閉鎖性の高い中海で垂下養殖されたサルボウガイの成長と生残
鈴木秀幸
1・山口啓子
2,*・瀬戸浩二
3Growth and Survival of the Ark Shell Scapharca kagoshimensis Cultured
in a Semi-enclosed Lagoon, Lake Nakaumi, Southwest Japan
Hideyuki S
UZUKI1, Keiko Y
AMAGUCHI2,*and Koji S
ETO3Abstract: Growth and survival of the ark shell Scapharca kagoshimensis cultured within net-bags were examined under various environmental conditions in Lake Nakaumi. In summer, ark shells cultured in the surface layer of water showed higher growth than those in the bottom layer of water. In winter, ark shells cultured in the bottom layer showed higher growth than those in the surface layer. The growth and survival of ark shells were mainly influenced by dissolved oxygen in summer, while in winter, their growth was influenced by water temperature. These results indicate that hanging culture of the ark shells is possible using net-bags without bottom sediment, and also suggest that management of culturing depth is efficient for their growth through the year.
Key words: Scapharca kagoshimensis; Hanging culture; Growth; Survival
2010年 8 月13日受付;2010年12月14日受理.
1
島根大学大学院生物資源科学研究科(Graduate School of Life and Environmental Science, Shimane University, 1060 Nishikawatsu-cho, Matsue, Shimane 690-8504, Japan).
2
島 根 大 学 生 物 資 源 科 学 部(Faculity of Life and Environmental Science, Shimane University, 1060 Nishikawatsu-cho, Matsue, Shimane 690-8504, Japan).
3
島 根 大 学 汽 水 域 研 究 セ ン タ ー(Research Center for Coastal Lagoon Environments, Shimane University, 1060 Nishikawatsu-cho, Matsue, Shimane 690-8504, Japan).
* 連絡先(Corresponding author): Tel & Fax: (+81) 852-32-6588; E-mail: [email protected](K. Yamaguchi).
わが国の沿岸海域では,埋立てや護岸工事といっ た開発や人為的富栄養化によって環境が悪化し,漁業 資源の減少が問題となっている(日比野 1984; 眞鍋ら 1994; 清水 1997; 佐藤・田北 2000)。島根県と鳥取県 の間に位置する中海も,開発による環境改変と富栄養 化が問題とされる閉鎖性の高い潟湖であり,1989年に 湖沼水質保全特別措置法の指定水域に指定された。 現 在の中海は,農地造成のための国営中海土地改良事業 (干拓・淡水化事業)により造られた干拓堤防によっ て仕切られ,本庄水域と中海本湖に分かれている(Fig. 1)。干拓堤防建設後,海水の流入経路が変化し(倉門 ら 1998),湖水と海水の交換が著しく低下し,貧酸素 水塊が拡大した(相崎 2000)。 中海の水産業は,1960年以前は平均で年間約4000 t の漁獲があったが,その後減少し,現在は500 t 前後 で推移している(相崎 2000; 森脇・道根 2007)。中で も上述の貧酸素の影響を受けやすい底生生物の甲殻 類や貝類は,減少が著しく,甲殻類は最盛期の70∼ 100 t から10 t 前後,貝類においては,かつて1500 t 近 くあった漁獲が,現在は20∼30 t で推移している(森 脇・道根 2007)。現在,中海で漁獲される貝類はアサ リ Ruditapes philippinarum が中心であり,かつて中 心であったサルボウガイ Scapharca kagoshimensis は 漁獲されていない(森脇・道根 2007)。しかし,漁獲 対象となるほどの資源量ではないが,サルボウガイ は現在も中海の一部に生息が確認されている(清川ら 1996,1997; 道根ら 2009)。島根県が行った採苗実験 では,中海奥部でサルボウガイ稚貝の出現が確認され ているが(安木ら 1994; 松本ら 1995),底生生物調査 では,中海奥部にサルボウガイはほとんど出現してい ない(清川ら 1997; 中村ら 1997)。中海奥部は,夏季 に貧酸素化し,底質は硫化水素が発生する還元的環境
となることから(Sakai et al. 2004),幼生の供給はあ るが,その多くが無効分散となっていると考えられる。 このため,サルボウガイ資源の再生には,稚貝の生残 率の高い飼育が必要である。 また,漁業による漁獲は,富栄養化した水域からの 栄養塩の系外除去につながる(Nakamura et al. 1988; 山口・相崎 2003)。特に二枚貝の多くはろ過食性で あることから,赤潮を抑制する効果があり(鬼塚ら 2002; 中田・山室 2004),サルボウガイ漁業の再興に より水質浄化,生態系構造の安定化,漁業資源とし ての活用と多面的な効果が期待できる(相崎・福地 1998)。 中海と同様,強内湾性である有明海では,二枚貝 漁業が盛んであるが,1980年代以降漁獲量は減少し ている(佐々木 2005a)。有明海は,自然環境の変動 による潮流の弱まりや,諫早湾干拓事業による潮受 け堤防の締め切りによって,河川水由来の低塩分水 の輸送パターンの変化で成層構造が生じやすくなり, 赤潮や貧酸素化が起こりやすくなったといわれてお り(程木 2005),漁獲量の減少と干拓事業による環 境悪化との関係が示唆されている(佐々木 2005b; 佐 藤ら 2006)。環境が悪化した有明海では,貧酸素水 塊を避けるため,タイラギ(リシケタイラギ Atrina lischkeana)を垂下養殖する試みがなされており(水産 総合研究センター 2009),サルボウガイについても研 究されている(圦本ら 2007)。