https://doi.org/10.18960/hozen.1803
西表島周辺における熱帯性海草ウミショウブ Enhalus acoroides (L.f.)
Royle の有性繁殖:分布北限域の開花時期、結果率および果実の生残
水谷 晃
1,*・井上 太之
1・玉本 満
2・北原 侑治
3・藤吉 正明
4・村上 智一
5中瀬 浩太
6・河野 裕美
1 1東海大学沖縄地域研究センター 2東海大学大学院海洋学研究科 3東海大学海洋学部 4東海大学教養学部 5 (研)防災科学技術研究所 6五洋建設株式会社環境事業部Sexual reproduction of the tropical seagrass Enhalus acoroides (L.f.) Royle around Iriomote Island, Japan: Flowering period, fruit set, and fruit survival at the northern limit of the species
Akira Mizutani1,*, Hiroyuki Inoue1, Michiru Tamamoto2, Yuji Kitahara3, Masaaki Fujiyoshi4, Tomokazu Murakami5,
Kota Nakase6 and Hiroyoshi Kohno1 1 Okinawa Regional Research Center, Tokai University 2 Graduate School of Marine Science and Technology, Tokai University
3 School of Marine Science and Technology, Tokai University 4 School of Humanities and Culture, Tokai University 5 National Research Institute for Earth Science and Disaster Resilience
6 Penta-Ocean Construction Co., Ltd
要旨:熱帯性海草ウミショウブ Enhalus acoroides の北限分布域における有性繁殖の季節性を明らかにすることを目的 とした。八重山諸島の西表島において 2013 年から 2017 年まで開花時期を観察した。2014 年と 2015 年には、4×4 m 調査区を 3 ヵ所ずつ設置して、開花から種子放出までの経過を記録した。開花は、1 年で比較的水温が高く、日照時 間が長く、日中の干潮位が低い 5 月から 11 月までにみられた。開花は日中の干潮位がより低い大潮周期に同調したが、 10 月以降ではその潮位は高く、受粉成功に至らなかった。シュートの年開花数は 1 回であった。開花から果実裂開 までの期間は、平均 84±6.5 日であった。雌の開花シュートのうち、開花 3 日後に花が残存した割合は 51.2%であり、 その後 60 日まで生残した割合は 9.8%、最終的に果実の裂開に至った割合は 9.3%であった。ほぼ周年にわたり開花・ 結実する熱帯域と比較して、開花期は短く、果実成熟にいたる有効な開花は開花期の前半に限られた。この結果、本 海域における有性繁殖による生産性はより低いことが示唆された。 キーワード:海草群落の保全、開花フェノロジー、干潮位、水温、日照時間
Abstract: The phenology of the sexual reproduction of the tropical seagrass Enhalus acoroides (L.f.) Royle was studied at the northernmost limit of its distribution. Flowering seasons were recorded from 2013 to 2017 at Iriomote Island, Yaeyama Islands, Japan. Fruit development was monitored in 2014 and 2015. The flowering season was from May to November, when the water * 〒 907-1541 沖縄県八重山郡竹富町上原 870-277 東海大学沖縄地域研究センター
Okinawa Regional Research Center, Tokai University, 870-277 Uehara, Taketomi, Yaeyama, Okinawa 907-1541, Japan
e-mail: [email protected] 2018 年 2 月 4 日受付、2019 年 5 月 16 日受理、2019 年 11 月 8 日早期公開(J-STAGE)
調査報告
はじめに
トチカガミ科ウミショウブ Enhalus acoroides (L.f.) Royle は、海水域に生育する沈水性の多年草で、インド 洋と西部太平洋の亜熱帯および熱帯域に広く分布する (Green and Short 2003)。日本では琉球列島南部の八重山 諸島にのみ生育し(Kuo et al. 2006;大場・宮田 2007; 当真 1999)、ウミショウブの分布北限にあたる。 ウミショウブ群落は、生物の餌場や産卵場となるなど (例えば、Nakamura et al. 2003;Nakamura and Sano 2004,
2005;土屋 1981)、沿岸生態系に重要な機能を提供し、 漁場として利用されている。また、夏季にウミショウブ が一斉に開花し、多数の雄花が水面を漂う様子は、環境 教育の材料や観光の資源としても利用されている。 本種は、熱帯域では緊急な保全が必要な種とはみなさ れていない(Short and Waycott 2010)。一方、日本の個 体群は、遺伝的にフィリピン東側の個体群と比較的近い とされるものの(Nakajima et al. 2014)、主要な生育地は 西表島に限られ(大場・宮田 2007;当真 1999)、環境省 や沖縄県のレッドデータブックでは絶滅危惧Ⅱ類(VU) として掲載されている(環境省自然環境局野生生物課希 少種保全推進室 2015;沖縄県文化環境部自然保護課 2018)。 西表島では、近年、草食性アオウミガメ Chelonia mydas の被食による群落の衰退や消失が急速に進行し (環境省那覇自然環境事務所・東海大学沖縄地域研究セ ン タ ー 2016, 2017, 2018; 水 谷 ほ か 2016; 竹 山 ほ か 2014)、葉の長さが極端に短く、葉の幅も減少すること で(水谷ほか 2016)、魚類の種数や個体数が著しく減少 するなど(Nakamura 2010)、群落構造の変化とそれに関 連した生物群集への影響が生じ始めている。