PLA
2induces several pharmacological ef- fects including presynaptic neurotoxicity, myotoxicity and cardiotoxicity as well as anticoagulation, hemolysis, hemorrhage, edema-induction and platelet-aggregation inhibition [3‒5]. Habu ( T. flavoviridis ) in- habiting in the southwestern islands of Ja- pan contains several PLA
2s and the related INTRODUCTION
Phospholipase A
2(EC 3.1.1.4) is the major component of most snake venoms [1], and the detailed insight into the structure and mechanism of these enzymes is available [2]. It catalyzes the hydrolysis of phospho- lipids at its sn -2 position. Snake venom
Structural Analysis of Two Isoforms of Subunit B in Phospholipase A
2Inhibitor PLI γ from the Serum of Habu, Trimeresurus flavoviridis
Narumi AOKI, Masanobu DESHIMARU, and Shigeyuki TERADA1)
(Received November
30
,2007
)Abstract
Two distinct types of phospholipase A2 inhibitors (PLIs), PLIα and PLIγ, are present in the serum of Habu (Trimeresurus flavoviridis). It is expected that, like other PLIγs, T. flavoviridis PLIγ may contain two homologous subunits, A- and B-chains (also called α- and β-chains).
Five proteins, termed PLI-I to PLI-V, were identified as the constituting subunits of two types of PLIs. Based on the primary structures and the molecular sizes, PLI-I was assigned to be the subunit A of T. flavoviridis PLIγ. In order to prove that PLI-II and PLI-III are the subunit B, we have purified these proteins from the same serum, and determined the complete amino acid sequences by peptide analysis. The cDNA encoding PLI-II was also cloned from a cDNA li- brary of the T. flavoviridis liver. The respective nucleotide sequence encoded
19
-residue signal sequences, followed by181
-residue proteins. Both PLI-II and PLI-III were glycosylated at Asn14, and showed a high homology to several B-chains of PLIγ identified in other snakes. Gel filtra- tion analysis showed that all the PLIs exist in the T. flavoviridis serum as the high molecular forms of more than60
kDa, and PLI-II and PLI-III also associate to form the corresponding ho- modimers.Key words: amino acid sequence, cDNA cloning, phospholipase A2 inhibitor, snake serum, Trimeresurus flavoviridis
1) Department of Chemistry, Faculty of Science, Fukuoka University, 8-19-1 Nanakuma, Jonan-ku, Fukuoka 814-0180, Japan
Abbreviations: BPI, basic protein I; BPII, basic protein II; MALDI, matrix-assisted laser desorption ionization; PLA2, phospholipase A2; PLI, phospholipase A2 inhibitor; PCR, polymerase chain reaction; RACE, rapid amplification of cDNA ends; SDS-PAGE, SDS-polyacrylamide gel electrophoresis; TFA, trifluoroacetic acid; TOF, time-of-flight; UTR, untranslated region.
proteins such as acidic [Asp
49]-PLA
2s (PLA2 and PLA-N), a basic [Asp
49]-PLA
2(PLA-B), and basic proteins I and II (BPI and BPII) [5
‒8].
Venomous snakes have PLA
2inhibitors (PLIs), proteins capable of inhibiting snake venom PLA
2s, in their blood sera [9]. The role of these proteins is postulated to pro- tect themselves from the leakage of their own venomous PLA
2s into their circulatory system. Many PLIs have been isolated from various snake sera and their primary structures have been determined [10‒
14]. PLIs can be divided into three groups (PLI
α, PLI
β, and PLI
γ) based on their structural characteristics. PLI
αs are 75‒
100-kDa glycoproteins composed of three or four subunits and have the carbohydrate- recognition domain of Ca
2+-dependent (C- type) lectins [12,16]. PLI
βis a 160-kDa glycoprotein composed of three identical 50 kDa subunits with leucine-rich repeats [17].
PLI
γs are oligomeric proteins consisting of glycosylated subunit A and non-glyco- sylated subunit B [12‒14,18]. The subunit B of Elaphe quadrivirgata PLI
γ, however, contains a suger chain at residue 12 [19].
