Sirt1 Overexpression in POMC Neurons Partially Rescues the Obese Phenotype Induced by Insulin-resistant Constitutively-nuclear FoxO1 in Male Mice.
POMC ニューロンにおける Sirt1 過剰発現は、インスリン抵抗性核内滞留型 FoxO1 に よる体重増加を改善する
指導教員 北村 忠弘 教授 平成25年12月作成
群馬大学大学院医学系研究科 平成22年度入学
高次機能統御系 細胞形態・情報統御学 代謝シグナル解析分野
Vina Yanti Susanti
学
位
論文Sirt1 Overexpression in POMC Neurons Partially Rescues the Obese Phenotype Induced by Insulin-resistant Constitutively-nuclear FoxO1 in Male Mice.
POMC ニューロンにおける Sirt1 過剰発現は、インスリン抵抗性核内滞留型 FoxO1 に よる体重増加を改善する
指導教員 北村 忠弘 教授 平成25年12月作成
群馬大学大学院医学系研究科 平成22年度入学
高次機能統御系 細胞形態・情報統御学 代謝シグナル解析分野
Vina Yanti Susanti
Thesis
Sirt1 Overexpression in POMC Neurons Partially Rescues the Obese Phenotype Induced by Insulin-resistant Constitutively-nuclear FoxO1 in Male Mice
Gunma University Graduate School of Medicine
Vina Yanti Susanti
Contents
Contents 1
Abbreviation 2
Introduction 3-4
Materials and methods 5-7
Results 8-10
Discussion 11-13
Summary 14
Acknowledgments 15
References 16-20
Legends and Figure 21-25
Submitted paper 26
Abbreviation
Sirt1: Sirtuin 1
FoxO1: Forkhead box O1
CN-FoxO1: Constitutively-nuclear FoxO1 POMC: Proopiomelanocortin
AgRP: Agouti-related protein NPY: Neuropeptide Y
α-MSH: α-melanocyte stimulating hormone MC3R: Melanocortin type-3 receptor MC4R: Melanocortin type-4 receptor IRS: Insulin receptor substrate IGF-I: Insulin-like growth factor 1 KI: Knock in
MEFs: Mouse embryonic fibroblasts MOI: Multiplicity of infection PCR: Polymerase chain reaction
SDS-PAGE: Sodium dodecyl sulfate polyacrylamide gel electrophoresis PBS: Sodium dodecyl sulfate polyacrylamide gel electrophoresis
PFA: Paraformaldehyde ARC: Arcuate nucleus
RER: Respiratory exchange rate
Introduction
Worldwide, the prevalence of obesity has nearly doubled since 1980. In the United States, this trend is especially alarming in children and adolescents.
According to the Centers for Disease Control, 18.1% of adolescents (12–19 years old) in the United States were considered obese in 2008, 3-fold more than in the 1980s (1). This alarming increase in the worldwide incidence of obesity is also associated with metabolic pathologies, including type-2 diabetes and heart disease.
Intense research efforts have focused on understanding the mechanisms underlying these disorders and on developing new treatment options. Despite the many new treatment options, obesity-related conditions have become the leading causes of global mortality and morbidity, emphasizing the need for more effective therapeutic and preventive measures (2).
Obesity results from an imbalance between energy intake and energy expenditure. The hypothalamus is the key brain region for controlling energy balance regulation as it integrates information about an individual’s nutritional state and the changing energy requirements of the peripheral tissues (3). The arcuate nucleus of the hypothalamus is the primary center for energy balance regulation, and it contains two types of neurons that are related to energy homeostasis:
orexigenic neuropeptide Y (NPY) and agouti-related protein- (AgRP-) containing neurons and proopiomelanocortin (POMC) and cocaine- and amphetamine- regulated transcript-containing neurons (3, 4). POMC neurons secrete, in addition to other neuropeptides, -melanocyte stimulating hormone (-MSH), which suppresses food intake and increases energy expenditure by activating the melanocortin type-3 receptor (MC3R) and the melanocortin type-4 receptor (MC4R) (5). AgRP is an inverse agonist for MC3R and MC4R that increases food intake and suppresses energy expenditure. POMC, AgRP, and MC3R/MC4R comprise the central melanocortin system, and genetic defects in this system are the most common defects in the monogenic form of obesity in rodents and humans (6-8).
Insulin signaling in the hypothalamus also regulates the energy balance. A lack of insulin receptors or insulin receptor substrate (IRS) 2 in the central nervous system increases food intake and body weight (9-11). The forkhead box-containing
protein of the O subfamily (FoxO)1 is a downstream transcription factor for insulin/IGF-I signaling (12). FoxO1 in the hypothalamus increases food intake via trans-activation of Agrp and/or Npy gene expression and via repression of Pomc gene expression (13, 14). A POMC neuron-specific phosphorylation mutant, constitutively-nuclear FoxO1 (CN-FoxO1) knock-in (KI) mice, exhibits mild obesity and hyperphagia (15). Conversely, Foxo1 ablation in POMC neurons results in decreased food intake, partly by increasing the expression of Cpe (which encodes -MSH), leading to increased -MSH production (16, 17).