ウミショウ ブの北限個体群とその藻場生態系の保全策を講じること と、それに資するための生態学的情報の集積が急務であ り、これまでに分布と生育環境(中瀬ほか 2015;竹山 ほか 2014)、種子や果実の分散(村上ほか 2014, 2015a, b)、 葉の成長とアオウミガメの摂餌による衰退(水谷ほか 2016)、西表島と石垣島の個体群の遺伝的多様性と連結 性(野原ほか 2018)、および障害物を用いたアオウミガ メの摂餌軽減手法の有効性(水谷ほか 2018)が報告さ れてきた。一方で、衰退や消失した群落の自然回復の見 込みを評価することや、将来的には人為的な再生技術を 検討することに必要な本海域におけるウミショウブの繁 殖に関する研究例はなかった。本種は、地下茎を伸ばす ことによる栄養繁殖(クローナル成長)と有性繁殖を行 うが、地理分布の中心である熱帯地域と異なり、環境の 季節変化が顕著な分布北限域では、これらの繁殖が可能 な季節が限られている可能性がある。 本研究では、分布北限域におけるウミショウブの有性 繁殖の特徴を明らかにすることを目的として、西表島北 西部沿岸の群落を対象に開花時期、開花量、および種子 放出までの期間と生残率について調査を行った。
材料と方法
調査地 西表島北西部に位置する祖納集落と干立集落の前に奥 行き 0.9 km の湾がある(図 1)。湾内の浅瀬の底質は砂 礫であり、与那田川河口を挟んで南西側(祖納側)と北 東側(干立側)に総面積約 10 ha のウミショウブを主と する群落が形成されている。2014 年と 2015 年現在、ア オウミガメによる食痕(葉が明確に短く、かつ先端が直 線的)(水谷ほか 2016;中西 2009)は、サンゴ礁との境 界付近にごく疎らにみられたものの、群落の中心部にま では至っておらず、比較的健全な状態であった。そこで、 当地を調査地とした。 2014 年(調査区 A-C)と 2015 年(調査区 D-F)に、4× 4 m のコドラートをそれぞれ 3 ヵ所ずつ設置した。各調 査区の高さは、気象庁西表島(24° 21’N, 123° 45’E)の temperature and sunshine duration were high and the daytime low-tide level was relatively low. The timing of flowering was synchronised with the spring tides associated with lower daytime tide levels during either full or new moons. After October, the tides were high and surface-pollination failed. The average period from flowering to mature fruit dehiscence was 84±6.5 days, and was shorter than in the tropics. The pollination success (proportion of female flowers remaining after 3 days) and fruit survivorship 2 months after flowering were 51.2% and 9.8%, respectively. The proportion of fruit that matured per female flower was 9.3%. The flowering season was shorter than in tropical regions and only flowers from the earlier part of the flowering season produced seeds. This suggests that sexual reproduction is lower at the northern limit than in the tropics.潮位表基準面から -15.5 から -18.5 cm であり、調査区間 の水深の差異は、2015 年に計測した限りでは 5 cm の範 囲内であった。 ウミショウブのシュート数は、各年 5 月に計数し、3 調査区合計で 2014 年は 2,540 shoots(45-61 shoots/m2) であり、2015 年が 2,651 shoots(44-68 shoots/m2)であ った。なお、群落内におけるウミショウブのシュート数 密度は、最も高い場所で 150-200 shoots/m2であるが(著 者ほか 未発表)、本調査では、個々のシュートの識別を 正確に行うために、シュート密度が比較的低い場所を選 定した。 開花と果実の観察 ウミショウブは雌雄異株である(図 2)。受粉の様式は、 雄性花水面媒と呼ばれ、大潮の日中の干潮時に放出され た雄花が水面を移動して、海面に出た雌花がそれら を取り込んで受粉する(図 2A-D)(Ackerman 2006;den Hartog 1970)。受粉しなかった雌花は開花後に脱落する が、受粉した雌花は、苞鞘の柄が螺旋状に巻き、シュー トの根元で果実(子房)が肥大成長し(図 2E)、成熟後 に不規則に裂開して種子を放出する(図 2F)(大場・宮 田 2007;田中 2015)。本調査では、まず、各月とも大潮 の最干潮日の 1 週間前に各調査区で雌雄の花序のあるシ ュートを数えて、その月の開花シュート数とした。雌花 序を有したシュートには、番号を記した耐水紙を巻いて 識別をした。受粉成功の判断基準として、3 日後に開花 したシュートを観察して、雌花の有無を記録した。さら に、60 日目以降に果実形成の有無を確認し、そして果 実が裂開(種子放出)するまでほぼ毎日観察した。これ らの調査を 2014 年 1 月から 12 月と 2015 年 5 月から 12 月に行った。 また、2013 年から 2017 年にかけて、特定の調査区は 設けてはいないものの、本群落内を大潮の干潮時に広範 囲に 1-2 時間程度泳ぐか、歩いて、雌雄の花序や開花(特 に雄花が水面を浮遊する様子)を観察し、開花の状況を 定性的に記録した。 環境要因 本研究では調査区において環境の直接的な観測は行っ ていないため、調査地から最も近くで観測された記録を 図 1.調査海域。黒丸 , 4×4 m の調査区(2014 年,A-C;2015 年,D-F);斜線,ウミショウブを優占 種とした海草藻場(竹山ほか 2014 より);破線,サンゴ礁縁。
Fig. 1. Study area. Solid circles, 4 × 4 m quadrats (2014, A–C; 2015, D–F); diagonal lines, seagrass meadow consisting predominantly of Enhalus acoroides (Takeyama et al. 2014); broken lines, edge of the coral reef.