The primary structure of both subunits has dual three-finger motifs and is related to the Ly-6 superfamily [20].
Five proteins named PLI-I to PLI-V in T. flavoviridis serum have already been identified by affinity chromatography [21].
PLI-I corresponds to a subunit A of PLI
γ, and the cDNA has already been cloned [21]. PLI-IV and PLI-V are the member of
α
type PLIs, and identical to PLI-A and PLI-B [10], respectively. The structures of PLI-II and PLI-III have not yet known. In the present study, we have isolated two PLA
2inhibitors, PLI-II and PLI-III, from T.
flavoviridis serum, and determined their complete amino acid sequences by peptide sequencing as well as by cDNA cloning of
PLI-II. We also discuss the subunit struc- ture of T. flavoviridis PLI
γ.
MATERIALS AND METHODS Materials : The blood of T. flavoviridis in the Amami Oshima islands was collected by decapitation. The serum was separated by centrifugation and stored at
−20
℃. Ethanol precipitation was carried out as described [22]. All other reagents were purchased from Wako Pure Chem. (Osaka).
Electrophoresis : SDS‒PAGE was carried out on 12.5% gels by Laemmli
's method [23], or on 10% polyacrylamide gels as described by Schagger and von Jagow [24]. Bovine serum albumin (67,000), ovalbumin (46,000), carbonic anhydrase (30,000), and chymo- trypsinogen (24,000) were used as molecu- lar weight markers. Prestained XL-ladder marker kit (Apro science) was also used.
After running the gels under a constant current, they were stained with 0.1% Coo- massie brilliant blue R-250 and destained with 10% acetic acid.
Mass spectrometric analysis : Mass spec- trum was measured on a Voyager DE-STR matrix-assisted laser desorption ionization time-of-flight (MALDI‒TOF) mass spec- trometer (PerSeptive Biosystems). Sample was dissolved in 0.1% trifluoroacetic acid
‒50% acetonitrile containing
α-cyano-4- hydroxycinnamic acid (10 mg/ml) as the matrix, and 2-
μl aliquots were analyzed.
Spectrum was calibrated by the molecular mass of apomyoglobin.
Reverse-phase HPLC : Preparative re- verse-phase HPLC was performed on a
μ
Bondashere 5
μ-C8-300Å column (1.9
×15 cm, Waters), and the proteins were eluted with a linear gradient of acetonitrile in 0.1%
trifluoroacetic acid (TFA) at a flow rate of 5 ml/min.
Protein sequencing : Protein was reduced
and S -pyridylethylated (PE) according to Friedman et al . [25] The PE-protein (500
μ
g each) was digested at 37ºC with bo- vine trypsin (E/S
=1:50) for 24 h in 20 mM Tris‒HCl (pH 9.0), Lys-C (E/S
=1:100) for 5 h in 2 M urea-20 mM Tris‒HCl (pH 9.0),
α
-chymotrypsin (E/S
=1:50) for 16 h in 1 M urea‒50 mM NH
4HCO
3, Asp-N (E/S
=1:100) for 16 h in 50 mM Tris‒HCl (pH 8.0), V8 protease (E/S
=1:30) for 16 h, or cleaved at room temperature by CNBr (100 equiva- lents) for 21 h in 70% HCOOH. The digests were lyophilized, dissolved in 0.1% TFA, and fractionated by reverse-phase HPLC on a YMC-Pack ODS column in 0.1% TFA with an appropriate gradient of acetonitrile.
The amino acid sequences of proteins were determined by an automatic protein se- quencing system PPSQ 21 (Shimadzu).
Analytical gel filtration : A sample was dis- solved in 0.2 M NaCl-50 mM phosphate buf- fer (pH 7.0), and put on a TSKgel G3000SW column (0.75
×30 cm, Tosoh). Elution was carried out by the same buffer at a flow rate of 1.0 ml/min. Elution was monitored at 280 nm and the peak area was deter- mined by 807-IT integrator (Jasco). Mo- lecular weight was calibrated by the reten- tion times of bovine serum albumin (67,000), ovalbumin (46,000), and soybean trypsin inhibitor (21,500).
