報 告
日本哺乳類学会北海道網走郡大空町で確認されたヒメヒナコウモリの
出産哺育コロニー
近藤 憲久
1, 福井 大
2, 倉野 翔史
3, 黒澤 春樹
4 1根室市歴史と自然の資料館2Division of Animal Resources, National Institute of Biological Resources, Korea
3東京農業大学産業生物学部動物生産管理学研究室 4道東コウモリ研究会
摘
要
北海道網走郡大空町にある旧大成小学校体育館で,コ
ウモリの出産哺育コロニーが発見された.本コロニーを
形成する個体の捕獲を行い,外部形態を精査したところ,
乳頭が 2 対あることからヒメヒナコウモリと同定した.
また,8 月以降にコロニー周辺で拾得された 2 個体のコ
ウモリについても,外部並びに頭骨計測値からヒメヒナ
コウモリと同定した.5 回にわたる捕獲調査の結果,本
種は 6 月下旬~ 7 月上旬に出産し,8 月上旬には幼獣が飛
翔を始めていた.本コロニーを形成する雌成獣は約 60 頭
であった.8 月以降は,成獣はほとんどいなくなり,幼
獣が大部分(96%)を占めていた.飛翔時の音声構造は,
FM-QCF
型であり,ピーク周波数の平均値は 26.1 kHz で
あったが,FM 成分と QCF 成分の比率は飛翔環境によっ
て大きく変化していた.ヒメヒナコウモリのねぐらおよ
び出産哺育個体群は国内初記録であり,今回の発見によ
り,本種の国内における繁殖・定着が明らかになった.
は じ め に
ヒメヒナコウモリ Vespertilio murinus は,北緯 35° から
北緯 62° のスカンジナビア,西・中央ヨーロッパ,ウラ
ル山脈,アゼルバイジャン,北イラン,アフガニスタン,
ロシア,モンゴル,極東ウスリー地方に分布する中型の
食虫性コウモリで(Baagøe 1986;Rydell and Baagøe 1994;
Blant and Jaberg
1995;Rakhmatulina 1995;Syvertsen et al.
1995;Tiunov 1997;Smith and Xie 2008;Dietz et al. 2009;
Orlova
2011),人工構造物や洞穴,岩の割れ目をねぐら
として利用する(Strelkov 1969;Schober et al. 1993;Rydell
and Baagøe
1994;Tiunov 1997;Dietz et al. 2009).季節移
動をほとんど行なわない個体群がある(Tiunov 1997;
Baagøe
2001;Baranauskas et al. 2006;Dietz et al. 2009;Suba
et al.
2010)一方で,標識調査によって 1,000 km を超える
長距離季節移動も確認されている(Masing 1989;Dietz et
al.
2009).移動の際には海洋上も飛翔し(Stansfield 1966;
Boshamer and Bekker
2008;Ahlen et al. 2009),1日で180 km
移動した例もある(Strelkov 1969).
日本では,2002 年に北海道礼文島で初めて確認された
(Satô and Maeda 2003)後,2005 年 12 月に青森県外ヶ浜
町,北海道羽幌町,北海道千歳市で相次いで確認された
が,いずれも単独個体であった(Kawai et al. 2010).これ
らはコウモリ類の移動時期である秋から冬にかけての日
本海側での発見であること,単独個体であること,遺伝
的にヨーロッパおよびロシア産個体との差が小さいこと
から,いずれも大陸からの迷行個体の可能性が指摘され
ている(Kawai et al. 2010).環境省によるレッドリスト
のランクでは,日本国内での生息・生態情報が極めて少
ないことから情報不足(DD)とされている(環境省 URL:
http://www.
env.go.jp/press/press.php?serial=8648;2011 年 12
月 1 日確認).
しかし,2011 年に北海道網走郡大空町で本種の出産哺
育コロニーが発見された.本報告では,その状況ならび
に,本種のエコーロケーションコール構造について記載
する.
調査地および方法
1.調査地
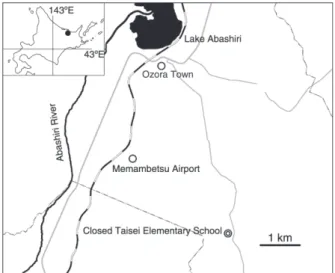
調査地は,北海道網走郡大空町の旧大成小学校敷地内
(北緯 43.85 度,東経 144.20 度)である.周囲は,農家
が点在する農耕地帯で,北北西 7.2 km に網走湖,6.7 km
に大空市街地が位置し,西 6.0 km に網走湖に注ぐ網走川
が流れている.また,北西 4.5 km に女満別空港がある
(Fig. 1).