しかし,圦本ら(2007) の実験は,殻長32 mm 以上の成貝を用いており,稚 貝からの垂下養殖については検討されていない。また, 日下部(1954)や高見・吉岡(1983),片山ら(1986)は, 殻長18 mm 以下の稚貝を用いて垂下養殖が試験され ているが,様々な環境条件での比較はされていない。 そこで本研究では,閉鎖性の高い中海をモデルフィー ルドとして稚貝からの垂下飼育実験と環境調査を行 い,各種環境下でのサルボウガイの成長と生残を検討 した。さらに,その評価のために桁曳き調査を行い, 天然で生息するサルボウガイの生育状態と比較した。 材料および方法 中海・本庄水域の概要 中 海 は 湖 水 面 積86.2 km2, 平 均 水 深5.4 m と 比 較 的 浅 い 内 湾 で あ る( 高 安 2001)。 狭 い 境 水 道( 幅 0.3 km,長さ7.5 km)から海水が流入するため,底層 は海水に近く,一方表層は,斐伊川から流入する淡水 が宍道湖を経て,大橋川から流入するため,中塩分で あり,成層構造が発達している(高安 2001)。中海では, 夏季に下層が貧酸素化する成層期と(伊達ら 1989; 森 脇・大北 2003),冬季に季節風の影響で鉛直混合が起 こりやすい循環期がある(Godo et al. 2001)。中海は, 塩分変動が大きく,しばしば表層は低塩分化し,底層 は,貧酸素化する(近藤ら 1994; 清家ら 1999)。一方, 本庄水域は極端に閉鎖性の強い水域であり,南西部の 開口部でのみ中海の表層水と本庄水域の水塊が交換さ れるため,塩分躍層ができにくい特徴がある(清家ほ か 1999; Nomura and Seto 2002)。
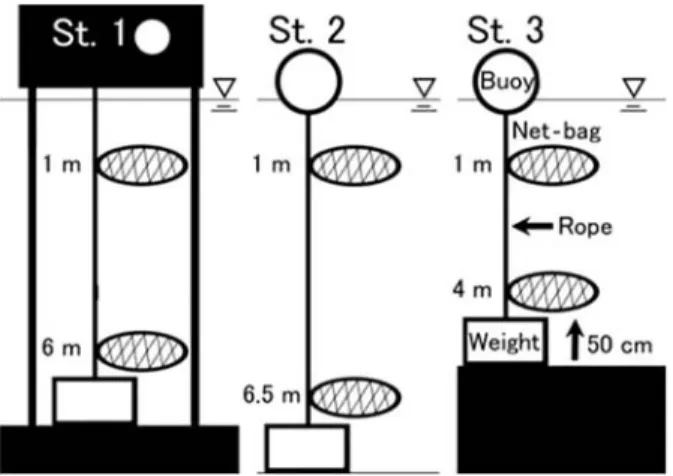
垂下飼育実験 実験には,殻長12.95±0.84 mm(平均値±標準偏差), 体重0.6±0.2 g のサルボウガイ稚貝を480個体用いた。 実験に使用したサルボウガイは,島根県水産技術セ ンターが2007年夏から秋に中海での採苗実験で採集し た稚貝を同年12月に譲り受け,島根大学汽水域研究セ ンター中海分室の流水水槽で飼育していた当歳貝とし た。実験は2008年 5 月から2009年 5 月まで行った。 実験地点は,St. 1 ∼ 3 の 3 地点とし,既存の研究(伊 達ら1989; 近藤ら1994; 清家ら 1999; Godo et al. 2001; Nomura and Seto 2002; 森脇・大北2003)から以下の ような特徴をもつ実験区とした(Fig. 1)。St. 1 は国土 交通省の中海湖心観測塔で,中海の中央に位置してい る代表的な地点であり,躍層が発達し夏季に底層が貧 酸素化する地点として設定した(平均水深6.5 m)。St. 2 は本庄水域内で,夏期に底層が貧酸素化するが,塩 分躍層があまり発達しない地点として設定した(平均 水深7.0 m)。St. 3 は境水道付近で,塩分躍層が発達す るが,境水道から酸素が豊富な海水が流入し,貧酸素 化しにくい地点として設定した(平均水深4.5 m)。 各実験区の表層と底層において,サルボウガイ稚貝 を40個体ずつ収容したネット(440 mm×290 mm)を 2 個ずつ垂下飼育し,合計12個のネットを設置した。
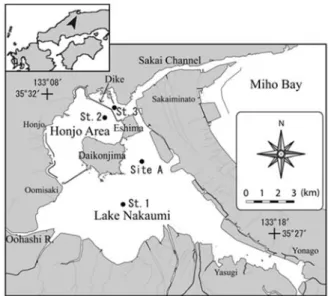
Fig. 1. Location of study sites (St.1, 2, 3 and Site A) in Lake Nakaumi, southwest Japan. St. 1~3, stations of cultur-ing ark shells; Site A, site of collectcultur-ing ark shells.
ネットはそれぞれロープ,コンクリートブロックおよ びブイを用いて設置した。ここで表層はそれぞれ水深 1 m に設置し,底層は,底質の影響を避けるため,湖 底から50 cm 程度上に設置した。St. 1 については国交 省の観測塔の建造物から垂下した(Fig. 2)。 測定項目 月に 1 回の頻度でサルボウガイの生残数,サイズ測 定,水質測定を行った(2009年 1 月は水質測定のみ)。 サルボウガイは研究室に持ち帰り,生存個体と死亡 個体に分け,生存個体についてはデジマチックキャリ パ(CD-S15C,Mitutoyo 社)を用いて殻長を,電子天 秤(EC-411G,ACCULAB 社)を用いて殻付湿重量を 測定した。測定後は,ネットを新しいものに交換し, 実験地点に再び設置した。成長は,実験終了時の各地 点,各層の殻長について比較し,さらに夏季(2008年 5 月∼2008年10月)と冬季(2008年11月∼2009年 4 月) に分けて殻長の成長量を比較した。成長量は,それぞ れ期間終了時の各個体の殻長から期間開始時の平均殻 長を引いて算出した。また,実験終了時に各条件の肥 満度を測定した。肥満度の測定は,末光ら(2001)の 方法に従って行った。殻長,成長量および肥満度につ いて,一元配置の分散分析(1-way ANOVA)を行い, 有意差が認められたため,Tukey の多重比較を行っ た。水質は,水温(℃),塩分(psu),溶存酸素(以 後,DO,mg/l)およびクロロフィル a(以後,Chl. a, ppb)について,多項目水質計(MINI SONDE 4, HYDROLAB社またはAAQ,JFE ALEC社)を用いて, 表層(水深1 m)と底層(湖底上0.5 m)を測定した。 桁曳き調査 江島の南(35°29´N,133°12´E 付近)に Site A を設 けた(Fig. 1)。Site A は,漁業者からサルボウガイの 生息情報があり,桁曳き調査で生息が確認された地点 である(道根ら 2009)。桁曳き調査は2008年 6 月25日, 7 月24日,8 月28日,10月 3 日 ,10月28日,11月10日, 12月17日,2009年 4 月29日,6 月 2 日,7 月10日,8 月 10日,9 月10日に行った。漁業者が所蔵していたサ ルボウガイの伝統的漁具であるサルボウ桁(桁幅110 cm,爪長15 cm,爪間隔15 mm,網目幅15 mm)を, 船で200 m を 2 ∼ 4 回曳き,底質中のサルボウガイを 採集した。なお,曳網距離の200 m は,ハンディ GPS (Map 76, Garmin 社)で計測した。調査には,島根大 学汽水域研究センターの調査船ルピアを使用した。ま た,桁曳きと同時期に湖底直上水(約10 cm)の水温, 塩分,DO について,多項目水質計(MINI SONDE 4a, HYDROLAB 社)を用いて測定した。2009年の 1 ∼ 3 月については桁曳きが行えなかったため,水質測 定のみ行った。 採集したサルボウガイは,デジマチックキャリパを 用いて殻長,殻高および殻幅を,電子天秤(PB153-S, METTLER TOLEDO 社)を用いて殻付湿重量を測定 した。測定したサルボウガイは殻長組成から大型個体 と小型個体,新規加入個体に分け,それぞれの肥満度 を測定した。 結 果 垂下飼育実験 Fig. 3 に 3 地点の各層におけるサルボウガイの生残 率の経月変化を示した。表層の生残率は,3 地点とも 実験終了時まで80%以上を維持した(Fig. 3A)。また Fig. 2. Schematic illustration of hanging culture system.