用いた。水温は、調査地から約 9.5 km 離れた西表島網 取湾の東海大学沖縄地域研究センターによる記録を用い た( 水 谷・ 崎 原 2015, 2016)。 水 温 計(HOBO 社 製、 Water Temp pro V2)は、同施設前の水深約 2m の底層に 設置され、10 分間隔で記録された。日照時間は、気象 庁により調査地から約 3.6 km 離れたアメダス観測地点 の西表で得られた記録(http://www.jma.go.jp/jma/menu/ menureport.html 最終確認日 2018 年 1 月 8 日)を参照した。 潮位は、NAO モデル(Matsumoto et al. 2000)を用いて、 2014 年 1 月から 2015 年 12 月までの祖納における天文 潮位を時間解像度 10 分で算出した。開花から果実裂開 までの期間中に八重山諸島に接近・通過した台風と、そ の時のアメダス観測地点西表における日平均風速を参照 した(http://www.jma.go.jp/jma/menu/menureport.html 最終 確認日 2018 年 1 月 8 日)。 統計解析 開花時期と関連する環境要因を検討するため、2014 年から 2015 年の月ごとの開花の有無を目的変数とし、 発育に対する影響を想定して開花前 1 ヵ月間の平均水温 (開花しなかった場合は新月前 1 ヵ月)、およびその月の 日中の最低干潮位(開花した場合は開花日の最低干潮位) を説明変数として二項分布誤差による一般化線形モデル (GLM)での単変量ロジスティック回帰分析を行った。 環境要因どうしの相関が高いと予想されたため変数選択 は行わなかった。 本研究では、受粉成功の指標として開花 3 日後の雌花 の残存を用いた。受粉成功を決める環境要因を明らかに するため、開花 3 日後の雌花残存率を目的変数とし、開 花時の最干潮位、水温、風速を説明変数とした多変量の ロジスティック回帰を行った。 果実生存率を決める環境要因を明らかにするため、開 花 3 日後の残存雌花数に対する 2 ヵ月後の果実の残存率 を目的変数とし、開花から 2 ヵ月間の平均水温、および その間の台風の有無(日平均風速 25 m/s 以上の暴風を 伴ったもの)を説明変数とし、多変量のロジスティック 回帰を行った。 いずれの解析でも環境要因のうち日照時間は水温と強 い正の相関が認められたため(r = 0.84-0.85)、解析から 除 外 し た。 解 析 は 統 計 ソ フ ト R version 3.4.1(R Development Core Team 2017)を使用した。
結 果
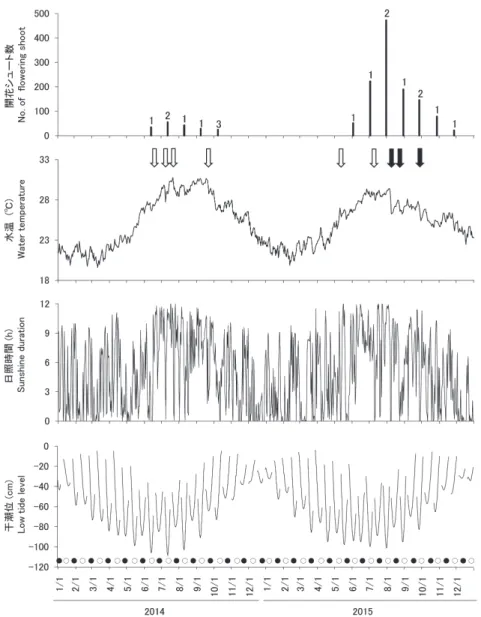
開花時期 2013 年から 2017 年までの定性的な観察を含めると、 ウミショウブの開花は 5 月から 11 月まで確認された(表 1, 2, 図 3)。また、年により新月と満月で日中の最干潮 位が異なるが、各年ともにより大きく引く大潮に同調し た開花周期がみられた。例えば、2014 年と 2015 年の開 花季節の大潮時の日中最干潮位は、満月の方が、その後 の新月よりも -20.8±9.05 cm(平均 ±SD、-31.1-1.1 cm) 低かった。 開花期間中は、一年のなかでも比較的水温が高く、日 照時間は長く、日中の干潮位が低い傾向があった。これ らのうち、開花時期に影響する要因を検討するため、水 図 2.ウミショウブ。A,雄の花序;B,雄花;C,雌の花序;D, 水面媒;E,開花 2 ヵ月後の果実;F,種子放出。Fig. 2. Enhalus acoroides. A, male inflorescence; B, female inflorescence; C, floating male flower; D, surface-pollinating; E, fruit 2 months after flowering; F, seed release.
温と高い相関がみられた日照時間を除いて単変量回帰し た結果、開花は水温と関連が大きく、日中の干潮位との 関連性は小さかった(表 3)。 2014 年と 2015 年の調査では、各大潮での開花日数は 1 日ないし最長 3 日間続くこともあった。また、年内に 複数回の開花が確認されたシュートは全くなかった。開 花数は 2014 年よりも 2015 年の方が多く、また 2015 年 は 7 月下旬に最も多かった。 受粉と果実の生残 2014 年と 2015 年に雌花が確認されたシュートは計 794 shoots であった。3 日後に雌花が残っていることを 受粉の指標とした場合、その割合は 2 年間の合計で 51.2%であり、また各開花周期によって変動が大きかっ た(表 2)。この比率に対し、受粉への影響が予想され る開花時の最干潮位、水温、および風速を用いた多変量 のロジスティック回帰を行った結果、最干潮位が低く、 水温が低く、風速が弱い場合に受粉の可能性は高まった 表 1.ウミショウブの開花と月周期。浮遊する雄花を観察して判断した開花量:○,多い;△,少ない;×,なし。NR,調査なし。 色つきは新月と満月で、日中の最低干潮がより低い月周期を示す。
Table 1. Flowering of Enhalus acoroides and lunar periodicity. Flowering intensity based on floating male flowers observed: ○ , large; △ , small; ×, none. NR, not researched. Coloured parts indicate lower tides in daytime between the new and full moons.