敷地内には公民館と体育館(23.5 m×14.5 m)の建物
があり,時折周辺住民が使用している.コウモリは,体
育館の南東(高さ約 4.5 m,横幅約 7.5 m:Fig. 2)およ
び南西側の板壁に多数ある隙間から出巣していた.
2.捕獲調査および計測
調査地に生息するコウモリの種同定,ならびにねぐら
の利用状況を把握するため,2011 年に 5 回(6 月 24 日,
7 月 8 日,8 月 7 日,8 月 18 日,9 月 3 日)にわたって,
カスミ網(36 mm メッシュ)を使用して出巣個体の捕獲
調査を行った.また,7 月 10 日には出巣個体数を目視に
よって数えた.
捕獲した個体の性および齢を確認した後,翼開長を携
帯メジャー(Convex35, Uchida;最小目盛 1 mm),前腕長
および体重をデジタルノギス(19975,シンワ測定,新
潟;最小目盛 0.01 mm)と体重計(ハンディミニ 1476,
タニタ,東京;最小目盛 0.1 g)を使用して計測した.雄
の場合は,陰茎長も計測した.齢は,Brunet-Rossinni and
Wilkinson
(2009)に基づき,中手骨・第一指骨間の関節
の化骨化度合いを参考に成獣(前年以前生まれ)と幼獣
(当年生まれ)に区別した.雌成獣の場合は,腹部が明ら
かに膨らんでいるものを妊娠中,腹部は膨らんでいない
が乳頭が肥大化して周囲の毛が抜けているものを授乳中
とした.さらに,ヒナコウモリ(V. sinensis)との明らか
な違いである乳頭の数(Schober et al. 1993;Smith and Xie
2008;佐藤 2011)を確認した.調査期間中に拾得死亡個
体があった場合は仮剥製を製作し,頭骨の計測ならびに
歯式の確認も行なった.なお,本調査は,環境省鳥獣捕
獲許可第 11-112 号に基づいて実施された.
3.音声録音
2011 年 7 月 25 日に,ねぐらから出巣したコウモリの発す
るエコーロケーションコールを,タイムエキスパンジョン
式バットディテクター(D-240x, Pettersson Elektronik AB,
Uppsala, Sweden
)に接続した IC レコーダー(D-09,ロー
ランド(株),静岡県浜松市)を用いて録音した.その
際,ねぐらから約 50 m 離れた地点において,出巣した個
体を目視で追跡し,十分にねぐらから離れて飛翔して
いるコウモリにバットディテクターのマイク部分を向け
てエコーロケーションコールを録音した.本研究で用い
たバットディテクターでは,一回の録音で 3.4 秒分の音
を 10 倍に引き延ばして(34 秒)録音することができる.
1 回の録音で記録された連続した一連のパルスを,1
シークエンスとして扱った.録音した音声は,音声解析
ソフト(Bat Sound 1.31, Petersson Elektronik AB, Uppsala,
Sweden
)を 用 い て ソ ナ グ ラ ム 化 さ れ(FFT size 512,
Hanning window
),各シークエンスの中の明瞭に録音さ
れている連続した全てのパルスについて,最高周波数
(FMAX),最低周波数(FMIN),最大音圧(ピーク)周
波数(PF),パルス長(D),パルス間隔(IPI),バンド
幅(Bandwidth=最高周波数-最低周波数)が測定された.
4.統計処理
前腕長,翼開長の雌雄間および齢間の比較を,雌雄お
よび齢を独立変数,各サイズを従属変数とした 1 元配置
Fig. 1. Location of the study site. The area covered by the black circle in the inset has been magnified.
Fig. 2. A photograph of the building where the maternity colony of Vespertilio murinus was roosting.
分散分析によって行った.P<0.05 を有意水準とし,計
算には表計算ソフト Excel 2007 を用いた.
結 果
1.捕獲結果および種の同定
5 回の捕獲調査で合計 58 個体のコウモリを捕獲した.
そのうち 1 個体(8 月 7 日捕獲)は乳頭が一対のヒナコウ
モリであったが,他の個体は全て乳頭を 2 対有していた
(Fig. 3).
6 月 24 日に捕獲された 3 個体はすべて妊娠中の雌,7 月
8 日に捕獲された 3 個体はすべて授乳中の雌であった
(Fig. 4).なお,7 月 10 日には,夕方照度 3.8 Lx の時点
から 26 分間で,成獣と見られる個体が約 60 個体出巣し
ていた.一方で,8 月 7 日以降に捕獲された 51 個体のう
ちの 49 個体(96%)は幼獣であった(Fig. 4).
