(不二製油㈱、大阪)粗タンパク質として85.5%含む。.
#3; 大豆ペプチド(ハイニュート-AM)(不二製油㈱、大阪)粗タンパク質として86.7%、
遊離アミノ酸2%以下、ジ、トリペプチド含量65%以上
#4; AIN-93G-MX組成
#5; AIN-93-VX組成
29
Table2-2 SPHを含む食餌がWistarラットの摂食量及び体重増加に及ぼす影響
CAH SPI SPH
(n) 5 5 5
初体重(g) 162.5±1.5 162.4±1.5 162.6±1.6
終体重(g) 230.7±3.8 238.6±3.5 225.9±2.9
食餌量(g/日) 15.8±0.2a 17.9±0.2b 16.4±0.4a
飼料効率 0.288±0.008a 0.284±0.009a 0.257±0.003b
各ポイントと誤差表示はそれぞれ平均と標準誤差を示す(n=5)。 a, b; Tukey HSD検定 (p < 0.05)。
30
第2章 脂肪組織における大豆ペプチドの生理機能
Fig.2-1 SPH摂取が血漿中の中性脂肪、総コレステロール及び血糖に及ぼす影響
各ポイントと誤差表示はそれぞれ平均と標準誤差を示す(n=5)。 a, b; Tukey HSD検定(p < 0.05)。
CAS SPI SPH
CAS SPI SPH CAS SPI SPH
B; 総コレステロール C; 血糖 A; 中性脂肪
mg/dL mg/dL mg/dL
31
Fig.2-2 SPH摂取が腹腔内脂肪及び褐色脂肪に及ぼす影響
CAS SPI SPH CAS SPI SPH
A; 腹腔内脂肪 B; 褐色脂肪
wt% wt%
32
第2章 脂肪組織における大豆ペプチドの生理機能
Fig.2-3 SPH摂取が褐色脂肪組織(BAT)のUCP-1mRNA発現に及ぼす影響 各ポイントと誤差表示はそれぞれ平均と標準誤差を示す(n=5)。
CAS SPI SPH
Arbitrary unit
33
Fig.2-4 SPH摂取が血中アディポネクチン量に及ぼす影響
各ポイントと誤差表示はそれぞれ平均と標準誤差を示す(n=5)。 a, b; Tukey HS検定(p <0.05)。
μg/mL
CAS SPI SPH
34
第2章 脂肪組織における大豆ペプチドの生理機能
第2項 初代内臓脂肪細胞におけるSPHの作用
細胞形態観察
コントロール群の形態観察から本試験系では初代内臓脂肪細胞としての分化状態 は問題なく、正常な細胞系試験であると判断された(Fig.2-5)。
培養期間を通した目視観察の結果、SPH 群ではコントロール群と比較して肥大が 抑制され脂肪蓄積した細胞が少ない傾向にあった。また、培養8日目のNE刺激にお いてコントロール群ではNE刺激後、細胞が肥大化し微細な脂肪滴を有する細胞が観 察されたが、SPH群では細胞の肥大化は観察されなかった(Fig.2-5)。
SPHがUCP-1遺伝子発現に及ぼす影響
初代内臓脂肪細胞に対する SPH 添加により、コントロール群と比べて有意な
(p<0.05)UCP-1の発現増加が認められた(Fig.2-6)。
35
Fig.2-5 SPH添加後、培養4日目、6日目、8日目並びにNE添加前後の内臓脂肪 細胞の顕微鏡写真
スケールバーは100μm
培養6日目
A: コントロール培養4日目 培養8日目NE添加前 培養8日目NE添加後
B: SPH 培養4日目 培養6日目 培養8日目NE添加前 培養8日目NE添加後
36
第2章 脂肪組織における大豆ペプチドの生理機能
Fig.2-6 SPHが内臓脂肪細胞のUCP-1 mRNA発現に及ぼす影響 Student’s-t検定 **p < 0.01
Control SPH
Arbitrary unit
**
37
第4節 考察
大豆ペプチドの摂取は、高脂肪食による肥満マウスにおいて食餌エネルギー制限 下で、他のタンパク質素材のペプチドと比べて優れた減量効果を示すとの報告があ る82)。その作用機序の一部は肝臓におけるTGの合成抑制によると考えられている が41)、エネルギー代謝系に及ぼす影響については未解明であった。そこで、本研究 では、高スクロース食での摂食試験を実施し、これら観点からの大豆ペプチド摂取 の効果を検証した。その結果、Wistar系ラット、6週齢を用いた2週間の試験期間で、
体重については減量効果が認められなかったが、飼料効率では大豆ペプチド摂取に よって明らかな抑制効果が認められたことから、長期間の摂取によって肥満の抑制 効果が期待される知見を得ることができた。
肥満は、運動量の低下によるエネルギー消費の減少や必要量以上のエネルギーの 摂取により誘因されると言われている。食品成分の中には、摂取しエネルギーの消 費を促す因子が存在することが数多く報告されている83)。例えば、カプサイシンを 摂取すると中枢神経を刺激することにより副腎皮質からノルアドレナリン分泌を促 進し、熱産生を誘導することが知られている84)。
大豆タンパク質摂取によってUCP-1の発現が増加するとの報告がなされているが
85,86)、本研究においてもコントロール群と比較して、大豆系タンパク質を摂取するこ
とによりBATにおけるUCP-1の発現が摂食後6時間で高くなる傾向が認められた。
UCP-1の誘導は絶食/再摂食後2時間で最高となり、誘導速度が極めて早いとの報
38
第2章 脂肪組織における大豆ペプチドの生理機能