Sirt1 is a protein deacetylase, and its targets include histones, transcription factors, and cofactors (18). Sirt1 improves insulin sensitivity by repressing protein tyrosine phosphatase-1B expression and by deacetylating IRS-2 in hepatocytes (19).
We reported previously that Sirt1 deacetylates FoxO1, enhances FoxO1 ubiquitination, and decreases FoxO1 protein levels (20). We also studied the roles of FoxO1 and Sirt1 in the mediobasal hypothalamus by stereotaxic microinjection of adenovirus-encoded Sirt1 and FoxO1 and found that Sirt1 can suppress hyperphagia and weight gain in the insulin-resistant CN-FoxO1 mutant (21).
However, the effects of FoxO1 and Sirt1 overexpression were not assessed either long-term or in a cell-type specific manner due to the limitations of the stereotaxic injection method. In a separate study, we reported that POMC neuron-specific Sirt1 overexpression prevents age-associated weight gain by stimulating energy expenditure in male mice (22). Therefore, in order to test the long-term effects of Sirt1 overexpression in POMC neurons, we asked whether the obese phenotype caused by the overexpression of CN-FoxO1 in POMC neurons could be rescued by Sirt1 overexpression. Toward this end, we analyzed POMC neuron-specific CN- FoxO1/Sirt1 double knock-in (DKI) mice.
Materials and Methods Animal Studies
Pomc-Cre mice and Rosa26CN-Foxo1 mice containing FLAG-tagged CN-FoxO1 inserted into the Rosa26 locus with a loxP-flanked stop sequence were described previously (15). Rosa26 Sirt1-WT mice were also described previously (22). DKI mice were generated by mating Pomc-Cre; Rosa26CN-Foxo1/+ mice with Pomc-Cre;
Rosa26Sirt1-WT/+ mice. Mice were housed in individual cages in a temperature- controlled facility with a 12-h light, 12-h dark cycle and with free access to water and normal chow (60% kcal of carbohydrate, 15% kcal of fat, and 25% kcal of protein) until the experiments. All animal care and experimental procedures were approved by the Institutional Animal Care and Use Committee at Gunma University. Body weight was measured weekly. Food intake was measured when mice were 25 and 26 weeks old. The average values were reported for body weight and food intake.
Cell culture, transfection, and adenovirus infection
Mouse embryonic fibroblasts (MEFs) were prepared as described previously (23).
Hypothalamic N41 cells (Cosmo Bio, Tokyo, Japan) and MEFs were maintained in high-glucose DMEM supplemented with 10% FBS. Transient transfection was performed using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. Adenovirus infection was performed using a standard protocol at an MOI of 50. The cells were harvested 24 h after transfection or infection.
RNA isolation and real time RT-PCR
We isolated RNA from tissue using RNAiso Plus (TAKARA, Otsu, Japan), and cDNAs were generated using the Improm II Reverse Transcription System (Promega, Madison, WI, USA). The generated cDNAs were subjected to real time PCR using the Light Cycler system and Light Cycler 480 SYBR Green I (Roche, Tokyo, Japan).
The primer sequences are listed in Supplemental Table 1. All results were normalized using -actin. We carried out each reaction in triplicate using a standard curve with the relevant cDNA for each primer set.
PCR genotyping
We extracted genomic DNA from tail samples using phenol/chloroform and genotyped them using Rosa26 genotyping PCR with ExTaq polymerase (TAKARA) as described previously (24). The primer sequences are listed in Supplemental Table 1.
For PCR identification of the Rosa26Sirt1-WT locus and the Rosa26CN-Foxo1 locus, the
Sirt1-1918F and M13F primer set and the Flag S1 and SE 1 primer set were used, respectively.
Western blot analyses
Proteins from cell lysates or MEFs were separated by SDS-PAGE, transferred to nitrocellulose membranes (Pall Life Sciences, Dreieich, Germany), and blotted with antibodies. The antibodies used in this study are listed in Supplemental Table 2.
Immunoreactive proteins were assessed using the LAS-4000 Image analyzer (Fuji Film, Tokyo, Japan) and densitometry using Image-J software.
Preparation of lysate from cell culture
Total cell lysates and nuclear lysates were prepared using lysis buffer supplemented with a Complete Mini Protease Inhibitor Cocktail tablet (Roche), 40 µM MG132 (benozyloxycarbonyl-L-leucyl-L-leucyl-L-leucinal; Sigma-Aldrich, St. Louis, MO, USA), and 0.25 µg/ml ubiquitin aldehyde (Peptide Institute, Minoh, Japan). To prepare cytoplasmic and nuclear lysates, cells were first washed with ice-cold PBS.