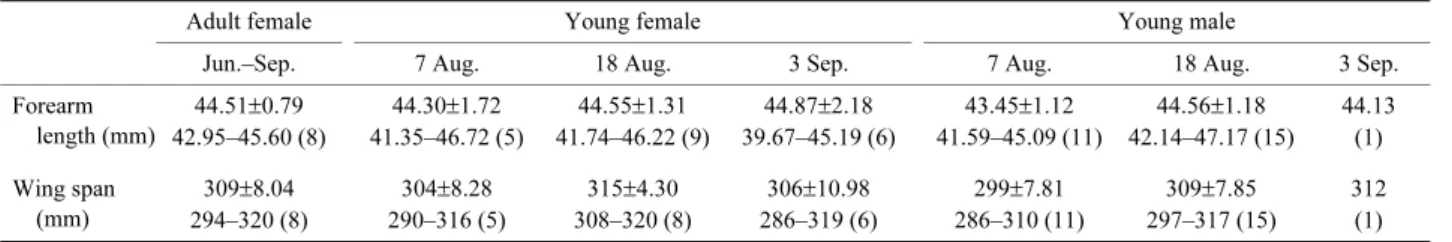
捕獲された全個体の前腕長は平均 44.06 mm で,雌成
獣と雌幼獣間ならびに幼獣の雌雄間には明瞭な差は見ら
れなかった(それぞれ F=0.30,P>0.05;F=0.54,P>
0.05)(Table 1).翼開長は平均 306.9 mm で,雌成獣と
雌幼獣間に有意差はなかった(F=0.12,P>0.05)が,
幼獣の雌雄間には有意差があった(F=4.59,P<0.05)
(Table 1).サンプル数が少なく統計解析はできなかった
ものの,成獣の体重は経産後に減少したが,その後は増
加傾向にあった.また,幼獣も同様に,8 月から 9 月に
かけて増加傾向にあった(Fig. 5).雄幼獣の陰茎の長さ
は,平均 8.1 mm(n=11)であった.
外部形態の特徴は,左右の耳介の基部が離れ,第 3 指
の第 2・3 指骨長は第 1 指骨長の 2 倍以下であり,第 5 指
の先端は第 4 指の第 1 指骨の先端に達していた.また,
鼻孔は筒状を呈しておらず,股間膜背面の毛の先端は黄
褐色であり,その分布は基部から 1/3 まであった.耳珠
幅が最大になる部位は耳珠の中間部より先端側であり,
耳介の最大幅は 8.5 mm であった.体色は,膜,耳介,顔
が,他の部分に比べて極端に黒かった.
2011 年 8 月 20 日と 12 月 15 日には,調査地および調査
地から南南東へ 2.2 km 離れた建物内で,8 月 18 日に標
識・放獣された死亡個体(2 頭)が拾得された.両個体
の外部計測値と頭骨の計測値を Table 2 に示す.歯式は
I
2/3+C1/1+P1/2+M3/3=32であり,特に前臼歯は上顎
に 1 対,下顎に 2 対で,吻部の上面両側には明らかに凹
みがあった.これらの標本は道東コウモリ研究会(根室
市)に保管されている(標本番号 NK0178,NK0180).
Table 1. Descriptive statistics of the external characteristics of captured bats in each season
Adult female Young female Young male
Jun.–Sep. 7 Aug. 18 Aug. 3 Sep. 7 Aug. 18 Aug. 3 Sep.
Forearm length (mm) 44.51±0.79 44.30±1.72 44.55±1.31 44.87±2.18 43.45±1.12 44.56±1.18 44.13 42.95–45.60 (8) 41.35–46.72 (5) 41.74–46.22 (9) 39.67–45.19 (6) 41.59–45.09 (11) 42.14–47.17 (15) (1) Wing span (mm) 309±8.04 304±8.28 315±4.30 306±10.98 299±7.81 309±7.85 312 294–320 (8) 290–316 (5) 308–320 (8) 286–319 (6) 286–310 (11) 297–317 (15) (1) Mean±SD values and ranges are shown. Numbers of measured individuals are given in parentheses.
Fig. 3. A photograph of an adult female of Vespertilio murinus. Two nipples on the left thorax are shown.
Fig. 4. Number of captured Vespertilio murinus in the closed Taisei Elementary School.
2.エコーロケーションコール構造
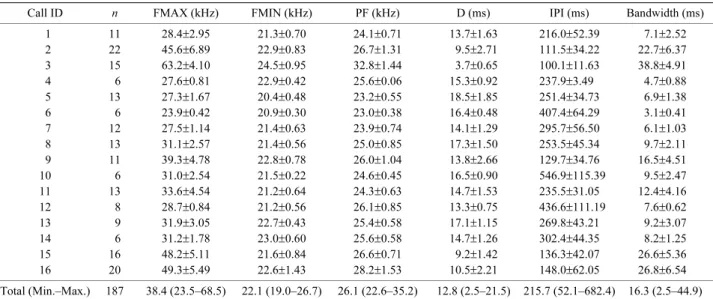
合計で 16 シークエンスのエコーロケーションコール
を録音することができた.また,各シークエンスで 6 ~
22 パルス,合計 187 パルスの測定を行なった(Table 3).