Synthesis of partial fragment of PLI cDNA . Total RNA was extracted from 0.5 g of T. flavoviridis liver by acid guanidinium- phenol-chloroform (AGPC) method, and reverse transcribed to synthesize cDNA first strands using adaptor-linked Oligo(dT) primer (5
'-GGCCACGCGTCGACTAGTAC- (dT)
17-3
'). cDNA obtained was used as the template for 3
'-RACE (rapid amplifica- tion of cDNA ends) reaction. Synthetic oligonucleotide, PLI-II_NS (5
'-CCNGGN- GARACNTGYAAYGGNACNATGATG-3
') and 3
'-Adp (5
'-GGCCACGCGTCGACTAG-
TAC-3
'), were used for PCR amplification.
PLI-II_NS primer was designed upon N-terminal amino acid sequence of PLI- II, and 3
'-Adp corresponded to the adaptor sequence within adaptor-linked oligo(dT) primer. Amplification product was once subcloned into plasmid vector, and its nu- cleotide sequence was determined. As the result, the sequence was confirmed to be the cDNA partial fragment for PLI-II. PLI- II cDNA fragment was radiolabeled with [
α-
32P]-dCTP (3000 Ci/mmol) using random primer DNA labeling kit (Takara Bio) and used for hybridization screening of cDNA library.
Construction of T. flavoviridis liver cDNA library: T. flavoviridis liver cDNA library was constructed using Creator SMART cDNA Library Construction kit (BD Biosci- ences) according to manufacturers
'instruc- tion. Briefly, cDNA first strand was syn- thesized using 1
μg of total RNA, followed by five cycles of PCR for non-specific en- richment of full-length cDNAs. The cDNA fragments were then ligated to pDNR-LIB vector. When the plasmid clones were used to transform E. coli JM109, resulting library contained 2.0
×10
6independent clones.
Cloning and sequence determination of
cDNA encoding PLI-II : Clones (9.6
×10
4)
from unamplified cDNA library were plated
on LB agar plates and bacterial colonies
were transferred onto Hybond-NX mem-
branes (GE Healthcare Bio-Science) and
fixed by UV irradiation. The resulted
replica membranes were prehybridized in
Church
ʼs hybridization solution at 65
℃for 30 min and then hybridized with ra-
diolabeled PLI-II cDNA overnight at 50
℃in Church
ʼs hybridization solution. Mem-
branes were finally washed twice for 15
min at 50
℃with 1
×SSC, 0.1% SDS and
hybridization signals were visualized using
BioImage Analyzer (Fuji Film). Accord- ingly, 14 bacterial colonies were isolated, cultured and their plasmids were purified by standard Alkali-SDS method. Nucleo- tide sequences of cDNA inserts were de- termined using ABI PRISM 377 DNA Se- quencing System (Applied Biosystems).
RESULTS
Purification of phospholipase A
2inhibitors : Ethanol fraction E
2.0of the T. flavoviridis serum was prepared as described previ- ously [22], and subjected to reverse-phase HPLC using a C8 column. Among several peaks observed in Fig. 1A, five peaks (I
−V) contained the PLI-related proteins as ana- lyzed by SDS-PAGE and the N-terminal sequencing. Since the protein in peak III was not homogeneous on SDS-PAGE (Fig.
1B), it was further purified by gel filtra- tion using a Sephacryl S-200HR column (data not shown). From the N-terminal sequence analysis, peaks I, IV, and V were assigned to be PLI-I [21], PLI-A, and PLI-B [10], respectively. We assumed that peaks II and III corresponded to PLI-II and PLI-III because of their molecular mass as well as the relative retention time on reverse-phase HPLC [21]. The apparent molecular mass- es of the purified PLI-I, PLI-II, and PLI-III were estimated to be 24, 24, and 25 kDa by SDS-PAGE, respectively (Fig. 1C). The exact masses were determined by MALDI
‒TOF mass spectrometry to be 22,653.9, 22,339.0, and 22,491.0 for PLI-I, PLI-II, and PLI-III, respectively.