Ice-cold cytoplasmic lysis buffer (5 mM PIPES (KOH) pH 8.0, 65 mM KCl, and 0.5% NP 40) was added, the cells were scraped off the dish using a cold plastic cell scraper, and the cell suspension was gently transferred into a pre-cooled microfuge tube. After constant agitation for 30 minutes at 4°C, samples were pelleted in a microcentrifuge at 4°C. The supernatant, which was the cytoplasmic lysate, was removed to a fresh tube on ice. Nuclear lysis buffer was added to the pellet. After constant agitation for 30 minutes at 4°C, the samples were centrifuged in a microcentrifuge at 4°C, and the supernatant, which was the nuclear lysate, was removed to a fresh tube.
Immunohistochemistry
For immunofluorescence analysis of samples from the hypothalamus, mice were transcardially perfused with saline followed by 4% paraformaldehyde (PFA) in 0.1 M phosphate-buffered saline (pH 7.4). The brains were dissected into 4% PFA at 4°C, soaked in 30% sucrose overnight, and frozen in OCT. The sections were washed in PBS (3 x 5 minutes) to quench endogenous peroxidase activity and then stained with anti-POMC antibody. Pictures were taken, and the number of neurons that were positive for POMC immunostaining were counted and marked digitally using Adobe Photoshop to prevent re-counting. The results are expressed as the number of POMC- positive, nucleus-containing cell bodies per hemisection (the average of the left and the right) or the sum of these cells in all five counted sections of the ARC.
Energy metabolism
Oxygen (O2) consumption and carbon dioxide (CO2) production were measured in individual 7-month-old mice using an Oxymax apparatus (Columbus Instruments, Columbus, OH, USA). Animals were maintained on 12-h light, 12-h dark cycle beginning at 0600 and 1800 h, respectively. O2 and CO2 were measured every 18 min for each animal over a period of 3 days. Total O2 consumption was calculated as the accumulated O2 uptake for each mouse. The respiratory exchange rate (RER) was calculated as VCO2/VO2. Heat generation was calculated as caloric value (CV) x VO2; CV=3.815 +1.232 x RER. Locomotor activity was measured using the ACTIMO-100 (Shinfactory, Fukuoka, Japan).
Results
Sirt1 overexpression in POMC neurons rescues the obese phenotype caused by CN- FoxO1 in male but not female mice.
In order to test the long-term effects of Sirt1 and CN-FoxO1 overexpression in POMC neurons, we asked whether the obese phenotype caused by the overexpression of CN-FoxO1 in POMC neurons could be rescued by Sirt1 overexpression by analyzing POMC neuron-specific CN-FoxO1/Sirt1 DKI mice. Consistent with previous studies, we found that CN-FoxO1 KI mice had an obese phenotype and that male Sirt1 KI mice had a lean phenotype (15, 21, 22). The DKI mice showed rescue of the obese phenotype starting at 12 weeks of age. This effect plateaued from 18 weeks to 21 weeks of age and was seen again from 22 weeks to 27 weeks of age (Fig.
1A). The increase in body weight in CN-FoxO1 KI male mice was accompanied by increases in body length and adiposity, and these phenotypes were partially rescued in DKI mice (Fig. 1B-D).
The rescue of CN-FoxO1-induced obesity by Sirt1 overexpression in POMC neurons was associated with the effect on energy expenditure
Because the rescued phenotype was observed only in male mice, we performed subsequent analyses using only data from male mice. In order to investigate the cause of the obese phenotype and the mechanisms underlying rescue of the phenotype in these mice, we analyzed food intake, oxygen consumption, and locomotor activity. Daily food intake was not different in the examined groups (Fig.
2A). Oxygen consumption (during light and dark phases and for 24 h) tended to be decreased in CN-FoxO1 KI mice, and this was partially rescued in DKI mice (Fig.
2B). The locomotor activity did not differ in the groups studied (Fig. 2C). These results suggest that the rescue of CN-FoxO1-induced obesity by Sirt1 overexpression in POMC neurons could be explained by an effect on energy expenditure. To elucidate the molecular mechanism underlying increased basal metabolism, we measured the expression levels of mitochondrial-related genes (Ucp1 and Pgc- in brown adipose tissue (BAT). Although the Ucp1 expression levels were not significantly altered, expression of Pgc- tended to be increased in DKI mice compared to CN-FoxO1 KI mice (Fig. 2D, E).
Sirt1 overexpression rescues the effect of CN-FoxO1 on the number of POMC neurons.
FoxO1 has been shown to regulate the number of POMC neurons (25).