告があることから87)、SPH摂取によるUCP-1の誘導を明らかにするためには、短時 間刺激での挙動をさらに検討する必要があるが、本章で得られた結果はBATにおけ る熱産生亢進作用にSPHが関与している可能性を強く示唆するものであった。
白色脂肪組織(WAT)は脂肪の単なる蓄積組織ではなく、種々のアディポサイト カインを分泌する代謝調節組織であることが報告されている88)。とりわけ、アディ ポネクチンは、肝臓における脂肪燃焼や筋肉における糖代謝亢進(糖取込み増強)
などあらゆる組織においてインスリン抵抗性の改善作用を示すことが報告されてい る89)。本研究において血中アディポネクチン量が大豆ペプチド摂取によって分泌増 大傾向を示したことから、SPHには血中アディポネクチン分泌を促す機能性ペプチ ドが含まれる可能性が強く示された。Masakiらはマウス(KK-Ay)を用いた試験に おいて、アディポネクチンの投与により褐色脂肪組織(BAT)でのUCP-1のmRNA 発現が誘導されることを報告していることから90)、SPIを酵素分解することによって 得られるペプチド、すなわちSPHが、WATでのアディポネクチン産生を介してBAT
のUCP-1の発現を活性化させている可能性が示された。
従来、UCP- 1は脂肪を熱に変換するBATにのみ発現すると考えられていたが、
CousinらはWATにおいてもある種の薬理的な刺激によりUCP-1が発現されること
を見いだした76)。WATでUCP-1の発現が活性化され、Browning化あるいはBRITE
(Brown in white)細胞に誘導されることは、肥満改善の点で注目されている77-79)。
本研究において、SPHによりNE刺激による白色脂肪細胞のUCP-1発現量がコント ロールと比較して2.5倍高くなったことから、SPHにはUCP-1発現を誘導するβ3ア
39
ドレナリン受容体のアゴニスト、あるいはPPARγのアゴニストとして作用するペプ チドが存在する可能性が初めて明らかとなった。
40
第2章 脂肪組織における大豆ペプチドの生理機能
第5節 小括
A soy protein isolate (SPI) diet has been reported to lowerthe serum and hepatic lipid levels, as well as to lowerthe weight gain and body fat accumulation forobesty-indced rat experiments.In Chaper 2, anenzymatichydrolysate of SPI (SPH) diet was also found to possess some physiological benefits,assimilar to SPI-diet. Namely,
Experiment I; Male Wistar rats (n=15) divided into CAS, SPI and SPH groups were maintained on a high-sucrose diet containing 20% crude protein. After two weeks, tissue weights, and uncoupling protein-1 (UCP-1) gene expression in brown adipose tissue (BAT) were evaluated, as well as plasma lipid and adiponectin levels.
Experiment II;Primary visceral adipose cells, a model for white adipose tissue (WAT), were cultured in medium containing SPH, and UCP-1expression was measured.
Following results were obtained.
Experiment I; Animals on SPH diet showeda significantly higher plasma adiponectin level than that of contrl group. SPH-dietsignificantly induced upregulation of UCP-1 expression in BAT.
Experiment II;Reduced adipocyte size andupreguraed UCP-1 mRNA expression by 100μg/mL SPH was observed for adipose cell-experiments.
These rsults strongly indicate that SPH could stimulate the UCP-1 mRNA expression in WATand thenelevate plasma adiponectin levels. These findings provide aninsight into the
41
regulation of adipose tissue function by peptides, and could be useful for developing anti-obesty foods by soybean peptides.