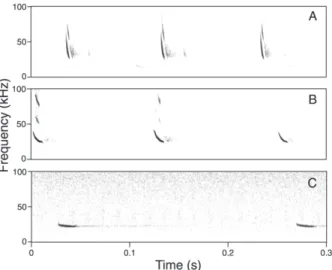
パルスの形状は,周波数が急激に変調する
FM(Fre-quency modulated
)成分と周波数が緩やかに変調する QCF
(Quasi-constant frequency)成分が組合わさった,いわゆ
る FM-QCF 型であった(Fig. 6).ただし,FM 成分と QCF
成分の幅の比率は,シークエンスごとに大きく異なって
いた(Table 3,Fig. 6).各パラメーターの平均値は,38.4
kHz
(最高周波数),22.1 kHz(最低周波数),26.1 kHz
(ピーク周波数),12.8 ms(パルス幅),215.7 ms(パル
ス間隔),16.3 kHz(バンド幅)であったが,最高周波
数,パルス長,パルス間隔,バンド幅は変異が大きかっ
た(Table 3,Fig. 6).
なお,本調査では,推定出産時期から 3 ~ 4 週間ほど
経った日にエコーロケーションコールの録音を行なって
いる.後述するように,ヨーロッパにおける幼獣の飛翔
開始時期が通常 34–35 日齢であることから,今回録音し
たエコーロケーションコールを発していた個体は成獣で
あると考えられる.
Fig. 5. Seasonal changes in the body weight of Vespertilio murinus between June and September.
Table 2. Measurements of the external and cranial characteristics of bats captured near the closed Taisei Elementary School
Table 3. Descriptive statistics for echolocation call sequences of Vespertilio murinus
Specimen Sex Age BW FAL WS GL BCL CM³ I¹M³ RW UMW ZW BCW MtdW BCH MdL I1M3
NK0178 M J 12.2 47.17 327.0 14.23 14.04 5.06 5.90 5.11 5.95 9.61 7.82 8.81 6.63 10.64 6.09 NK0180 F J 10.6 45.00 309.0 14.46 14.19 5.36 5.91 5.19 6.37 9.86 7.83 8.94 6.68 11.00 6.24 M, male; F, female; J, juvenile; BW, body weight; FAL, forearm length; WS, wing span; GL, greatest length of skull; BCL, condylobasal length; CM³, length of upper tooth row (from C to M³); I¹M³, length of upper tooth row (from I¹ to M³); RW, rostral width; UMW, width across upper molars; ZW, Zygomatic width; BCW, braincase width; MtdW, mestoid width; BCH, height of braincase; MdL, mandible length; I1M3, length of lower tooth
row (from I1 to M3). Units of all measurements except BW (g) are mm.
Call ID n FMAX (kHz) FMIN (kHz) PF (kHz) D (ms) IPI (ms) Bandwidth (ms)
1 11 28.4±2.95 21.3±0.70 24.1±0.71 13.7±1.63 216.0±52.39 7.1±2.52 2 22 45.6±6.89 22.9±0.83 26.7±1.31 9.5±2.71 111.5±34.22 22.7±6.37 3 15 63.2±4.10 24.5±0.95 32.8±1.44 3.7±0.65 100.1±11.63 38.8±4.91 4 6 27.6±0.81 22.9±0.42 25.6±0.06 15.3±0.92 237.9±3.49 4.7±0.88 5 13 27.3±1.67 20.4±0.48 23.2±0.55 18.5±1.85 251.4±34.73 6.9±1.38 6 6 23.9±0.42 20.9±0.30 23.0±0.38 16.4±0.48 407.4±64.29 3.1±0.41 7 12 27.5±1.14 21.4±0.63 23.9±0.74 14.1±1.29 295.7±56.50 6.1±1.03 8 13 31.1±2.57 21.4±0.56 25.0±0.85 17.3±1.50 253.5±45.34 9.7±2.11 9 11 39.3±4.78 22.8±0.78 26.0±1.04 13.8±2.66 129.7±34.76 16.5±4.51 10 6 31.0±2.54 21.5±0.22 24.6±0.45 16.5±0.90 546.9±115.39 9.5±2.47 11 13 33.6±4.54 21.2±0.64 24.3±0.63 14.7±1.53 235.5±31.05 12.4±4.16 12 8 28.7±0.84 21.2±0.56 26.1±0.85 13.3±0.75 436.6±111.19 7.6±0.62 13 9 31.9±3.05 22.7±0.43 25.4±0.58 17.1±1.15 269.8±43.21 9.2±3.07 14 6 31.2±1.78 23.0±0.60 25.6±0.58 14.7±1.26 302.4±44.35 8.2±1.25 15 16 48.2±5.11 21.6±0.84 26.6±0.71 9.2±1.42 136.3±42.07 26.6±5.36 16 20 49.3±5.49 22.6±1.43 28.2±1.53 10.5±2.21 148.0±62.05 26.8±6.54 Total (Min.–Max.) 187 38.4 (23.5–68.5) 22.1 (19.0–26.7) 26.1 (22.6–35.2) 12.8 (2.5–21.5) 215.7 (52.1–682.4) 16.3 (2.5–44.9) n, number of measured pulses; FMAX, maximum frequency; FMIN, minimum frequency; PF, frequency of maximum energy; D, duration; IPI, inter-pulse interval. Means±SD values are shown.