調査年 月周期
Jan-Apr May Jun Jul Aug Sep Oct Nov Dec Year Lunar periodicity
2013 新月 New NR NR × × △ △ NR NR NR 満月 Full NR NR 〇 〇 〇 〇 NR NR NR 2014 新月 New × × × × × × × × × 満月 Full × × 〇 〇 〇 〇 〇 × × 2015 新月 New NR × × × × × × × × 満月 Full NR × 〇 〇 〇 〇 〇 〇 × 2016 新月 New NR × 〇 〇 〇 〇 〇 〇 × 満月 Full NR △ △ △ △ NR NR NR × 2017 新月 New NR 〇 〇 〇 〇 〇 〇 × NR 満月 Full NR × △ △ △ △ △ × NR 表 2.ウミショウブの開花から果実の裂開までの概要。シュート数は 3 調査区(4×4 m/ 調査区)の合計。
Table 2. Summary from flowering to fruit dehiscence in Enhalus acoroides. Numbers of shoots are shown for three quadrats (4 × 4 m/quadrat). 調査年 Year シュート数 Number of shoot 開花日 Flowering date 開花数 Number of flowering shoot
3 日後の 雌花残存数 Number of female flowers remaining after three days 2 ヵ月後の 結果数 Number of fruit after two months
裂開果数 Number of dehiscing fruit
開花から果実裂開 までの日数 Days from flowering
to fruit dehiscing, mean ± SD (min-max) 果実裂開まで の台風数 Number of typhoons before fruit dehiscing 雄 Male 雌 Female 合計 Total 2014 2540 6/13 12 23 35 15 65.2% 11 47.8% 11 47.8% 77±1.6 (74-80) 0 7/12, 13 24 32 56 6 18.8% 6 18.8% 6 18.8% 75±2.2 (71-78) 0 8/10 13 30 43 14 46.7% 4 13.3% 4 13.3% 76±2.7 (72-79) 0 9/8 8 21 29 18 85.7% 2 9.5% 2 9.5% 101±4.2 (98-104) 0 10/7, 8, 9 8 17 25 0 0% 0 0% 0 0% - 0 小計 Subtotal 65 123 188 53 43.1% 23 18.7% 23 18.7% 79±7.3 (71-104) 2015 2651 6/2 15 37 52 25 67.6% 11 28.7% 10 27.0% 83±3.3 (78-88) 2 7/2 75 148 223 113 76.3% 7 4.7% 7 4.7% 84±3.0 (78-88) 3 7/30, 31 269 204 473 111 54.4% 26 12.7% 24 11.8% 87±3.9 (74-92) 3 8/29 72 118 190 77 65.3% 10 8.5% 9 7.6% 89±4.6 (84-99) 1 9/26, 27 55 91 146 29 31.9% 1 1.8% 0 0% - 1 10/27 23 56 79 0 0% 0 0% 0 0% - 0 11/26 5 17 22 0 0% 0 0% 0 0% - 0 小計 Subtotal 514 671 1185 355 52.9% 55 8.2% 50 7.5% 86±4.8 (74-99) 5191 合計 Total 579 794 1373 408 51.2% 78 9.8% 73 9.3% 84±6.5 (71-104)
(表 4)。また、開花時の干潮位が -80 cm 以下であれば結 実するが、それより高い水位では開花しても結実しなか った(図 4)。 開花から果実の裂開までの期間は、平均 84±6.5 日間 (71-104 日間)であった(表 2)。また、開花から果実裂 開までの着果シュートの残存率は 9.3%(開花 3 日後か らでは 17.8%)であった。2 ヵ月目から果実裂開までの 着果シュートの残存率は 93.6%であった。開花の月別に みると、6 月に開花したシュートでは 30-50%が果実裂 開に至ったのに対して、9 月下旬以降に開花したシュー 図 3.ウミショウブの開花シュート数、日平均水温、日照時間、および日中の最低干潮位の変化。大潮の 1 週間前に 花序を有した場合を開花とした。黒棒の上の数字はその大潮で開花がみられた日数を表した。黒棒は、その大潮 中に 3 調査区で開花した雄と雌の合計シュート数を示した。干潮位のグラフ上の白丸と黒丸は、それぞれ満月と 新月を示す。矢印は八重山諸島に接近した台風を表し、黒は日平均風速 25 m/s 以上(最大瞬間風速 40 m/s 以上) を意味する。
Fig. 3. Changes in the number of flowering shoots of Enhalus acoroides, daily average water temperature, sunshine duration, and lowest daytime tide level. Flowering was defined as having an inflorescence 1 week before the spring tide. Numbers above the black bars indicate the days of flowering observed during each tidal phase. Black bars show the total number of male and female flowering shoots in three quadrats (4 × 4 m/quadrat) during the tidal phase. White and black circles on the tide graph indicate the full and new moon, respectively. Arrows indicate a typhoon approaching Yaeyama Islands, and black indicates a daily average wind speed of 25 m/s and maximum instantaneous wind speed ≥ 40 m/s.