Sequence analysis of PLI-II and PLI-III : The N-terminal sequences of S -pyridyl- ethylated PLI-II and PLI-III were directly determined and shown in Fig. 2. Both pro- teins had a common sequence of XXCXXC, where X is any amino acid residue other than Cys. This agreed with the structural
feature of N-terminals in all the PLI
γs.
The complete amino acid sequences of two inhibitors were determined on the basis of the results of sequence analysis of peptide fragments generated by several enzymatic and chemical digestions. The sequence of PLI-II consisting of 181 amino acids was determined by the combination of BrCN- cleavage and tryptic digestion (Fig. 2A). A residue 14 could not be determined by Ed- man degradation, suggesting that it might be the Asn residue having a carbohydrate chain.
The entire primary structure of PLI-III
Fig. 1. (A) Purification of PLA2 inhibi-tors by reverse-phase HPLC on a μBondashere
5
μ-C8
-300
Å column (1
.9
×
15
cm). (B) SDS-PAGE analysis of proteins in peak I‒V in Fig.1
A.12
.5
% gel was used. M, marker proteins. (C) SDS-PAGE analysis of purified pro- teins on a10
% gel. M, prestained XL- ladder marker; I, PLI-I; II, PLI-II; III, PLI-III.was also determined similarly by several enzymatic digestions using Lys-C, chymo- trypsin, and V8 protease. The arrangement of the fragments and the complete amino acid sequence are summarized in Fig. 2B.
PLI-III was a very similar protein to PLI- II having 181 residues. It may also have a sugar chain at residue 14.
Cloning of cDNA encoding PLI-II: T. fla- voviridis liver cDNA was synthesized using an adaptor-linked oligo(dT) primer. Partial cDNA fragment for PLI-II was then ampli- fied by PCR using the oligonucleotide PLI- IIa_NS that was originally designed based on N-terminal amino acid sequence of PLI- II (Fig. 2). As a result, a DNA fragment of the expected size (approx. 0.7 kb) was ob- tained. The nucleotide sequence was con- firmed to be the partial fragment of cDNA in accord with the amino acid sequences of PLI-II. The cDNA library was screened with PLI-II DNA fragment as a probe. Six positive clones were obtained from 9.6
×10
4plaques by washing in high stringency (0.2
×
SSC containing 0.1% SDS at 65
℃). Fig.
3 shows the nucleotide sequence of full- length cDNA for PLI-II. It consisted of 920 bp, including a 118-bp 5
ʼ-untranslated re- gion (UTR), 603-bp open reading frame en- coding 200 amino acids including a poten- tial signal peptide of 19 amino acids, and 202-bp 3
ʼ-UTR. The nucleotide sequence in the mature protein-coding region of PLI-II showed 53.9% identity to PLI-I [21]. Amino acid sequence deduced from the nucleotide sequence of PLI-II cDNA was agreed well to the protein sequence as cited in Fig. 2.
Comparison of amino acid sequences of PLIs : The amino acid sequences of T. fla- voviridis PLI-II and PLI-III were compared with those of several subunit proteins that constitute PLI
γs from the Elapidae, Hy- drophidae, Boidae, and Colubridae families (Fig. 4). Sequences of subunits A and B are separately shown and the conserved residues are also marked separately. Nev-
Fig. 2. Sequence determination of T. flavoviridis phospholipase A2 inhibitors. Sugar-attached Asn residues are marked by closed circles. Sequences determined directly from the intact pro- teins are underlined.
ertheless, most of the Cys residues are located at the same positions in both sub- units except Cys
105and Cys
113of subunit B.
The T. flavoviridis PLI-I has the common features of PLI
γ-subunit A: the presence of highly conserved 16 cysteine residues and N-linked carbohydrate site at Asn
157(the position 158 in the sequence (1) in Fig. 4A).
It showed the highest similarity to Agkis- trodon blomhoffii siniticus PLI
γ-subunit A (87% identity; the sequence (2) in Fig. 4A) [26].