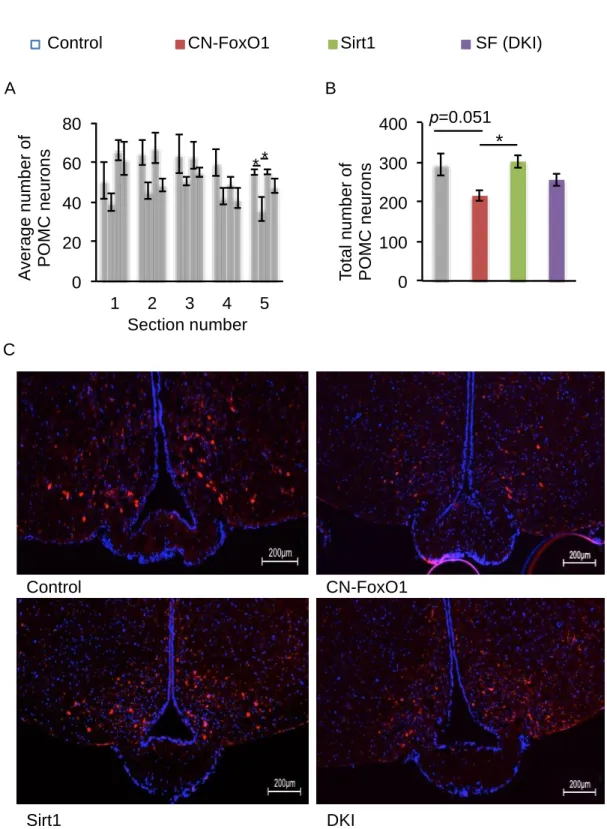
Therefore, we tested whether Sirt1 overexpression rescued the effect of CN-FoxO1 overexpression on the number of POMC neurons by counting the number of POMC neurons that showed positive immunostaining in five sections that spanned the arcuate nucleus of the hypothalamus where POMC neurons are located. Contrary to previous studies (25), we found that the number of POMC neurons (both the average number and the total number) tended to be decreased in CN-FoxO1 KI mice; this was partially rescued in DKI mice (Fig. 3A, B). Sirt1 by itself did not have any significant effect on the number of POMC neurons, which is consistent with our previous finding (22). No obvious morphological change in POMC neurons was observed (Fig. 3C). Therefore, Sirt1 overexpression in POMC neurons rescued the obese phenotype caused by CN- FoxO1, possibly by normalizing the effect of CN-FoxO1 on the number of POMC neurons.
Sirt1 overexpression decreases FoxO1 acetylation and protein levels without affecting the nuclear localization of CN-FoxO1.
We reported previously that deacetylation of the FoxO1 protein affects its stability by promoting poly-ubiquitination of FoxO1 (20). Insulin signaling promotes the nuclear exclusion of FoxO1 through Akt-mediated phosphorylation of FoxO1;
since CN-FoxO1 is the phosphorylation mutant of these Akt substrate sites, this mutant leads to insulin-resistance (15). However, it has been reported that the acetylation status of FoxO1 can override the nuclear/cytoplasmic localization signals conferred by phosphorylation: phosphorylated FoxO1 can be retained in the nucleus by deacetylation, and dephosphorylated FoxO1 can be targeted to the cytoplasm through acetylation (26). Therefore, Sirt1 overexpression could promote CN-FoxO1 deacetylation and affect either protein stability and/or the nuclear localization of CN- FoxO1. In order to address these questions, we performed in vitro experiments using hypothalamic N41 cells and MEFs to assess the effect of Sirt1 overexpression on FoxO1 acetylation, protein level, and subcellular localization. We found that Sirt1 overexpression in hypothalamic N41 cells promoted FoxO1 deacetylation and decreased the CN-FoxO1 protein level without affecting its nuclear localization (Fig.
4A-C). We also confirmed that the degree of Sirt1 overexpression achieved in MEFs via infection with adenovirus encoding the Cre recombinase was sufficient to
decrease the level of FoxO1 protein in the nucleus of DKI MEFs without affecting the FoxO1 protein level in the cytoplasm (Fig. 4D). We conclude that Sirt1 overexpression decreases FoxO1 acetylation and the FOXO1 protein level without affecting the nuclear localization of CN-FoxO1. The may explain in part why Sirt1 overexpression rescues the obese phenotype caused by CN-FoxO1 overexpression in POMC neurons.