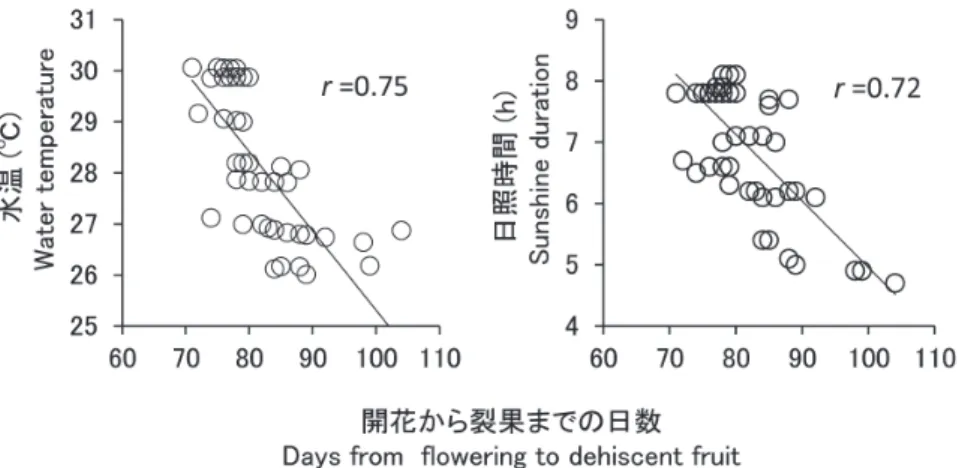
トでは果実裂開に至るものはほとんどなかった(図 5)。 開花 3 日目から 2 ヵ月目までの着果シュートの残存率 に影響する可能性がある要因として、2 ヵ月間の平均水 温と同期間中の台風の有無を説明変数とした回帰を行っ たところ、水温との関連がみられ、台風の有無との関連 はみられなかった(表 5)。 開花から果実の裂開までの期間について、平均水温お よび平均日照時間と回帰分析を行った結果、いずれの要 因とも負の相関がみられた(図 6)。
考 察
東南アジアやオーストラリアなど、ウミショウブの分 布中心域である熱帯海域では、開花はほぼ 1 年を通して 確認されている(Brounds and Heijs 1986;Erftemeijer 1993;Kenyon et al. 1997;Rattanchot et al. 2016;Rollon et al. 2003;Verheij and Erftemeijer 1993)。より高緯度に 位置し亜熱帯域に属する八重山諸島では、水温や日長の 季節変化が大きく、それらに制限される北限個体群特有 のフェノロジーがみられた。北限個体群の開花時期は 5 月から 11 月までに限られ、種子生産への寄与は開花期 の前半に限られる。これらのことから、ウミショウブの 北限個体群の有性繁殖による生産性は、熱帯と比べれば 低い可能性がある。 表 3.開花時期を決める要因。月ごとの開花の有無を目的変数、開花前 1 ヵ月間の平均 水温と日中の最低干潮位を説明変数とした単変量ロジスティック回帰分析の結果。 Table 3. Factors determining the flowering period. Results of univariate logistic regression analyses using presence/absence of flowering as the objective variable and the average water temperature 1 month before flowering and lowest daytime tide level on the flowering day as explanatory variables.説明変数 回帰係数 切片
P Explanatory variable Regression coeffcient Intercept
水温 Water temperature 1.3457 -34.3676 0.0198 最低干潮位 Lowest tide level -0.05833 -4.31864 0.0114
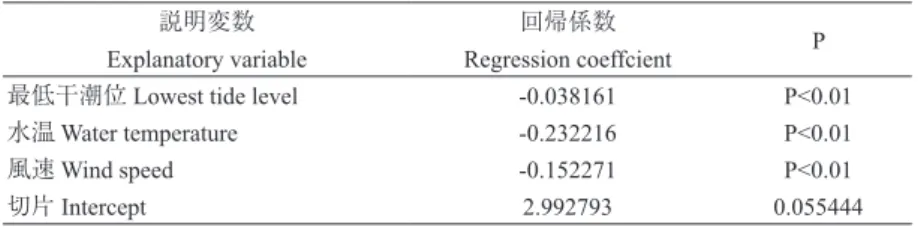
表 4.受粉成功を決める要因。受粉成功の指標として開花 3 日後の雌花の残存を用い た。開花 3 日後の雌花残存の割合を目的変数とし、開花時の日中の干潮位、水温、 風速を説明変数とした多変量ロジスティック回帰分析の結果。
Table 4. Factors determining successful pollination. A female flower remaining after 3 days was regarded as successful pollination. Results of the multivariate logistic regression analysis using the rate of female flowers remaining as the objective variable and the lowest tide level, water temperature, and wind speed at flowering as explanatory variables.
説明変数 回帰係数
P Explanatory variable Regression coeffcient
最低干潮位 Lowest tide level -0.038161 P<0.01 水温 Water temperature -0.232216 P<0.01 風速 Wind speed -0.152271 P<0.01 切片 Intercept 2.992793 0.055444
図 4.受粉の成功を指標する開花 3 日後の雌花残存率と開花日 の日中の最低干潮位との関係。
Fig. 4. The relationship between the proportion of female flowers remaining after 3 days and lowest tide on the flowering date.