On the other hand, the T. flavoviridis PLI-II and PLI-III were highly homologous (more than 53% identities) to other subunits
B of PLI
γs (Fig. 4B). In addition, all the cysteine residues were invariable. A sugar chain is attached at residues 14 of T. flavo- viridis PLI
γ-B (PLI-II and PLI-III) as well as at residue 12 of E. quadrivirgata PLI
γ-B [19]. Although G. blomhoff ii siniticus PLI
γ-B has a putative glycosylation site at residue 14, the protein was not glycosyl- ated [17]. Other proteins have no sugar chain.
Oligomer formation of PLIs : Analytical gel filtration using a TSKgel G3000SW column showed that PLI-I, PLI-II, and PLI-III were eluted at 9.07, 8.84, and 9.13 min which cor- responded to the molecular masses of 25,
Fig. 3. Nucleotide sequence of PLI-II. Deduced amino acid sequence is cited below the nucleotide sequence. Start and stop codons are boxed. Polyadenylation signal is underlined.
Fig. 4. Comparison of the amino acid sequences of γ-type phospholipase A2 inhibitors. (A) Sub- units A, (1) [accession No. BAA24503] T. flavoviridis PLI-I (subunit A), (2) [BAA86970] Gloy- dius blomhoffii siniticus subunit A, (3) [AAF23778] Notechis ater α-subunit 1, (4) [AAF23779] Notechis ater α-subunit 2, (5) [AAF23780] Notechis ater α-subunit 3, (6) [CAB56616] Notechis scutatus α-subunit ii, (7) [CAB56617] Notechis scutatus α-subunit iii, (8) [AAF23781] Oxyura- nus scutellatus α-subunit 1, (9) [AAF23782] Oxyuranus scutellatus α-subunit 2, (10) [AAF23783] Pseudonaja textilis α-subunit, (11) [AAF23785] Oxyuranus microlepidotus α-subunit 2, (12) [BAA83078] Elaphe quadrivirgata subunit A, (13) [AAB32582] Naja kaouthia 31 kDa sub- unit, (14) [AAA19162] Crotalus durissus terrificus CNF, (15) [AAR04437] Lachesis muta muta α-subunit 1, and (16) [AAR04438] Lachesis muta muta α-subunit 2. (B) Subunits B, (17) T.
flavoviridis subunit B1 (PLI-II), (18) T. flavoviridis subunit B2 (PLI-III), (19) [BAA86971] Gloydius blomhoffii siniticus subunit B, (20) [AAF21051] Pseudonaja textilis β-subunit 2, (21) [AAF21050] Pseudonaja textilis β-subunit 1, (22) [AAF21047] Oxyuranus scutellatus subunit B, (23) [AAF21046] Notechis ater subunit B, (24) [BAA83079] Elaphe quadrivirgata subunit B, (25) [AAB32583] Naja kaouthia 25 kDa subunit. Cysteine residues are indicated in white-on- black type. Conserved residues in each of subunit A and B are marked by asterisks. Carbo- hydrate-attatched Asn residues are underlined.
30, and 24 kDa, respectively, and also gave an additional peak corresponding to the molecular mass of 100, 56, or 50 kDa, re- spectively (data not shown). These results indicate that PLI-I is likely to exist as an oligomer, and PLI-II and PLI-III prefer to form the homodimers.
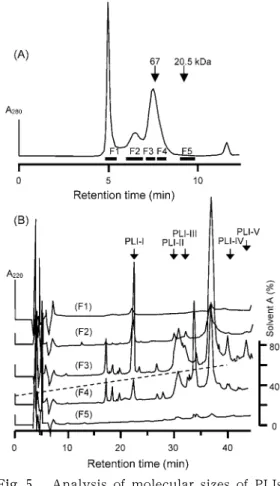
When T. flavoviridis serum was analyzed by gel filtration under the same conditions, four protein peaks were eluted at 5.00, 6.64, 7.54, and 11.68 min (Fig. 5A). The last peak contained no protein. To examine the presence of PLIs in these peaks on gel filtration, effluents from the column were divided into five fractions (F1 to F5) as shown in Fig. 5A, and the fractions were subjected directly to an analytical reverse- phase HPLC. As shown in Fig. 5B, no monomeric PLI was found in fraction F5.