A
B
Survivalrate (%) 100 80 60 40 20 0 100 80 60 40 20 0 2008 2009 Month M J J A S O N D J F M A MFig. 3. Monthly changes in sur vival rate of ark shells cultured in surface and bottom layers of water at each sta-tion. ○, St. 1; □, St. 2; ▲, St. 3. A, surface layer; B, bottom layer.
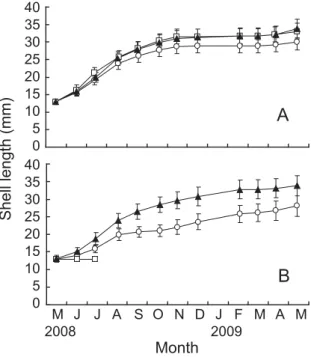
底層においては,St. 2 の生残率は 7 月に80%以上で あったが,8 月には 0%となった(Fig. 3B)。St. 1およ び St. 3 は,12月まで徐々に生残率は低下したが,実 験終了時まで60%を維持した。 3 地点の各層におけるサルボウガイの殻長の経月変 化を Fig. 4 に示した。殻長の平均値は,実験開始時に 12.95 mm であったが,実験終了時の表層において, St. 1 は30.04 mm,St. 2 は33.10 mm,St. 3 は33.81 mm となった(Fig. 4A)。表層は,5 ∼11月の 6 ヶ月間で およそ2.2∼2.4倍に達したが,11∼ 5 月の 6 ヶ月間で は,1.1倍となった。一方底層においては,実験終了 時に St. 1 は28.02 mm,St. 3 は33.80 mm となった(Fig. 4B)。底層は,5 ∼11月の 6 ヶ月間で殻長はおよそ1.7 ∼2.3倍に達し,11∼ 5 月の 6 ヶ月間では,1.1∼1.3倍 となった。また,St. 2 の底層は 8 月に全滅したが,5 ∼ 6 月に成長は全くみられなかった。 Fig. 5 に実験終了時(2009年 5 月)における殻長の 平均値と多重比較の結果を示した。表層においては, St. 2(33.10±2.32 mm) と St. 3(33.81±2.66 mm) で差はみられなかったが(P > 0.05),St. 1(30.04± 2.27 mm)は St. 2 および St. 3 に比べ成長が小さかっ た(P <0.01,Fig. 5)。底層においては,St. 3(33.83 ±2.89 mm)は St. 1(28.01±2.87 mm)よりも成長し ていた(P <0.01)。地点ごとにみると,St. 1 は表層 と底層で成長に差がみられた(P <0.01)。一方,St. 3 は表層と底層に差はみられなかった(P > 0.05)。また, 表層,底層ともに St. 1 の成長は,他の 2 地点よりも 小さかった(P <0.01)。
A
B
Shell length (mm) 40 35 30 25 20 15 10 5 0 40 35 30 25 20 15 10 5 0 2008 2009 Month M J J A S O N D J F M A MFig. 4. Monthly changes in mean shell length of ark shells cultured in surface and bottom layers at each station (bars = standard deviation). ○, St. 1; □, St. 2; ▲, St. 3. A, surface layer; B, bottom layer.
a b c c c Shell length (mm) Experimental condition St.1S St.1B St.2S St.3S St.3B 40 35 30 25 20 15 10 5 0 0 5 10 15 20 25 - 2 0 2 4 6 8 a a a b b b c c a b Growth increment (mm) Growth increment (mm) Summer Winter Experimental condition St.1S St.1B St.2S St.3S St.3B St.1S St.1B St.2S St.3S St.3B Fig. 5. Mean shell length of ark shells for each experi-mental condition in May 2009 (bars = standard devia-tion). S, sur face layer; B, bottom layer. St. 1 S, n=66; St. 1 B, n=47; St. 2 S, n=75; St. 3 S, n=54; St. 3 B, n=26. Alphabetical letters indicate the results of Tukey’s mul-tiple comparison test (a<b<c, P<0.05).
Fig. 6. Mean growth increment of shell length of ark shells for each experimental condition from (Summer, May 2008 to October 2008; St. 1 S, n=68; St. 1 B, n=57; St. 2 S, n=76; St. 3 S, n=58; St. 3 B, n=30) and (Winter, November 2008 to April 2009; St. 1 S, n=68; St. 1 B, n=57; St. 2 S, n=76; St. 3 S, n=58; St. 3 B, n=30). Bars=standard deviation S, surface layer; B, bottom layer. Alphabetical let-ters indicate the results of Tukey’s multiple comparison test (a<b<c, P<0.05).