開花フェノロジー
開花は、水温のほかに、濁りや水深に伴う光合成有効 放射の相違によって時間的・空間的に異なることや (McMillan 1982;Rollón et al. 2003)、開花量はシュート 密度に伴うキャノピー下での光の強さには関連しない (Rattanchot and Prathep 2011;Rattanchot et al. 2016)とい
った報告がある。 一方、分布の北限海域にある八重山諸島では、開花時 期は 5-9 月(当真 1999;横地 1985)、7-8 月(大場・宮 田 2007)、あるいは 9-1 月(Kuo et al. 2006)と異なる記 述がなされており、詳細は明らかにされていなかった。 本研究では、西表島において開花時期が 5 月から 11 月 までであることを確認した。また、開花は、満月と新月 の大潮のうち、日中の干潮位がより低い周期に同調した。 本種の受粉成功には、潮位が影響すると考えられており (Brouns and Heijs 1986)、雄性花水面媒を行う本種にと って、大潮周期での開花は受粉成功を高める繁殖戦略の 一つであると解釈される。 一方で、3 月や 4 月でも日中の干潮位は、5 月から 11 月と同様に低い年もあるが、ウミショウブの開花はみら れなかった。開花時期と水温や潮位の関係を解析したと ころ、水温がより強い関連を示した。また、日照時間は 水温と強い相関があった。 受粉と果実成熟のフェノロジー パプアニューギニアにおいて、受粉率(雌の開花シュ ート数に対して 1 ヵ月後に果実を有したシュート数の割 合)は、大潮の日中の干潮位が低い月では 65-78%で、 高い月では 33% に下がるといった記録がある(Brouns and Heijs 1986)。本調査地では受粉に関係する開花 3 日 後の残存率が、開花時の最干潮位と強い関連を示し(表 4)、また日中の干潮位が -80 cm よりも低い 9 月以前で は 57.9%(18.8-85.7%)で、日中の干潮位が -80 cm よ りも高い 10 月と 11 月では 0%であった。観察した限り では、雌花は最干潮時に水面まで達しておらず、この 時期の潮位では結実に至る可能性は低いと考えられる。 本調査地では、開花から果実裂開までの期間は、平均 84 日間(71-101 日間)であった。しかし、パプアニュ ーギニアでは、果実の成熟期間が 3-5 ヵ月、主に 4 ヵ月 で本海域よりも長かった(Brouns and Heijs 1986)。繁殖 図 5.雌の開花量と裂果量の季節変化。A,2014 年と 2015 年の
雌の年開花数に対する各月開花数の割合の平均;B,各開 花月の雌の開花数に対する果実裂開数の割合。
Fig. 5. The seasonal variation in the amount of female flowering and fruit dehiscence. A, average ratio of monthly number of female flowers to annual number of flowers in 2014 and 2015; B, ratio of the number of dehiscent fruit to the number of flowers in each month.
表 5.果実の生残率を決める要因。開花 3 日後の残存雌花を初期状態として 2 ヵ月後の 果実の残存割合を目的変数とし、開花から 2 ヵ月間の平均水温と台風(最大瞬間風 速 25 m/s 以上)の有無を説明変数とした多変量ロジスティック回帰分析の結果。 Table 5. Factors determining fruit survival. Results of the multivariate logistic regression
analysis using the proportion of fruit remaining 2 months after flowering to the female flowers remaining as the objective variable and the average water temperature and presence/absence of typhoons with daily wind speeds ≥ 25 m/s as explanatory variables.
説明変数 回帰係数
P Explanatory variable Regression coeffcient
水温 Water temperature 1.183 P<0.01 台風の有無 Presence/absence of Typhoon 0.1498 0.497 切片 Intercept -34.1804 P<0.01
成長が可能な季節が限られた北限の個体群では短い成熟 期間が適応的である可能性がある。 シュートあたり種子生産 熱帯域では、ひとつのシュートの開花回数が年に最大 3.4 回(Rollón et al. 2003)、果実あたりの種子数は、平 均 9 個(3-15 個)(Rollón et al. 2003)や多くが 7 個ない し 9 個(2-11 個)(Brouns and Heijs 1986)といった記録 がある。本調査ではシュートの開花回数は全て年 1 回で あった。また、西表島で調べられた果実あたりの種子数 は、2013 年 6-10 月の開花では平均 8.8 個(1-17 個)(村 上ほか 2014)であり、また 2014 年 6-9 月の開花では、 月平均 4.6 個(3-8 個)、9.5 個(5-13 個)、9.4 個(5-12 個)、 10.2 個(5-12 個)であった(村上ほか 2015b)。したが って、熱帯域と比較して、果実あたりの種子数は同等で あるが、ひとつのシュートによる年間の種子生産数は少 なく、これも北限個体群の特徴の一つといえる。 年変動 開花数は年によって変動した(表 2)。また、9 月以前 の残存率は、開花周期によって変動が大きかった。受粉 成功に影響する要因として潮位の他に、その時々の風が 挙げられており(Brouns and Heijs 1986)、本解析でもそ の関連性がみられた(表 4)。一方で、本解析では水温 との負の関連も認められたが、熱帯性のウミショウブに とって低温が受粉を促進することはしられておらず、他 の要因によるみかけの効果であるかもしれない。群落内 のウミショウブを含めた海草全体のキャノピー被度と結 実率には非線形的な関係があり、被度 50%程度で増加 することも報告されている(Vermaat et al. 2004)。ウミ ショウブの受粉成功には、その時々の潮汐、風況、群落 内の被度などが相互に関係しながら影響すると考えられ る。 2015 年 7 月から 8 月は、果実の成熟期に台風を経験し、 開花 3 日後から果実裂開までの生残率が 6.1-21.6%で顕 著に低く、台風後の観察では、果実が海岸に打ち上がっ ていることもあった。また、同様の海岸への果実漂着は、 11 月以降に北東季節風が卓越して海況が悪化した日の 後でも観察された。本研究では果実の生残を妨げる要因 として暴風を伴う台風の有無を検討したが、統計的な有 意差はみいだされなかった。このような年による有性繁 殖の違いは今後の研究課題であり、長期の継続調査が必 要である。果実の表面や花茎に魚類によると思われる食 痕が確認されることもあったが、この重要性や変動につ いても今後の課題である。
謝 辞
本調査は、東海大学沖縄地域研究センター・崎原健氏 をはじめ、同学海洋学部の学生諸氏に協力頂いた。統計 解析には統計数理研究所・山本誉士氏と水産大学校・南 條楠土氏に教示頂いた。英文は水谷麻希子氏に校閲して 頂いた。また、地域の方々からは各地のウミショウブ群 落の衰退や消失に関する情報を提供して頂いた。本稿の 図 6.開花から果実の裂開までの日数と水温および日照時間との関係。水温と日照時間は、 各シュートの開花から果実裂開までの期間の平均を示した。Fig. 6. The relationship between the days from flowering to fruit dehiscence and the water temperature or sunshine duration.