考 察
旧世界の温帯域において,2 対の乳頭を有するコウモリ
類は極めて稀で(Schober et al. 1993;Kunz and Hood 2000;
Smith and Xie
2008),日本国内でこれまでに確認された種
の中ではヒメヒナコウモリのみである.本研究で捕獲さ
れたコウモリは,一個体のヒナコウモリを除く個体全て
が 2 対の乳頭を有していた(Fig. 3)ことに加え,前腕長,
耳介の形状および幅,体色(膜,耳介,顔)の形態的特
徴や,拾得された幼獣 2 個体の外部計測値と頭骨の計測
値(Table 2)と歯式,吻部の上面両側には明らかに凹み
があったことから,ヒメヒナコウモリと同定される(Satô
and Maeda
2003;前田 2005;Kawai et al. 2010;佐藤 2011).
従って,今回確認されたコロニーは,日本国内で初記録
となるヒメヒナコウモリの出産哺育個体群である.
ヒメヒナコウモリは旧北区の温帯域以北に広く分布す
る種である.今回得られた計測データは,ヨーロッパ産
個体のもの[前腕長:39–49 mm,体重:9.8–23.0 g,翼
開長:270–310 mm,陰茎長:7–8 mm,頭骨基底全長:
14.0–16.0 mm(Schober et al. 1993;Rydell and Baagøe
1994)]と明瞭な違いは見られず,本種における形態の地
理的変異は少ないものと思われる.
ヒメヒナコウモリの前腕長は,野外飛翔を始める
34 ~ 35 日齢以降に成長が鈍化してほぼ成獣大に達する
(Kozhurina 1999).本報告では成獣と幼獣間に前腕長の差
は見られなかったが,これは,計測した幼獣がすべて飛
翔開始後であったためであると考えられる.また,Safi
(2006)は,スイスのヒメヒナコウモリで成体の前腕長の
雌雄間には有意差がないことを報告しており,本報告で
の結果と矛盾しない.体重について,ヒメヒナコウモリ
も含めた温帯域のコウモリ類では,分散期から越冬直前
にかけて増加することが知られている(Beasley et al.
1984;Rydell and Baagøe 1994).本報告で見られた 8 月か
ら 9 月にかけての体重増加も,越冬前の脂肪蓄積による
ものと考えられる.
今回のコロニーにおける出産時期は,捕獲の状況から
みて 6 月末から 7 月上旬と推定される.ヨーロッパにお
ける出産時期も同様に 6 月下旬から 7 月上旬であること
が知られている(Rydell and Baagøe 1994).コロニーの規
模(雌成獣の個体数)は,約 60 個体であり,これについ
てもヨーロッパから知られるコロニーの規模(30–140 個
体)と大きく変わらない(Blant and Jaberg 1995).
8 月 7 日以降に捕獲された個体のほとんどが幼獣で
あった(Fig. 4)ことから,この時期までには授乳期は終
わり,雌成獣はすでに分散を開始していたものと思われ
る.ヨーロッパにおけるヒメヒナコウモリの哺育期間は,
通常 34–35 日であり,早い個体では 25 日齢から飛翔を
開始する(Kozhurina 1999).本研究の結果は,日本に
おけるヒメヒナコウモリの哺育期間も同様に 30 日程度
であることを示唆する.また,ヒナコウモリと同様に
(Funakoshi and Uchida 1981),母獣は哺育が終了すると幼
獣より先に分散を始める傾向にあると考えられる.