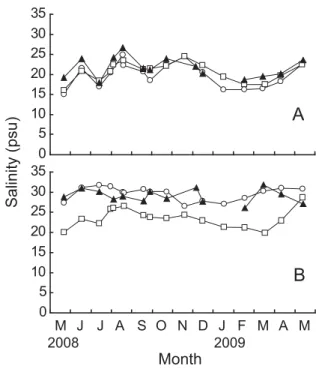
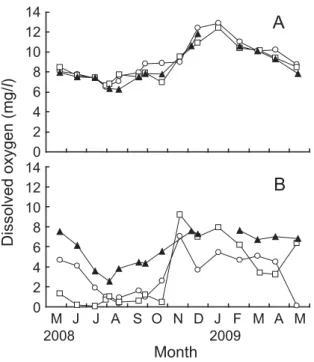
Fig. 6 に 夏 季(2008年 5 月 ∼2008年10月) と 冬 季 (2008年11月∼2009年 4 月)における殻長の成長量の 平均値と多重比較の結果を示した。夏季は,St. 1 と St. 3 はともに表層の成長量(St. 1, 14.79±1.85 mm; St. 3, 16.92±2.18 mm)が底層の成長量(St. 1, 8.09± 1.61 mm; St. 3, 15.42±2.18 mm)よりも高く,それぞ れ1.8倍,1.1倍であった(P <0.05,Fig. 6A)。冬季は 逆に,底層の成長量(St. 1, 4.66±2.75 mm; St. 3, 3.44 ±2.78 mm)が表層の成長量(St. 1, 0.50±2.05 mm; St. 3, 1.23±2.35 mm)よりも高く,St. 1 は9.4倍,St. 3 は2.8倍であった(P <0.05,Fig. 6B)。 Fig. 7 に実験終了時における肥満度の平均値と多重 比較の結果を示した。表層においては,3 地点とも差 はみられなかった(St. 1, 0.054±0.006; St. 2, 0.057± 0.005; St. 3, 0.054±0.004; P > 0.05,Fig. 7)。底層でも St. 1とSt. 3で差はみられなかった(St. 1, 0.035±0.005; St. 3, 0.039±0.005; P > 0.05)。地点ごとにみると,St. 1 と St. 3 はともに表層の肥満度は底層よりも高く,そ れぞれ1.6倍,1.4倍であった(P <0.01)。 Figs. 8 ∼11に水質測定の結果を示した。水温は, 表層において地点による差はほとんどなく,何れも 8 月上旬に30℃を上回り,1 月に約 6℃まで低下した (Fig. 8A)。底層水温は,表層と同様の変動を示した が,2009年 1 月の St. 1 と St. 2 では,それぞれ10℃, 6℃ と な っ た(St. 3 に つ い て は 欠 測,Fig. 8B)。 塩 分は表層において年間を通じて15∼26 psu を示した (Fig. 9A)。底層においては,St. 2 は表層塩分と同じ 範囲内にあったが,St. 1 とSt. 3 は20∼32 psu を示し た(Fig. 9B)。DO は, 表 層 に お い て 年 間 を 通 し て 6 mg/l(=84%)以上を示した(Fig. 10A)。底層 DO は, 10月まで St. 3,St. 1,St. 2 の順で高く,St. 3 は2.5∼ 7.6 mg/l(=35∼106%),St. 1 は0.9∼7.1 mg/l(=13 ∼99%),St. 2 は0.1∼9.2 mg/l(=1∼127%)を示し た(Fig. 10B)。Chl. a は,表層においては地点による 差はほとんどみられず,1.62∼10.86 ppb を示した(Fig. 11A)。底層 Chl. a は,St. 2 で 9 月と 1 月に20 ppb を 超える値を示したが,それ以外では 3 地点とも0.85∼ 13.50 ppb を示した(Fig. 11B)。 b a b b a St.1S St.1B St.2S St.3S St.3B Experimental condition Condition index 0.07 0.06 0.05 0.04 0.03 0.02 0.01 0.00 0 5 10 15 20 25 30 35 0 5 10 15 20 25 30 35
A
B
Water temperature ( ℃ ) 2008 2009 Month M J J A S O N D J F M A MFig. 7. Mean condition index of ark shells for each experi-ment condition in May 2009 (bars = standard deviation). S, surface layer; B, bottom layer. St. 1 S, n=10; St. 1 B, n=10; St. 2 S, n=9; St. 3 S, n=10; St. 3 B, n=10. Alphabetical let-ters indicate the results of Tukey’s multiple comparison test (a<b<c, P<0.05).
Fig. 8. Monthly changes in water temperature of surface and bottom layers at each station. ○, St. 1; □, St. 2; ▲, St. 3. A, surface layer; B, bottom layer.
0 5 10 15 20 25 30 35 0 5 10 15 20 25 30 35
A
B
2008 2009 Month M J J A S O N D J F M A M Salinity (psu)Fig. 9. Monthly changes in salinity of surface and bottom layers at each station. ○, St. 1; □, St. 2; ▲, St. 3. A, sur-face layer; B, bottom layer.