Discussion
In order to investigate the long-term effects of Sirt1 and CN-FoxO1 overexpression in POMC neurons, we asked whether the obese phenotype caused by overexpression of CN-FoxO1 in POMC neurons could be rescued by Sirt1 overexpression by analyzing POMC neuron-specific CN-FoxO1/Sirt1 DKI mice. We found that Sirt1 overexpression in POMC neurons rescued the obese phenotype caused by CN-FoxO1 in male but not female mice. We thus focused our analyses on male mice. We found that the rescue of CN-FoxO1-induced obesity by Sirt1 overexpression in POMC neurons was associated with an effect on energy expenditure and that Sirt1 overexpression rescued the effect of CN-FoxO1 on the number of POMC neurons. These findings suggest possible mechanisms to explain the observed phenotypes in male mice. We also confirmed that Sirt1 overexpression decreases FoxO1 acetylation and the FoxO1 protein level without affecting the nuclear localization of CN-FoxO1, which provides a molecular mechanism: Sirt1 overexpression in POMC neurons partially rescued the obese phenotype induced by insulin-resistance due to CN-FoxO1 in male mice by decreasing the amount of overexpressed CN-FoxO1.
Overexpression of Sirt1 rescued the obese phenotype caused by CN-FoxO1 overexpression in male mice but not in female mice. The obese phenotype induced by CN-FoxO1 was not significant in the female mice in our study; in contrast, it was observed in both male and female mice in the previous study (15). This may be due to differences in the genetic background of the mice in the two studies. Furthermore, Sirt1 overexpression in POMC neurons had a lean effect that was observed only in male mice in our previous study (22). Sex hormones may differentially regulate FoxO1, Sirt1, and/or POMC neurons in male and female mice, but further studies will be needed to address this issue.
In the current study, the effect of Sirt1 overexpression on body weight appeared to be due to an effect on energy expenditure in the mice. POMC neurons and AgRP neurons send fibers to similar target neurons, such as to the paraventricular nucleus of the hypothalamus; the MC4R is crucial for mediating the regulation of food intake via these neurons (27). However, some of these targets are directly innervated only by POMC neurons in the ARC, such as the sympathetic preganglionic neurons of the intermediolateral nucleus of the thoracic spinal cord (28, 29).
Postganglionic neurons that innervate the interscapular brown adipose tissue receive
projections from the intermediolateral nucleus (30). Re-expression of MC4R in cholinergic neurons normalizes energy expenditure without affecting food intake (31), providing a pathway by which POMC neurons can regulate energy expenditure. In fact, Sirt1 overexpression in POMC neurons enhances sympathetic tone in the adipose tissue of male mice (22). Therefore, the results of this study are consistent with previous findings that POMC neurons exert stronger effects on energy expenditure than on food intake in terms of regulating the energy balance.
FoxO1 regulates the number of POMC neurons, and POMC neuron-specific FoxO1 knock-out mice have 23% fewer POMC neurons than controls (25). Contrary to our expectations, POMC neuron-specific CN-FoxO1 KI mice had fewer POMC neurons than control mice, and this reduction was rescued in DKI mice (Fig. 3). One possible explanation for the differing results is that the genetic background of the mice differs in the two studies. Another possibility is that the loss of function (FoxO1 knock-out) and the gain of function (CN-FoxO1 KI) differentially affect the FoxO1 target genes. Because FoxO1 regulates both cell proliferation and survival (20, 32), the lack of FoxO1 may promote POMC neuronal cell death, whereas overexpression of CN-FoxO1 may suppress the proliferation of POMC neurons and/or their progenitors.
Deacetylation of FoxO1 is reported to promote nuclear retention of FoxO1, thereby promoting its activity as a transcription factor (26). We reported previously that the deacetylation of FoxO1 destabilizes the protein by promoting degradation through the ubiquitin-proteasome system (20). Therefore, in order to test the net effect of FoxO1 deacetylation by Sirt1 in our mice, we tested the effect of Sirt1 overexpression in hypothalamic N41 cells and in MEFs infected with adenovirus encoding the Cre recombinase. Our results indicated that the degree of Sirt1 overexpression achieved in our model system decreased the CN-FoxO1 protein level, leading to the rescue of phenotypes caused by the overexpression of CN-FoxO1 in POMC neurons.
Based on these results, we conclude that Sirt1 overexpression in POMC neurons partially rescues the obesity phenotype caused by CN-Foxo1 by decreasing FoxO1 acetylation and FoxO1 protein levels. Further, Sirt1 can partially reverse obesity caused by central insulin resistance in male mice. Because central insulin resistance is a factor that contributes to the disruption of energy homeostasis (33),
Sirt1 activation may provide a possible solution for obesity that is caused by defective energy balance regulation.
Summary
Based on these results we conclude that Sirt1 overexpression in POMC neurons partially rescues obesity caused by CN-Foxo1 by decreasing FoxO1 acetylation and protein level and that Sirt1 can partially reverse obesity caused by the central insulin resistance in male mice. Because the central insulin resistance is one of the contributing factors for the disruption of energy homeostasis (33), Sirt1 activation could provide a possible solution for obesity caused by the defective regulation of energy balance.
Acknowledgements
We thank Ms. C. Osawa for excellent technical assistance, Ms. Kanako Ohtani for secretarial assistance, and the members of the Kitamura lab for constructive discussions of the data. We also thank the Biosignal Genome Resource Center (Gunma University) for the use of the facility. We thank Dr. Jun Nakae at Keio University for providing us with Rosa 26CN-FoxO1 mice.