本種のエコーロケーションコールは,典型的な
FM-QCF
構造を有していた(Fig. 6).海外産の本種のエコー
ロケーションコールも同様の構造を有しており,そのパ
ラメーター計測値についても明瞭な差は見られない
(Schaub and Schnitzler 2007).北海道内では,同様の構造
のエコーロケーションコールを発する種としてヤマコウ
モリ(Nyctalus aviator),ヒナコウモリ,キタクビワコウ
モ リ(Eptesicus nilssonii),ク ロ オ オ ア ブ ラ コ ウ モ リ
(Hypsugo alaschanicus)が 知 ら れ る(Fukui et al. 2004;
福井未発表).計測パラメーターのうち,比較的変異が
小さい PF に注目すると,ヒメヒナコウモリの平均 PF
(26.1 kHz)はキタクビワコウモリ(30.5 kHz)より低く,
ヒナコウモリ(24.2 kHz)よりもわずかに高い傾向があ
る.一般的に,コウモリ類では体サイズの大きな種はピー
ク周波数が低い超音波音声を発する傾向にある(特に同
属や近縁種において)が(例えば,Barclay and Brigham
1991;Bogdanowicz et al. 1999;Jones 1999;Holderied and
von Helversen
2003),本結果もこの傾向を支持するもの
である.なお,捕獲調査でヒナコウモリが 1 個体確認さ
れているため,今回録音された音声の中にヒナコウモリ
のものが含まれている可能性を否定できないが,捕獲数
Fig. 6. Sonograms of Vespertilio murinus that seem to be flying close to obstacles (A), some distance from obstacles (B) and in open spaces (C).
の比率から,その可能性は極めて低いと考えられる.
本調査で録音されたパルスは前述のように FM-QCF 構
造を有していたが,FM 成分と QCF 成分の幅の比率は,
シークエンスごとに異なっていた(Fig. 6).これは,録
音されたパルスを発していた個体が飛翔していた環境の
違いによると考えられる.FM 成分は障害物に近い空間,
QCF
成分は開けた空間で対象物を探索・定位するのに適
しており,FM-QCF 構造の音声を利用するコウモリは,
障害物までの距離に応じて FM 成分と QCF 成分の比率を
変化させて利用していることが知られている(Schnitzler
and Kalko
2001).ドイツのヒメヒナコウモリにおいても,
障害物までの距離に応じて本研究で見られたような音声
構 造 の 変 化 が あ る こ と が 分 か っ て い る(Schaub and
Schnitzler
2007).本調査地敷地内および隣接地には,ね
ぐらとなっている体育館の他に,公民館や水道施設の建
物や樹木もあり,それらの近くを飛翔した際により FM
成分が多い音声(Fig. 6A)を発していたと思われる.
日本における本種の記録は,本研究以前は 4 個体が拾
得されたのみで,いずれも大陸からの迷行個体の可能性
が高いと思われていた(Kawai et al. 2010).しかし,本研
究結果から,日本国内での出産哺育が明らかになった.
また,過去の 4 例や,今回死体として拾得された 2 個体
のうち 1 個体が秋から冬にかけてのものであることか
ら,日本国内で越冬している可能性が高い.ヒメヒナコ
ウモリは,1,000 km を超える長距離移動をすることが知
られており(Masing 1989;Dietz et al. 2009),国内におけ
る分布域が北海道および青森県に限らない可能性も考え
られる.
本種は外部形態がヒナコウモリと酷似しており,捕獲
を行なって乳頭の数を確認しなければ種の同定は難し
く,特に雄では乳頭が痕跡しかない(近藤ほか 2012).数
年前まではヒメヒナコウモリが国内に生息しているとは
考えられておらず,日本産コウモリ類の種同定では乳頭
数が考慮されることはなかった.さらに,今回の捕獲調
査でヒナコウモリ 1 頭が本種に混じって捕獲されたこと
は,本種がヒナコウモリと混群を形成する可能性がある
ことを示唆する.従って,ヒメヒナコウモリを捕獲した
にも関わらず,ヒナコウモリと誤同定されていた可能性
も十分に考えられるため,今後は細心の注意が必要であ
る.日本における本種の現状を明らかにするためにも,よ
り詳細な分布・生態・分類に関する調査研究が待たれる.
謝 辞
調査にあたって,以下の方々に御協力頂いた.深く感
謝申し上げる.秋葉暢康(大空町役場),石原新太郎(大
空町),葛木いく(大空町),河合久仁子(北海道大学北
方生物圏フィルド科学センター),佐藤雅彦(利尻博物
館),須貝昌太郎(本庄市),鈴木 薫(大空町朝日公民
館長),高塚慎太朗(網走市),武田令子(根室市歴史と
自然の資料館),中島宏章(札幌市),橋本幸太郎(網走
市),原田博之(大空町大成公民館長),森 利博(根室
市)(50 音順,敬称略).