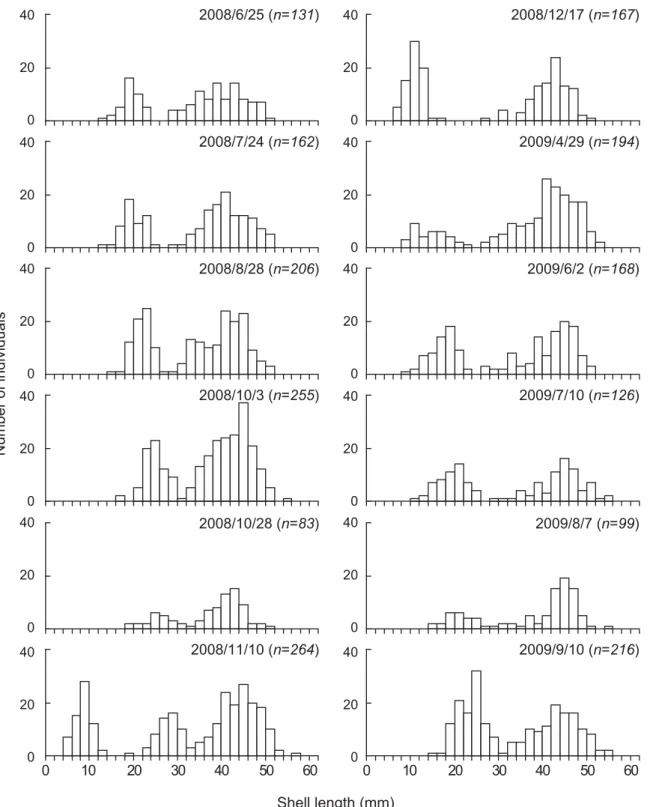
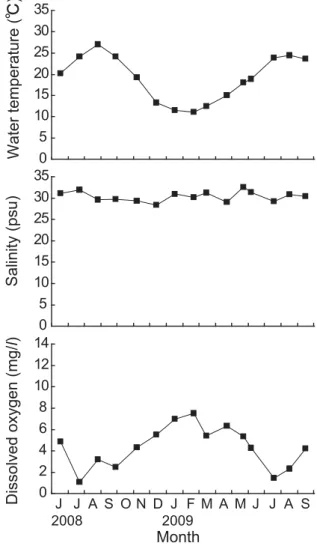
桁曳き調査 Fig. 12に2008年 6 月 ∼2009年 9 月 ま で に 桁 曳 き (Site A)にて採取した個体の殻長組成の推移を示し た。2008年 6 月の殻長組成では,明瞭に 2 つのサイズ グループに分かれ,殻長約43 mm にピークをもつ大 型の個体群と約19 mm にピークをもつ小型の個体群 が存在していた。月を経るごとに,小型の個体群のピー クは大型の個体群のピークに近づいていた。11月には 殻長約 8 mm にピークをもつ新規加入個体が確認され た。2008年12月以降は11月までみられた大型および小 型個体を明瞭に区別することができなくなった。大型 の個体群のピークや平均殻長には,大きな変動はみら れなかった。 Fig. 13にサルボウガイの肥満度の経月変化を示し た。大型個体と小型個体で肥満度の変化が異なってい た。大型個体の平均値は2008年 6 月に0.026であった が,8 月に0.016 に低下した。小型個体は 6 ∼ 8 月まで 約0.015 で減少はみられなかった。12月になると,大型, 小型個体ともに肥満度は増加した。12月以降は大型, 小型個体は同様の変化を示し,2009年 5 月にピークが みられ,その後 9 月まで減少した。新規加入個体は, 2008年12月に0.027になり,その後2009年 9 月まで減 少し0.017となった。 Fig. 14に Site A の底層での水温,塩分および DO の 経月変化を示した。水温は 8 月に27℃まで上昇し,そ の後徐々に低下して 2 月に11℃となった。塩分は,期 間中変動は少なく30 psu 前後で推移した。DO は,7 ∼ 9 月にかけて 1 ∼ 3 mg/l(=14∼42%)の貧酸素状 態となったが,それ以外の期間は 4 ∼ 6 mg/l(=56∼ 84%)を示した。 考 察 St. 2 の底層において,8 月にサルボウガイの斃死が 起こり,生残率が 0%となった(Fig. 3)。この斃死と 環境要因との関係を考えると,水温や塩分の変動は少 なく,Chl. a も他の地点と同程度であったことから, 貧酸素が一因とみられる(Figs. 8 ∼11)。一般に底生 の貝類は1.5 ml/l(=30%,2.1 mg/l)以下の貧酸素で 危険になるとされる(柳 1989)。その貧酸素に St. 3 は ならなかったが,St. 1 は 7 ∼ 9 月,St. 2 は 5 ∼10月に なり,St. 2 は著しい貧酸素状態であった。11月以降は, 3 地点とも DO は十分に回復していた。St. 2 は,実験 開始時から貧酸素状態であったが 1 ∼ 2 ヶ月生残した ことから,サルボウガイの貧酸素耐性は高いが,2 ヶ 月以上の貧酸素状態には耐えられなかったことがわ かる。また,表層では 3 地点とも80%以上の生残率を 維持したが,底層では夏以降生残率は大きく低下した (Fig. 3)。St. 1 と St. 3の底層においても,貧酸素化し た夏期に生残率が低下していた。このように夏季の底 層では DO がサルボウガイの生存を制限しているこ とが窺えた(Fig. 10)。 次に成長についてみると,全滅した St. 2 の底層を 除き,サルボウガイに成長がみられた(Fig. 4)。底 0 5 10 15 20 25 30 0 5 10 15 20 25 30
A
B
Chlorophyll a (ppb) 2008 2009 Month M J J A S O N D J F M A MFig. 11. Monthly changes in chlorophyll a of surface and bottom layers at each station. ○, St. 1; □, St. 2; ▲, St. 3. A, surface layer; B: bottom layer.
A
B
Dissolved oxygen (mg/ l) 2008 2009 Month M J J A S O N D J F M A M 14 12 10 8 6 4 2 0 14 12 10 8 6 4 2 0Fig. 10. Monthly changes in dissolved oxygen of surface and bottom layers at each station. ○, St. 1; □, St. 2; ▲, St. 3. A, surface layer; B, bottom layer.
層の成長を地点間で比較すると,実験開始から明ら かに St. 3,St. 1,St. 2 の順で成長が小さくなってい た(Fig. 4)。この時期,3 地点で DO が明らかに異な り,成長に大きく影響したと考えられる(Fig. 10)。 近縁種であるアカガイ S. broughtonii は,DO が平均 2 ml/l(≒2.9 mg/l)になると成長が阻害されることが 知られている(中西 1981)。サルボウガイは多くの二 枚貝と同様に,換水を行い,それとともに鰓で水中の 懸濁物を捕捉し,ろ過摂食している(菊池 2003)。し かし,一般に二枚貝は環境水の著しい低酸素化が起こ ると,換水量は減少する(Sobral and Widdows 1997; 山元ら1998,1999,2006)。実験開始時(2008年 5 月) から St. 2 の底層の生残率が 0%となるまでの 2 ヶ月間 の Chl. a をみると,5 月の時点で St. 2 は,5 ppb 以上 と明らかに他の 2 地点よりも高かったが,まったく成 長がみられなかった(Figs. 4, 11)。これは著しい貧酸 素状態(最低0.1 mg/l, 1%)であったため,サルボウ ガイが換水を行わなかったためによると考えられた。 0 10 20 30 40 50 60 0 10 20 30 40 50 60 40 20 0 40 20 0 40 20 0 40 20 0 40 20 0 40 20 0 40 20 0 40 20 0 40 20 0 40 20 0 40 20 0 40 20 0 Number of individuals Shell length (mm) 2008/6/25 (n=131) 2008/12/17 (n=167) 2008/7/24 (n=162) 2009/4/29 (n=194) 2008/8/28 (n=206) 2009/6/2 (n=168) 2008/10/3 (n=255) 2009/7/10 (n=126) 2008/10/28 (n=83) 2009/8/7 (n=99) 2008/11/10 (n=264) 2009/9/10 (n=216)