査読者ならびに編集者の方々には多大な教示と助言を賜 った。これらの方々に深謝致します。なお、本研究は 2013-2015 年度沖縄地域研究センター研究教育助成を受 けたほか、東海大学、国立研究開発法人防災科学技術研 究所、五洋建設株式会社との共同研究として実施された。
引用文献
Ackerman JD (2006) Sexual reproduction of seagrasses: Pollination in the marine context. In: Larkum AWD, Orth RJ, Duarte CM (eds.) Seagrasses: Biology, Ecology and Conservation. 89-109. Springer, Dordrecht
Brouns JJWM, Heijs FML (1986) Production and biomass of the seagrass Enhalus acoroides (L.f.) Royle and its epiphytes. Aquatic Botany, 25:21-45. https://doi. org/10.1016/0304-3770(86)90038-0
den Hartog C (1970) The Seagrass of the World. North Holland Publishing Company, Amsterdam
Green EP, Short FT (2003) World Atlas of Seagrasses. Prepared by the UNEP World Conservation Monitoring Centre. University of California Press, Berkeley
環境省那覇自然環境事務所, 東海大学沖縄地域研究セン ター (2016) 平成27年度西表島崎山湾・網取湾自然環 境保全地域におけるウミショウブの減少要因等調査業 務報告書. 環境省那覇自然環境事務所, 那覇 環境省那覇自然環境事務所, 東海大学沖縄地域研究セン ター (2017) 平成28年度西表島崎山湾・網取湾自然環 境保全地域におけるウミショウブの減少要因等調査業 務報告書. 環境省那覇自然環境事務所, 那覇 環境省那覇自然環境事務所, 東海大学沖縄地域研究セン ター (2018) 平成30年度西表島崎山湾・網取湾自然環 境保全地域におけるウミショウブの減少要因等対策検 討業務報告書. 環境省那覇自然環境事務所, 那覇 環境省自然環境局野生生物課希少種保全推進室 (編) (2015) レッドデータブック2014 −日本の絶滅のおそ れのある野生生物− 8 植物Ⅰ (維管束植物). ぎょうせ い, 東京
Kenyon RA, Conacher CA, Poiner IR (1997) Seasonal growth and reproduction of Enhalus acoroides (L.f.) Royle in a shallow bay in the western Gulf of Carpentaria, Australia. Journal of Marine and Freshwater Research, 48: 335-342. https://doi.org/10.1071/MF96106
Kuo J, Kanamoto Z, Iizumi H, Aioi K, Mukai H (2006) Seagrasses from Nansei Islands, Southern Japanese Archpelago: Species composition, distribution and biogeography. Marine Ecology, 27:290-298. https://doi. org/10.1111/j.1439-0485.2006.00098.x
Matsumoto K, Takanezawa T, Ooe M (2000) Ocean tide models developed by assimilating TOPEX/POSEIDON altimeter data into hydrodynamical model: A global model and a regional around Japan. Journal of Oceanography, 56:567-581. https://doi.org/10.1023/A:1011157212596
McMillan C (1982) Reproductive physiology of tropical seagrasses. Aquatic Botany, 14:245-258. https://doi. org/10.1016/0304-3770(82)90102-4 水谷 晃, 井上 太之, 井上 嵩裕, 竹中 康進, 河野 裕美 (2018) アオウミガメによるウミショウブの摂餌を軽減 する手法の有効性の評価. 東海大学沖縄地域研究セン ター所報 西表島研究, 2017:33-45 水谷 晃, 崎原 健 (2015) 東海大学沖縄地域研究センター における気象観測 (2014年) −網取湾の風向・風速・ 気温・湿度・雨量・海水温, ならびに浦内川河口域の 水温・塩分−. 東海大学沖縄地域研究センター所報 西 表島研究, 2014:68-79 水谷 晃, 崎原 健 (2016) 東海大学沖縄地域研究センター における気象観測 (2015年) −網取湾の風向・風速・ 気温・湿度・雨量・海水温ならびに台風13号, 15号, 21 号の記録−. 東海大学沖縄地域研究センター所報 西表 島研究, 2015: 74-84 水谷 晃, 崎原 健, 川田 菜摘, 河野 裕美 (2016) アオウミ ガメの摂餌がウミショウブの株に与える影響 −葉の 切り戻し実験と食痕藻場の経過観察による検証−. 東 海大学沖縄地域研究センター所報 西表島研究, 2015: 42-51 村上 智一, 河野 裕美, 岩崎 彩加, 蔵持 登士江, 下川 信 也, 木村 賢史 (2014) 生態調査に基づく西表島網取湾 におけるウミショウブ種子の分散力の数値解析. 土木 学会論文集B3 (海洋開発), 70:I_1074-I_1079. https://doi. org/10.2208/jscejoe.70.I_1074 村上 智一, 河野 裕美, 玉本 満, 水谷 晃, 下川 信也 (2015a) 西表島北西部のウミショウブ群落における果実分 散の可能性に関する数値解析. 土木学会論文集B2 (海岸工学), 71:I_1351-I_1356. https://doi.org/10.2208/ kaigan.71.I_1351 村上 智一, 河野 裕美, 玉本 満, 水谷 晃, 下川 信也 (2015b) 数値シミュレ–ションを用いた西表島北西部のウミシ ョウブ群落における種子の分散力評価. 土木学会論文 集B3 (海洋開発), 70:I_951-I_956. https://doi.org/10.2208/ jscejoe.71.I_951
Nakajima Y, Matsuki Y, Lian C, Fortes MD, Uy WH, Campos WL, Nakaoka M, Nadaoka K (2014) The Kuroshio Current influences genetic diversity and population genetic structure of a tropical seagrass, Enhalus acoroides. Molecular Ecology, 23:6029-6044. https://doi.org/10.1111/ mec.12996