Address all correspondence and request for reprints to: Tsutomu Sasaki or Tadahiro Kitamura, Metabolic Signal Research Center, Institute for Molecular and Cellular Regulation, Gunma University, 3-39-15 Showa-machi, maebashi-shi, Gunma 371- 8512, Japan. Email: [email protected] or [email protected]
This study was supported by a fellowship grant (to V.Y.S); a Grants-in-Aid for Scientific Research (C) (to T.S.); a Grant-in-Aid for Scientific Research (B)(to T.K.);
and a grant from the Global Centers of Excellence program from the Ministry of Education, Culture, Sports, Science, and Technology of Japan.
Disclosure Summary: The authors declare no conflicts of interest, financial or otherwise.
References
1. Ogden CL, Carroll MD, Curtin LR, Lamb MM, Flegal KM 2010 Prevalence of high body mass index in US children and adolescents, 2007- 2008. JAMA 303:242-249
2. Hummasti S, Hotamisligil GS 2010 Endoplasmic reticulum stress and inflammation in obesity and diabetes. Circ Res 107:579-591
3. Horvath TL 2005 The hardship of obesity: a soft-wired hypothalamus. Nat Neurosci 8:561-565
4. Woods SC, D'Alessio DA 2008 Central control of body weight and appetite. J Clin Endocrinol Metab 93:S37-50
5. Coll AP, Farooqi IS, Challis BG, Yeo GS, O'Rahilly S 2004 Proopiomelanocortin and energy balance: insights from human and murine genetics. J Clin Endocrinol Metab 89:2557-2562
6. Barsh GS, Farooqi IS, O'Rahilly S 2000 Genetics of body-weight regulation.
Nature 404:644-651
7. Vaisse C, Clement K, Guy-Grand B, Froguel P 1998 A frameshift mutation in human MC4R is associated with a dominant form of obesity. Nat Genet 20:113-114
8. Yeo GS, Lank EJ, Farooqi IS, Keogh J, Challis BG, O'Rahilly S 2003 Mutations in the human melanocortin-4 receptor gene associated with severe familial obesity disrupts receptor function through multiple molecular mechanisms. Hum Mol Genet 12:561-574
9. Burks DJ, Font de Mora J, Schubert M, Withers DJ, Myers MG, Towery HH, Altamuro SL, Flint CL, White MF 2000 IRS-2 pathways integrate female reproduction and energy homeostasis. Nature 407:377-382
10. Kubota N, Terauchi Y, Tobe K, Yano W, Suzuki R, Ueki K, Takamoto I, Satoh H, Maki T, Kubota T, Moroi M, Okada-Iwabu M, Ezaki O, Nagai R, Ueta Y, Kadowaki T, Noda T 2004 Insulin receptor substrate 2 plays a crucial role in beta cells and the hypothalamus. J Clin Invest 114:917-927 11. Bruning JC, Gautam D, Burks DJ, Gillette J, Schubert M, Orban PC,
Klein R, Krone W, Muller-Wieland D, Kahn CR 2000 Role of brain insulin receptor in control of body weight and reproduction. Science 289:2122-2125 12. Accili D, Arden KC 2004 FoxOs at the crossroads of cellular metabolism,
differentiation, and transformation. Cell 117:421-426
13. Kim MS, Pak YK, Jang PG, Namkoong C, Choi YS, Won JC, Kim KS, Kim SW, Kim HS, Park JY, Kim YB, Lee KU 2006 Role of hypothalamic Foxo1 in the regulation of food intake and energy homeostasis. Nat Neurosci 9:901-906
14. Kitamura T, Feng Y, Kitamura YI, Chua SC, Jr., Xu AW, Barsh GS, Rossetti L, Accili D 2006 Forkhead protein FoxO1 mediates Agrp-dependent effects of leptin on food intake. Nat Med 12:534-540
15. Iskandar K, Cao Y, Hayashi Y, Nakata M, Takano E, Yada T, Zhang C, Ogawa W, Oki M, Chua S, Jr., Itoh H, Noda T, Kasuga M, Nakae J 2010 PDK-1/FoxO1 pathway in POMC neurons regulates Pomc expression and food intake. Am J Physiol Endocrinol Metab 298:E787-798
16. Plum L, Lin HV, Dutia R, Tanaka J, Aizawa KS, Matsumoto M, Kim AJ, Cawley NX, Paik JH, Loh YP, DePinho RA, Wardlaw SL, Accili D 2009
The obesity susceptibility gene Cpe links FoxO1 signaling in hypothalamic pro-opiomelanocortin neurons with regulation of food intake. Nat Med 15:1195-1201
17. Ropelle ER, Pauli JR, Prada P, Cintra DE, Rocha GZ, Moraes JC, Frederico MJ, da Luz G, Pinho RA, Carvalheira JB, Velloso LA, Saad MA, De Souza CT 2009 Inhibition of hypothalamic Foxo1 expression reduced food intake in diet-induced obesity rats. J Physiol 587:2341-2351 18. Imai S, Armstrong CM, Kaeberlein M, Guarente L 2000 Transcriptional
silencing and longevity protein Sir2 is an NAD-dependent histone deacetylase.