St. 1 と St. 3 では,Chl. a から餌量が成長に関係した 可能性も考えられるが,富栄養湖である中海で餌不足 は考え難い。なぜなら,中海の懸濁物量は上層と下層 で同程度か下層が高く(伊達ら1989; 橋谷ら1991), Hiratsuka et al.(2007)によると,中海の懸濁物質は 主に植物プランクトンおよび,植物プランクトン起源 のデトリタスである。すなわち,底層では光が届かず Chl. a は低いが,植物プランクトンの死骸やデトリタ スが多く,餌として供給されていると考えられる。こ れらのことから,貧酸素化が起こる夏季は,DO が成 長を制限している主な要因と考えられた。 Fig. 5 よりサルボウガイの各条件における 1 年間の 成長差は小さいが,表層と底層で,時期によって成長 に異なる特徴がみられた(Fig. 4)。夏季(2008年 5 月 ∼2008年10月)と冬季(2008年11月∼2009年 4 月)で は,夏季は底層で成長量が小さく,逆に冬季は底層の 成長量が高いことが明らかとなった(Fig. 6)。宮本・ 初田(2008)の実験によると,サルボウガイは水温 8℃ では濾過活動が認められない。本実験では表層の最低 水温が 5℃であるのに対して,St. 1 底層は10℃であっ た(Fig. 8)。また,貧酸素状態は2008年11月には解消 されていた(Fig. 10)。このことから,冬季において 底層で高い成長がみられたのは,表層に比べ水温が低 下しにくく,濾過活性が表層より高く維持されたこと が一因と考えられた。 St. 1 での成長は表層,底層ともに他の地点と比べ, 明らかに小さかった(Fig. 5)。この原因としては,底 層は先述したように貧酸素化が主な要因と考えられる が,表層は塩分が影響した可能性が考えられた。本研 究での月 1 回程度の測定では,3 地点の表層間におけ る塩分差は0.1∼4.3 psu と僅かではあったが,一般に 中海湖心は河川からの低塩分水が直接流入するため, 境水道や本庄水域に比べ低塩分化しやすいことが知 られている(Kondo et al. 1990; 近藤ら 1994; 清家ら 1999)。一方,Chl. a は 3 地点とも同程度であったこ とから(Fig. 11),St. 1 は餌量による制限があったと は考え難い。以上のことから,低塩分化のストレスが St. 1 底層の成長を制限した一因と考えられた。また, St. 3 の 1 年間の成長に表層と底層で差はみられず, どちらも高かった(Fig. 5)。中海の湾口の水道では, 上潮時に鉛直混合が起こりやすいことが知られており (宇野ら 2001),St. 3 でも水がよく混ざることで St. 1 よりも水質の差が少なく,表層と底層の成長差が小さ くなったと考えられた。このように,閉鎖性の高い内 湾でサルボウガイの垂下養殖を行う際には,成層構造 の影響で成長と生残が変化することが示された。 Site A の殻長組成の推移より,天然個体は大型の 個体群と小型の個体群,新たに加入した個体群に分 かれていた(Fig. 12)。11月に新規加入した個体群は 2008年産卵の年級群と考えられ,2009年 6 月には殻 長約18 mm にピークを持つ個体群となっていた。こ のことから,2008年 6 月に約19 mm にピークのあっ Condition index 2008 2009 Month J J A S O N D J F M A M J J A S 0.045 0.040 0.035 0.030 0.025 0.020 0.015 0.010 0.005 0.000 0 5 10 15 20 25 30 35 0 5 10 15 20 25 30 35 0 2 4 6 8 10 12 14 2008 2009 Month J J A S O N D J F M A M J J A S Water temperature ( ℃ ) Salinity (psu) Dissolved oxygen (mg/ l)
Fig. 13. Monthly changes in mean condition index of ark shells collected from Site A (bars = standard deviation). ●, large-size individuals; □, small-size individuals; △, newly recruited individuals.
Fig. 14. Monthly changes in water temperature, salinity and dissolved oxygen of the bottom overlying water at Site A.
た小型個体は,2007年産卵の年級群で,1 年間の成長 を表していると判断できる。垂下飼育実験の個体と 桁曳きで得られた2007年産卵の年級個体(小型個体) を,2008年11月時点で比較すると,天然個体は殻長約 28 mm にピークが見られたが,表層で垂下飼育した 個体の殻長は,おおよそ30 mm 以上であり,垂下飼 育した個体の成長が勝っていた。 Site A の大型個体と小型個体では,肥満度の変化に 差がみられた(Fig. 13)。大型個体は 7 月から 8 月に かけて肥満度が低下したが,小型個体は低下しなかっ た。一般に,肥満度の減少には生殖活動による放卵 放精の影響があることが明らかとなっている(鳥羽ら 1993)。畑(1948)によると,1946年の中海では 7 月 下旬に親貝のほとんどが成熟しており,田中(1954)は, 有明海産のサルボウガイは 7 ∼ 8 月が産卵盛期であっ たとしている。圦本ら(2007)は,6 ∼ 8 月の肥満度 の低下は,生殖活動による可能性が高いと述べており, これらのことから,大型個体の肥満度の低下は,生殖 活動によるものと考えられた。一方,サルボウガイは 満 2 ヶ年で親貝となるが(日下部 1954),小型個体は 2007年産卵の生後 1 年の個体であり,未成熟であった ため肥満度が低下しなかったと考えられた。大型個体 の肥満度は,12月から 4 月にかけて増加しており,サ ルボウガイは冬季∼春季に肥満度が増加することがわ かった。また,垂下飼育した個体の肥満度は,表層で 高い傾向があったが(Fig. 7),実験個体の肥満度と同 時期の天然の小型個体の肥満度(2009年 6 月 2 日)を 比較すると,天然個体に比べ底層の実験個体の肥満度 はやや高く,表層の実験個体の肥満度においては,1.9 倍の平均0.055と極めて高い値であった。 天然個体群で 最も高い値は2009年 4 月の0.032であり,表層で垂下 飼育した個体の肥満度の方が明らかに高く,身太りし ていることが明らかとなった。 本研究では,サルボウガイをネットに入れるのみで 砂などを用いずに実験を行った。結果として,閉鎖性 の高い中海においては,サルボウガイは底質がなくて も十分成長し,生残できた。そして何より,天然個体 よりも大きく成長し,身太りしていた。本研究の結果 から,夏期は表層に垂下し,冬季は底層に垂下すると いったように,成層構造を考慮して時期によって垂下 する水深を変えることで年間を通して成長が見込め, 採苗から 1 年で十分に出荷サイズに育成が可能と考え られる。 