中西 喜栄 (2009) 石垣島の海草藻場で確認されたアオウ ミガメの食み跡. 沖縄生物学会誌, 47:19-23
Nakamura Y (2010) Patterns in fish response to seagrass bed loss at the southern Ryukyu Islands, Japan. Marine Biology, 157:2397-2406
Nakamura Y, Horinouchi M, Nakai T, Sano M (2003) Food habits of fishes in a seagrass bed on a fringing coral reef at Iriomote Island, southern Japan. Ichthyological Research, 50:15-22. https://doi.org/10.1007/s102280300002
risk for juvenile fishes in a seagrass bed compared with an adjacent coral area? Bulletin of Marine Science, 74:477-482
Nakamura Y, Sano M (2005) Comparison of invertebrate abundance in a seagrass bed and adjacent coral and sand areas at Amitori Bay, Iriomote Island, Japan. Fisheries Science, 71:543-550. https://doi.org/10.1111/j.1444-2906.2005.00998.x 中瀬 浩太, 村上 智一, 鵜飼 亮行, 水谷 晃, 下川 信也, 河野 裕美 (2015) 外力再現計算を用いた西表島北西部海域 におけるウミショウブ群落分布域の物理条件解析. 土 木学会論文集B3 (海洋開発), 71:I_957-I_962. https://doi. org/10.2208/jscejoe.71.I_957 野原 健司, 水谷 晃, 丸 琢哉, 鈴木 舞子, 武島 弘彦, 河野 裕 美 (2018) 西表島崎山湾および石垣島野底沿岸におけ るウミショウブ群落の遺伝的多様性と連結性. 東海大 学沖縄地域研究センター所報 西表島研究, 2017:23-32 大場 達之, 宮田 昌彦 (2007) 日本海草図譜. 北海道大学出 版会, 札幌 沖縄県文化環境部自然保護課 (編) (2018) 改定・沖縄県 の絶滅のおそれのある野生生物 第3版 (菌類編・植物 編) −レッドデータおきなわ−. 沖縄県文化環境部自 然保護課, 那覇
Rattanchot E, Prathep A (2011) Temporal variation in growth and reproduction of Enhalus acoroides (L.f.) Royle in a monospecific meadow in Haad Chao Mai National Park, Trang Province, Thailand. Botanica Marina, 54:201-207. https://doi.org/10.1515/bot.2011.018
Rattanchot E, Short FT, Prathep A (2016) Enhalus acoroides responses to experimental shoot density reductions in Haad Chao Mai National Park, Trang Province, Thailand. Marine Ecology, 37:411-418
R Development Core Team (2017) R: A Language and Environment for Statistical Computing. R Foundation for
Statistical Computing, Vienna
Rollón RN, Steveninck EDR, Vierssen W (2003) Spatio-temporal variation in sexual reproduction of the tropical seagrass Enhalus acoroides (L.f.) Royle in Cape Bolinao, NW Philippines. Aquatic Botany, 76:339-354. https://doi. org/10.1016/S0304-3770(03)00070-6
Short FT, Waycott M (2010) Enhalus acoroides. The IUCN Red List of Threatened Species 2010:e.T173331A6992567. http://dx.doi.org/10.2305/IUCN.UK.2010-3.RLTS. T173331A6992567.en 竹山 佳奈, 河野 裕美, 蔵持 登士江, 岩崎 彩加, 村上 智 一, 木村 賢史, 鵜飼 亮行, 中瀬 浩太 (2014) 西表島 北西部における熱帯性海草ウミショウブの広域分 布及び生育環境条件について. 土木学会論文集B3 (海洋開発), 70:I_1068-I_1073. https://doi.org/10.2208/ jscejoe.70.I_1068 田中 法生 (2015) トチカガミ科. (大橋 広好, 門田 裕一, 木 原 浩, 邑田 仁, 米倉 浩司 編) 改定新版 日本の野生植物 1 ソテツ科-カヤツリグサ科, 120-121, 平凡社, 東京 当真 武 (1999) 琉球列島の海草 −Ⅱ. 種類と分布. 沖縄生 物学会誌, 37:75-92 土屋 正弘 (1981) 沖縄西表島網取湾におけるアオリイカ の産卵について. 東海大学海洋研究所資料, 3:53-75 Verheij E, Erftemeijer PLA (1993) Distribution of seagrasses
and associated macroalgae in South Sulawesi, Indonesia. Blumea-Biodiversity, Evolution and Biogeography of Plants, 38:45-64
Vermaat JE, Rollón RN, Lacap CDA, Billot C, Alberto F, Nacorda HME, Wiegman F, Terrados J (2004) Meadow fragmentation and reproductive output of the SE Asian seagrass Enhalus acoroides. Journal of Sea Research, 52:321-328. https://doi.org/10.1016/j.seares.2004.04.002 横地 洋之 (1985) 海に咲く花 −西表島のウミショウブに