引 用 文 献
Ahlen, I., Baagøe, H. J. and Bach, L. 2009. Behavior of Scandinavian bats during migration and foraging at sea. Journal of Mammalogy 90: 1318–1323.
Baagøe, H. J. 1986. Summer occurrence of Vespertilio murinus Linne–1758 and Eptesicus serotinus (Schreber–1780) (Chiroptera, Mammalia) on Zealand, Denmark, based on records of roosts and registrations with bat detectors. Annual Naturhistorisches Museum Wien 88/89: B281–B291.
Baagøe, H. J. 2001. Danish bats (Mammalia: Chiroptera): Atlas and analysis of distribution, occurrence and abundance. Steenstrupia 26: 1–117.
Baranauskas, K., Grikieniene, J. and Masing, M. 2006. Particoloured bat Vespertilio murinus (Chiroptera) found hibernating in Lithua-nia for the first time. Ekologija 4: 31–33.
Barclay, R. M. R. and Brigham, R. M. 1991. Prey detection, dietary niche breadth, and body size in bats: why are aerial insectivorous bats so small? The American Naturalist 137: 693–703.
Beasley, L. J., Pelz, K. M. and Zucker, I. 1984. Circannual rhythms of body weight in pallid bats. American Journal of Physiology 246: R955–R958.
Blant, J-D. and Jaberg, C. 1995. Confirmation of the reproduction of Vespertilio murinus L., in Switzerland. Myotis 32–33: 203–208. Bogdanowicz, W., Fenton, M. B. and Daleszczyk, K. 1999. The
rela-tionships between echolocation calls, morphology and diet in insectivorous bats. Journal of Zoology, London 247: 381–393. Boshamer, J. P. C. and Bekker, J. P. 2008. Nathusius’ pipistrelles
(Pipistrellus nathusii) and other species of bats on offshore plat-forms in the Dutch sector of the North Sea. Lutra 51: 17–36. Brunet-Rossinni, A. K. and Wilkinson, G. S. 2009. Methods for age
estimation and the study of senescence in bats. In (T. H. Kunz and S. Parsons, eds.) Ecological and Behavioral Methods for the Study of Bats, pp. 315–325. The Johns Hopkins University Press, Baltimore.
Dietz, C., von Helversen, O. and Nill, D. 2009. Bats of Britain, Europe and Northwest Africa. A and C Black Publishers Ltd., London, 400 pp.
Fukui, D., Agetsuma, N. and Hill, D. A. 2004. Acoustic identification of eight species of bat (Mammalia: Chiroptera) inhabiting forests of southern Hokkaido, Japan: potential for conservation monitor-ing. Zoological Science 21: 947–955.
breeding season and postnatal growth in the Namie’s frosted Bat, Vespertilio superans superans. Japanese Journal of Ecology 31: 67–77.
Holderied, M. W. and von Helversen, O. 2003. Echolocation range and wingbeat period match in aerial-hawking bats. Proceedings of the Royal Society of London, Series B 270: 2293–2299. Jones, G. 1999. Scaling of echolocation call parameters in bats.
Journal of Experimental Biology 202: 3359–3367.
Kawai, K., Fukui, D., Satô, M., Harada, M. and Maeda, K. 2010. Vespertilio murinus Linnaeus, 1758 confirmed in Japan from morphology and mitochondrial DNA. Acta Chiropterologica 12: 463–470.
近藤憲久・中島宏章・倉野翔史・黒澤春樹.2012.ヒメヒナコ ウモリ(Vespertilio murinus)の雄の識別点.根室市歴史と 自然の資料館紀要 24: 61–63.
Kozhurina, E. I. 1999. Early postnatal ontogeny and main periods of the development in parti-coloured bats Vespertilio murinus (Chiroptera: Vespertilionidae). Folia Zoologica 48:33–48. Kunz, T. H. and Hood, W. R. 2000. Parental care and postnatal growth
in the Chiroptera. In (E. G. Crichton and P. H. Krutzsch, eds.) Reproductive Biology of Bats, pp. 415–468. Academic Press, Cambridge.
前田喜四雄.2005.日本産翼手目(コウモリ)検索表.日本の 哺乳類改訂版(阿部 永,監修),pp. 159–169.東海大学出 版会,秦野.
Masing, M. 1989. A long-distance flight of Vespetilio murinus from Estonia. Myotis 27: 147–150.
Orlova, M. V. 2011. Ectoparasite associations of bats from the Urals (Russia). Hystrix, the Italian Journal of Mammalogy 22: 105– 110.