42
第3章 肝臓における大豆ペプチドの生理機能
第3章 肝臓における大豆ペプチドの生理機能
第1節 緒言
肥満の進行に伴い、脂肪組織、特に内臓脂肪から肝臓への遊離脂肪酸流入が増加 し、肝臓でのトリグリセリド(TG)の合成が促進される。また、内臓脂肪の肥大化 に伴うインスリン感受性の低下によっても同様の事象が起こることが知られている。
肝臓での脂肪酸の供給過多は、超低密度リポタンパク質(very low density lipoprotein, VLDL)の合成・分泌を促進させ91)、血中のTG濃度を上昇させることから、血中の 過剰のTGは、動脈硬化の進展を亢進させる要因の一つと考えられている。従って、
食品の持つ機能性を活用して血中の TG 濃度を正常値に低下させることが強く望ま れている。
Aoyamaらは、大豆ペプチドは大豆タンパク質と比較して肝臓でのTG低下作用が
強く発現することをラットによる動物試験によって明らかにした。しかしながら、
大豆ペプチド中に生理活性ペプチドが存在する可能性を示したが 30)、活性本体の同 定には至っていない。そこで、本章では脂質、特にTG低下作用を示す大豆タンパク 質由来ペプチドの検索と同定を目的とした。
TG低下ペプチドを単離同定するに当たり、まず大豆タンパク質のアミノ酸配列に 注目し、疎水性アミノ酸含量の高いペプチド画分と親水性アミノ酸含量の高い画分 とに分画し、両画分におけるTG低下作用を検証した。疎水性アミノ酸含量の高い画
43
分と親水性アミノ酸に富む画分に分画するに当たっては疎水性吸着樹脂を用いた。
TG低下作用の検証にはHepG2を用いた細胞培養系を用い、細胞内のTG合成量 を指標とした。HepG2細胞は、ヒト肝臓実質細胞が示す多様な生物学的機能 92)と血 漿リポタンパク質の合成及び分泌の栄養応答性調節機能 93,94)を保持しており、ヒト 肝臓実験モデルとして広く利用されている。また、TG低下作用を動物レベルで検証 するため、肥満・高脂血症の病態モデルである Otsuka Long-Evans Tokushima Fatty
(OLETF)ラットを用いた肥満モデル系を用いて検証した。
食事由来のタンパク質は胃液や膵液中のプロテアーゼにより分解された後、アミ ノ酸はアミノ酸輸送体を通じて吸収され、ペプチドは PepT1 と呼ばれるペプチド輸 送体を介して吸収される42,43)。PepT1は基質特異性が低いが、分子量に依存しており、
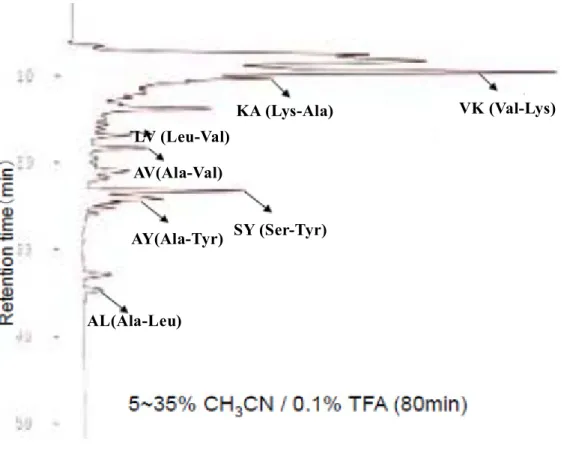
ジペプチド、トリペプチドの鎖長までを認識する 44)特徴がある。従って、体内に取 り込まれ、門脈を通じて肝臓に到達しTG低下作用を発揮することが出来る最小のペ プチド単位は、ジペプチドかトリペプチドであると考えられる。そこで、本研究で は、活性画分をゲルろ過クロマトグラフィー(GPC-HPLC)によってジ、トリペプチ ド画分に予め分画し、活性が発現する最小単位であるジペプチドあるいはトリペプ チドでTG低下作用を示すペプチドを単離し同定することを目的とした。
ところで、アポリポタンパク質B100(ApoB100)は肝臓で生合成されるVLDLの 必須構成タンパク質であり、ApoB100 の細胞内代謝には TG 合成が強く関与するこ とが知られている95-100)。また、ApoB100濃度は動脈硬化症の発症率と正の相関を示 すことが知られており、その過剰分泌は独立した危険因子と考えられている101)。
44
第3章 肝臓における大豆ペプチドの生理機能
そこで、本研究ではHepG2細胞でのTG合成低下作用の指標に、ApoB100分泌の 指標を加え、大豆ペプチド中のジ、トリペプチド画分からTG低下ペプチドの単離・
同定を試みた。
45