一方,塩分については宮本・初田(2008)は,低 塩分はサルボウガイの濾過活動に不適であるとしてお り,山城(1959)や福原ら(1986)は,サルボウガイ 稚貝はおおよそ10‰以下で生息に不適と述べている。 しかし本研究の調査時は,例年(清家ら 1986; 近藤ら 1994; 清家ら 1999; 宇野ら 2001)よりも塩分の変動が 少なく,表層は15 psu 以上で安定していた。このこと から,表層がより低塩分化した際に,生残率が低下す ることが懸念され,この点については,今後の検討課 題である。 要 約 中海の環境条件が異なる 3 地点で,表層と底層に 垂下したネットにサルボウガイを入れ,2008年 5 月か ら2009年 5 月まで飼育し,成長,生残,肥満度を比較 した。また,これと同時に中海に生息する天然個体と の比較も行った。その結果,夏季は表層で成長が大き く,冬季は底層で成長が大きかった。夏季の成層期で は,DO がサルボウガイの成長・生残を制限する一因 であり,冬季の循環期では,水温が成長を制限する一 因であった。表層で垂下飼育したサルボウガイは天然 個体に比べ成長が速く,肥満度も明らかに高かった。 閉鎖性の高い中海では,サルボウガイは潜砂しなくて も成長し,夏季は表層に垂下し,冬季は底層に垂下す ることで,年間を通して高い成長率が見込めることが わかった。 謝 辞 本研究を遂行するにあたり,島根県水産技術セン ターの勢村 均・佐々木正両氏には,供試稚貝を快 くお譲りいただいた。鳥取県衛生環境研究所の宮本 康博士には,様々なご助言,ご協力をいただいた。中 海漁業協同組合の石倉正夫氏には,桁曳きの技術指導 をしていただいた。国土交通省出雲河川事務所には, 実験場所を快くご提供いただいた。島根大学水圏生態 学研究室の皆様には,分析処理でご協力いただいた。 なお、本研究は科学研究費補助金(挑戦的萌芽研究 19651007)「沿岸域環境モニタリングのための生物情 報解読手法の開発」の一部を用いて行った。以上の機 関と方々に,この場を借りて厚く御礼申し上げる。 文 献 相崎守弘・福地美和(1998)ヤマトシジミを用いた汽水性 汚濁水域の浄化.用水と排水,40,894-898. 相崎守弘(2000)湖沼生態系の保全と管理.環境修復のた め の 生 態 工 学( 須 藤 隆 一 編 ), 講 談 社, 東 京,pp. 113-138. 伊達善夫・橋谷 博・清家 泰・近藤邦男・奥村 稔・藤 永 薫(1989)12年間の定期調査からみた中海・宍道 湖の水質−季節変化,経年変化,平均値−.山陰地域 研究(自然環境),5,89-102.
福原 修・Waspada・梅沢 敏・野上和彦(1986)サルボウ 養殖種苗の塩分耐性.南西水研報,20,1-12.
Godo, T., K. Kato, H. Kamiya and Y. Ishitobi (2001) Observation of wind-induced two-layer dynamics in Lake Nakaumi, a coastal lagoon in Japan. Limnology, 2, 137-143. 橋谷 博・清家 泰・近藤邦男・奥谷 稔・藤永 薫・伊 達善夫(1991)月 1 回の定期調査からみた中海・宍道 湖の水質.山陰地域研究(自然環境),7,93-104. 畑 久三(1948)中海に於けるモガヒの自然發生に就て. 日水誌,13,248-250. 日比野雅俊(1984)三河湾−汚濁と集水域の自然環境.陸 水と人間活動 多摩川・霞ヶ浦・諏訪湖・中海・三河湾・ 琵琶湖(門司正三・高井康雄編),東京大学出版会,東京, pp. 209-254.
Hiratsuka, J., M. Yamamuro and Y. Ishitobi (2007) Long-term change in water transparency before and after the loss of eelgrass beds in an estuarine lagoon, Lake Nakaumi, Japan. Limnology, 8, 53-58. 程木義邦(2005)有明海浅海定線調査データでみられる表 層低塩分水輸送パターンの変化.有明海の生態系再生 をめざして(日本海洋学会編),恒星社厚生閣,東京, pp. 55-61. 片山勝介・池田善平・三宅与志雄(1986)異なる種苗を用 いて養殖したサルボウの成長とへい死.岡山水試報,1, 50-54. 菊池泰二(2003)ベントスとはどういうものか.海洋ベン トスの生態学(日本ベントス学会編),東海大学出版会, 神奈川,pp. 1-32. 清川智之・向井哲也・山根恭道・中村幹雄(1996)中海・ 宍道湖底質改良実証実験事業サルボウ天然稚貝発生状 況調査.島根県水産試験場事業報告(平成 8 年度),pp. 205-206. 清川智之・重本欣史・中村幹雄(1997)宍道湖・中海水産 振興対策調査事業有用水産動物調査(アサリ・サルボ ウ).島根県水産試験場事業報告(平成 9 年度),pp. 235-237.
Kondo, K., Y. Seike and Y. Date (1990) Relationships between phytoplankton occurrence and salinity or water tempera-ture in brackish lake Nakanoumi, 1979-1986. Jpn. J.
Limnol., 51, 173-184. 近藤邦男・清家 泰・橋谷 博(1994)汽水湖中海におけ る栄養塩類および植物プランクトンの鉛直分布を支配 する塩分躍層の役割.陸水学雑誌,55,47-60. 倉門由紀子・三瓶良和・高安克己・徳岡隆夫・井内美郎(1998) 中海および浜名湖表層堆積物の有機炭素・窒素・イオ ウ濃度分布.LAGUNA(汽水域研究),5,123-135. 日下部台次郎(1954)モガイとその養殖.水産増殖叢書 (6), 東京大学農学部水産学科,東京,75 pp. 眞鍋武彦・長井 敏・堀 豊(1994)厳しさを増す沿岸漁業. 水産と環境(清水 誠編),水産学シリーズ 103,恒星 社厚生閣,東京,pp. 9-18. 松本洋典・中村幹雄・山根恭道・向井哲也・清川智之(1995) 中海・宍道湖底質改良実証実験事業サルボウ天然稚貝 発生状況調査.島根県水産試験場事業報告(平成 7 年 度),pp. 125-134. 道根 淳・原 修一・木村 秀・青山喜久雄(2009)中海 におけるサルボウガイの桁曳き網調査.島根水技セ研 報,2,71-73. 宮本 康・初田亜希子(2008)塩分と水温に応じたサルボ ウ(Scapharca kagoshimensis)の濾過速度と生残率の変 化.LAGUNA(汽水域研究),15,13-18. 森脇晋平・大北晋也(2003)中海に出現する貧酸素水塊の 海況学的特性と海洋構造.LAGUNA(汽水域研究), 10,27-34. 森脇晋平・道根 淳(2007)中海における漁獲量変動.島 根水技セ研報,1,41-48.