Rakhmatulina, I. K. 1995. Bat’ attachment to different shelters in the eastern Transcaucasia. Myotis 32-33: 197–202.
Rydell, J. and Baagøe, H. J. 1994. Vespertilio murinus. Mammalian Species 467: 1–6+3 figs.
Safi, K. 2006. Die Zweifarbfledermaus in der Schweiz—Status und Grundlagen für den Schutz. Haupt Berne, Bern, 100 pp. 佐藤雅彦.2011.ヒメヒナコウモリ.コウモリ識別ハンドブッ
ク改訂版(佐野 明・福井 大,監修),p. 34.文一総合出 版,東京.
Satô, M. and Maeda, K. 2003. First record of Vespertilio murinus Lin-naeus, 1758 (Vespertilionidae, Chiroptera) from Japan. Bulletin of the Asian Bat Research Institute 3: 10–14.
Schaub, A. and Schnitzler, H-U. 2007. Echolocation behavior of the bat Vespertilio murinus reveals the border between the habitat types “edge” and “open space”. Behavioral Ecology and Socio-biology 61: 513–523.
Schnitzler, H-U. and Kalko, E. K. V. 2001. Echolocation by insect-eating bats. BioScience 51: 557–569.
Schober, W., Grimmberger, E. and Stebbings, R. E. 1993. Bats of Britain and Europe. Hamlyn Publishing Group Limited, London, 224 pp.
Smith, A. T. and Xie, Y. 2008. A guide to the Mammals of China. Princeton University Press, Princeton, 544 pp+60 Pl.
Stansfield, G. 1966. Parti-coloured bat Vespetilio murinus L. from a North Sea drilling rig. Journal of Zoology, London 150: 491– 492.
Strelkov, P. P. 1969. Migratory and stationary bats (Chiroptera) of the European part of the Soviet Union. Acta Zoologica Cracoviensia 16: 393–440+4 figs.
Suba, J., Vietniece, D. and Petersons, G. 2010. The parti-coloured bat Vespertilio murinus in Riga (Latvia) during autumn and winter. Environmental and Experimental Biology 8: 93–96.
Syvertsen, P. O., Stormark, T. A., Nordseth, M. and Starholm, T. 1995. A tentative assessment of bat diversity and distribution in Norway. Myotis 32–33: 183–191.
Tiunov, M. P. 1997. Rukokrylye Dal’nego Vostoka Rossii. Dal’nauka, Vladivostok, 134 pp. (in Russian).
ABSTRACT
A maternity colony of Vespertilio murinus in Ozora, Abashiri District, Hokkaido
Norihisa Kondo
1,*, Dai Fukui
2, Syohji Kurano
3and Haruki Kurosawa
41Nemuro Museum of History and Nature, Hanasaki-minato 209, Nemuro, Hokkaido 087-0032, Japan
2Division of Animal Resources, National Institute of Biological Resources, Environmental Research Complex, Gyeoungseo-dong, Seo-gu, Incheon
404-708, Korea
3Faculty of Bio-Industry, Tokyo University of Agriculture, Yasaka 196, Abashiri, Hokkaido 099-2493, Japan 4Dohtoh Bat Research Group, Motomachi 38-54, Bihoro, Hokkaido 092-0063, Japan
*E-mail: [email protected]
A maternity roost of bats was found in a closed elementary school in Ozora, Abashiri District, Hokkaido. After measurements
of their external and cranial characteristics and the number of nipples, the bats in the colony were identified as Vespertilio murinus.
In July, about 60 adult females emerged from the roost. According to the results of a capture survey, the adult female gave birth
in late June and early July. In August, almost all of the members of the colony were volant young, and the adult females had
already dispersed. The echolocation call of flying V. murinus had a typical frequency-modulated/quasi-constant frequency (FM/
QCF) structure (mean peak frequency=26.1 kHz), although the ratio between FM and QCF components varied with the flying
spaces of the recorded individuals. To our knowledge, this is the first time that a roost of V. murinus has been found in Japan,
and we believe that V. murinus is a resident species in Japan.
Key words: Hokkaido, Ozora Town, Vespertilio murinus, maternity colony, echolocation call
受付日:2012 年 2 月 20 日,受理日:2012 年 5 月 28 日
著 者:近藤憲久,〒087-0032 根室市花咲港209番地 根室市歴史と自然の資料館 [email protected] 福井 大,Division of Animal Resources, National Institute of Biological Resources, Environmental Research Complex, Gyeoungseo-dong, Seo-gu, Incheon 404-708, Korea
倉野翔史,〒099-2493 網走市八坂196 東京農業大学産業生物学部動物生産管理学研究室 黒澤春樹,〒092-0063 美幌町元町38-54 道東コウモリ研究会