Most of the PLIs were detected in fractions F3, and a small amount in F2 and F4 as well. The major components of T. flavo- viridis serum proteins in these fractions are summarized in Table 1. The apparent mo- lecular-mass range of F3 was a range from 65 to 130 kDa. This result clearly indicates that all of the PLIs are not monomeric but exist in the high-molecular-mass form in the serum as previously reported [9].
DISCUSSION
Two groups of PLIs have been identified in the serum of T. flavoviridis , PLI
αand PLI
γ[21]. PLI
γhave the two repeats of a unit termed the three-finger motif [20], which has also been found in urokinase- type plasminogen activator receptor [27], Ly-6 [28], CD59 [29] and neurotoxins [30].
In the present study, we have purified five proteins (PLI-I to PLI-V) from the serum of T. flavoviridis . Though the fraction III con- tained two proteins (Fig. 1B), a pure PLI-III was obtained after a contaminating 47-kDa
protein was removed by gel filtration. This 47-kDa protein was identified to be a novel HSF-like protein (unpublished data). Ana- lytical gel filtration showed that the resul- tant each of PLI-I, PLI-II, and PLI-III gave an additional peak in the molecular mass range from 50 to 100 kDa being attributable to its dimer or oligomer.
Sequence analysis showed that PLI- IV and PLI-V corresponded to PLI-A and PLI-B [10], respectively, and are known to
Fig. 5. Analysis of molecular sizes of PLIs.
(A) Analytical gel filtration of T. fla- voviridis serum on a TSKgel G
3000
SW column (0
.75
×30
cm) in0
.2
M NaCl‒50
mM phosphate buffer (pH7
.0
). (B) Reverse-phase HPLC of gel filtration fractions on a SepaxBio-C8
column (0
.46
×25
cm). Arrows show the reten- tion times of several PLIs.be the subunits of
α-type PLIs. Other pro- teins, PLI-I to PLI-III, were assigned to the subunits of PLI
γ. According to Nobuhisa et al. [21], PLI-I is a major component of in- hibitory proteins against three basic PLA
2isozymes in T. flavoviridis venom, that is, a basic PLA-B, BPI and BPII [7,8]. PLI
αis known to bind mainly to PLA2 (a major [Asp
49] PLA
2in the venom) [6]. Although PLI-II and PLI-III are thought to be the components of hetrooligomeric PLI
γ[21], their structures were not yet elucidated.
Therefore, we have determined their com- plete amino acid sequences by peptide analysis as well as the cDNA cloning.
As the results, PLI-II and PLI-III have been proved to be the isoforms of T. flavo- viridis PLI
γ-B since they showed a high homology to the subunits B from several other snakes. However, we have no evi- dence that PLI-I and PLI-II/III can associate to form a high molecular-mass heterooligo- mer in the blood plasma of T. flavoviridis . Moreover, as far as we know, the organiza- tion of T. flavoviridis PLI
γhas not been reported. Though the molar ratio of 25- to 20-kDa subunits was reported to approxi- mately 2 in A. blomhoffii siniticus PLI
γ[17], the contents of two subunits in the T. flavoviridis serum are greatly different and seem unexplainable by the formation of 2:1 heterooligomer. Considering that
three subunits of T. flavoviridis PLI
γwere detected in a wide range of the serum frac- tions (F2‒F4 in Fig. 5) on the analytical gel filtration, most of PLI
γmay be composed of PLI-I alone and a part of PLI-I may exist as the heterooligomer with the subunits B.
When various amounts of the purified PLI-II or PLI-III were mixed with a fixed amount of PLI-I and the mixtures were then analyzed by the analytical gel filtra- tion, any new peak corresponding to the complex could not be observed (data not shown). Because our PLIs were purified by reverse-phase HPLC under the acidic conditions, they could be significantly damaged. When PLI-I was purified under the mild conditions, it gave a single peak at approximately 120 kDa and no peak of 25 kDa was detected (data not shown).