Nakamura, M., M. Yamamuro, M. Ishikawa and H. Nishimura (1988) Role of the bivalve Corbicula japonica in the nitro-gen cycle in a mesohaline lagoon. Marine Biol., 99, 369-374. 中村幹雄・清川智之・山根恭道・内田 浩・福井克也(1997) 宍道湖・中海水産振興対策検討調査事業 宍道湖・中 海の底質環境と底生生物調査.島根県水産試験場事業 報告(平成 9 年度),pp. 205-228. 中西雅幸(1981)アカガイの成長におよぼす水温,塩分, 溶 存 酸 素 濃 度 の 影 響 に つ い て. 京 都 海 セ 研 報,5, 23-28. 中田喜三郎・山室真澄(2004)宍道湖・中海を対象とした 生態系モデル−懸濁物食性二枚貝の効果.海洋と生物, 26,267-278.
Nomura, R. and K. Seto(2002)Influence of man-made con-struction on environmental conditions in brackish Lake Nakaumi, southwest Japan, foraminiferal evidence. J.
Geol. Soc. Jpn., 108, 394-409.
鬼塚 剛・柳 哲雄・門谷 茂・山田真知子・上田直子・ 鈴木 學(2002)ムラサキイガイ養殖による洞海湾浄 化の試み.海の研究,11,403-417.
Sakai, S., M. Nakaya and K. Takayasu(2004)Hydrogen sulfide distribution in bottom and pore waters during an anoxic period in Lake Nakaumi, Japan. LAGUNA (ReCLE), 11, 65-68. 佐々木克之(2005a)漁業生産.有明海の生態系再生をめざ し て( 日 本 海 洋 学 会 編 ), 恒 星 社 厚 生 閣, 東 京,pp. 37-38. 佐々木克之(2005b)有明海環境変化と生態系異変の総括. 有明海の生態系再生をめざして(日本海洋学会編),恒 星社厚生閣,東京,pp. 167-173. 佐藤正典・田北 徹(2000)有明海の生物相と環境.有明 海の生きものたち 干潟・河口域の生物多様性(佐藤 正典編),海遊舎,東京,pp. 10-35. 佐藤慎一・金澤 拓・萩山竜馬(2006)諫早湾干拓事業に 伴う二枚貝類の変化.海洋と生物,28,618-624. 清家 泰・近藤邦男・伊達善夫・石田裕三郎(1986)汽水湖・ 中海における窒素代謝Ⅱ−夏期における底泥表層部で の脱窒特性−.陸水学雑誌,47,133-141. 清家 泰・奥村 稔・藤永 薫・大谷修司・千賀有希子・ 岡 宏樹(1999)中海本庄水域の水質特性.LAGUNA(汽 水域研究),6,1-9. 清水 誠(1997)水産生物.東京湾の生物誌(沼田 眞・ 風呂田利夫編),築地書館,東京,pp. 143-155.
Sobral, P. and J. Widdows (1997) Influence of hypoxia and anoxia on the physiological responses of the clam
Ruditapes decussatus from southern Portugal. Marine
Biol., 127, 455-461. 末光健治・山口啓子・相崎守弘(2001)ヤマトシジミの大 量斃死機構に関する基礎的研究Ⅱ.LAGUNA(汽水域 研究),8,39-46. 水産総合研究センター(2009)大型二枚貝タイラギの養殖 技術の開発に成功.FRANEWS,17,32. 高見東洋・吉岡貞範(1983)サルボウガイ養殖試験.山口 県内海水試報,11,26-39. 高安克己(2001)汽水域の科学,たたら書房,米子,183 pp. 田中彌太郎(1954)有明海産重要二枚貝の産卵期–Ⅰ サル ボウについて.日水誌,19,1157-1160. 鳥羽光晴・夏目 洋・山川 紘(1993)東京湾船橋地先に おけるアサリの生殖周期.日水誌,59,15-22. 宇野誠高・田中秀典・大塚泰介・高安克己・鮎川和泰(2001) 中海中浦水道における水質動向.LAGUNA(汽水域研 究),8,15-22. 山口啓子・相崎守弘(2003)底生生物の浄化作用−ヤマト シジミを中心として−.エコテクノロジーによる河川・ 湖沼の水質浄化−持続的な水環境の保全と再生−(島 谷幸宏・細見正明・中村圭吾編),ソフトサイエンス社, 東京,pp. 31-43. 山元憲一・半田岳志・中村真敏・田村晃一・韓青渓(1997) ミドリイガイの呼吸に及ぼす低酸素の影響.水産増殖, 46,523-527. 山元憲一・安達 智・河邊 博(1998)アコヤガイの呼吸 に及ぼす低酸素の影響.水産増殖,47,539-544. 山元憲一・半田岳志・西岡 晃(2006)リシケタイラギの 換水に及ぼす低酸素の影響.水産増殖,54,319-323. 山城隆文(1959)サルボウの生態学的研究−Ⅰサルボウ稚 貝 の 低 比 重 海 水 に 対 す る 抵 抗 力. 水 産 増 殖,6(3), 27-32. 柳 哲雄(1989)シンポジウム「貧酸素水塊」のまとめ. 沿岸海洋研究ノート,26,141-145. 安木 茂・中村幹雄・向井哲也・山根恭道・松本洋典(1994) 中海・宍道湖底質改良実証実験事業–Ⅰ(中海の実証実 験区).島根県水産試験場事業報告(平成 6 年度),pp. 158-168. 圦本達也・那須博史・鳥羽瀬憲久・前野幸男(2007)有明 海においてカゴ飼育されたサルボウの成長,生残およ び摂餌状態.水産増殖,55,535-540.