Nature 403:795-800
19. Sun C, Zhang F, Ge X, Yan T, Chen X, Shi X, Zhai Q 2007 SIRT1 improves insulin sensitivity under insulin-resistant conditions by repressing PTP1B. Cell Metab 6:307-319
20. Kitamura YI, Kitamura T, Kruse JP, Raum JC, Stein R, Gu W, Accili D 2005 FoxO1 protects against pancreatic beta cell failure through NeuroD and MafA induction. Cell Metab 2:153-163
21. Sasaki T, Kim HJ, Kobayashi M, Kitamura YI, Yokota-Hashimoto H, Shiuchi T, Minokoshi Y, Kitamura T 2010 Induction of hypothalamic Sirt1 leads to cessation of feeding via agouti-related peptide. Endocrinology 151:2556-2566
22. Sasaki T, Kikuchi O, Shimpuku M, Susanti VY, Yokota-Hashimoto H, Taguchi R, Shibusawa N, Sato T, Tang L, Amano K, Kitazumi T, Kuroko M, Fujita Y, Maruyama J, Lee YS, Kobayashi M, Nakagawa T, Minokoshi Y, Harada A, Yamada M, Kitamura T 2013 Hypothalamic Sirt1
prevents age-associated weight gain by improving leptin sensitivity in mice.
Diabetologia, in press, DOI: 10. 1007/s00125-013-3140-5
23. Sasaki T, Maier B, Bartke A, Scrable H 2006 Progressive loss of SIRT1 with cell cycle withdrawal. Aging Cell 5:413-422
24. Soriano 1999 Generalized LacZ expression with the Rosa 26 Cre reporter strain Nature genetics 21:70-71
25. Plum L, Lin HV, Aizawa KS, Liu Y, Accili D 2012 InsR/FoxO1 signaling curtails hypothalamic POMC neuron number. PLoS One 7:e31487
26. Qiang L, Banks AS, Accili D 2010 Uncoupling of acetylation from phosphorylation regulates FoxO1 function independent of its subcellular localization. J Biol Chem 285:27396-27401
27. Balthasar N, Dalgaard LT, Lee CE, Yu J, Funahashi H, Williams T, Ferreira M, Tang V, McGovern RA, Kenny CD, Christiansen LM, Edelstein E, Choi B, Boss O, Aschkenasi C, Zhang CY, Mountjoy K, Kishi T, Elmquist JK, Lowell BB 2005 Divergence of melanocortin pathways in the control of food intake and energy expenditure. Cell 123:493- 505
28. Elias CF, Lee C, Kelly J, Aschkenasi C, Ahima RS, Couceyro PR, Kuhar MJ, Saper CB, Elmquist JK 1998 Leptin activates hypothalamic CART neurons projecting to the spinal cord. Neuron 21:1375-1385
29. Swanson LW, Kuypers HG 1980 A direct projection from the ventromedial nucleus and retrochiasmatic area of the hypothalamus to the medulla and spinal cord of the rat. Neurosci Lett 17:307-312
30. Bamshad M, Song CK, Bartness TJ 1999 CNS origins of the sympathetic nervous system outflow to brown adipose tissue. Am J Physiol 276:R1569- 1578
31. Rossi J, Balthasar N, Olson D, Scott M, Berglund E, Lee CE, Choi MJ, Lauzon D, Lowell BB, Elmquist JK 2011 Melanocortin-4 receptors expressed by cholinergic neurons regulate energy balance and glucose homeostasis. Cell Metab 13:195-204
32. Kitamura T, Nakae J, Kitamura Y, Kido Y, Biggs WH, 3rd, Wright CV, White MF, Arden KC, Accili D 2002 The forkhead transcription factor Foxo1 links insulin signaling to Pdx1 regulation of pancreatic beta cell growth.
J Clin Invest 110:1839-1847
33. Plum L, Belgardt BF, Bruning JC 2006 Central insulin action in energy and glucose homeostasis. J Clin Invest 116:1761-1766
Figure and Legends
Fig. 1. Sirt1 overexpression in POMC neurons rescues the obese phenotype of CN-FoxO1 in male but not female mice. A, Changes in the body weight of control mice, CN-FoxO1 KI mice, Sirt1 KI mice, and double-KI male (left) (n=8–12) and female (right) mice (n=7–15). Body weight was measured weekly from 4 weeks until 27 weeks of age. B, The body length of mice in the four groups (n=8-12). C and D, The weight of the brown adipose tissue (BAT) (C) and perigonadal white adipose tissue (WAT) (D) of the studied mice (n=8-12). Data are reported as mean ± SEM. *, p<0.05 by ANOVA.