Some snakes have the isoforms of either subunit A (
α-chain), B
(β-chain), or both (Fig. 4). For example, Oxyuranus scutel- latus, Notechis ater , and N. scutatus have several isoforms of subunit A, and O. mi- crolepidotus, Pseudonaja textilis , and T. fla- voviridis possess the isoforms of subunit B [32]. The isoforms are found for both sub- units of O. microlepidotus PLI
γ(Sekuloski et al., unpublished data). This enables to expand the repertoires of the specificity by the combination of many isoforms, and seems to the results of diversification of
Table 1. Characterization of fractions of T. flavoviridis serum on an analytical gel filtration.Fraction Molecular size
(kDa)a) Componentsb)
F
1
>500
High molecular mass proteins F2 130
‒340
GlobulinsF
3 65
‒130
Serum albumin, PLI-I, PLI-II, PLI-III, PLI-A, PLI-B, HSFF
4 30
‒65
serotriflin, SSP-1
to SSP-5
F5 18
‒30
Low molecular mass compoundsa) The column was calibrated by thyroglobin (
669
kDa), apoferritin (443
kDa), β-amylase (200
kDa), bovine serum albumin (67
kDa), ovalbumin (46
kDa), soybean Kunitz trypsin inhibitor (20
.5
kDa), and SSP-1
(1
.0
kDa).b) HSF, habu serum factor [
22
]: SSP, small serum protein.PLI
γgenes to adapt the rapid evolution of the target enzyme, PLA
2[33].
Recently, the nucleotide sequence of the cDNA encoding T. flavoviridis PLI- II was uploaded by So et al. [34] on the DDBJ sequence data bank: accession No.
BAF75726. Three nucleotide was different from our sequence: A
323TGCCT
328instead of our sequence A
323CGTCT
328. This results in the change of the primary structure of PLI-II from Thr
50-Ser
51to Met
50-Pro
51. These nucleotide substitution may reflect the in- traspecies diversification, or represent the similar gene on a different allele.
REFERENCES
[1] E . A . D e n n i s , J. Biol. Chem. , 269,
13057
-13060
(1994
).[2] D. L. Scott and P. B. Sigler, in :“Advances in Protein Chemistry”, 45,
53
-88
(1994
).[3] G. A. Boffa, M. C. Boffa, and J. J.
Winchenne, Biochim. Biophys. Acta, 429,
828
‒838
(1976
).[4] J. E. Fletcher, B.E. Rapuano, E. Condrea, C. C. Yang, and P. Rosenberg, Toxicol.
Appl. Pharmaco
l
., 59,375
-388
(1981
).[5] H. Kihara, R. Uchikawa, S. Hattori, and M. Ohno, Biochem. Int., 28,
895
-903
(1992
).[6] S. Tanaka, N. Mohri, H. Kihara, and M.
Ohno, J. Biochem., 99,
281
-289
(1986
).[7] S. -Y. Liu, K., Yoshizumi, N. Oda, M.
Ohno, F. Tokunaga, S. Iwanaga, and H.
Kihara, J. Biochem., 107,
400
-408
(1990
).[8] K. Yoshizumi, S. -Y. Liu, T. Miyata, S.
Saita, M. Ohno, S. Iwanaga, and H. Ki- hara, Toxicon, 28,
43
-54
(1990
).[9] R. D. Dunn and K. W. Broady, Biochim.
Biophys. Acta, 1533,
29
-37
(2001
).[
10
] S. Inoue, H. Kogaki, K. Ikeda, Y. Same- jima, and T. Omori-Satoh, J. Biol. Chem., 266,1001
-1007
(1991
).[
11
] S. Lizano, B. Lomonte, J. W. Fox, and J. M. Gutierrez, Biochem. J., 326,853
-859
(1997
).[
12
] N. Ohkura, S. Inoue, K. Ikeda, and K.Hayashi, J. Biochem., 113,
413
-419
(1993
).[
13
] N. Ohkura, S. Inoue, K. Ikeda, and K.Hayashi, Biochem. Biophys. Res. Common., 200,
784
-788
(1994
).[
14
] J. Perales, C. Villela, G. B. Domont, V.Choumet, B. Saliou, H. Moussatche, C.
Bon, and G. Faure, Eur. J. Biochem., 227,
19
-26
(1995
).[
15
] S. Inoue, A. Shimada, N. Ohkura, K.Ikeda, Y. Samejima, T. Omori-Satoh, and K. Hayashi, Biochem. Mol. Biol. Int., 41,
529
-537
(1997
).[
16
] H. Kogaki, S. Inoue, K. Ikeda, Y. Same- jima, T. Omori-Satoh, and K. Hayashi, J.Biochem., 106,
966
-971
(1991
).[
17
] N. Ohkura, H. Okuhara, S. Inoue, K.Ikeda, and K. Hayashi, Biochem. J., 325,
527
-531
(1997
).[
18
] C. L. Fortes-Dial, Y. Lin, J. Ewell, C. R.Diniz, and T. -Y. Liu, J. Biol. Chem., 269,
15646
-15651
(1994
).[
19
] K. Okumura, K. Masui, S. Inoue, K.Ikeda, and K. Hayashi, Biochem. J., 341,
165
-171
(1999
).[
20
] N. Ohkura, S. Inoue, K. Ikeda, and K.Hayashi, Biochem. Biophys. Res. Commun., 204,
1212
-1218
(1994
).[
21
] I. Nobuhisa, S. Inamasu, M. Nakai, A.Tatsui, T. Mimori, T. Ogawa, Y. Shi- mohigashi, Y. Fukumaki, S. Hattori, H.
Kihara, M. Ohno, Eur. J. Biochem., 249,
838
-845
(1997
).[
22
] M. Deshimaru, C. Tanaka, A. Tokunaga, M. Goto, and S. Terada, Fukuoka Univ.Sci. Rep., 33,
45
-53
(2003
).[
23
] U. K. Laemmli, Nature, 227,680
-685
(1970
).[
24
] H. Schagger, G. von Jagow, Anal. Bio- chem., 166,368
-379
(1987
).[
25
] M. Friedman, L. H. Krull, J. F. Cavins, J.Biol. Chem., 245
3868
-3871
(1970
).[
26
] K. Okumura, S. Inoue, N. Ohkura, K.Ikeda, and K. Hayashi, IUBMB Life, 48,
99
-104
(1999
).[
27
] Y. Wang, J. Dang, L. K. Johnson, J. J.Selhamer, and W. F. Doe, Eur. J. Biochem., 227,
116
-122
(1995
).[
28
] T. J. Fleming, C. O'hUigin, and T. R.Malek, J. Immunol., 150,
5379
-5390
(1993
).[
29
] J. G. Petranka, D.E. Fleenor, K. Sykes, R.E. Kaufman, and W. F. Rosse, Proc. Natl.
Acad. Sci. U.S.A., 89,
7876
-7879
(1992
).[
30
] N. Fuse, T. Tsuchiya, Y. Nonomura, A.Menez, and T. Tamiya, Eur. J. Biochem., 193,
629
-633
(1990
).[
32
] P. G. Hains, B. Nield, S. Sekuloski, R.Dunn, and K. Broady, J. Mol. Biol., 312,
875
-884
(2001
).[
33
] K. Nakashima, I. Nobuhisa, M. Deshima- ru, M. Nakai, T. Ogawa, Y. Shimohigashi, Y. Fukumaki, M. Hattori, Y. Sakaki, S.Hattori, and M. Ohno, Proc. Natl. Acad.
Sci. U.S.A., 92,
5605
-5609
(1995
).[
34
] S. So, T. Chijiwa, I. Nobuhisa, N. Oda- Ueda, S. Hattori, and M. Ohno, DDBJ/EMBL/GenBank database, http://www.
ddbj.nig.ac.jp

![Fig. 4 . Comparison of the amino acid sequences of γ -type phospholipase A 2 inhibitors. (A) Sub- Sub-units A, ( 1 ) [accession No. BAA 24503 ] T](https://thumb-ap.123doks.com/thumbv2/123deta/6406072.2138281/7.773.118.654.127.786/fig-comparison-amino-sequences-phospholipase-inhibitors-units-accession.webp)