Fig. 2. Food intake, oxygen consumption, and locomotor activity of the male mice in the study. A, Daily food intake at 25–26 weeks of age. B and C, O2 consumption (VO2) (B) and locomotor activity (C) in control mice, CN-FoxO1 KI mice, Sirt1 KI mice, and DKI mice. D and E, The expression levels of Ucp1 and Pgc1 in mouse brown adipose tissue (BAT) at 28 weeks of age as assessed by real-time RT-PCR.
Fig. 3. Sirt1 and FoxO1 regulate the number of POMC neurons. A and B, The average number (A) and the total number (B) of POMC neurons in the arcuate nucleus of the indicated mice. C. Representative arcuate nucleus sections with POMC neurons from control, CN-FoxO1, Sirt1, and DKI mice. Sections were immunostained with anti-POMC (red) and stained with DAPI (blue). 400-fold magnification. Data are reported as mean ± SEM. *, p<0.05 by ANOVA.
Fig. 4. Sirt1 affects the acetylation and amount of CN-FoxO1 protein but not its nuclear localization. A, Sirt1 overexpression decreases Foxo1 acetylation in hypothalamic N41 cells. B, Sirt1 overexpression decreases CN-FoxO1 protein in hypothalamic N41 cells. C, Sirt1 overexpression does not affect CN-FoxO1 localization in hypothalamic N41 cells. D, The amount of nuclear FoxO1 protein is decreased in Ad-Cre-infected DKI MEFs (SF) compared to CN-FoxO1 MEF (F) mice.
Figure 1
Female body weight (g)
Age (weeks)
Male BAT weight (g) Female BAT weight (g)
15 20 25 30 35 40
4 8 12 16 20 24 28
** *
*
10 15 20 25 30
4 8 12 16 20 24 28
Female body length (cm) 0 25 50 75 100 125
0.00 0.05 0.10 0.15
0.00 0.05 0.10 0.15
0.0 0.1 0.2 0.3
Female perigonadal WAT weight (g) 0.0
0.1 0.2 0.3 0.4
Male perigonadal WAT weight (g) Male body weight (g) Male body length (cm) 0 25 50 75 100
125 *
* *
* *
* * * * * * * * * * * * *
A
B
C
D
Control CN-FoxO1 Sirt1 SF (DKI)
Age (weeks)
28 weeks 28 weeks
28 weeks 28 weeks
28 weeks 28 weeks
Figure 2
0 2 4 6
Food intake (g/day)
0 20000 40000 60000
light dark 24h Cumulative VO2 (L/g/period)
0 2000 4000 6000
light dark 24h Locomotor activity (103 counts/period)
6 4 2
0 1 2
0 1 2
BATUcp-1/β-actin (A.U.) BATPGC-1α/β-actin (A.U.)
A B C
D E
6 4 2
Control CN-FoxO1 Sirt1 SF (DKI)
Figure 3
0 20 40 60 80
1 2 3 4 5
* *
Section number A
0 100 200 300 400
* p=0.051 B
Average number of POMC neurons Total number of POMC neurons
Control CN-FoxO1
Sirt1 DKI
C
Control CN-FoxO1 Sirt1 SF (DKI)
Figure 4
Ac-FoxO1 FoxO1 Sirt1 α-Tubulin
70 70 96 45 Ad-HA-FoxO1:
Ad-Sirt1 : -
- + + + N41 cells (whole cell lysate)
- - - Ad GFP
Ad CN-FoxO1 Ad Sirt 1
+ - -
- + -
- +
- +
- +
Sirt1 96
HA-CN-FoxO1 70
β-Actin 42
N41 cells (whole cell lysate)
FoxO1 Sirt1 B23 α-Tubulin
70 96 32 45
MEFs WT S F SF WT S F SF A
C
Mouse embryonic fibroblasts (MEFs) with Ad-Cre (fractionated)
B
D
N41 cells (fractionated) -
+ + - + +
- -
- -
Sirt1
HA-CN-FoxO1 Ad GFP
Ad CN-FoxO1 Ad Sirt 1
+ +
Nuclear
+ + Cytoplasmic
- +
- -
- - - -
B23
- -
- - + - -
96 70 32 -
Cytoplasmic Nuclear
WT: Wild Type S: Sirt1
F: FoxO1
SF: Sirt1/FoxO1
Submitted Paper
Sirt1 Overexpression in POMC Neurons Partially Rescues the Obese Phenotype Induced by Insulin-resistant Constitutively-nuclear FoxO1 in Male Mice.
Endocrinology (投稿中)